Volatile Compound Screening Using HS-SPME-GC/MS on Saccharomyces eubayanus Strains under Low-Temperature Pilsner Wort Fermentation

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Strains Used in this Study
2.2. Micro-Fermentation Assay
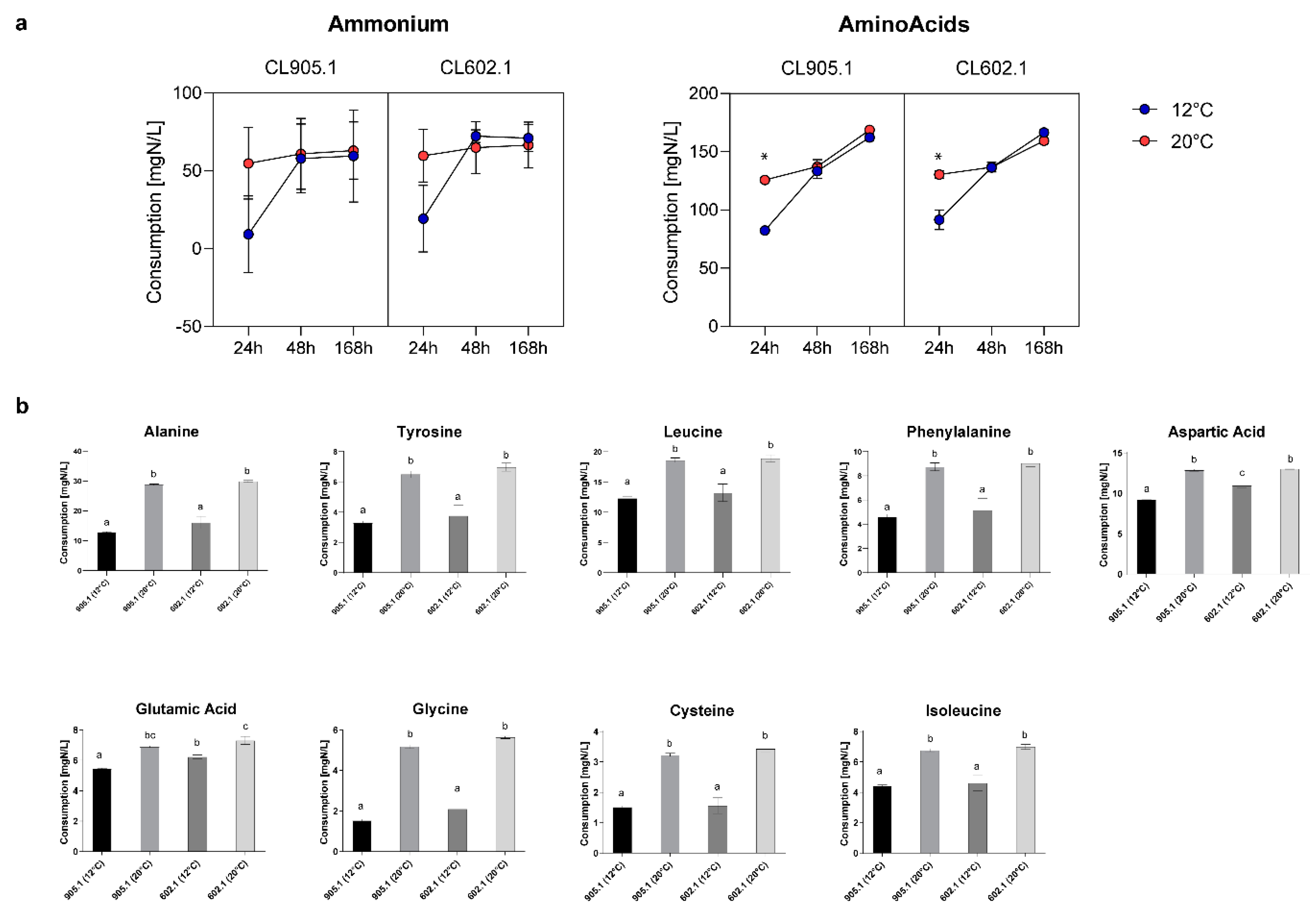
2.3. Ammonium and Amino Acid Analysis
2.4. HS-SPME-GC-MS Analysis
2.5. Statistical Analyses
3. Results
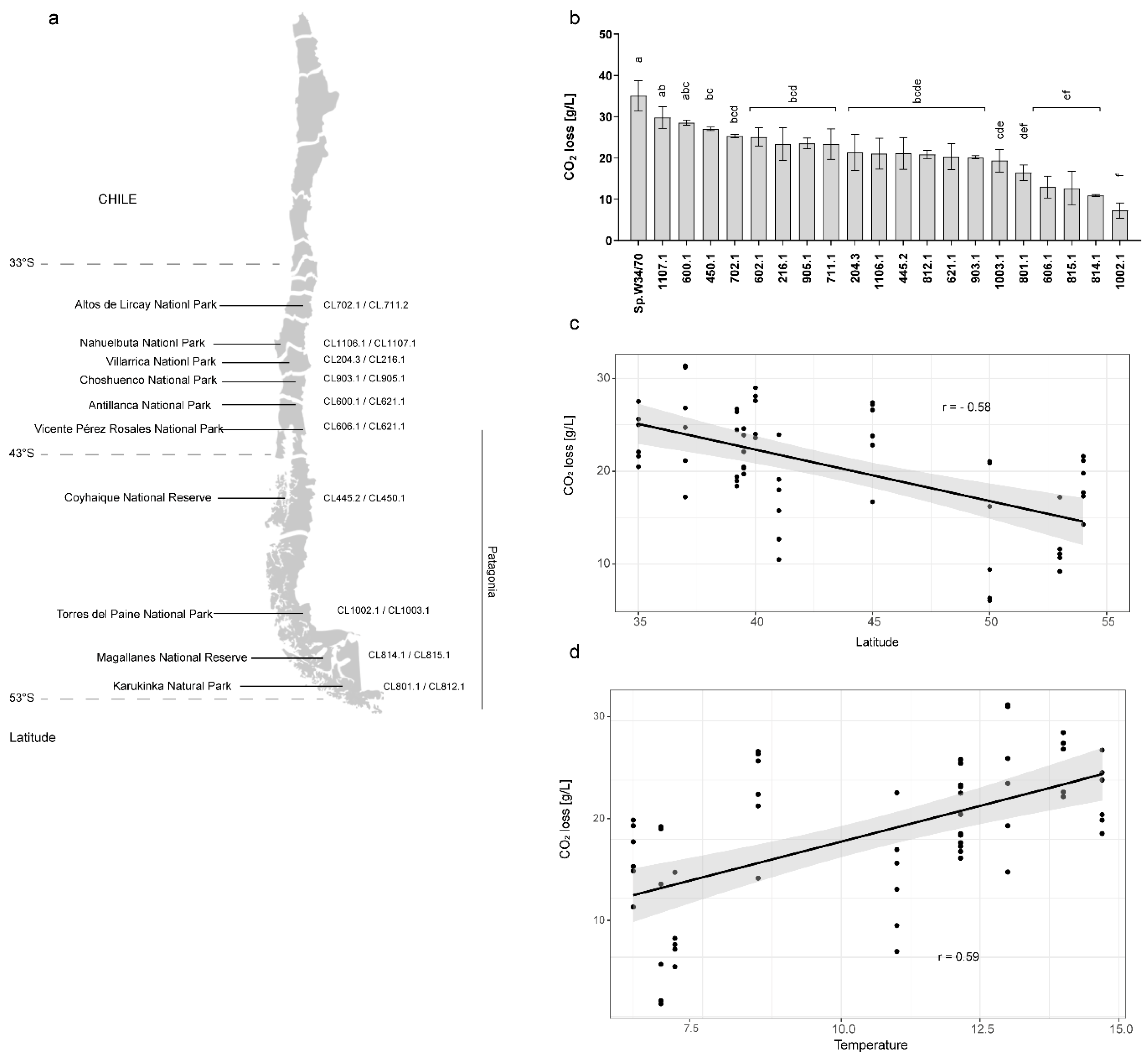
3.1. Differences in Fermentation Capacity Across S. eubayanus Isolates
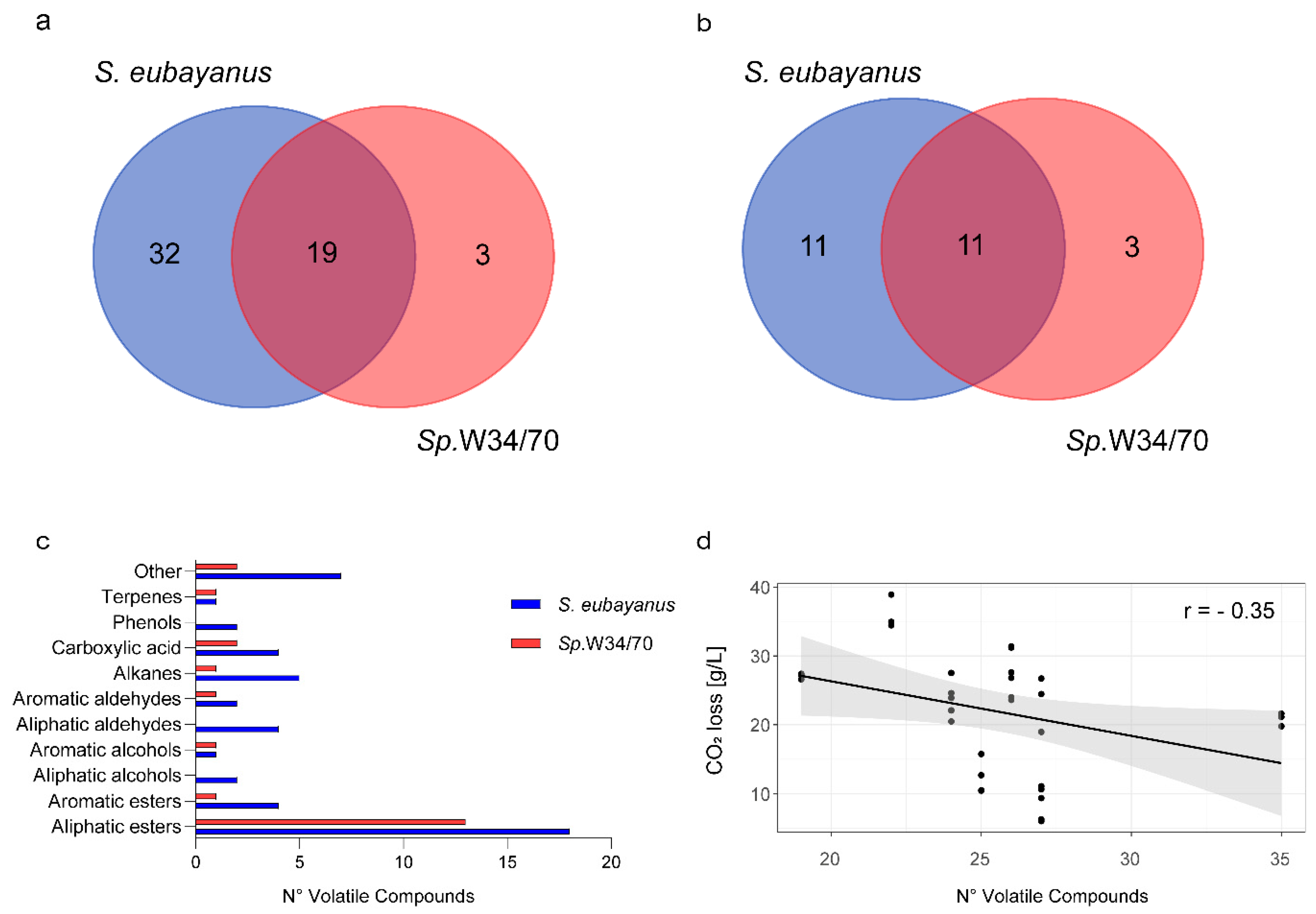
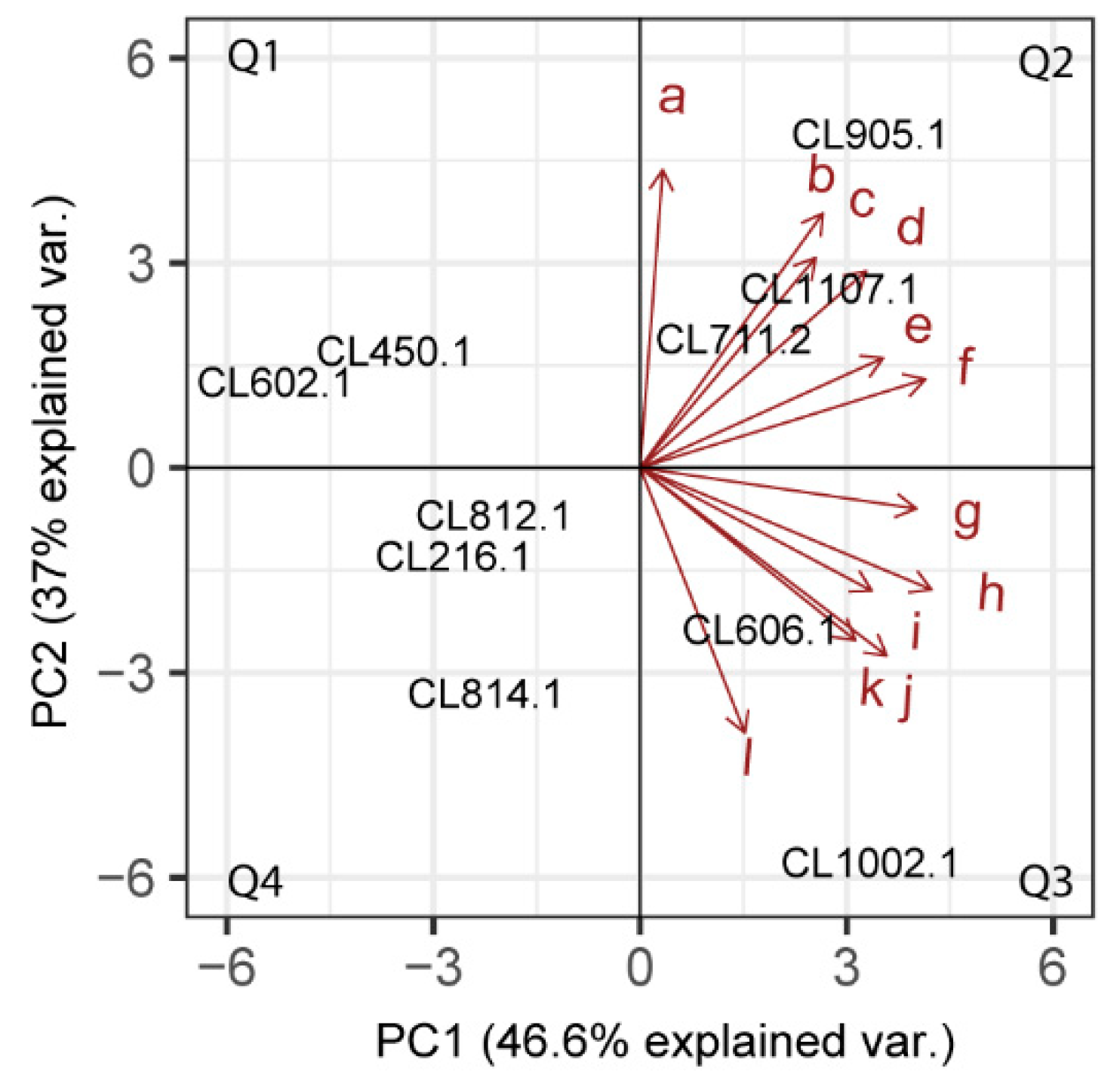
3.2. Volatile Compound Production in S. eubayanus Isolates
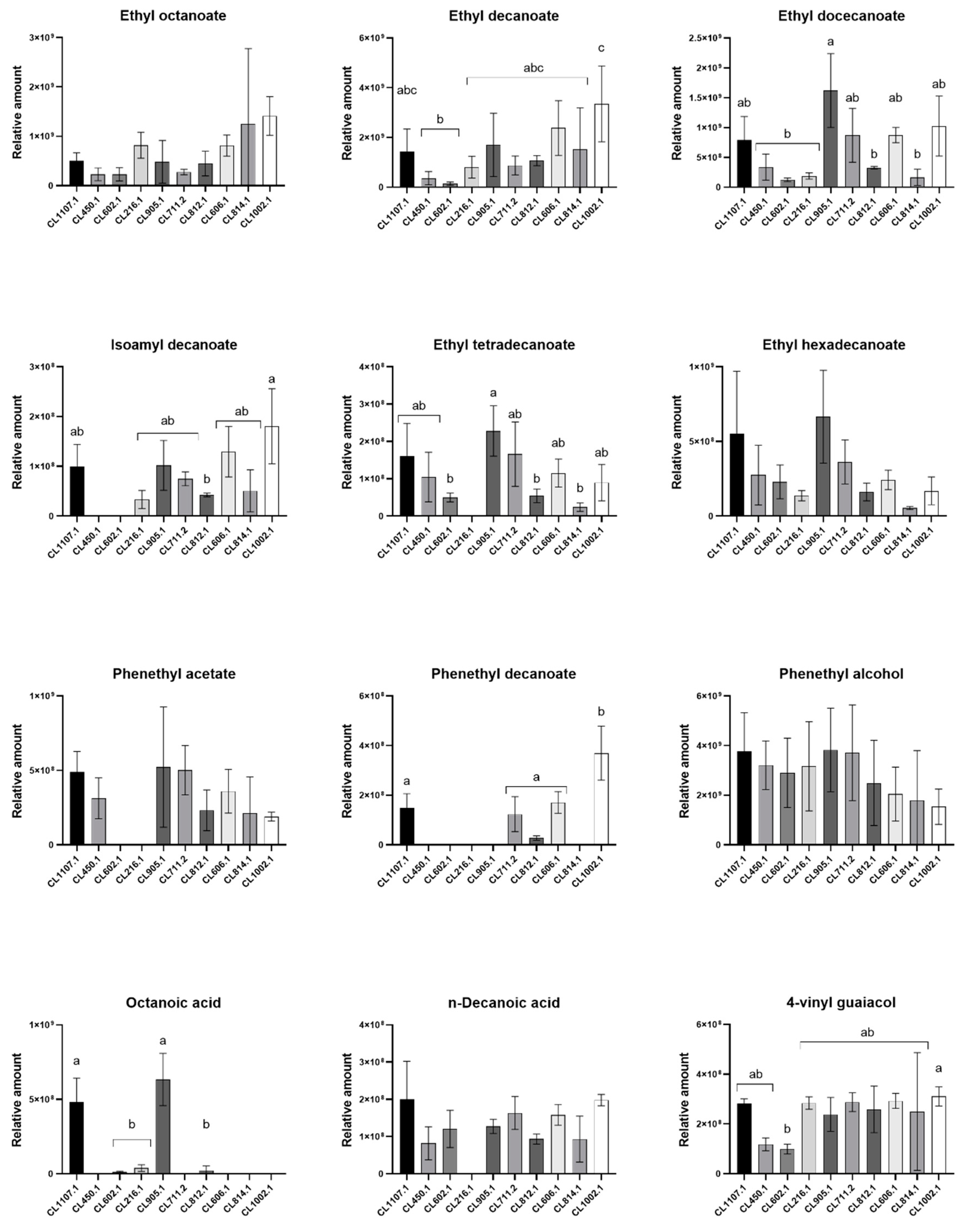
3.3. Main Volatile Compounds Identified in Fermented Wort
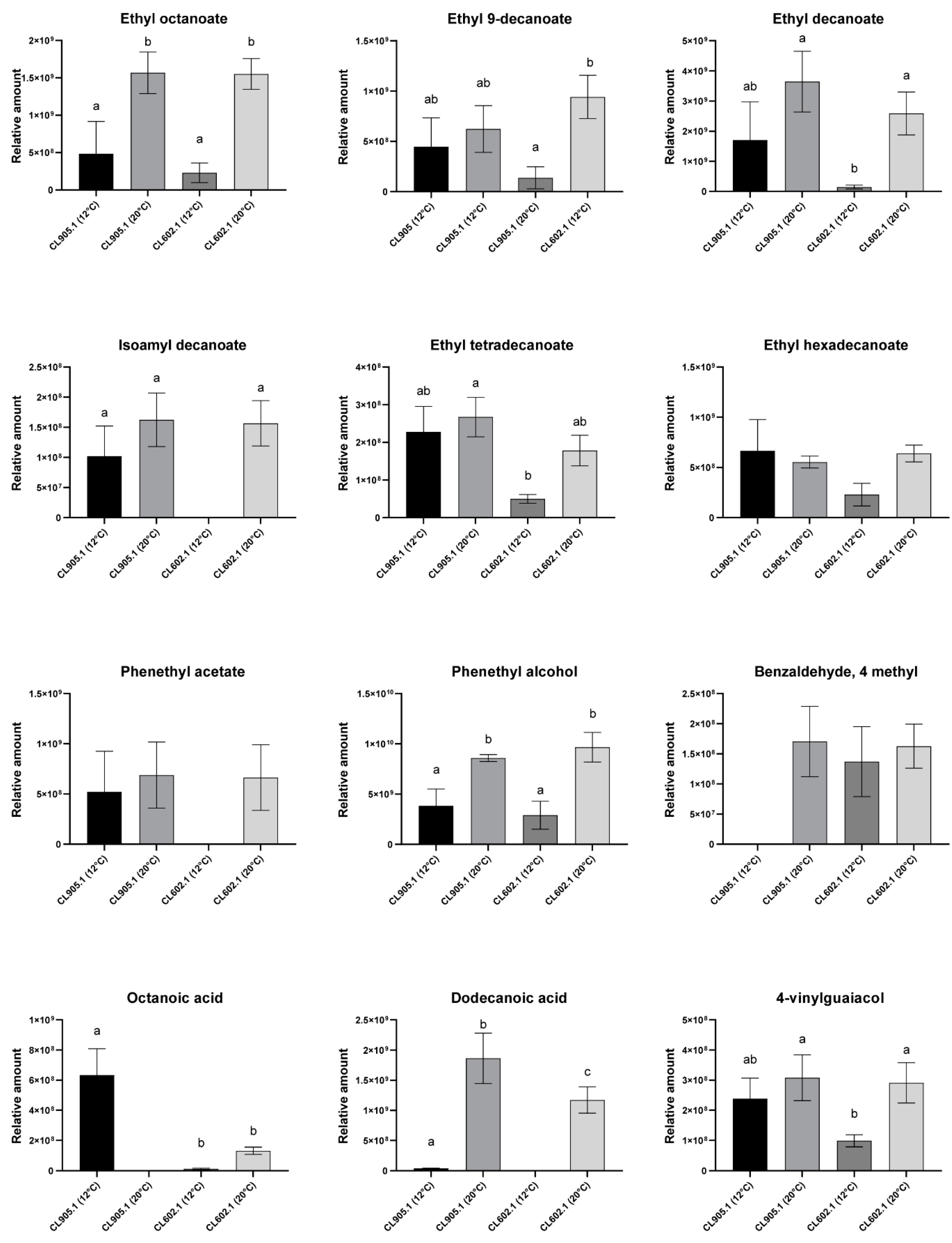
3.4. Fermentation Temperature Impacts the Volatile Compound Profile in S. eubayanus Strains
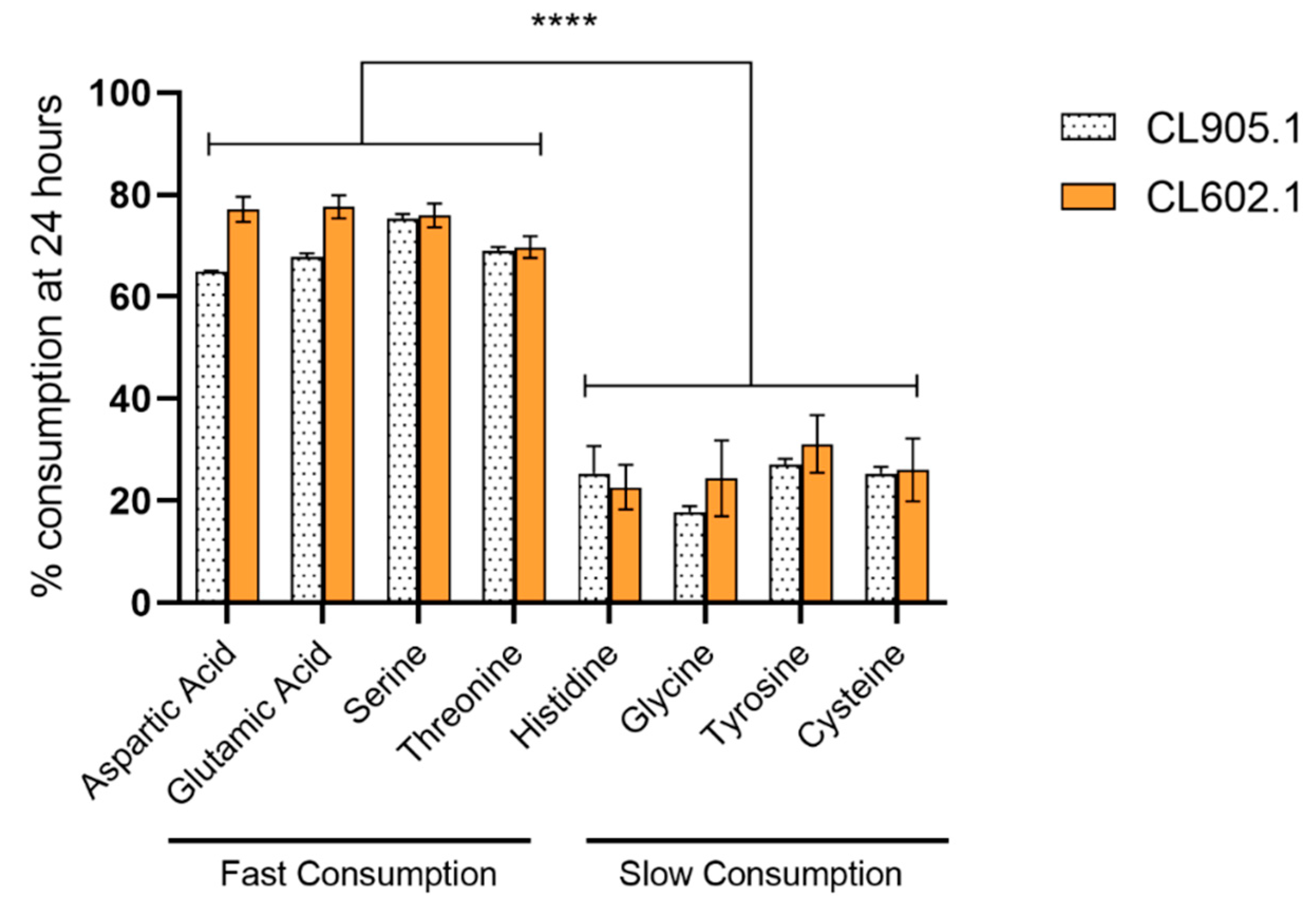
3.5. Nitrogen Consumption Differentially Impacts the Prodcution of Volatile Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Pires, E.J.; Teixeira, J.A.; Branyik, T.; Vicente, A.A. Yeast: The soul of beer’s aroma—A review of flavour-active esters and higher alcohols produced by the brewing yeast. Appl. Microbiol. Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef] [PubMed]
- Olaniran, A.O.; Hiralal, L.; Mokoena, M.P.; Pillay, B. Flavour-active volatile compounds in beer: Production, regulation and control. J. Inst. Brew. 2017, 123, 13–23. [Google Scholar] [CrossRef]
- Gallone, B.; Mertens, S.; Gordon, J.L.; Maere, S.; Verstrepen, K.J.; Steensels, J. Origins, evolution, domestication and diversity of Saccharomyces beer yeasts. Curr. Opin. Biotechnol. 2018, 49, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.; Miks, M.H.; de Carvalho, B.T.; Foulquie-Moreno, M.R.; Thevelein, J.M. The molecular biology of fruity and floral aromas in beer and other alcoholic beverages. FEMS Microbiol. Rev. 2019, 43, 193–222. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.P.; Hittinger, C.T. Evolution of a novel chimeric maltotriose transporter in Saccharomyces eubayanus from parent proteins unable to perform this function. PLoS Genet. 2019, 15, e1007786. [Google Scholar] [CrossRef]
