Uncoupling Foam Fractionation and Foam Adsorption for Enhanced Biosurfactant Synthesis and Recovery


Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Culture Conditions
2.3. Fermentation Setup and Procedure
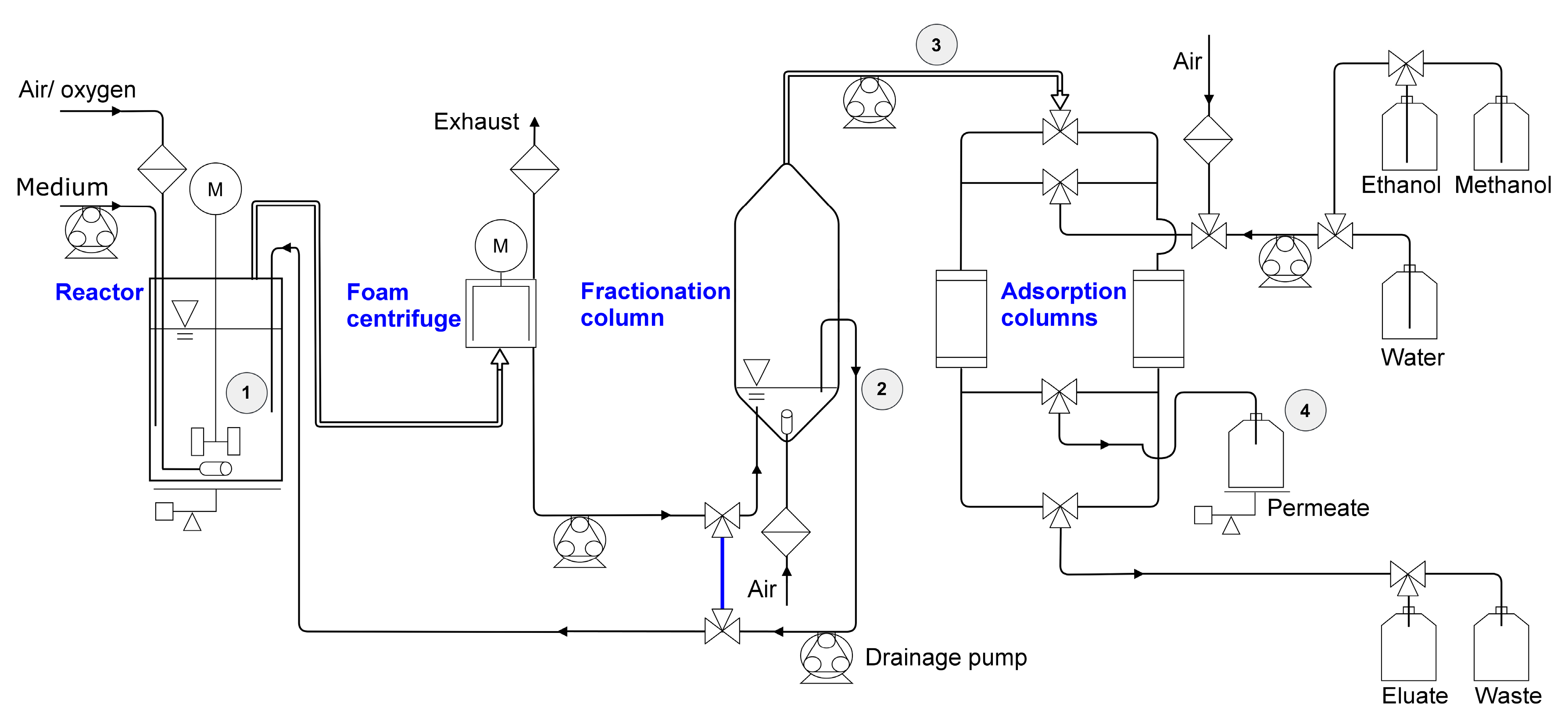
2.3.1. Setup and Procedure for 2 L Scale
2.3.2. Setup and Procedure for 9 L Scale
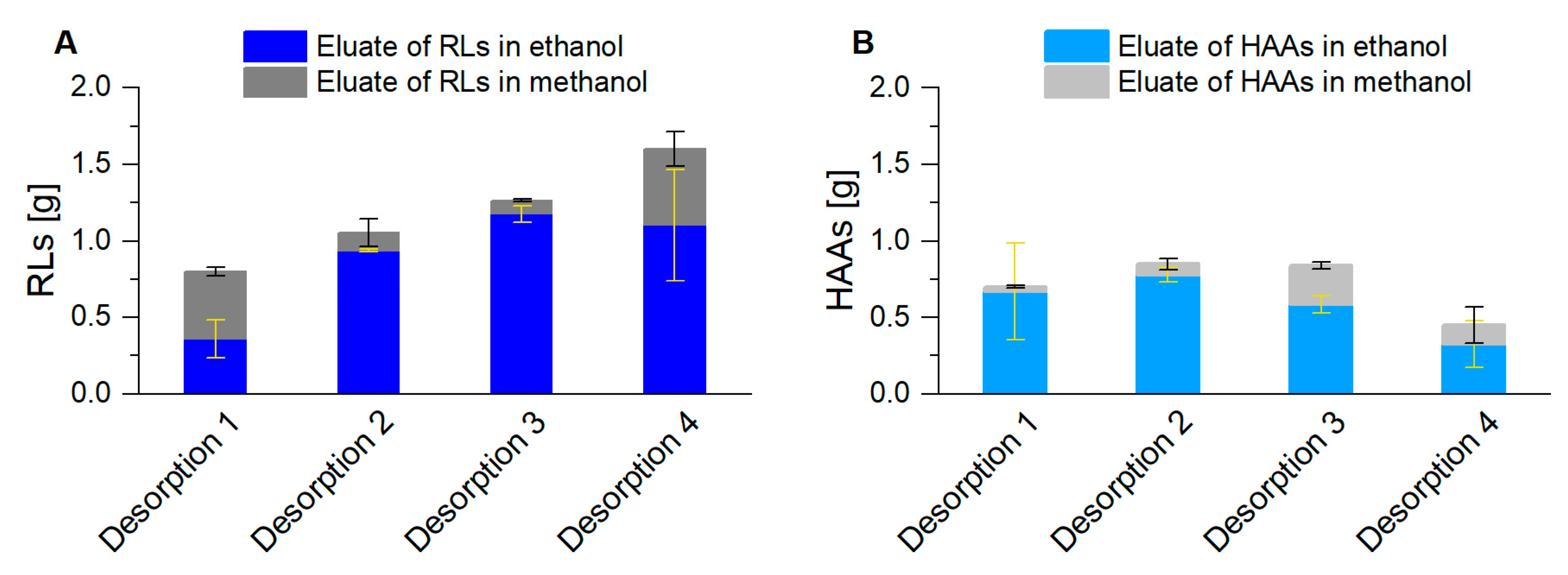
2.3.3. Determination of HAA Adsorption Capacity
2.4. Sampling and Analytics
2.5. Data Analysis
3. Results
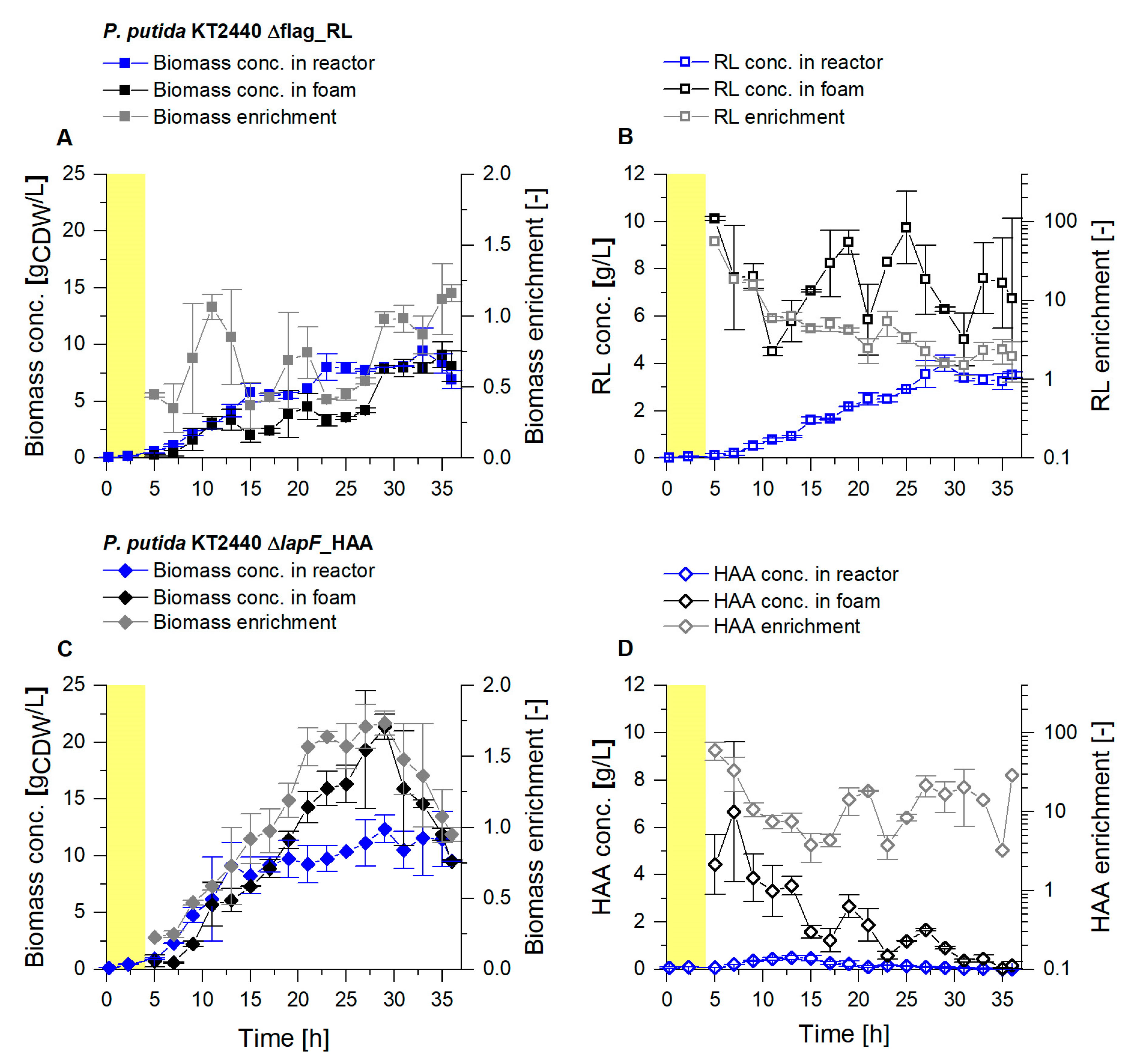
