Drosophila Sexual Attractiveness in Older Males Is Mediated by Their Microbiota


Abstract
1. Introduction
2. Materials and Methods
2.1. Manipulation of the Microbiota
2.2. Preference in No Choice Mating Trials
2.3. Preference in Choice (Competitive Mating) Mating Trials
2.4. Measurement of Male Activity
2.5. Data Analysis
3. Results
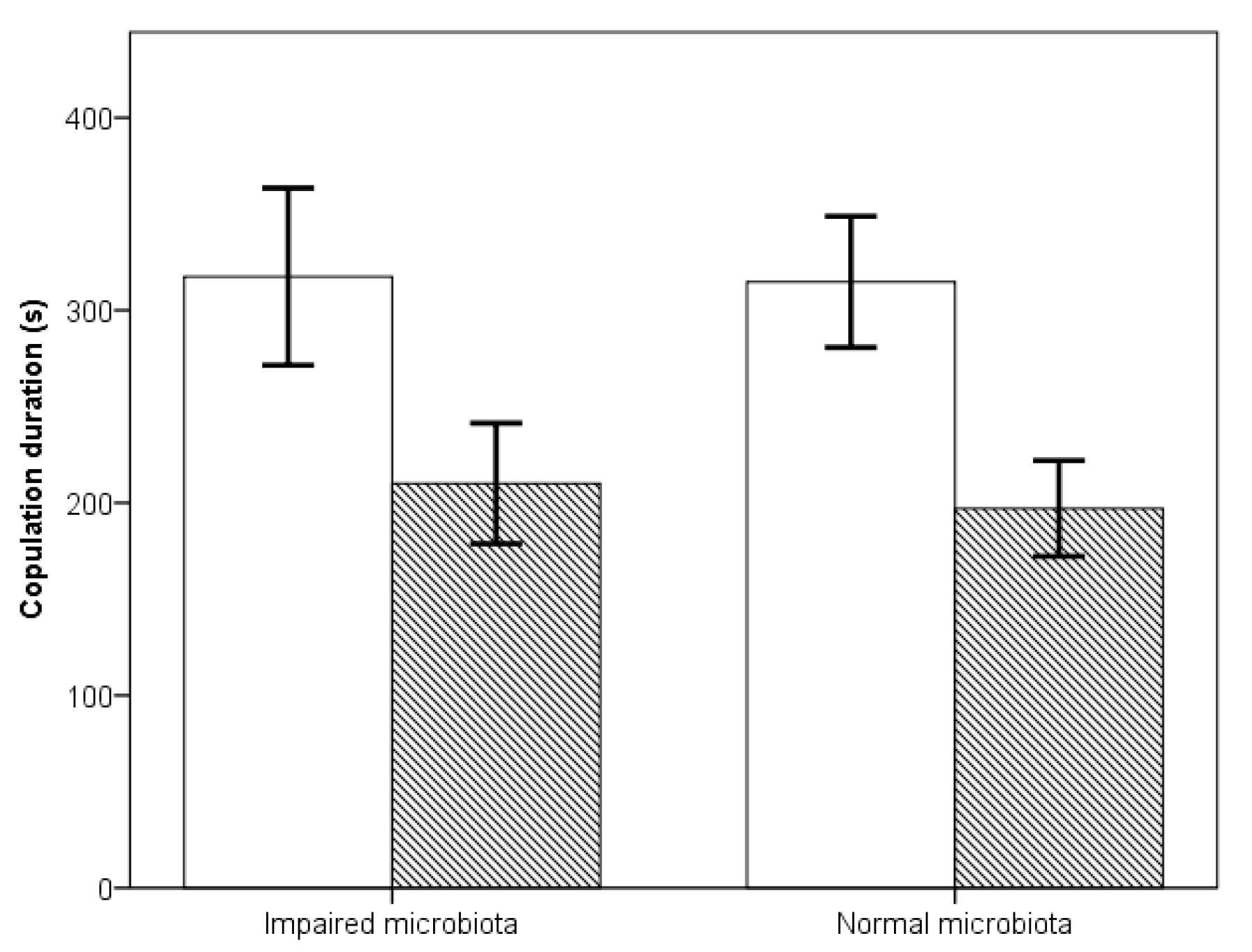
3.1. Preference in No Choice Mating Trials
3.2. Preference in Choice Competitive Mating Trials
3.3. Rapid Iterative Negative Geotaxis (RING) Test of Male Activity
3.4. Gut Microbiota
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Trivers, R. Parental investment and sexual selection. In Sexual selection and the descent of man 1871-1971; Campbell, B., Ed.; Aldine Press: Chicago, IL, USA, 1972; pp. 139–179. [Google Scholar]
- Byers, B.A.; Waits, L. Good genes sexual selection in nature. Proc. Natl. Acad. Sci. USA 2006, 103, 1.6.3.4.3–1.6.3.4.5. [Google Scholar] [CrossRef]
- Suzaki, Y.; Katsuki, M.; Miyatake, T.; Okada, Y. Male courtship behavior and weapon trait as indicators of indirect benefit in the bean bug, Riptortus pedestris. PLoS ONE 2013, 8, e83278. [Google Scholar] [CrossRef]
- Jones, T.M.; Balmford, A.; Quinell, R.J. Adaptive female choice for middle-aged mates in a lekking sandfly. Proc. R. Soc. Lond. B 2000, 267, 681–686. [Google Scholar] [CrossRef]
- López, P.; Aragón, P.; Martin, J. Responses of female lizards, Lacerta monticola, to males’ chemical cues reflect their mating preference for older males. Behav. Ecol. Sociobiol. 2003, 55, 73–79. [Google Scholar] [CrossRef]
- Kleindorfer, S. Nesting success in Darwin’s small tree finch, Camarhynchus parvulus: Evidence of female preference for older males and more concealed nests. Anim. Behav. 2007, 74, 795–804. [Google Scholar] [CrossRef]
- Manning, J.T. Choosy females and correlates of male age. J. Theor. Biol. 1985, 116, 349–354. [Google Scholar] [CrossRef]
- Brooks, R.; Kemp, D.J. Can older male deliver the good genes? Trends Ecol. Evol. 2001, 16, 308–313. [Google Scholar] [CrossRef]
- Proulx, S.R.; Day, T.; Rowe, L. Older males signal more reliably. Proc. R. Soc. Lond. B 2002, 269, 2291–2299. [Google Scholar] [CrossRef]
- Bonduriansky, R.; Brassil, C.E. Rapid and costly ageing in wild male flies. Nature 2002, 420, 377. [Google Scholar] [CrossRef]
- Price, D.K.; Hansen, T.F. How does offspring quality change with age in male Drosophila melanogaster? Behav. Genet. 1998, 28, 395–402. [Google Scholar] [CrossRef]
- Hansen, T.F.; Price, D.K. Good genes and old age: Do old mates provide superior genes? J. Evol. Biol. 1995, 8, 759–778. [Google Scholar] [CrossRef]
- Moulin, B.; Rybak, F.; Aublin, T.; Jallon, J.M. Compared ontogenesis of courtship song components of males from the sibling species, D. melanogaster and D. simulans. Behav. Genet. 2001, 31, 299–308. [Google Scholar] [CrossRef]
- Avent, T.D.; Price, T.A.R.; Wedell, N. Age-based female preference in the fruit fly Drosophila pseudoobscura. Anim. Behav. 2008, 75, 1413–1421. [Google Scholar] [CrossRef]