- Gibson, B.; Geertman, J.A.; Hittinger, C.T.; Krogerus, K.; Libkind, D.; Louis, E.J.; Magalhaes, F.; Sampaio, J.P. New yeasts-new brews: Modern approaches to brewing yeast design and development. FEMS Yeast Res. 2017, 17. [Google Scholar] [CrossRef]
- Dzialo, M.C.; Park, R.; Steensels, J.; Lievens, B.; Verstrepen, K.J. Physiology, ecology and industrial applications of aroma formation in yeast. FEMS Microbiol Rev. 2017, 41, S95–S128. [Google Scholar] [CrossRef]
- Lentz, M. The Impact of Simple Phenolic Compounds on Beer Aroma and Flavor. Fermentation 2018, 4, 20. [Google Scholar] [CrossRef]
- Cubillos, F.A.; Gibson, B.; Grijalva-Vallejos, N.; Krogerus, K.; Nikulin, J. Bioprospecting for brewers: Exploiting natural diversity for naturally diverse beers. Yeast 2019, 36, 383–398. [Google Scholar] [CrossRef]
- Giannetti, V.; Boccacci Mariani, M.; Torrelli, P.; Marini, F. Flavour component analysis by HS-SPME/GC–MS and chemometric modeling to characterize Pilsner-style Lager craft beers. Microchem. J. 2019, 149. [Google Scholar] [CrossRef]
- Barbosa, C.; Mendes-Faia, A.; Mendes-Ferreira, A. The nitrogen source impacts major volatile compounds released by Saccharomyces cerevisiae during alcoholic fermentation. Int. J. Food Microbiol. 2012, 160, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Schneiderbanger, H.; Koob, J.; Poltinger, S.; Jacob, F.; Hutzler, M. Gene expression in wheat beer yeast strains and the synthesis of acetate esters. J. Inst. Brew. 2016, 122, 403–411. [Google Scholar] [CrossRef]
- Bamforth, C.W. Progress in Brewing Science and Beer Production. Annu. Rev. Chem. Biomol. Eng. 2017, 8, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Meier-Dörnberg, T.; Hutzler, M.; Michel, M.; Methner, F.-J.; Jacob, F. The Importance of a Comparative Characterization of Saccharomyces cerevisiae and Saccharomyces pastorianus Strains for Brewing. Fermentation 2017, 3, 41. [Google Scholar] [CrossRef]
- Hittinger, C.T.; Steele, J.L.; Ryder, D.S. Diverse yeasts for diverse fermented beverages and foods. Curr. Opin. Biotechnol. 2018, 49, 199–206. [Google Scholar] [CrossRef]
