Revealing Fungal Communities in Alpine Wetlands through Species Diversity, Functional Diversity and Ecological Network Diversity

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Sample Collection
2.2. Soil Properties Analyses
2.3. DNA Extraction, PCR Amplification and Sequence Data Preprocessing
2.4. Statistical Analysis
3. Results
3.1. Geographical Parameters and Soil Properties
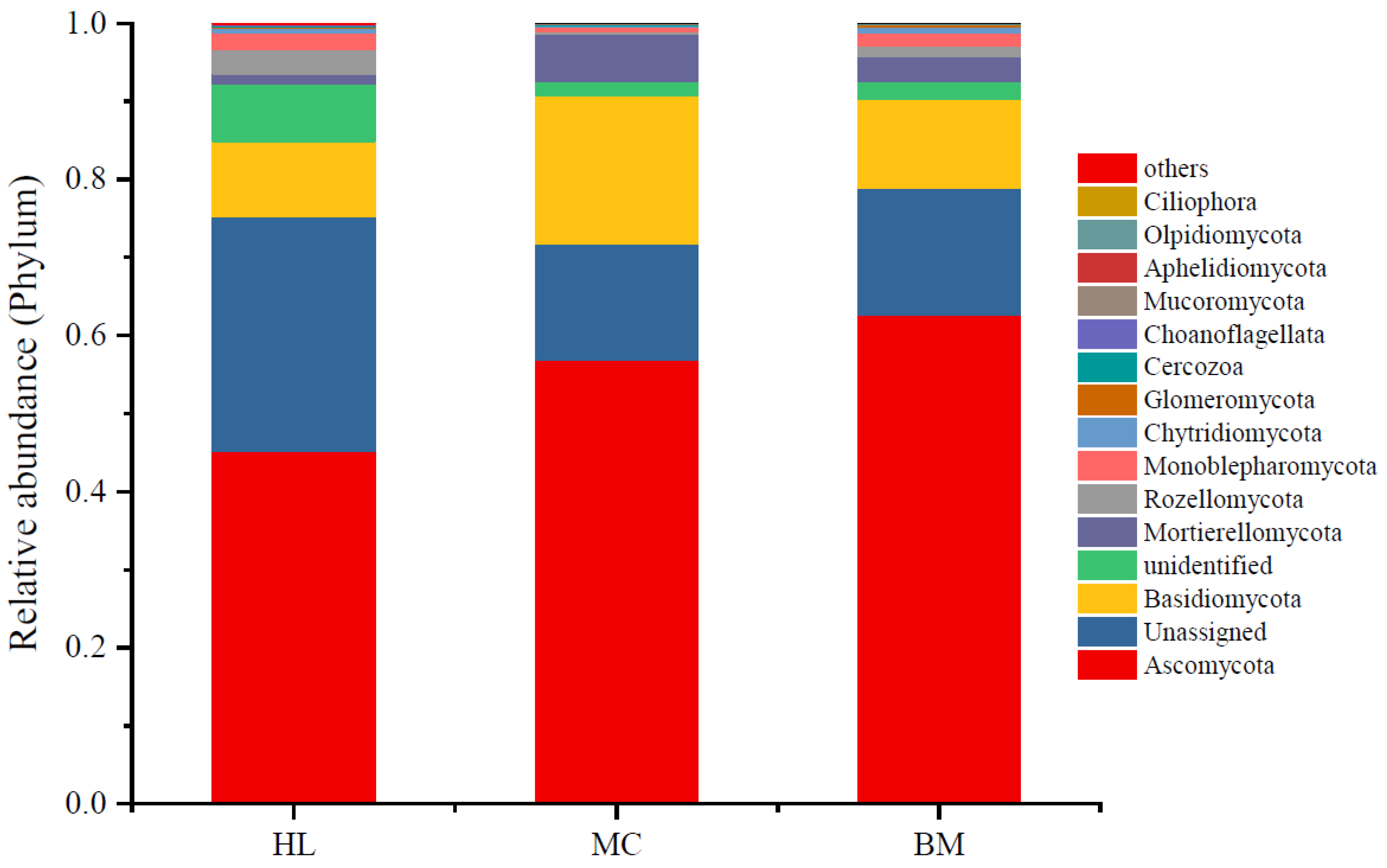
3.2. Fungal Community Composition
3.3. Fungal Community Diversity
3.4. The Trophic Strategies of Fungal Communities
3.5. Analysis of Fungal Ecological Network
3.6. Geographic Sampling Location, Soil Properties and Fungal Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Geml, J.; Morgado, L.N.; Semenova-Nelsen, T.A.; Schilthuizen, M. Changes in richness and community composition of ectomycorrhizal fungi among altitudinal vegetation types on Mount Kinabalu in Borneo. New Phytol. 2017, 215, 454–468. [Google Scholar] [CrossRef] [PubMed]
- Weißbecker, C.; Heintz-Buschart, A.; Bruelheide, H.; Buscot, F.; Wubet, T. Linking Soil Fungal Generality to Tree Richness in Young Subtropical Chinese Forests. Microorganisms 2019, 7, 547. [Google Scholar]
- Gehring, C.A.; Wolf, J.E.; Theimer, T.C. Terrestrial vertebrates promote arbuscular mycorrhizal fungal diversity and inoculum potential in a rain forest soil. Ecol. Lett. 2002, 5, 540–548. [Google Scholar] [CrossRef]
- Johnson, D.; Vandenkoornhuyse, P.J.; Leake, J.R.; Gilbert, L.; Booth, R.E.; Grime, J.P.; Young, J.P.W.; Read, D.J. Plant communities affect arbuscular mycorrhizal fungal diversity and community composition in grassland microcosms. New Phytol. 2004, 161, 503–515. [Google Scholar] [CrossRef]
- Wang, K.B.; Zhang, Y.W.; Tang, Z.S.; Shangguan, Z.P.; Chang, F.; Jia, F.A.; Chen, Y.P.; He, X.H.; Shi, W.Y.; Deng, L. Effects of grassland afforestation on structure and function of soil bacterial and fungal communities. Sci. Total Environ. 2019, 676, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Cullings, K.; Makhija, S. Ectomycorrhizal fungal associates of Pinus contorta in soils associated with a hot spring in Norris Geyser Basin, Yellowstone National Park, Wyoming. Appl. Environ. Microbiol. 2001, 67, 5538–5543. [Google Scholar] [CrossRef] [PubMed]
- Massini, J.G.; Channing, A.; Guido, D.M.; Zamuner, A.B. First report of fungi and fungus-like organisms from Mesozoic hot springs. Palaios 2012, 27, 55–62. [Google Scholar] [CrossRef]
- Chen, Y.L.; Deng, Y.; Ding, J.Z.; Hu, H.W.; Xu, T.L.; Li, F.; Yang, G.B.; Yang, Y.H. Distinct microbial communities in the active and permafrost layers on the Tibetan Plateau. Mol. Ecol. 2017, 26, 6608–6620. [Google Scholar] [CrossRef]