- Somashekar, K.; Krishna, M.S. Evidence of female preference for older males in Drosophila bipectinata. Zool. Stud. 2011, 50, 1–15. [Google Scholar]
- Wedell, N.; Ritchie, M.G. Male age, mating status and nuptial gift quality in a bush cricket. Anim. Behav. 2004, 67, 1059–1065. [Google Scholar] [CrossRef]
- Shelly, T.E.; Edu, J.; Pahio, E. Female medflies mate selectively with young males but gain no apparent fitness benefits. J. Ins. Behav. 2011, 24, 55–66. [Google Scholar] [CrossRef]
- Papanastasiou, S.A.; Diamantidis, A.D.; Nakas, C.T.; Carey, J.R.; Papadopoulos, N.T. Dual reproductive cost of aging in male medflies: Dramatic decrease in mating competitiveness and gradual reduction in mating performance. J. Ins. Physiol. 2011, 57, 1368–1374. [Google Scholar] [CrossRef]
- Liu, X.P.; Xu, J.; He, H.M.; Xue, F.S. Male age affects female mate preference and reproductive performance in the cabbage beetle, Colaphellus bowringi. J. Ins. Behav. 2011, 24, 83–93. [Google Scholar] [CrossRef]
- Verburgt, L.; Ferreira, M.; Ferguson, J.W.H. Male field cricket song reflects age, allowing females to prefer younger males. Anim. Behav. 2011, 81, 19–29. [Google Scholar] [CrossRef]
- Brucker, R.M.; Bordenstein, S.R. Speciation by Symbiosis. Trends Ecol. Evol. 2012, 27, 443–451. [Google Scholar] [CrossRef]
- Markov, A.V.; Lazebny, O.E.; Goryacheva, I.I.; Antipin, M.I.; Kulikov, A.M. Symbiotic bacteria affect mating choice in Drosophila melanogaster. Anim. Behav. 2009, 77, 1011–1017. [Google Scholar] [CrossRef]
- Sharon, G.; Segal, D.; Ringo, J.M.; Hefetz, A.; Zilber-Rosenberg, I.; Rosenberg, E. Commensal bacteria play a role in mating preference of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2010, 107, 20051–20056. [Google Scholar] [CrossRef]
- Walsh, B.S.; Heys, C.; Lewis, Z. Gut microbiota influences female choice and fecundity in the nuptial gift-giving species, Drosophila subobscura (Diptera: Drosophilidae). Eur. J. Entomol. 2017, 114, 439–445. [Google Scholar] [CrossRef]
- Lizé, A.; McKay, R.; Lewis, Z. Kin recognition in Drosophila: The importance of ecology and gut bacteria. ISME J. 2014, 8, 469–477. [Google Scholar] [CrossRef]
- Najarro, M.A.; Sumethasorn, M.; Lamoureux, A.; Turner, T.L. Choosing mates based on the diet of your ancestors: Replication of non-genetic assortative mating in Drosophila melanogaster. Peer J. 2015, 3, e1173. [Google Scholar] [CrossRef]
- Chandler, J.A.; Lang, J.M.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial communities of diverse Drosophila species: Ecological context of a host-microbe model system. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef]
- Wong, C.A.N.; Chaston, J.M.; Douglas, A.E. The inconstant gut microbiota of Drosophila species revealed by 16S rRNA gene analysis. ISME J. 2013, 7, 1922–1932. [Google Scholar] [CrossRef]
- Wong, C.A.N.; Ng, P.; Douglas, A.E. Low-diversity bacterial community in the gut of the fruitfly Drosophila melanogaster. Environ. Microbial. 2011, 2511, 1889–1900. [Google Scholar] [CrossRef]