- Holt, S.; Mukherjee, V.; Lievens, B.; Verstrepen, K.J.; Thevelein, J.M. Bioflavoring by non-conventional yeasts in sequential beer fermentations. Food Microbiol. 2018, 72, 55–66. [Google Scholar] [CrossRef]
- Libkind, D.; Hittinger, C.T.; Valerio, E.; Goncalves, C.; Dover, J.; Johnston, M.; Goncalves, P.; Sampaio, J.P. Microbe domestication and the identification of the wild genetic stock of lager-brewing yeast. Proc. Natl. Acad. Sci. USA. 2011, 108, 14539–14544. [Google Scholar] [CrossRef]
- Peris, D.; Sylvester, K.; Libkind, D.; Goncalves, P.; Sampaio, J.P.; Alexander, W.G.; Hittinger, C.T. Population structure and reticulate evolution of Saccharomyces eubayanus and its lager-brewing hybrids. Mol. Ecol. 2014, 23, 2031–2045. [Google Scholar] [CrossRef]
- Langdon, Q.K.; Peris, D.; Eizaguirre, J.I.; Opulente, D.A.; Buh, K.V.; Sylvester, K.; Jarzyna, M.; Rodriguez, M.E.; Lopes, C.A.; Libkind, D.; et al. Postglacial migration shaped the genomic diversity and global distribution of the wild ancestor of lager-brewing hybrids. PLoS Genet. 2020, 16, e1008680. [Google Scholar] [CrossRef]
- Bing, J.; Han, P.J.; Liu, W.Q.; Wang, Q.M.; Bai, F.Y. Evidence for a Far East Asian origin of lager beer yeast. Curr. Biol. 2014, 24, R380–R381. [Google Scholar] [CrossRef]
- Gayevskiy, V.; Goddard, M.R. Saccharomyces eubayanus and Saccharomyces arboricola reside in North Island native New Zealand forests. Environ. Microbiol. 2016, 18, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Nespolo, R.F.; Villarroel, C.A.; Oporto, C.I.; Tapia, S.M.; Vega, F.; Urbina, K.; De Chiara, M.; Mozzachiodi, S.; Mikhalev, E.; Thompson, D.; et al. An Out-of-Patagonia migration explains the worldwide diversity and distribution of Saccharomyces eubayanus lineages. PLoS Genet. 2020. [Google Scholar] [CrossRef] [PubMed]
- Mardones, W.; Villarroel, C.A.; Krogerus, K.; Tapia, S.M.; Urbina, K.; Oporto, C.I.; O’Donnell, S.; Minebois, R.; Nespolo, R.; Fischer, G.; et al. Molecular profiling of beer wort fermentation diversity across natural Saccharomyces eubayanus isolates. Microb. Biotechnol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, J.P.; Goncalves, P. Natural populations of Saccharomyces kudriavzevii in Portugal are associated with oak bark and are sympatric with S. cerevisiae and S. paradoxus. Appl. Environ. Microbiol. 2008, 74, 2144–2152. [Google Scholar] [CrossRef] [PubMed]
