Delivery of Heterologous Proteins, Enzymes, and Antigens via the Bacterial Type III Secretion System

Abstract
1. Introduction
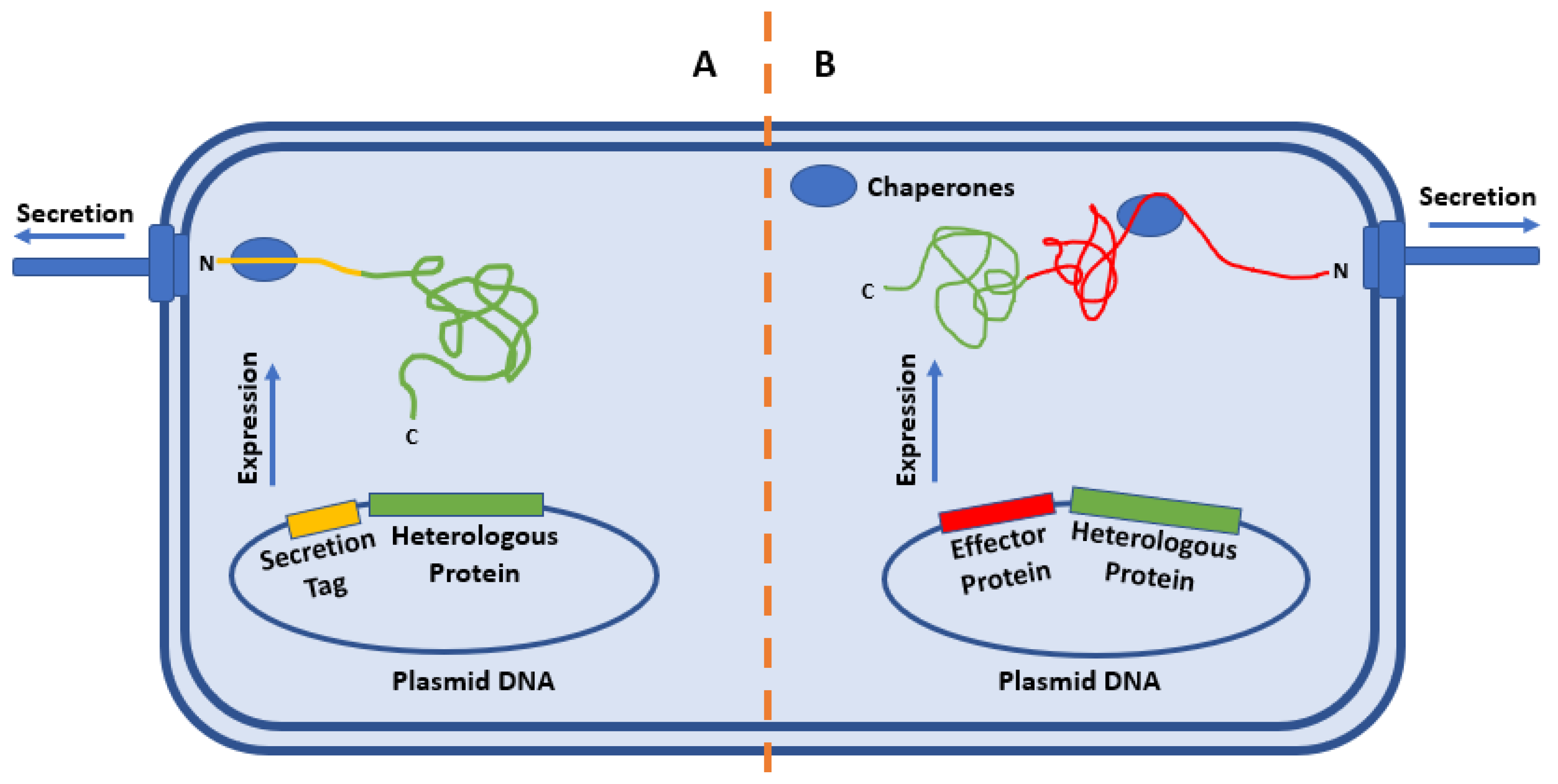
2. Strategies for the Secretion of Heterologous Proteins
2.1. “Secretion Tag”
2.2. Whole-Protein Fusions
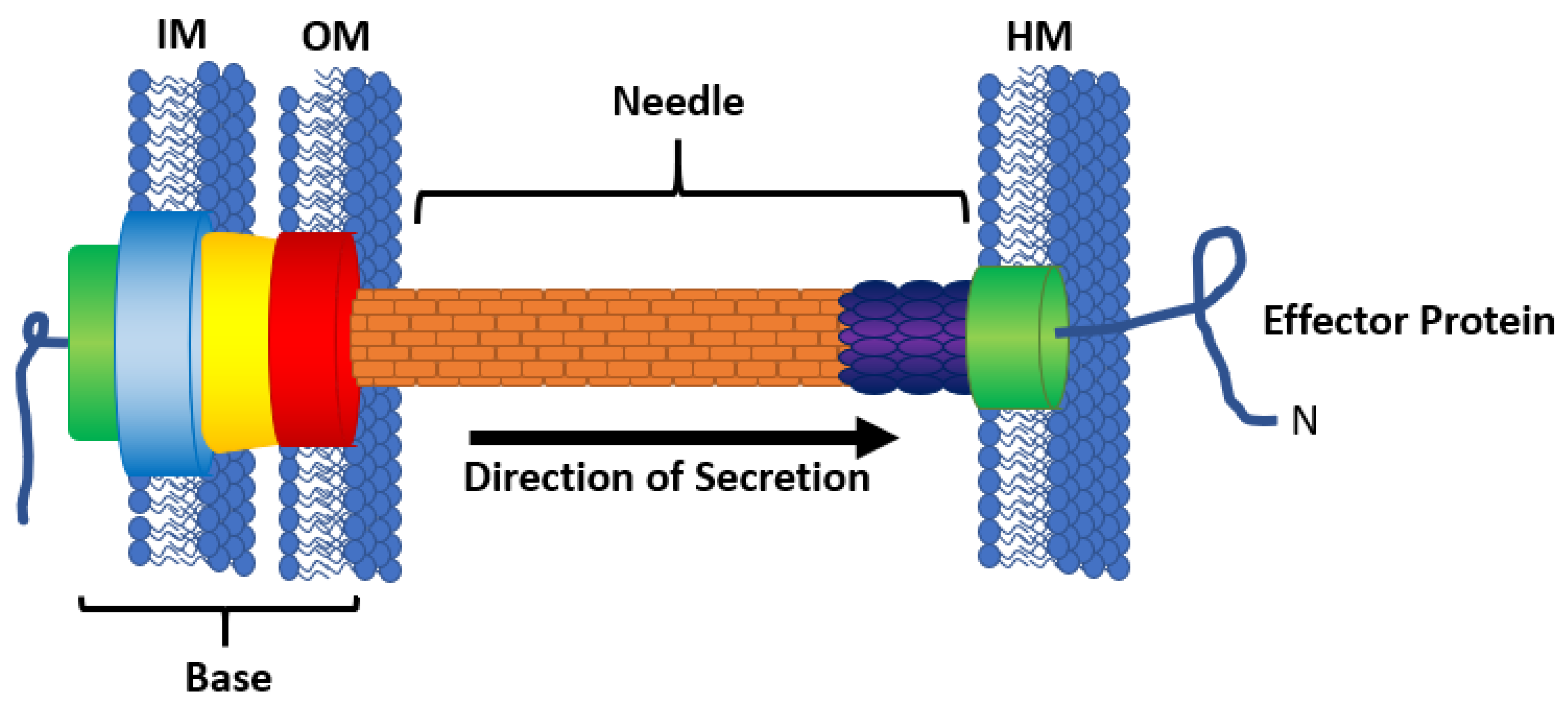
3. Physics of Substrate Secretion
3.1. Secretion Efficiency
3.2. Protein Folding
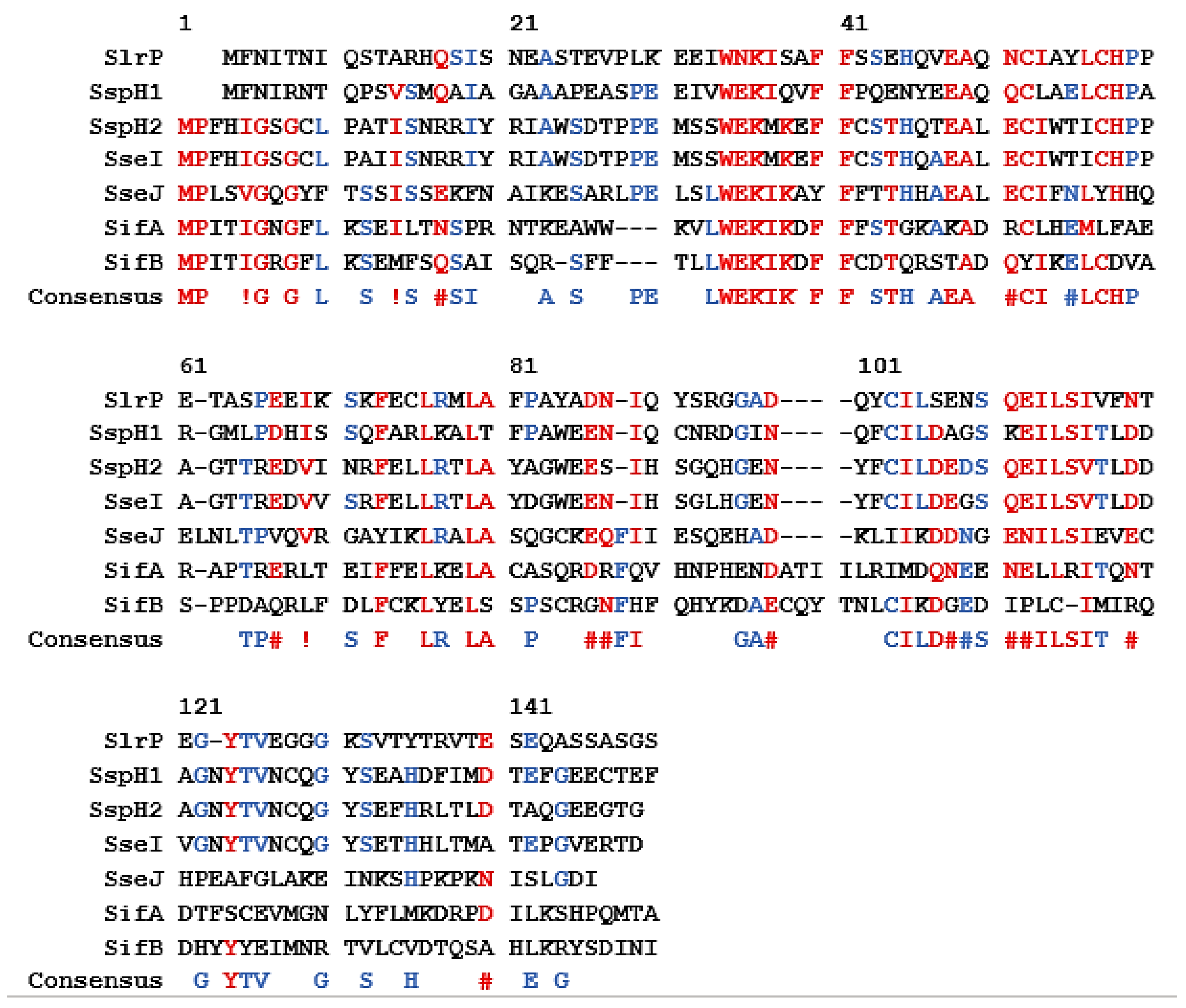
4. Delivery of Antigenic Proteins
5. Translocation vs. Secretion
6. Enzyme and Analyte Secretion for Monitoring T3SS Activity
6.1. CyaA Translocation
6.2. β-Lactamase (Bla) Translocation
6.3. Phospholipase Secretion
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moraes, T.F.; Spreter, T.; Strynadka, N.C. Piecing together the type III injectisome of bacterial pathogens. Curr. Opin. Struct. Biol. 2008, 18, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Buttner, D. Protein export according to schedule: Architecture, assembly, and regulation of type III secretion systems from plant- and animal-pathogenic bacteria. Microbiol. Mol. Biol. Rev. 2012, 76, 262–310. [Google Scholar] [CrossRef]
- Sekiya, K.; Ohishi, M.; Ogino, T.; Tamano, K.; Sasakawa, C.; Abe, A. Supermolecular structure of the enteropathogenic Escherichia coli type III secretion system and its direct interaction with the EspA-sheath-like structure. Proc. Natl. Acad. Sci. USA 2001, 98, 11638–11643. [Google Scholar] [CrossRef]
- Gao, X.; Mu, Z.; Yu, X.; Qin, B.; Wojdyla, J.; Wang, M.; Cui, S. Structural insight into conformational changes induced by ATP binding in a type III secretion-associated ATPase from Shigella flexneri. Front. Microbiol. 2018, 9, 1468. [Google Scholar] [CrossRef] [PubMed]
- Akeda, Y.; Galán, J.E. Chaperone release and unfolding of substrates in type III secretion. Nature 2005, 437, 911–915. [Google Scholar] [CrossRef] [PubMed]
- Andrade, A.; Pablo, J.; Espinosa, N.; Pe, G.; Gonza, B. Enzymatic characterization of the enteropathogenic Escherichia coli type III secretion ATPase EscN. Arch. Biochem. Biophys. 2007, 468, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Hodgkinson, J.L.; Horsley, A.; Stabat, D.; Simon, M.; Johnson, S.; Da Fonseca, P.C.A.; Morris, E.P.; Wall, J.S.; Lea, S.M.; Blocker, A.J. Three-dimensional reconstruction of the Shigella T3SS transmembrane regions reveals 12-fold symmetry and novel features throughout. Nat. Struct. Mol. Biol. 2009, 16, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Blocker, A.; Jouihri, N.; Larquet, E.; Gounon, P.; Ebel, F.; Parsot, C.; Sansonetti, P.; Allaoui, A. Structure and composition of the Shigella flexneri “needle complex”, a part of its type III secreton. Mol. Microbiol. 2001, 39, 652–663. [Google Scholar] [CrossRef]