- Brummel, T.; Ching, A.; Seroude, L.; Simon, A.F.; Benzer, S. Drosophila lifespan enhancement by exogenous bacteria. Proc. Natl. Acad. Sci. USA 2004, 101, 12974–12979. [Google Scholar] [CrossRef]
- Clark, R.I.; Salazar, A.; Yamada, R.; Fitz-Gibbon, S.; Morselli, M.; Alcaraz, J.; Rana, A.; Rera, M.; Pellegrini, M.; Ja, W.W.; et al. Distinct shifts in microbiota composition during Drosophila aging impair intestinal function and drive mortality. Cell Rep. 2015, 12, 1656–1667. [Google Scholar] [CrossRef]
- Galenza, A.; Hutchinson, J.; Campbell, S.D.; Hazes, B.; Foley, E. Glucose modulates Drosophila longevity and immunity independent of the microbiota. Biol. Open. 2016, 5, 165–173. [Google Scholar] [CrossRef]
- Lee, H.-Y.; Lee, S.-H.; Lee, J.-H.; Lee, W.-J.; Min, K.-Y. The role of commensal microbes in the lifespan on Drosophila melanogaster. Aging 2019, 11, 4611–4640. [Google Scholar] [CrossRef]
- Buchon, N.; Broderick, N.A.; Chakrabarti, S.; Lemaitre, B. Invasive and indigenous microbiota impact intestinal stem cell activity through multiple pathways in Drosophila. Genes Dev. 2009, 23, 2333–2344. [Google Scholar] [CrossRef]
- Ryu, J.H.; Kim, S.H.; Lee, H.Y.; Bai, J.Y.; Nam, Y.D.; Bae, J.W.; Lee, D.G.; Shin, S.C.; Ha, E.M.; Lee, W.J. Innate immune homeostasis by the homeobox gene caudal and com- mensal-gut mutualism in Drosophila. Science 2008, 319, 777–782. [Google Scholar] [CrossRef]
- Ren, C.; Webster, P.; Finkel, S.E.; Tower, J. Increased internal and external bacterial load during Drosophila aging without life-span trade-off. Cell Metab. 2007, 6, 144–152. [Google Scholar] [CrossRef]
- Dhole, S.; Pfennig, K.S. Age-dependent male mating investment in Drosophila pseudoobscura. PLoS ONE 2014, 9, e88700. [Google Scholar] [CrossRef]
- Lin, X.-L.; Kang, Z.-W.; Pan, Q.-J.; Liu, T.-X. Evaluation of five antibiotics on larval gut bacterial diversity of Plutella xylostella (Lepidoptera: Plutellidae). Insect Sci. 2015, 22, 619–628. [Google Scholar] [CrossRef]
- Graf, E.; Benz, G. Toxicity of streptomycin and terramycin, and influence on growth and developmental time of Drosophila melanogaster. Experientia 1970, 26, 1339. [Google Scholar] [CrossRef]
- Heys, C.; Lizé, A.; Blow, F.; White, L.; Darby, A.; Lewis, Z. The effect of gut microbiota elimination in Drosophila melanogaster: A how-to guide for host-microbiota studies. Ecol. Evol. 2018, 8, 4150–4161. [Google Scholar] [CrossRef]
- Broderick, N.A.; Lemaitre, B. Gut associated microbes of Drosophila melanogaster. Gut Microb. 2012, 3, 307–321. [Google Scholar] [CrossRef]
- Lizé, A.; Doff, R.J.; Smaller, E.A.; Lewis, Z.; Hurst, G.D.D. Perception of male-male competition influences Drosophila copulation behaviour even in species where females rarely remate. Biol. Lett. 2012, 8, 35–38. [Google Scholar] [CrossRef]
- Lizé, A.; Price, T.A.R.; Heys, C.; Lewis, Z.; Hurst, G.D.D. Extreme cost of rivalry in a monandrous species: Male-male interactions result in failure to acquire mates and reduced longevity. Proc. R. Soc. Lond. B 2014, 281, 20140631. [Google Scholar] [CrossRef]