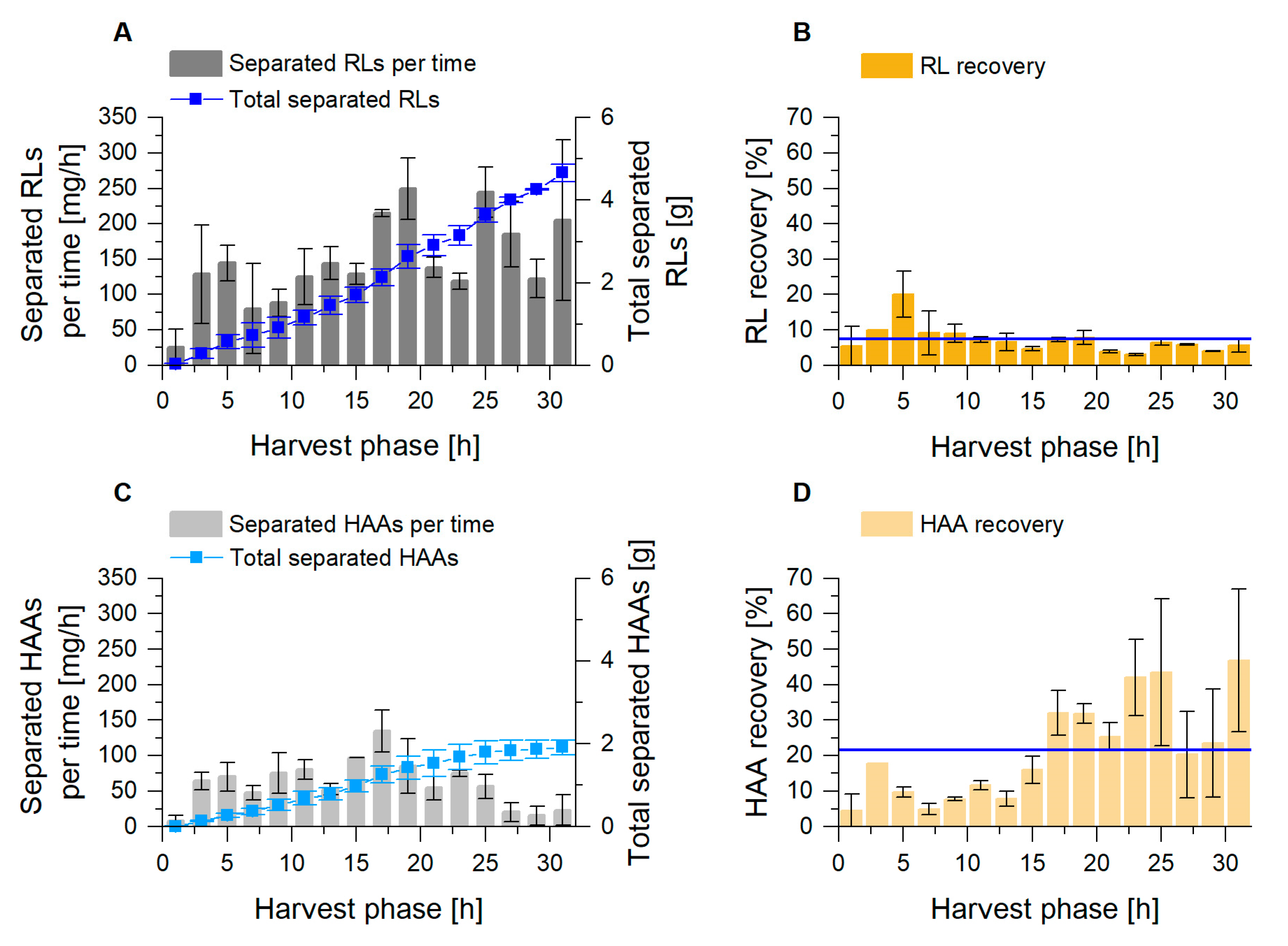
3.1. Enhanced Fractionation Performance by Uncoupling Its Operation from Fermentation
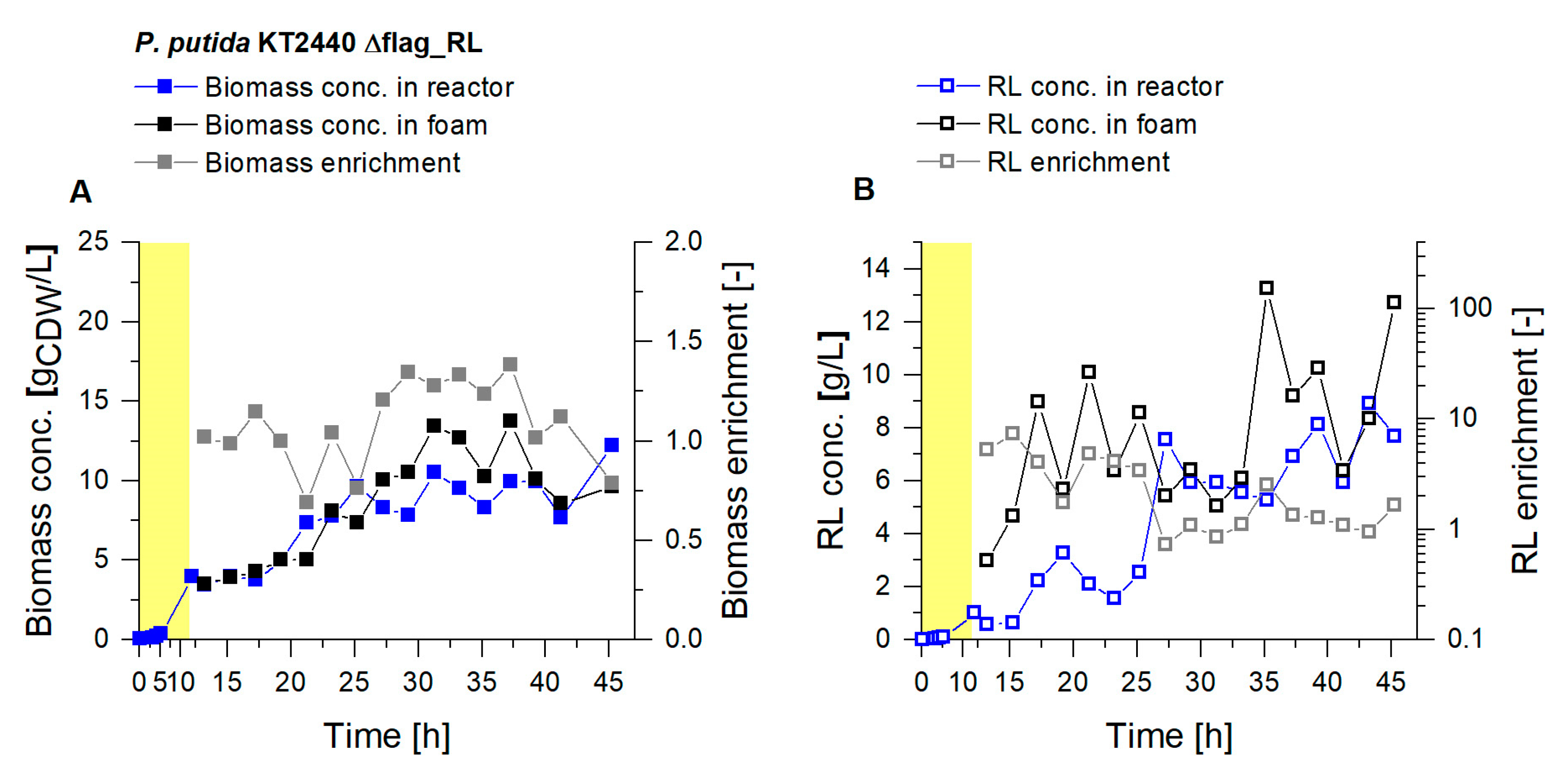
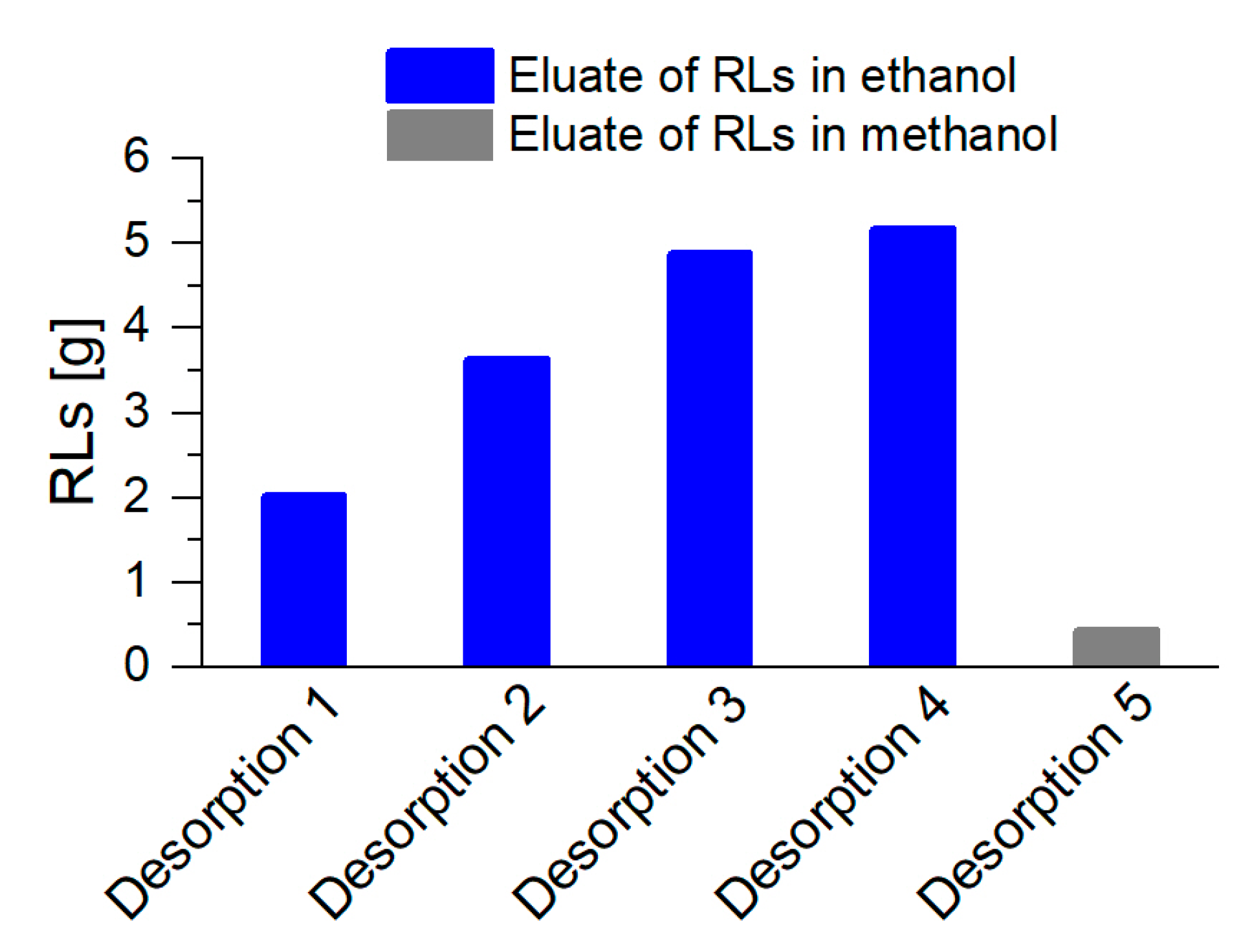
3.2. Coupled Foam Adsorption Allows In Situ Product Removal
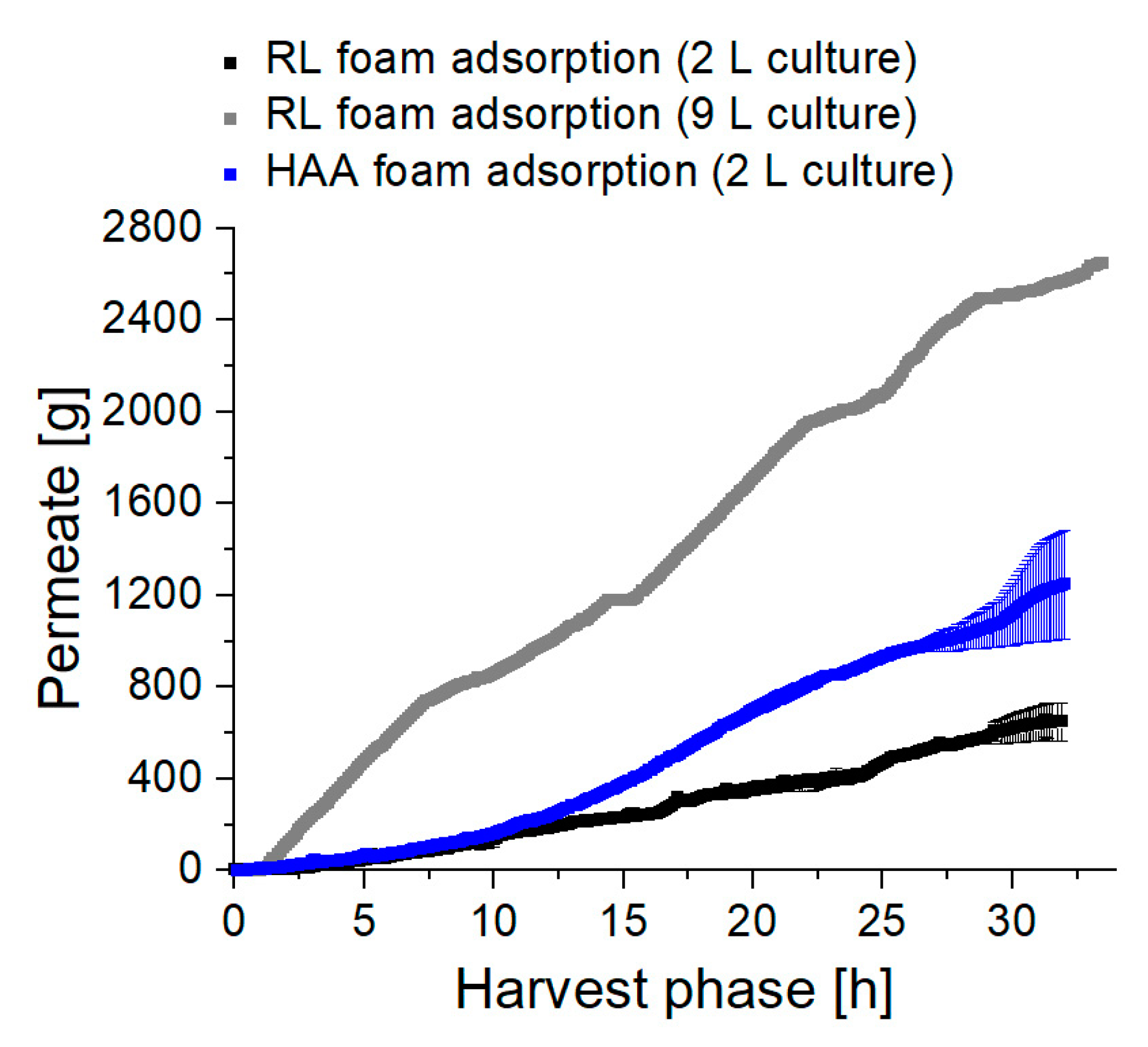
3.3. Up-Scaling of Fermentation Volume Showcases Robustness of the Presented Process
3.3.1. Higher Foam Quantities Decrease the Efficiency of Foam Fractionation
3.3.2. Fractionated Foam Causes Steady Permeate Flow through the Adsorption Column