- Warawa, J.; Finlay, B.B.; Kenny, B. Type III secretion-dependent hemolytic activity of enteropathogenic Escherichia coli. Infect. Immun. 1999, 67, 5538–5540. [Google Scholar] [CrossRef]
- Jarvis, K.G.; Giron, J.A.; Jerse, A.E.; McDaniel, T.K.; Donnenberg, M.S.; Kaper, J.B. Enteropathogenic Escherichia coli contains a putative type III secretion system necessary for the export of proteins involved in attaching and effacing lesion formation. Proc. Natl. Acad. Sci. USA 1995, 92, 7996–8000. [Google Scholar] [CrossRef]
- Shaw, R.K.; Cleary, J.; Murphy, M.S.; Frankel, G.; Knutton, S. Interaction of enteropathogenic Escherichia coli with human intestinal mucosa: Role of effector proteins in brush border remodeling and formation of attaching and effacing lesions. Infect. Immun. 2005, 73, 1243–1251. [Google Scholar] [CrossRef]
- Holmes, A.; Mühlen, S.; Roe, A.J.; Dean, P. The EspF effector, a bacterial pathogen’s Swiss army knife. Infect. Immun. 2010, 78, 4445–4453. [Google Scholar] [CrossRef] [PubMed]
- Sory, M.P.; Cornelis, G.R. Translocation of a hybrid YopE-adenylate cyclase from Yersinia enterocolitica into HeLa cells. Mol. Microbiol. 1994, 14, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Marketon, M.M.; Depaolo, R.W.; Debord, K.L. Plague bacteria target immune cells during infection. Science 2005, 309, 1739–1742. [Google Scholar] [CrossRef]
- Mou, X.; Souter, S.; Du, J.; Reeves, A.Z.; Lesser, C.F. Synthetic bottom-up approach reveals the complex interplay of Shigella effectors in regulation of epithelial cell death. Proc. Natl. Acad. Sci. USA 2018, 115, 6452–6457. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Iwatsuki, M.; Nagai, T.; Matsumoto, A.; Takahashi, Y.; Shiomi, K.; Omura, S.; Abe, A. A small-molecule inhibitor of the bacterial type III secretion system protects against in vivo infection with Citrobacter rodentium. J. Antibiot. (Tokyo) 2011, 64, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Hocking, D.M.; Cheng, C.; Dogovski, C.; Perugini, M.A.; Holien, J.K.; Parker, M.W.; Hartland, E.L.; Tauschek, M.; Robins-Browne, R.M. Disarming bacterial virulence through chemical inhibition of the DNA binding domain of an AraC-like transcriptional activator protein. J. Biol. Chem. 2013, 288, 31115–31126. [Google Scholar] [CrossRef]
- Franzin, F.M.; Sircili, M.P. Locus of enterocyte effacement: A pathogenicity island involved in the virulence of enteropathogenic and enterohemorragic Escherichia coli subjected to a complex network of gene regulation. Biomed. Res. Int. 2015, 2015, 1–10. [Google Scholar] [CrossRef]