- White, C.; Zainasheff, J. Yeast: The Practical Guide to Beer Fermentation; Brewers Publications: Boulder, CO, USA, 2010. [Google Scholar]
- Santander, R.; Creixell, W.; Sánchez, E.; Tomic, G.; Silva, J.R.; Acevedo, C.A. Recognizing Age at Slaughter of Cattle from Beef Samples Using GC/MS–SPME Chromatographic Method. Food Bioprocess. Technol. 2012, 6, 3345–3352. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Lenguage and Environment for Statisctical Computing; R. Foundation for Statistical Computing, 2008; Available online: http://finzi.psych.upenn.edu/R/library/dplR/doc/intro-dplR.pdf (accessed on 18 May 2020).
- Jara, M.; Cubillos, F.A.; Garcia, V.; Salinas, F.; Aguilera, O.; Liti, G.; Martinez, C. Mapping genetic variants underlying differences in the central nitrogen metabolism in fermenter yeasts. PLoS ONE 2014, 9, e86533. [Google Scholar] [CrossRef]
- Crepin, L.; Nidelet, T.; Sanchez, I.; Dequin, S.; Camarasa, C. Sequential use of nitrogen compounds by Saccharomyces cerevisiae during wine fermentation: A model based on kinetic and regulation characteristics of nitrogen permeases. Appl. Environ. Microbiol. 2012, 78, 8102–8111. [Google Scholar] [CrossRef]
- Gobert, A.; Tourdot-Marechal, R.; Sparrow, C.; Morge, C.; Alexandre, H. Influence of nitrogen status in wine alcoholic fermentation. Food Microbiol. 2019, 83, 71–85. [Google Scholar] [CrossRef]
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and Divergence of Saccharomyces cerevisiae Beer Yeasts. Cell 2016, 166, 1397–1410 e1316. [Google Scholar] [CrossRef]
- Bellut, K.; Arendt, E.K. Chance and Challenge: Non-Saccharomyces Yeasts in Nonalcoholic and Low Alcohol Beer Brewing—A Review. J. Am. Soc. Brew. Chem. 2019, 77, 77–91. [Google Scholar] [CrossRef]
- Rodriguez, M.E.; Orozco, H.; Cantoral, J.M.; Matallana, E.; Aranda, A. Acetyltransferase SAS2 and sirtuin SIR2, respectively, control flocculation and biofilm formation in wine yeast. FEMS Yeast Res. 2014, 14, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Pinho, O.; Ferreira, I.M.; Santos, L.H. Method optimization by solid-phase microextraction in combination with gas chromatography with mass spectrometry for analysis of beer volatile fraction. J. Chromatogr. A 2006, 1121, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Bamforth, C.W. The microbiology of malting and brewing. Microbiol. Mol. Biol. Rev. 2013, 77, 157–172. [Google Scholar] [CrossRef]