- Hu, W.; Zhang, Q.; Li, D.; Cheng, G.; Mu, J.; Wu, Q.; Niu, F.; An, L.; Feng, H. Diversity and community structure of fungi through a permafrost core profile from the Qinghai-Tibet Plateau of China. J. Basic Microbiol. 2014, 54, 1331–1341. [Google Scholar] [CrossRef]
- Bohrer, K.E.; Friese, C.F.; Amon, J.P. Seasonal dynamics of arbuscular mycorrhizal fungi in differing wetland habitats. Mycorrhiza 2004, 14, 329–337. [Google Scholar] [CrossRef]
- Wolfe, B.E.; Weishampel, P.A.; Klironomos, J.N. Arbuscular mycorrhizal fungi and water table affect wetland plant community composition. J. Ecol. 2006, 94, 905–914. [Google Scholar] [CrossRef]
- Averill, C.; Bhatnagar, J.M.; Dietze, M.C.; Pearse, W.D.; Kivlin, S.N. Global imprint of mycorrhizal fungi on whole-plant nutrient economics. Proc. Natl. Acad. Sci. USA 2019, 116, 23163–23168. [Google Scholar] [CrossRef]
- Clemmensen, K.E.; Finlay, R.D.; Dahlberg, A.; Stenlid, J.; Wardle, D.A.; Lindahl, B.D. Carbon sequestration is related to mycorrhizal fungal community shifts during long-term succession in boreal forests. New Phytol. 2015, 205, 1525–1536. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, E.A.; Horton, T.R. Evidence that saprotrophic fungi mobilise carbon and mycorrhizal fungi mobilise nitrogen during litter decomposition. New Phytol. 2007, 173, 447–449. [Google Scholar] [CrossRef] [PubMed]
- Ma, A.; Zhuang, X.; Wu, J.; Cui, M.; Lv, D.; Liu, C.; Zhuang, G. Ascomycota members dominate fungal communities during straw residue decomposition in arable soil. PLoS ONE 2013, 8, e66146. [Google Scholar] [CrossRef] [PubMed]
- Thormann, M.N. Diversity and function of fungi in peatlands: A carbon cycling perspective. Can. J. Soil Sci. 2006, 86, 281–293. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Root, R.B. The niche exploitation pattern of the blue-gray gnatcatcher. Ecol. Monogr. 1967, 37, 317–350. [Google Scholar] [CrossRef]
- Kyaschenko, J.; Clemmensen, K.E.; Hagenbo, A.; Karltun, E.; Lindahl, B.D. Shift in fungal communities and associated enzyme activities along an age gradient of managed Pinus sylvestris stands. ISME J. 2017, 11, 863–874. [Google Scholar] [CrossRef]
- Frąc, M.; Hannula, S.E.; Bełka, M.; Jędryczka, M. Fungal biodiversity and their role in soil health. Front. Microbiol. 2018, 9, 707. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A.; Horton, T.R. Socialism in soil? The importance of mycorrhizal fungal networks for facilitation in natural ecosystems. J. Ecol. 2009, 97, 1139–1150. [Google Scholar] [CrossRef]
- White, J.F.; Kingsley, K.L.; Verma, S.K.; Kowalski, K.P. Rhizophagy cycle: An oxidative process in plants for nutrient extraction from symbiotic microbes. Microorganisms 2018, 6, 95. [Google Scholar] [CrossRef] [PubMed]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and associated fungi drive long-term carbon sequestration in boreal forest. Science 2013, 339, 1615–1618. [Google Scholar] [CrossRef] [PubMed]