- Croxen, M.A.; Finlay, B.B. Molecular mechanisms of Escherichia coli pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar] [CrossRef]
- Park, D.; Lara-Tejero, M.; Waxham, M.N.; Li, W.; Hu, B. Visualization of the type III secretion mediated Salmonella – host cell interface using cryo-electron tomography. eLife 2018, 7, e39514. [Google Scholar] [CrossRef]
- Figueira, R.; Holden, D.W. Functions of the Salmonella pathogenicity island 2 (SPI-2) type III secretion system effectors. Microbiology 2012, 2, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Li, Z. Conserved type III secretion system exerts important roles in Chlamydia Trach. Int. J. Exp. Pathol. 2014, 7, 5404–5414. [Google Scholar]
- Keyser, P.; Elofsson, M.; Rosell, S.; Wolf-Watz, H. Virulence blockers as alternatives to antibiotics: Type III secretion inhibitors against Gram-negative bacteria. J. Intern. Med. 2008, 264, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Avican, K.; Fahlgren, A.; Erttmann, S.F.; Nuss, A.M.; Dersch, P.; Fallman, M.; Edgren, T.; Wolf-watz, H. Increased plasmid copy number is essential for Yersinia T3SS function and virulence. Science 2016, 353, 492–495. [Google Scholar] [CrossRef]
- Pan, N.J.; Brady, M.J.; Leong, J.M.; Goguen, J.D. Targeting type III secretion in Yersinia pestis. Antimicrob. Agents Chemother. 2009, 53, 385–392. [Google Scholar] [CrossRef]
- Heine, S.J.; Franco-Mahecha, O.L.; Khandra, T.; Drachenberg, C.B.; van Roosmalen, M.L.; Leenhouts, K.; Picking, W.L.; Pasetti, M.F. A combined YopB and LcrV subunit vaccine elicits protective immunity against Yersinia infection in adult and infant mice. J. Immunol. 2019, 202, 2005–2016. [Google Scholar] [CrossRef]
- NCDPH North Carolina Department of Health and Human Services: E. coli. Available online: https://epi.dph.ncdhhs.gov/cd/diseases/ecoli.html (accessed on 17 April 2020).
- Centers for Disease Control and Prevention (CDC). Salmonella. Available online: https://www.cdc.gov/salmonella/index.html (accessed on 17 April 2020).
- Centers for Disease Control and Prevention (CDC). Pseudomonas Aeruginosa in Healthcare Settings. Available online: https://www.cdc.gov/hai/organisms/pseudomonas.html (accessed on 17 April 2020).
- Centers for Disease Control and Prevention (CDC). Yersinia Enterocolitica Questions and Answers. Available online: https://www.cdc.gov/yersinia/faq.html (accessed on 17 April 2020).