- Krogerus, K.; Magalhaes, F.; Vidgren, V.; Gibson, B. Novel brewing yeast hybrids: Creation and application. Appl. Microbiol. Biotechnol. 2017, 101, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Olaniran, A.O.; Maharaj, Y.R.; Pillay, B. Effects of fermentation temperature on the composition of beer volatile compounds, organoleptic quality and spent yeast density. Electro. J. Biotechnol. 2011, 14. [Google Scholar] [CrossRef]
- Hiralal, L.; Olaniran, A.O.; Pillay, B. Aroma-active ester profile of ale beer produced under different fermentation and nutritional conditions. J. Biosci. Bioeng. 2014, 117, 57–64. [Google Scholar] [CrossRef]
- Tokpohozin, S.E.; Fischer, S.; Becker, T. Selection of a new Saccharomyces yeast to enhance relevant sorghum beer aroma components, higher alcohols and esters. Food Microbiol. 2019, 83, 181–186. [Google Scholar] [CrossRef]
- Rossouw, D.; Naes, T.; Bauer, F.F. Linking gene regulation and the exo-metabolome: A comparative transcriptomics approach to identify genes that impact on the production of volatile aroma compounds in yeast. BMC Genom. 2008, 9, 530. [Google Scholar] [CrossRef]
- Eder, M.; Sanchez, I.; Brice, C.; Camarasa, C.; Legras, J.L.; Dequin, S. QTL mapping of volatile compound production in Saccharomyces cerevisiae during alcoholic fermentation. BMC Genom. 2018, 19, 166. [Google Scholar] [CrossRef]
- Lee, K.; Hahn, J.S. Interplay of Aro80 and GATA activators in regulation of genes for catabolism of aromatic amino acids in Saccharomyces cerevisiae. Mol. Microbiol. 2013, 88, 1120–1134. [Google Scholar] [CrossRef]
- Saerens, S.M.; Delvaux, F.; Verstrepen, K.J.; Van Dijck, P.; Thevelein, J.M.; Delvaux, F.R. Parameters affecting ethyl ester production by Saccharomyces cerevisiae during fermentation. Appl. Environ. Microbiol. 