- Fincham, W.N.W.; Dunn, A.M.; Brown, L.E.; Hesketh, H.; Roy, H.E. Invasion success of a widespread invasive predator may be explained by a high predatory efficacy but may be influenced by pathogen infection. Biol. Invasions 2019, 21, 3545–3560. [Google Scholar] [CrossRef]
- Liu, L.; Yu, S.; Xie, Z.-P.; Staehelin, C. Distance-dependent effects of pathogenic fungi on seedlings of a legume tree: Impaired nodule formation and identification of antagonistic rhizosphere bacteria. J. Ecol. 2016, 104, 1009–1019. [Google Scholar] [CrossRef]
- Moisan, K.; Cordovez, V.; van de Zande, E.M.; Raaijmakers, J.M.; Dicke, M.; Lucas-Barbosa, D. Volatiles of pathogenic and non-pathogenic soil-borne fungi affect plant development and resistance to insects. Oecologia 2019, 190, 589–604. [Google Scholar] [CrossRef]
- Tofangsazi, N.; Hogg, B.N.; Portman, S.L.; Pratt, P.D. Tritrophic Interactions Between an Invasive Weed (Lepidium latifolium), an Insect Herbivore (Bagrada hilaris), and a Plant Pathogenic Fungus (Albugo lepidii). Environ. Entomol. 2019, 48, 1317–1322. [Google Scholar] [CrossRef]
- Viswanathan, A.; Ghazoul, J.; Honwad, G.; Kumar, N.A.; Bagchi, R. The effects of rainforest fragment area on the strength of plant—Pathogen interactions. Biol. Lett. 2019, 15. [Google Scholar] [CrossRef]
- He, D.; Shen, W.; Eberwein, J.; Zhao, Q.; Ren, L.; Wu, Q.L. Diversity and co-occurrence network of soil fungi are more responsive than those of bacteria to shifts in precipitation seasonality in a subtropical forest. Soil Biol. Biochem. 2017, 115, 499–510. [Google Scholar] [CrossRef]
- Wagg, C.; Schlaeppi, K.; Banerjee, S.; Kuramae, E.E.; van der Heijden, M.G. Fungal-bacterial diversity and microbiome complexity predict ecosystem functioning. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, Y.; Luo, F.; He, Z.; Yang, Y. Phylogenetic molecular ecological network of soil microbial communities in response to elevated CO2. Mbio 2011, 2, e00122-11. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Jiang, Y.-H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, J.A.; Steele, J.A. Community structure of marine bacterioplankton: Patterns, networks, and relationships to function. Aquat. Microb. Ecol. 2008, 53, 69–81. [Google Scholar] [CrossRef]
- Martiny, J.B.H.; Bohannan, B.J.M.; Brown, J.H.; Colwell, R.K.; Fuhrman, J.A.; Green, J.L.; Horner-Devine, M.C.; Kane, M.; Krumins, J.A.; Kuske, C.R.; et al. Microbial biogeography: Putting microorganisms on the map. Nat. Rev. Microbiol. 2006, 4, 102–112. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, M.A. The nineteenth century roots of ‘everything is everywhere’. Nat. Rev. Microbiol. 2007, 5, 647–651. [Google Scholar] [CrossRef] [PubMed]