- Li, J.; Lv, C.; Sun, W.; Li, Z.; Han, X.; Li, Y.; Shen, Y. Cytosporone B, an inhibitor of the type III secretion system of Salmonella enterica serovar Typhimurium. Antimicrob. Agents Chemother. 2013, 57, 2191–2198. [Google Scholar] [CrossRef]
- Bzdzion, L.; Krezel, H.; Wrzeszcz, K.; Grzegorek, I.; Nowinska, K.; Chodaczek, G.; Swietnicki, W. Design of small molecule inhibitors of type III secretion system ATPase EscN from enteropathogenic Escherichia coli. Acta Biochim. Pol. 2017, 64, 49–63. [Google Scholar] [CrossRef]
- Bohnhoff, B.Y.M.; Miller, C.P.; Martin, W.R. Resistance of the mouse’s intestinal tract to experimental Salmonella infection I. factors which interfere with the initiation of infection. J. Exp. Med. 1964, 120, 805–816. [Google Scholar] [CrossRef]
- Kauppi, A.M.; Nordfelth, R.; Uvell, H.; Wolf-Watz, H.; Elofsson, M. Targeting bacterial virulence: Inhibitors of type III secretion in Yersinia. Chem. Biol. 2003, 10, 241–249. [Google Scholar] [CrossRef]
- Marshall, N.C.; Finlay, B.B. Targeting the type III secretion system to treat bacterial infections. Expert Opin. Targets. 2014, 18, 137–152. [Google Scholar] [CrossRef]
- Green, C.A.; Kamble, N.S.; Court, E.K.; Bryant, O.J.; Hicks, M.G.; Lennon, C.; Fraser, G.M.; Wright, P.C.; Stafford, G.P. Engineering the flagellar type III secretion system: Improving capacity for secretion of recombinant protein. Microb. Cell Fact. 2019, 18, 10. [Google Scholar] [CrossRef] [PubMed]
- Wieser, A.; Magistro, G.; Nörenberg, D.; Hoffmann, C.; Schubert, S. First multi-epitope subunit vaccine against extraintestinal pathogenic Escherichia coli delivered by a bacterial type 3 secretion system (T3SS). Int. J. Med. Microbiol. 2012, 302, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, K.J.; Bevington, J.L.; Rosales, S.L.; Burdette, L.A.; Valdivia, E.; Tullman-Ercek, D. Proteins adopt functionally active conformations after type III secretion. Microb. Cell Fact. 2016, 15, 213. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.L.; Chakravarthy, S.; Mathieu, J.; Helmann, T.C.; Stodghill, P.; Swingle, B.; Martin, G.B.; Collmer, A. Pseudomonas syringae pv. tomato DC3000 type III secretion effector polymutants reveal an interplay between HopAD1 and AvrPtoB. Cell Host Microbe 2015, 17, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Kvitko, B.H.; Park, D.H.; Velásquez, A.C.; Wei, C.F.; Russell, A.B.; Martin, G.B.; Schneider, D.J.; Collmer, A. Deletions in the repertoire of Pseudomonas syringae pv. tomato DC3000 type III secretion effector genes reveal functional overlap among effectors. PLoS Pathog. 2009, 5, e1000388. [Google Scholar]