2008, 74, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Curiel, J.A.; Salvado, Z.; Tronchoni, J.; Morales, P.; Rodrigues, A.J.; Quiros, M.; Gonzalez, R. Identification of target genes to control acetate yield during aerobic fermentation with Saccharomyces cerevisiae. Microb. Cell Fact. 2016, 15, 156. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, C.; Falco, V.; Mendes-Faia, A.; Mendes-Ferreira, A. Nitrogen addition influences formation of aroma compounds, volatile acidity and ethanol in nitrogen deficient media fermented by Saccharomyces cerevisiae wine strains. J. Biosci. Bioeng. 2009, 108, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Bely, M.; Rinaldi, A.; Dubourdieu, D. Influence of assimilable nitrogen on volatile acidity production by Saccharomyces cerevisiae during high sugar fermentation. J. Biosci. Bioeng. 2003, 96, 507–512. [Google Scholar] [CrossRef]
- Beltran, G.; Novo, M.; Guillamon, J.M.; Mas, A.; Rozes, N. Effect of fermentation temperature and culture media on the yeast lipid composition and wine volatile compounds. Int. J. Food Microbiol. 2008, 121, 169–177. [Google Scholar] [CrossRef]
- Kodama, Y.; Omura, F.; Miyajima, K.; Ashikari, T. Control of Higher Alcohol Production by Manipulation of the BAP2 Gene in Brewing Yeast. J. Am. Soc. Brew. Chem. 2018, 59, 157–162. [Google Scholar] [CrossRef]
- Deed, R.C.; Deed, N.K.; Gardner, R.C. Transcriptional response of Saccharomyces cerevisiae to low temperature during wine fermentation. Antonie. Van. Leeuwenhoek. 2015, 107, 1029–1048. [Google Scholar] [CrossRef]
- Hill, A.; Stewart, G. Free Amino Nitrogen in Brewing. Fermentation 2019, 5, 22. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relative Amount (×106) and SEM | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strains | ||||||||||||
| Chemical Group | Compound Name | RI | CL1107.1 | CL1002.1 | CL905.1 | CL814.1 | CL812.1 | CL711.2 | CL606.1 | CL602.1 | CL450.1 | CL216.1 |
| Aliphatic esters | Ethyl octanoate | 1192.64 | 506.1 ± 91.8 | 1400 ± 226.4 | 483.1 ± 250.2 | 1300 ± 1100 | 446.4 ± 144 | 276.7 ± 31.7 | 812.9 ± 123.5 | 228.8 ± 76 | 230.2 ± 73.7 | 819.5 ± 151.7 |
| Ethyl decanoate | 1391.77 | 1400 ± 522.4 | 3300 ± 879.8 | 1700 ± 734.6 | 1500 ± 1200 | 1100 ± 118.7 | 870.3 ± 219.4 | 2400 ± 635.3 | 141.7 ± 38.3 | 362 ± 152.8 | 802.2 ± 253.5 | |