4. Discussion
4.1. Combining Previous Knowledge with Recent Findings Enabled the Design of a Highly Efficient System for Biosurfactant Production
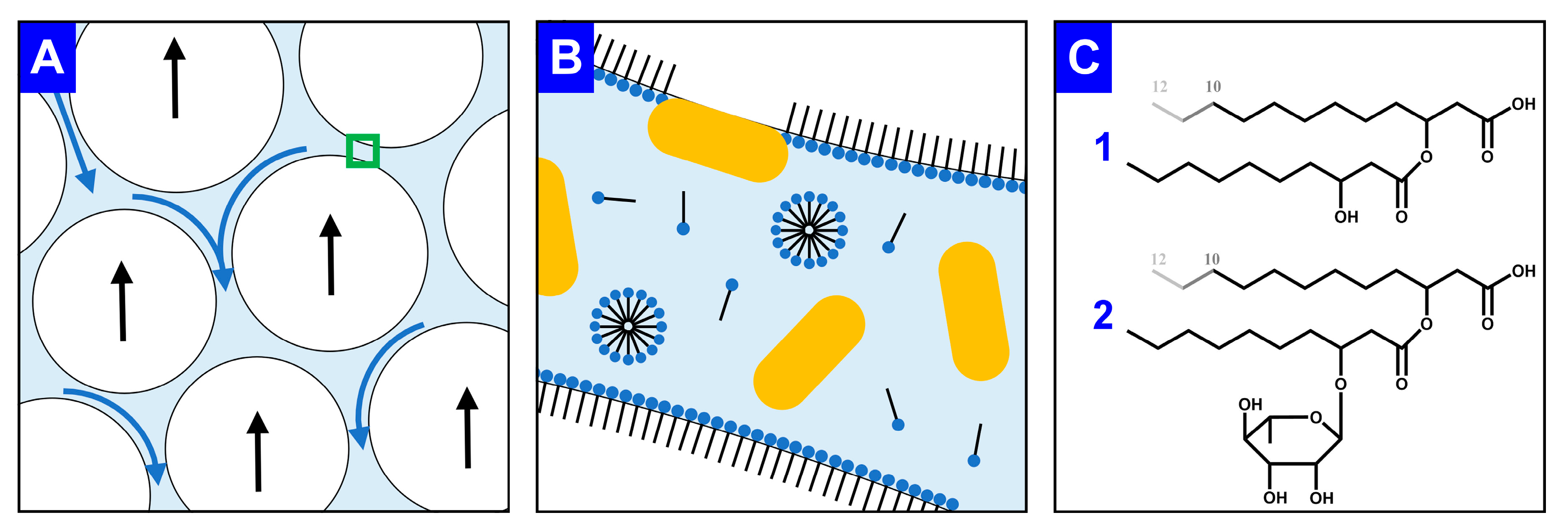
4.2. Fermentation Products Impact Foam Formation and Stabilization
4.3. Further Technical Enhancements in Foam Fractionation Are Foreseeable
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Henkel, M.; Geissler, M.; Weggenmann, F.; Hausmann, R. Production of microbial biosurfactants: Status quo of