- Collmer, A.; Badel, J.L.; Charkowski, A.O.; Deng, W.L.; Fouts, D.E.; Ramos, A.R.; Rehm, A.H.; Anderson, D.M.; Schneewind, O.; van Dijk, K.; et al. Pseudomonas syringae Hrp type III secretion system and effector proteins. Proc. Natl. Acad. Sci. USA 2000, 97, 8770–8777. [Google Scholar] [CrossRef]
- Zhou, J.M.; Chai, J. Plant pathogenic bacterial type III effectors subdue host responses. Curr. Opin. Microbiol. 2008, 11, 179–185. [Google Scholar] [CrossRef]
- Munera, D.; Crepin, V.F.; Marches, O.; Frankel, G. N-terminal type III secretion signal of enteropathogenic Escherichia coli translocator proteins. J. Bacteriol. 2010, 192, 3534–3539. [Google Scholar] [CrossRef]
- Mills, E.; Baruch, K.; Charpentier, X.; Kobi, S.; Rosenshine, I. Real-time analysis of effector translocation by the type III secretion system of enteropathogenic Escherichia coli. Cell Host Microbe 2008, 3, 104–113. [Google Scholar] [CrossRef]
- Wiesand, U.; Sorg, I.; Amstutz, M.; Wagner, S.; van den Heuvel, J.; Lührs, T.; Cornelis, G.R.; Heinz, D.W. Structure of the type III secretion recognition protein YscU from Yersinia enterocolitica. J. Mol. Biol. 2009, 385, 854–866. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Niemann, G.S.; Brown, R.N.; Mushamiri, I.T.; Nguyen, N.T.; Taiwo, R.; Stufkens, A.; Smith, R.D.; Adkins, J.N.; McDermott, J.E.; Heffron, F. RNA type III secretion signals that require Hfq. J. Bacteriol. 2013, 195, 2119–2125. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P. Process of protein transport by the type III secretion system. Microbiol. Mol. Biol. Rev. 2004, 68, 771–795. [Google Scholar] [CrossRef] [PubMed]
- Stebbins, C.E.; Galán, J.E. Maintenance of an unfolded polypeptide by a cognate chaperone in bacterial type III secretion. Nature 2001, 414, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Schlumberger, M.C.; Müller, A.J.; Ehrbar, K.; Winnen, B.; Duss, I.; Stecher, B.; Hardt, W.D. Real-time imaging of type III secretion: Salmonella SipA injection into host cells. Proc. Natl. Acad. Sci. USA 2005, 102, 12548–12553. [Google Scholar] [CrossRef]
- Miao, E.A.; Miller, S.I. A conserved amino acid sequence directing intracellular type III secretion by Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 2000, 97, 7539–7544. [Google Scholar] [CrossRef]
- Zurawski, D.V.; Stein, M.A. SseA acts as the chaperone for the SseB component of the Salmonella pathogenicity island 2 translocon. Mol. Microbiol. 2003, 47, 1341–1351. [Google Scholar] [CrossRef]
- Widmaier, D.M.; Tullman-Ercek, D.; Mirsky, E.A.; Hill, R.; Govindarajan, S.; Minshull, J.; Voigt, C.A. Engineering the Salmonella type III secretion system to export spider silk monomers. Mol. Syst. Biol. 2009, 5, 1–9. [Google Scholar]