- Qi, D.; Guo, S.; Zhao, X.; Yang, J.; Tangi, W. Genetic diversity and historical population structure of Schizopygopsis pylzovi (Teleostei: Cyprinidae) in the Qinghai-Tibetan Plateau. Freshw. Biol. 2007, 52, 1090–1104. [Google Scholar] [CrossRef]
- Zhang, Y.; Cui, M.; Duan, J.; Zhuang, X.; Zhuang, G.; Ma, A. Abundance, rather than composition, of methane-cycling microbes mainly affects methane emissions from different vegetation soils in the Zoige alpine wetland. Microbiologyopen 2019, 8, e00699. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, A.; Liu, W.; Bai, Z.; Zhuang, X.; Zhuang, G. The occurrence of putative nitric oxide dismutase (Nod) in an alpine wetland with a new dominant subcluster and the potential ability for a methane sink. Archaea Int. Microbiol. J. 2018. [Google Scholar] [CrossRef]
- Wang, L.; Abbott, R.J.; Zheng, W.; Chen, P.; Wang, Y.; Liu, J. History and evolution of alpine plants endemic to the Qinghai-Tibetan Plateau: Aconitum gymnandrum (Ranunculaceae). Mol. Ecol. 2009, 18, 709–721. [Google Scholar] [CrossRef]
- Starke, R.; Capek, P.; Morais, D.; Callister, S.J.; Jehmlich, N. The total microbiome functions in bacteria and fungi. J. Proteom. 2020, 213, 5. [Google Scholar] [CrossRef]
- Ren, G.; Ma, A.; Zhang, Y.; Deng, Y.; Zheng, G.; Zhuang, X.; Zhuang, G.; Fortin, D. Electron acceptors for anaerobic oxidation of methane drive microbial community structure and diversity in mud volcanoes. Environ. Microbiol. 2018, 20, 2370–2385. [Google Scholar] [CrossRef] [PubMed]
- Che, R.; Wang, S.; Wang, Y.; Xu, Z.; Wang, W.; Rui, Y.; Wang, F.; Hu, J.; Tao, J.; Cui, X. Total and active soil fungal community profiles were significantly altered by six years of warming but not by grazing. Soil Biol. Biochem. 2019, 139, 107611. [Google Scholar] [CrossRef]
- Malik, A.A.; Chowdhury, S.; Schlager, V.; Oliver, A.; Puissant, J.; Vazquez, P.G.M.; Jehmlich, N.; von Bergen, M.; Griffiths, R.I.; Gleixner, G. Soil fungal: Bacterial ratios are linked to altered carbon cycling. Front. Microbiol. 2016, 7, 1247. [Google Scholar] [CrossRef]
- Leifheit, E.F.; Verbruggen, E.; Rillig, M.C. Arbuscular mycorrhizal fungi reduce decomposition of woody plant litter while increasing soil aggregation. Soil Biol. Biochem. 2015, 81, 323–328. [Google Scholar] [CrossRef]
- Wilson, G.W.T.; Rice, C.W.; Rillig, M.C.; Springer, A.; Hartnett, D.C. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: Results from long-term field experiments. Ecol. Lett. 2009, 12, 452–461. [Google Scholar] [CrossRef]
- Guimera, R.; Sales-Pardo, M.; Amaral, L.A.N. Classes of complex networks defined by role-to-role connectivity profiles. Nat. Phys. 2007, 3, 63–69. [Google Scholar] [CrossRef]