| Ethyl dodecanoate | 1443.83 | 793.6 ± 226 | 1000 ± 289.6 | 1600 ± 357.4 | 164.5 ± 96.9 | 325.8 ± 11.7 | 868.5 ± 260.4 | 872.2 ± 73.8 | 123.7 ± 17 | 335.3 ± 125.3 | 186 ± 30.2 | |
| Isoamyl decanoate | 1590.63 | 99.7 ± 25.7 | 180.5 ± 43.5 | 102.1 ± 28.9 | 50.8 ± 30 | 42.4 ± 2.1 | 74.9 ± 8.1 | 129.4 ± 29.3 | N/D | N/D | 33.5 ± 10.5 | |
| Ethyl tetradecanoate | 1790.87 | 160.3 ± 50.9 | 89.8 ± 28.2 | 228.2 ± 38.9 | 23.7 ± 8 | 54.3 ± 10.5 | 166 ± 50 | 115.2 ± 21.6 | 49.7 ± 6.8 | 104.5 ± 38.3 | N/D | |
| Ethyl hexadecanoate | 1991.92 | 552 ± 241.4 | 169 ± 53.7 | 666 ± 179.7 | 53.4 ± 7.4 | 161.3 ± 34.4 | 363 ± 85.2 | 242.5 ± 37.5 | 229.4 ± 65.1 | 274.3 ± 115.2 | 136.9 ± 20 | |
| Aromatic esters | Phenethyl acetate | 1250.12 | 490.8 ± 79.1 | 189.8 ± 17 | 522.4 ± 233.1 | 212.9 ± 172.2 | 232.1 ± 78.9 | 501.7 ± 95.5 | 360.2 ± 84.7 | N/D | N/D | 312.6 ± 79.2 |
| Phenethyl decanoate | 2062.65 | 149.5 ± 32.8 | 369.7 ± 62.8 | N/D | N/D | 27.7 ± 5.3 | 123.7 ± 40.7 | 170.8 ± 25.3 | N/D | N/D | N/D | |
| Aromatic alcohol | Phenethyl Alcohol | 1116.19 | 2900 ± 1600 | 1500 ± 409.3 | 3800 ± 971.5 | 1800 ± 1400 | 2500 ± 992.4 | 3700 ± 1100 | 2000 ± 627 | 2900 ± 805 | 3200 ± 565.2 | 3200 ± 1000 |
| Carboxylic acid | Octanoic acid | 1171.39 | 483 ± 91.8 | N/D | 633.5 ± 101.3 | N/D | 22.9 ± 17.4 | N/D | N/D | 12.5 ± 2.4 | N/D | 39 ± 13.1 |
| n-Decanoic acid | 1360.8 | 200 ± 59 | 198 ± 8.8 | 127.5 ± 11 | 93.3 ± 43.8 | 93.4 ± 7.8 | 163.3 ± 25.6 | 158.3 ± 16.1 | N/D | 120.4 ± 28.9 | 81.9 ± 25.6 | |
| Phenols | 4-vinylguaiacol | 1317.45 | 281.4 ± 11.2 | 310.9 ± 22.3 | 238.7 ± 39.4 | 250.1 ± 167.4 | 258.7 ± 53.9 | 287.5 ± 21.8 | 292.7 ± 17.6 | 98.8 ± 11.4 | 118.1 ± 15 | 284.1 ± 14.5 |
| Relative Abundance (×106) and SEM | ||||||
|---|---|---|---|---|---|---|
| Strain | ||||||
| CL905.1 | CL602.1 | |||||
| Chemical Group | Compound | RI | 12 °C | 20 °C | 12 °C | 20 °C |
| Aliphatic esters | Ethyl octanoate | 1192.63881 | 483.1 ± 250.2 | 1566.67 ± 251.66 | 228.8 ± 76 | 1566.67 ± 120.19 |
| Ethyl 9-decenoate | 1384.56268 | 448.5 ± 165.1 | 622.93 ± 232.23 | 137.5 ± 63.5 | 622.93 ± 116.55 | |
| Ethyl decanoate | 1391.77014 | 1700 ± 734.6 | 3666.67 ± 1011.60 | 141.7 ± 38.3 | 3666.67 ± 409.61 | |