- Cardenal-Muñoz, E.; Ramos-Morales, F. Analysis of the expression, secretion and translocation of the Salmonella enterica type III secretion system effector SteA. PLoS ONE 2011, 6, e26930. [Google Scholar] [CrossRef]
- Metcalf, K.J.; Tullman-Ercek, D.; Azam, A.; Valdivia, E. Using transcriptional control to increase titers of secreted heterologous proteins by the type III secretion system. Appl. Environ. Microbiol. 2014, 80, 5927–5934. [Google Scholar] [CrossRef]
- Andersson, K.; Carballeira, N.; Magnusson, K.E.; Persson, C.; Stendahl, O.; Wolf-Watz, H.; Fällman, M. YopH of Yersinia pseudotuberculosis interrupts early phosphotyrosine signalling associated with phagocytosis. Mol. Microbiol. 1996, 20, 1057–1069. [Google Scholar] [CrossRef] [PubMed]
- Sansonetti, P.; Nhieu, V.G.T.; Enninga, J. Secretion of type III effectors into host cells in real time. Nat. Publ. Gr. 2005, 2, 959–965. [Google Scholar]
- Liang, S.T.; Xu, Y.C.; Dennis, P.; Bremer, H. mRNA composition and control of bacterial gene expression. J. Bacteriol. 2000, 182, 3037–3044. [Google Scholar] [CrossRef] [PubMed]
- Mitarai, N.; Sneppen, K.; Pedersen, S. Ribosome collisions and translation efficiency: Optimization by codon usage and mRNA destabilization. J. Mol. Biol. 2008, 382, 236–245. [Google Scholar] [CrossRef]
- Proshkin, S.; Rahmouni, R.; Mironov, A.; Nudler, E. Cooperation between translating ribosomes and RNA polymerase in transcription elongation. Science 2010, 328, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Dohlich, K.; Zumsteg, A.B.; Goosmann, C.; Kolbe, M. A substrate-fusion protein is trapped inside the type III secretion system channel in Shigella flexneri. PLoS Pathog. 2014, 10, e1003881. [Google Scholar] [CrossRef]
- Niebuhr, K.; Jouihri, N.; Allaoui, A.; Gounon, P.; Sansonetti, P.J.; Parsot, C. IpgD, a protein secreted by the type III secretion machinery of Shigella flexneri, is chaperoned by IpgE and implicated in entry focus formation. Mol. Microbiol. 2000, 38, 8–19. [Google Scholar] [CrossRef]
- Tsai, C.L.; Burkinshaw, B.J.; Strynadka, N.C.J.; Tainer, J.A. The Salmonella type III secretion system virulence effector forms a new hexameric chaperone assembly for export of effector/chaperone complexes. J. Bacteriol. 2015, 197, 672–675. [Google Scholar] [CrossRef]
- Carleton, H.A.; Lara-Tejero, M.; Liu, X.; Galán, J.E. Engineering the type III secretion system in non-replicating bacterial minicells for antigen delivery. Nat. Commun. 2013, 4, 1590. [Google Scholar] [CrossRef]