- Berry, D.; Widder, S. Deciphering microbial interactions and detecting keystone species with co-occurrence networks. Front. Microbiol. 2014, 5, 219. [Google Scholar] [CrossRef]
- Wu, B.; Tian, J.; Bai, C.; Xiang, M.; Sun, J.; Liu, X. The biogeography of fungal communities in wetland sediments along the Changjiang River and other sites in China. ISME J. 2013, 7, 1299–1309. [Google Scholar] [CrossRef]
- Ge, Y.; He, J.-Z.; Zhu, Y.-G.; Zhang, J.-B.; Xu, Z.; Zhang, L.-M.; Zheng, Y.-M. Differences in soil bacterial diversity: Driven by contemporary disturbances or historical contingencies? ISME J. 2008, 2, 254–264. [Google Scholar] [CrossRef]
- Williams, S.E.; Marsh, H.; Winter, J. Spatial scale, species diversity, and habitat structure: Small mammals in Australian tropical rain forest. Ecology 2002, 83, 1317–1329. [Google Scholar] [CrossRef]
- Wang, F.P.; Shi, G.X.; Nicholas, O.; Yao, B.Q.; Ji, M.F.; Wang, W.Y.; Ma, Z.; Zhou, H.K.; Zhao, X.Q. Ecosystem nitrogen retention is regulated by plant community trait interactions with nutrient status in an alpine meadow. J. Ecol. 2018, 106, 1570–1581. [Google Scholar] [CrossRef]
- Wu, J.; Li, N.; Wang, X. Nitrogen deposition strengthens the relationship between plants and the soil fungal community in alpine steppe, North Tibet. Appl. Soil Ecol. 2020, 147, 103441. [Google Scholar]
- Cassman, N.A.; Leite, M.F.; Pan, Y.; De Hollander, M.; Van Veen, J.A.; Kuramae, E.E. Plant and soil fungal but not soil bacterial communities are linked in long-term fertilized grassland. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wooliver, R.C.; Senior, J.K.; Potts, B.M.; Van Nuland, M.E.; Bailey, J.K.; Schweitzer, J.A. Soil fungi underlie a phylogenetic pattern in plant growth responses to nitrogen enrichment. J. Ecol. 2018, 106, 2161–2175. [Google Scholar] [CrossRef]
- Zachow, C.; Berg, C.; Müller, H.; Meincke, R.; Komon-Zelazowska, M.; Druzhinina, I.S.; Kubicek, C.P.; Berg, G. Fungal diversity in the rhizosphere of endemic plant species of Tenerife (Canary Islands): Relationship to vegetation zones and environmental factors. ISME J. 2009, 3, 79–92. [Google Scholar] [CrossRef]
- Nevarez, L.; Vasseur, V.; Le Madec, A.; Le Bras, M.; Coroller, L.; Leguérinel, I.; Barbier, G. Physiological traits of Penicillium glabrum strain LCP 08.5568, a filamentous fungus isolated from bottled aromatised mineral water. Int. J. Food Microbiol. 2009, 130, 166–171. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | MC (%) | TC (g/kg) | TN (g/kg) | TP (g/kg) | AN (mg/kg) | NN (mg/kg) | C/N Rate | |
|---|---|---|---|---|---|---|---|---|
| HL1 | 7.00 | 136.37 | 170.83 | 13.89 | 0.80 | 0.76 | 1.33 | 12.30 |
| HL2 | 6.97 | 200.47 | 172.35 | 13.61 | 0.90 | 1.17 | 1.10 | 12.66 |