| Isoamyl decanoate | 1644.71354 | 102.1 ± 28.9 | 162.40 ± 44.51 | N/D | 162.40 ± 21.72 | |
| Ethyl tetradecanoate | 1786.82537 | 228.2 ± 38.9 | 267.33 ± 52.60 | 49.7 ± 6.8 | 267.33 ± 23.47 | |
| Ethyl hexadecanoate | 1991.92208 | 666 ± 179.7 | 553.43 ± 59.25 | N/D | 553.43 ± 48.51 | |
| Aromatic esters | Phenethyl acetate | 1250.12471 | 522.4 ± 233.1 | 687.80 ± 328.51 | N/D | 687.80 ± 189.00 |
| Aromatic alcohols | Phenethyl Alcohol | 1116.18753 | 3800 ± 971.5 | 8600.00 ± 346.41 | 2900 ± 805 | 8600.00 ± 851.14 |
| Aromatic aldehydes | Benzaldehyde, 4-methyl | 1083.5138 | N/D | 170.50 ± 58.41 | 42.1 ± 6 | 170.50 ± 21.10 |
| Carboxylic acids | Octanoic acid | 1171.38687 | 633.5 ± 101.3 | N/D | 12.5 ± 2.4 | 131.80 ± 13.44 |
| Dodecanoic acid | 1556.73878 | 38.3 ± 3.4 | 1866.67 ± 404.15 | N/D | 1866.67 ± 135.36 | |
| Phenols | 4-vinylguaiacol | 1317.44761 | 238.7 ± 39.4 | 308.03 ± 76.01 | 98.8 ± 11.4 | 290.93 ± 38.64 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urbina, K.; Villarreal, P.; Nespolo, R.F.; Salazar, R.; Santander, R.; Cubillos, F.A. Volatile Compound Screening Using HS-SPME-GC/MS on Saccharomyces eubayanus Strains under Low-Temperature Pilsner Wort Fermentation. Microorganisms 2020, 8, 755. https://doi.org/10.3390/microorganisms8050755
Urbina K, Villarreal P, Nespolo RF, Salazar R, Santander R, Cubillos FA. Volatile Compound Screening Using HS-SPME-GC/MS on Saccharomyces eubayanus Strains under Low-Temperature Pilsner Wort Fermentation. Microorganisms. 2020; 8(5):755. https://doi.org/10.3390/microorganisms8050755
Chicago/Turabian StyleUrbina, Kamila, Pablo Villarreal, Roberto F. Nespolo, Ricardo Salazar, Rocio Santander, and Francisco A. Cubillos. 2020. "Volatile Compound Screening Using HS-SPME-GC/MS on Saccharomyces eubayanus Strains under Low-Temperature Pilsner Wort Fermentation" Microorganisms 8, no. 5: 755. https://doi.org/10.3390/microorganisms8050755
APA StyleUrbina, K., Villarreal, P., Nespolo, R. F., Salazar, R., Santander, R., & Cubillos, F. A. (2020). Volatile Compound Screening Using HS-SPME-GC/MS on Saccharomyces eubayanus Strains under Low-Temperature Pilsner Wort Fermentation. Microorganisms, 8(5), 755. https://doi.org/10.3390/microorganisms8050755