- Rüssmann, H.; Shams, H.; Poblete, F.; Fu, Y.; Galán, J.E.; Donis, R.O. Delivery of epitopes by the Salmonella type III secretion system for vaccine development. Science 1998, 281, 565–568. [Google Scholar]
- Xu, C.; Zhang, B.Z.; Lin, Q.; Deng, J.; Yu, B.; Arya, S.; Yuen, K.Y.; Huang, J.D. Live attenuated Salmonella Typhimurium vaccines delivering SaEsxA and SaEsxB via type III secretion system confer protection against Staphylococcus aureus infection. BMC Infect. Dis. 2018, 18, 195. [Google Scholar] [CrossRef] [PubMed]
- Aguilera-Herce, J.; García-Quintanilla, M.; Romero-Flores, R.; McConnell, M.J.; Ramos-Morales, F. A Live Salmonella vaccine delivering PcrV through the type III secretion system protects against Pseudomonas aeruginosa. mSphere 2019, 4, e00116–e00119. [Google Scholar] [CrossRef] [PubMed]
- Yount, J.S.; Tsou, L.K.; Dossa, P.D.; Kullas, A.L.; van der Velden, A.W.M.; Hang, H.C. Visible fluorescence detection of type III protein secretion from bacterial pathogens. J. Am. Chem. Soc. 2010, 132, 8244–8245. [Google Scholar] [CrossRef]
- Popa, C.M.; Tabuchi, M.; Valls, M. Modification of bacterial effector proteins inside eukaryotic host cells. Front. Cell. Infect. Microbiol. 2016, 6, 73. [Google Scholar] [CrossRef] [PubMed]
- Jeyaharan, D.; Aston, P.; Garcia-Perez, A.; Schouten, J.; Davis, P.; Dixon, A.M. Soluble expression, purification and functional characterisation of carboxypeptidase G2 and its individual domains. Protein Expr. Purif. 2016, 127, 44–52. [Google Scholar] [CrossRef]
- Tao, H.; Fan, S.; Jiang, S.; Xiang, X.; Yan, X.; Zhang, L. Small molecule inhibitors specifically targeting the type III secretion system of Xanthomonas oryzae on rice. Int. J. Mol. Sci. 2019, 20, 971. [Google Scholar] [CrossRef]
- Khokhani, D.; Zhang, C.; Li, Y.; Wang, Q.; Zeng, Q.; Yamazaki, A.; Hutchins, W.; Zhou, S.; Chen, X.; Yang, C. Discovery of plant phenolic compounds that act as type III secretion system inhibitors or inducers of the fire blight pathogen, Erwinia amylovora. Appl. Environ. Microbiol. 2013, 79, 5424–5436. [Google Scholar] [CrossRef]
- Pendergrass, H.A.; May, A.E. Natural product type III secretion system inhibitors. Antibiotics 2019, 8, 162. [Google Scholar] [CrossRef]
- May, A.E.; Khosla, C. Discovery and mechanism of type III secretion system inhibitors. Isr. J. Chem. 2013, 53, 577–587. [Google Scholar] [CrossRef]
- Crawford, J.A.; Kaper, J.B. The N-terminus of enteropathogenic Escherichia coli (EPEC) Tir mediates transport across bacterial and eukaryotic cell membranes. Mol. Microbiol. 2002, 46, 855–868. [Google Scholar] [CrossRef]