- Dobzhansky, T.; Epling, C. Contributions to the Genetics, Taxonomy and Ecology of Drosophila Pseudoobscura and its Relatives; Carnegie Institution of Washington: Washington, DC, USA, 1944. [Google Scholar]
- Speith, H.T. Courtship behavioiur in Drosophila. Annu. Rev. Entomol. 1974, 19, 385–405. [Google Scholar] [CrossRef]
- Economos, A.C.; Miquel, J.; Binnard, R.; Kessler, S. Quantitative analysis of mating behaviour in ageing male Drosophila melanogaster. Mech. Ageing Dev. 1979, 10, 233–240. [Google Scholar] [CrossRef]
- Noor, M.A.F.; Coyne, J.A. Genetics of a difference in cuticular hydrocarbons between Drosophila pseudoobscura and D. persimilis. Genet. Res. 1996, 68, 117–123. [Google Scholar] [CrossRef]
- Lefranc, A.; Bundgaard, J. The influence of male and female body size on copulation duration and fecundity in Drosophila melanogaster. Hereditas 2000, 132, 243–247. [Google Scholar] [CrossRef]
- Verspoor, R.; Cuss, M.; Price, T.A.R. Age based mate choice in the monandrous fruit fly Drosophila subobscura. Anim. Behav. 2015, 102, 199–207. [Google Scholar] [CrossRef]
- Prathibha, M.; Krishna, M.S.; Jayaramu, S.C. Male age influence on male reproductive success in Drosophila ananassae (Diptera: Drosophilidae). Ital. J. Zool. 2011, 78, 168–173. [Google Scholar] [CrossRef][Green Version]
- Ehrman, L. Mating success and genotype frequency in Drosophila. Anim. Behav. 1966, 14, 332–339. [Google Scholar] [CrossRef]
- Ehrman, L.; Petit, C. Genotype frequency and mating success in the willistoni species group of Drosophila. Evolution 1968, 22, 649–658. [Google Scholar] [CrossRef]
- Byrant, E.H.; Kence, A.; Kimball, K.T. A rare-male advantage in the housefly induced by wing clipping and some general considerations for Drosophila. Genetics 1980, 96, 975–993. [Google Scholar]
- Knoppien, P. The rare male mating advantage: An artefact caused by marking procedures? Am. Nat. 1984, 123, 862–866. [Google Scholar] [CrossRef]
- Dodd DMB. Reproductive isolation as a consequence of adaptive divergence in Drosophila pseudoobscura. Evolution 1989, 43, 1308–1311. [Google Scholar] [CrossRef]
- Nichols, C.D.; Becnel, J.; Pandey, U.B. Methods to assay Drosophila behaviour. J. Vis. Exp. 2012, 61, 3795. [Google Scholar]
- Brennan, P.L.R.; Richard, O.P. The limits of sexual conflict in the narrow sense: New insights from waterfowl biology. Phil. Trans. R. Soc. B 2012, 367, 2324–2338. [Google Scholar] [CrossRef]
- Gargano, J.W.; Martin, I.; Bhandari, P.; Grotewiel, M.S. Rapid Iterative Negative Geotaxis (RING): A new method for assessing age related locomotor decline in Drosophila. Exp. Geront. 2005, 40, 386–395. [Google Scholar] [CrossRef]
- Schretter, C.E.; Vielmetter, J.; Bartos, I.; Marka, Z.; Argade, S.; Mazmanian, S.K. A gut microbial factor modulates locomotor behaviour in Drosophila. Nature 2018, 563, 402–406. [Google Scholar] [CrossRef]
- Price, T.A.R.; Bretman, A.J.; Snook, R.R.; Hurst, G.D.D.; Wedell, N. Sex ratio distorter reduces sperm competitive ability in an insect. Evolution 2008, 62, 1644–1652. [Google Scholar] [CrossRef]