| HL3 | 7.02 | 162.36 | 112.62 | 8.65 | 0.87 | 0.86 | 0.69 | 13.02 |
| HL4 | 7.02 | 159.86 | 117.17 | 8.90 | 0.90 | 1.27 | 0.66 | 13.17 |
| HL5 | 7.11 | 163.56 | 128.61 | 12.06 | 0.98 | 1.22 | 0.95 | 10.66 |
| HL6 | 7.00 | 162.31 | 150.29 | 12.10 | 0.87 | 0.71 | 0.61 | 12.42 |
| HL7 | 6.99 | 150.57 | 164.77 | 12.12 | 0.90 | 1.19 | 1.42 | 13.59 |
| HL8 | 7.21 | 182.62 | 130.36 | 8.83 | 0.55 | 0.85 | 0.66 | 14.76 |
| MC1 | 5.82 | 88.74 | 55.05 | 5.19 | 1.30 | 0.61 | 1.07 | 10.61 |
| MC2 | 6.11 | 54.62 | 43.10 | 3.84 | 0.95 | 0.54 | 1.52 | 11.22 |
| MC3 | 5.53 | 47.06 | 38.45 | 3.39 | 1.02 | 0.40 | 2.62 | 11.34 |
| MC4 | 6.09 | 41.58 | 36.95 | 3.28 | 0.94 | 0.44 | 1.35 | 11.27 |
| MC5 | 5.41 | 79.61 | 59.53 | 5.53 | 0.96 | 0.52 | 2.59 | 10.76 |
| MC6 | 5.42 | 98.63 | 33.26 | 5.75 | 1.20 | 0.51 | 1.83 | 5.78 |
| MC7 | 5.37 | 71.94 | 51.35 | 4.96 | 0.87 | 0.47 | 1.50 | 10.35 |
| MC8 | 5.75 | 86.20 | 58.05 | 5.44 | 0.73 | 0.63 | 1.49 | 10.67 |
| BM1 | 6.43 | 46.31 | 19.53 | 2.12 | 1.04 | 3.12 | 0.23 | 9.21 |
| BM2 | 6.48 | 41.90 | 16.38 | 1.99 | 1.11 | 4.94 | 0.33 | 8.23 |
| BM3 | 6.57 | 63.22 | 27.43 | 2.94 | 0.94 | 6.39 | 0.29 | 9.33 |
| BM4 | 5.76 | 80.70 | 37.05 | 3.67 | 1.05 | 5.02 | 0.17 | 10.10 |
| BM5 | 6.16 | 69.98 | 26.23 | 2.85 | 1.17 | 6.32 | 0.22 | 9.20 |
| BM6 | 6.67 | 42.05 | 15.73 | 1.94 | 0.69 | 4.87 | 0.19 | 8.11 |
| BM7 | 6.00 | 65.41 | 30.53 | 3.41 | 1.08 | 5.21 | 0.19 | 8.95 |
| BM8 | 6.60 | 56.40 | 21.08 | 2.62 | 1.29 | 3.68 | 0.24 | 8.05 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, F.; Ma, A.; Zhou, H.; Liang, Y.; Yin, J.; Ma, K.; Zhuang, X.; Zhuang, G. Revealing Fungal Communities in Alpine Wetlands through Species Diversity, Functional Diversity and Ecological Network Diversity. Microorganisms 2020, 8, 632. https://doi.org/10.3390/microorganisms8050632
Xie F, Ma A, Zhou H, Liang Y, Yin J, Ma K, Zhuang X, Zhuang G. Revealing Fungal Communities in Alpine Wetlands through Species Diversity, Functional Diversity and Ecological Network Diversity. Microorganisms. 2020; 8(5):632. https://doi.org/10.3390/microorganisms8050632
Chicago/Turabian StyleXie, Fei, Anzhou Ma, Hanchang Zhou, Yu Liang, Jun Yin, Ke Ma, Xuliang Zhuang, and Guoqiang Zhuang. 2020. "Revealing Fungal Communities in Alpine Wetlands through Species Diversity, Functional Diversity and Ecological Network Diversity" Microorganisms 8, no. 5: 632. https://doi.org/10.3390/microorganisms8050632
APA StyleXie, F., Ma, A., Zhou, H., Liang, Y., Yin, J., Ma, K., Zhuang, X., & Zhuang, G. (2020). Revealing Fungal Communities in Alpine Wetlands through Species Diversity, Functional Diversity and Ecological Network Diversity. Microorganisms, 8(5), 632. https://doi.org/10.3390/microorganisms8050632