- Negrea, A.; Bjur, E.; Ygberg, S.E.; Elofsson, M.; Wolf-Watz, H.; Rhen, M. Salicylidene acylhydrazides that affect type III protein secretion in Salmonella enterica serovar Typhimurium. Antimicrob. Agents Chemother. 2007, 51, 2867–2876. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Qiu, J.; Luo, Z.Q.; Deng, X. The herbal compound thymol protects mice from lethal infection by Salmonella Typhimurium. Front. Microbiol. 2018, 9, 1002. [Google Scholar] [CrossRef] [PubMed]
- Bartra, S.S.; Lorica, C.; Qian, L.; Gong, X.; Bahnan, W. Chromosomally-encoded Yersinia pestis type III secretion effector proteins promote infection in cells and in mice. Front. Cell. Infect. Microbiol. 2019, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Pinaud, L.; Sansonetti, P.J.; Phalipon, A. Host cell targeting by enteropathogenic bacteria T3SS effectors. Trends Microbiol. 2018, 26, 266–283. [Google Scholar] [CrossRef] [PubMed]
- Felise, H.B.; Nguyen, H.V.; Pfuetzner, R.A.; Barry, K.C.; Stona, R.; Blanc, M.P.; Bronstein, P.A.; Kline, T.; Miller, S.I. An inhibitor of Gram-negative bacterial virulence protein secretion. Cell Host Microbe 2009, 4, 325–336. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Organism | Protein | Heterologous (Y/N) | Data | Rate | Ref |
|---|---|---|---|---|---|
| S. typhimurium | ADF-1 | Y | 9% protein secreted | 1.8 mg L−1 hr−1 Max conc. at 6 hr. | [52] |
| ADF-2 | Y | 17% protein secreted | |||
| ADF-3 | Y | 7.6% protein secreted | |||
| SipA | N | Initiation within 10–90 s. | 7–60 molecules sec−1 | [49] | |
| Active secretion for 100–600 s. | |||||
| DH | Y | 28 mg L−1 titer | [54] | ||
| Y. pseudotuberculosis | YopH | N | Initiation within 30 s | [55] | |
| S. flexneri | IpaB | N | ~50% secreted within 240 s | [56] | |
| IpaC | N |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pendergrass, H.A.; May, A.E. Delivery of Heterologous Proteins, Enzymes, and Antigens via the Bacterial Type III Secretion System. Microorganisms 2020, 8, 777. https://doi.org/10.3390/microorganisms8050777
Pendergrass HA, May AE. Delivery of Heterologous Proteins, Enzymes, and Antigens via the Bacterial Type III Secretion System. Microorganisms. 2020; 8(5):777. https://doi.org/10.3390/microorganisms8050777
Chicago/Turabian StylePendergrass, Heather A., and Aaron E. May. 2020. "Delivery of Heterologous Proteins, Enzymes, and Antigens via the Bacterial Type III Secretion System" Microorganisms 8, no. 5: 777. https://doi.org/10.3390/microorganisms8050777
APA StylePendergrass, H. A., & May, A. E. (2020). Delivery of Heterologous Proteins, Enzymes, and Antigens via the Bacterial Type III Secretion System. Microorganisms, 8(5), 777. https://doi.org/10.3390/microorganisms8050777