- Ferveur, J.F. Cuticular hydrocarbons: Their evolution and roles in Drosophila pheromonal communication. Behav. Genet. 2005, 35, 279–295. [Google Scholar] [CrossRef]
- Blomquist, G.J. Biosynthesis of cuticular hydrocarbons. In Insect Hydrocarbons Biology, Biochemistry, and Chemical Ecology, Blomquist G.J. and Bagnères A-G., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 35–52. [Google Scholar]
- Scott, D.; Shields, A.; Straker, M.; Dalrymple, H.; Dhillion, P.K.; Harbinder, S. Variation in the male pheromones and mating success of wild caught Drosophila melanogaster. PLoS ONE 2011, 6, e23645. [Google Scholar] [CrossRef]
- Nunes, T.M.; Turatti, L.C.C.; Mateus, S.; Nascimento, F.S.; Lopes, N.P.; Zucchi, R. Cuticular hydrocarbons in the stingless bee Schwarziana quadripunctata (Hymenoptera, Apidae, Meliponini): Differences between colonies, castes and age. Genet. Mol. Res. 2009, 8, 589–595. [Google Scholar] [CrossRef]
- Hugo, L.E.; Kay, B.H.; Eaglesham, G.K.; Holling, N.; Ryan, P.A. Investigation of cuticular hydrocarbons for determining the age and survivorship of Australasian mosquitoes. Am. J. Trop. Med. Hyg. 2006, 74, 464–474. [Google Scholar] [CrossRef]
- Kuo, T.H.; Yew, J.Y.; Fedina, T.Y.; Dreiesewerd, K.; Dierick, H.A.; Pletcher, S.D. Ageing modulates cuticular hydrocarbons and sexual attractiveness in Drosophila melanogaster. J. Exp. Biol. 2012, 215, 814–821. [Google Scholar] [CrossRef]
- Heys, C.; Lizé, A.; Colinet, H.; Price, T.A.R.P.; Prescott, M.; Ingleby, F.; Lewis, Z. Evidence that the microbiota counteracts male outbreeding strategy by inhibiting sexual signalling in females. Front. Ecol. Evol. 2018, 6, 29. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Diet | Age | Replicate | Number of colonies |
|---|---|---|---|
| Normal | Young | 1 | 1 |
| Normal | Young | 2 | 3 |
| Normal | Young | 3 | 2 |
| Antibiotic | Young | 1 | 0 |
| Antibiotic | Young | 2 | 0 |
| Antibiotic | Young | 3 | 0 |
| Normal | Old | 1 | 6396 |
| Normal | Old | 2 | 8528 |
| Normal | Old | 3 | 12,428 |
| Antibiotic | Old | 1 | 0 |
| Antibiotic | Old | 2 | 0 |
| Antibiotic | Old | 3 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heys, C.; Lizé, A.; Lewis, Z.; Price, T.A.R. Drosophila Sexual Attractiveness in Older Males Is Mediated by Their Microbiota. Microorganisms 2020, 8, 168. https://doi.org/10.3390/microorganisms8020168
Heys C, Lizé A, Lewis Z, Price TAR. Drosophila Sexual Attractiveness in Older Males Is Mediated by Their Microbiota. Microorganisms. 2020; 8(2):168. https://doi.org/10.3390/microorganisms8020168
Chicago/Turabian StyleHeys, Chloe, Anne Lizé, Zenobia Lewis, and Tom A. R. Price. 2020. "Drosophila Sexual Attractiveness in Older Males Is Mediated by Their Microbiota" Microorganisms 8, no. 2: 168. https://doi.org/10.3390/microorganisms8020168
APA StyleHeys, C., Lizé, A., Lewis, Z., & Price, T. A. R. (2020). Drosophila Sexual Attractiveness in Older Males Is Mediated by Their Microbiota. Microorganisms, 8(2), 168. https://doi.org/10.3390/microorganisms8020168