rhamnolipid and surfactin towards large-scale production. Biotechnol. J. 2017, 12. [Google Scholar] [CrossRef]
- Mulligan, C.N. Environmental applications for biosurfactants. Environ. Pollut. 2005, 133, 183–198. [Google Scholar] [CrossRef]
- Fenibo, E.O.; Ijoma, G.N.; Selvarajan, R.; Chikere, C.B. Microbial Surfactants: The next generation multifunctional biomolecules for applications in the petroleum industry and Its associated environmental remediation. Microorganisms 2019, 7, 581. [Google Scholar] [CrossRef]
- Kosaric, N.; Vardar-Sukan, F. Biosurfactants; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2015. [Google Scholar]
- Ines, M.; Dhouha, G. Glycolipid biosurfactants: Potential related biomedical and biotechnological applications. Carbohydr. Res. 2015, 416, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Paulino, B.N.; Pessoa, M.G.; Mano, M.C.; Molina, G.; Neri-Numa, I.A.; Pastore, G.M. Current status in biotechnological production and applications of glycolipid biosurfactants. Appl. Microbiol. Biotechnol. 2016, 100, 10265–10293. [Google Scholar] [CrossRef] [PubMed]
- Roelants, S.; Solaiman, D.K.Y.; Ashby, R.D.; Lodens, S.; Van Renterghem, L.; Soetaert, W. Production and applications of sophorolipids. In Biobased Surfactants; Elsevier Inc.: New York, NY, USA, 2019; pp. 65–119. [Google Scholar] [CrossRef]
- Evonik Commercializes Biosurfactants; Evonik Industries AG: Essen, Germany, June 2016; pp. 1–3. Available online: https://corporate.evonik.com/en/evonik-commercializes-biosurfactants-101537.html (accessed on 1 November 2020).
- Banat, I.M.; Franzetti, A.; Gandolfi, I.; Bestetti, G.; Martinotti, M.G.; Fracchia, L.; Smyth, T.J.; Marchant, R. Microbial biosurfactants production, applications and future potential. Appl. Microbiol. Biotechnol. 2010, 87, 427–444. [Google Scholar] [CrossRef]
- Hubert, J.; Plé, K.; Hamzaoui, M.; Nuissier, G.; Hadef, I.; Reynaud, R.; Guilleret, A.; Renault, J.-H. New perspectives for microbial glycolipid fractionation and purification processes. C. R. Chim. 2012, 15, 18–28. [Google Scholar] [CrossRef]
- Müller, M.M.; Kugler, J.H.; Henkel, M.; Gerlitzki, M.; Hormann, B.; Pohnlein, M.; Syldatk, C.; Hausmann, R. Rhamnolipids- next generation surfactants? J. Biotechnol. 2012, 162, 366–380. [Google Scholar] [CrossRef]
- Irorere, V.U.; Tripathi, L.; Marchant, R.; McClean, S.; Banat, I.M. Microbial rhamnolipid production: A critical re-evaluation of published data and suggested future publication criteria. Appl. Microbiol. Biotechnol. 2017, 101, 3941–3951. [Google Scholar] [CrossRef] [PubMed]
- Henkel, M.; Müller, M.M.; Kügler, J.H.; Lovaglio, R.B.; Contiero, J.; Syldatk, C.; Hausmann, R. Rhamnolipids as biosurfactants from renewable resources: Concepts for next-generation rhamnolipid production. Process Biochem. 2012, 47, 1207–1219. [Google Scholar] [CrossRef]
- Müller, M.M.; Hörmann, B.; Syldatk, C.; Hausmann, R. Pseudomonas aeruginosa PAO1 as a model for rhamnolipid production in bioreactor systems. Appl. Microbiol. Biotechnol. 2010, 87, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, F.G.; Johnson, M.J. A glyco-lipide produced by Pseudomonas aeruginosa. J. Am. Chem. Soc. 1949, 71, 4124–4126. [Google Scholar] [CrossRef]
- Déziel, E.; Lépine, F.; Dennie, D.; Boismenu, D.; Mamer, O.A.; Villemur, R. Liquid chromatography/mass spectrometry analysis of mixtures of rhamnolipids produced by Pseudomonas aeruginosa strain 57RP grown on mannitol or naphthalene. BBA-Mol. Cell. Biol. L 1999, 1440, 244–252. [Google Scholar] [CrossRef]
- Ochsner, U.A.; Fiechter, A.; Reiser, J. Isolation, characterization, and expression in Escherichia coli of the Pseudomonas aeruginosa rhlAB genes encoding a rhamnosyltransferase involved in rhamnolipid biosurfactant synthesis. J. Biol. Chem. 1994, 269, 19787–19795. [Google Scholar]
- Sekhon Randhawa, K.K.; Rahman, P.K. Rhamnolipid biosurfactants-past, present, and future scenario of global market. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef]
- Soberon-Chavez, G.; Gonzalez-Valdez, A.; Soto-Aceves, M.P.; Cocotl-Yanez, M. Rhamnolipids produced by Pseudomonas: From molecular genetics to the market. Microb. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Bahia, F.M.; de Almeida, G.C.; de Andrade, L.P.; Campos, C.G.; Queiroz, L.R.; da Silva, R.L.V.; Abdelnur, P.V.; Correa, J.R.; Bettiga, M.; Parachin, N.S. Rhamnolipids production from sucrose by engineered Saccharomyces cerevisiae. Sci. Rep. 2018, 8, 2905. [Google Scholar] [CrossRef]
- Cabrera-Valladares, N.; Richardson, A.P.; Olvera, C.; Trevino, L.G.; Deziel, E.; Lepine, F.; Soberon-Chavez, G. Monorhamnolipids and 3-(3-hydroxyalkanoyloxy)alkanoic acids (HAAs) production using Escherichia coli as a heterologous host. Appl. Microbiol. Biotechnol. 2006, 73, 187–194. [Google Scholar] [CrossRef]
- Wittgens, A.; Tiso, T.; Arndt, T.T.; Wenk, P.; Hemmerich, J.; Müller, C.; Wichmann, R.; Küpper, B.; Zwick, M.; Wilhelm, S.; et al. Growth independent rhamnolipid production from glucose using the non-pathogenic Pseudomonas putida KT2440. Microb. Cell Fact. 2011, 10. [Google Scholar] [CrossRef]
- Tiso, T.; Ihling, N.; Kubicki, S.; Biselli, A.; Schonhoff, A.; Bator, I.; Thies, S.; Karmainski, T.; Kruth, S.; Willenbrink, A.-L.; et al. Integration of genetic and process engineering for optimized rhamnolipid production using Pseudomonas putida. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef]
- Blesken, C.C.; Bator, I.; Eberlein, C.; Heipieper, H.J.; Tiso, T.; Blank, L.M. Genetic cell-surface modification for optimized foam fractionation. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Blank, L.M.; Tiso, T.; Germer, A. Extracellular Production of Designer Hydroxyalkanoyloxy Alcanoic Acids with Recombinant Bacteria. World Patent WO2017006252A1, 6 July 2016. [Google Scholar]
- Clean and Green; ELEMENTS Journal; Evonik Industries AG: Essen, Germany, 2020; pp. 36–41.
- Long, X.W.; Sha, R.Y.; Meng, Q.; Zhang, G.L. Mechanism study on the severe foaming of rhamnolipid in fermentation. J. Surfactants Deterg. 2016, 19, 833–840. [Google Scholar] [CrossRef]
- Sodagari, M.; Invally, K.; Ju, L.K. Maximize rhamnolipid production with low foaming and high yield. Enzym. Microb. Technol. 2018, 110, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Sha, R.; Meng, Q.; Jiang, L. The addition of ethanol as defoamer in fermentation of rhamnolipids. J. Chem. Technol. Biotechnol. 2012, 87, 368–373. [Google Scholar] [CrossRef]
- Sarachat, T.; Pornsunthorntawee, O.; Chavadej, S.; Rujiravanit, R. Purification and concentration of a rhamnolipid biosurfactant produced by Pseudomonas aeruginosa SP4 using foam fractionation. Bioresour. Technol. 2010, 101, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, P.; Li, X. Foam Fractionation-Principles and Process Design, 1st ed.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2014. [Google Scholar]
- Etoc, A.; Delvigne, F.; Lecomte, J.P.; Thonart, P. Foam Control in Fermentation Bioprocess. Appl. Biochem. Biotechnol. 2006, 129–132, 392–404. [Google Scholar] [CrossRef]
- Vardar-Sukan, F. Foaming: Consequences, prevention and destruction. Biotechnol. Adv. 1998, 16, 913–948. [Google Scholar] [CrossRef]
- Sodagari, M.; Ju, L.-K. Cells were a more important foaming factor than free rhamnolipids in fermentation of Pseudomonas aeruginosa E03-40 for high rhamnolipid production. J. Surfactants Deterg. 2013, 17, 573–582. [Google Scholar] [CrossRef]
- Chayabutra, C.; Ju, L.K. Polyhydroxyalkanoic acids and rhamnolipids are synthesized sequentially in hexadecane fermentation by Pseudomonas aeruginosa ATCC 10145. Biotechnol. Prog. 2001, 17, 419–423. [Google Scholar] [CrossRef]
- Beuker, J.; Barth, T.; Steier, A.; Wittgens, A.; Rosenau, F.; Henkel, M.; Hausmann, R. High titer heterologous rhamnolipid production. AMB Express 2016, 6, 124. [Google Scholar] [CrossRef]
- Zhu, L.; Yang, X.; Xue, C.; Chen, Y.; Qu, L.; Lu, W. Enhanced rhamnolipids production by Pseudomonas aeruginosa based on a pH stage-controlled fed-batch fermentation process. Bioresour. Technol. 2012, 117, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Özdemir, G.; Peker, S.; Helvaci, S.S. Effect of pH on the surface and interfacial behavior of rhamnolipids R1 and R2. Colloids Surf. A Physicochem. Eng. Asp. 2004, 234, 135–143. [Google Scholar] [CrossRef]
- Ramos, J.L.; Sol Cuenca, M.; Molina-Santiago, C.; Segura, A.; Duque, E.; Gomez-Garcia, M.R.; Udaondo, Z.; Roca, A. Mechanisms of solvent resistance mediated by interplay of cellular factors in Pseudomonas putida. FEMS Microbiol. Rev. 2015, 39, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, U.A.; Hembach, T.; Fiechter, A. Production of rhamnolipid biosurfactants. Adv. Biochem. Eng. Biotechnol. 1996, 53, 89–118. [Google Scholar] [CrossRef]
- Najmi, Z.; Ebrahimipour, G.; Franzetti, A.; Banat, I.M. In situ downstream strategies for cost-effective bio/surfactant recovery. Biotechnol. Appl. Biochem. 2018, 65, 523–532. [Google Scholar] [CrossRef]
- Demling, P.; von Campenhausen, M.; Grütering, C.; Tiso, T.; Jupke, A.; Blank, L.M. Selection of a recyclable in situ liquid–liquid extraction solvent for foam-free synthesis of rhamnolipids in a two-phase fermentation. Green Chem. 2020. [Google Scholar] [CrossRef]
- Kronemberger, F.A.; Borges, C.P.; Freire, D.M.G. Fed-Batch Biosurfactant Production in a Bioreactor. Int. Rev. Chem. Eng. 2010, 2, 513–518. [Google Scholar]
- Xu, N.; Liu, S.; Xu, L.; Zhou, J.; Xin, F.; Zhang, W.; Qian, X.; Li, M.; Dong, W.; Jiang, M. Enhanced rhamnolipids production using a novel bioreactor system based on integrated foam-control and repeated fed-batch fermentation strategy. Biotechnol. Biofuels 2020, 13, 80. [Google Scholar] [CrossRef]
- Long, X.; Shen, C.; He, N.; Zhang, G.; Meng, Q. Enhanced rhamnolipids production via efficient foam-control using stop valve as a foam breaker. Bioresour. Technol. 2017, 224, 536–543. [Google Scholar] [CrossRef]
- Reiling, H.E.; Thanei-Wyss, U.; Guerra-Santos, L.H.; Hirt, R.; Kappeli, O.; Fiechter, A. Pilot plant production of rhamnolipid biosurfactant by Pseudomonas aeruginosa. Appl. Environ. Microbiol. 1986, 51, 985–989. [Google Scholar] [CrossRef]
- Ankenbauer, A.; Schafer, R.A.; Viegas, S.C.; Pobre, V.; Voss, B.; Arraiano, C.M.; Takors, R. Pseudomonas putida KT2440 is naturally endowed to withstand industrial-scale stress conditions. Microb. Biotechnol. 2020, 13, 1145–1161. [Google Scholar] [CrossRef] [PubMed]
- Nikel, P.I.; Chavarria, M.; Danchin, A.; de Lorenzo, V. From dirt to industrial applications: Pseudomonas putida as a Synthetic Biology chassis for hosting harsh biochemical reactions. Curr. Opin. Chem. Biol. 2016, 34, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Merz, J.; Zorn, H.; Burghoff, B.; Schembecker, G. Purification of a fungal cutinase by adsorptive bubble separation: A statistical approach. Colloids Surf. A 2011, 382, 81–87. [Google Scholar] [CrossRef]
- Lemlich, R. Adsubble processes: Foam fractionation and bubble fractionation. J. Geophys. Res. 1972, 77, 5204–5210. [Google Scholar] [CrossRef]
- de Vries, A.J. Foam stability. Part I. Structure and stability of foams. Recl. Trav. Chim. Pays-Bas 1958, 77, 81–91. [Google Scholar] [CrossRef]
- Tiso, T.; Zauter, R.; Tulke, H.; Leuchtle, B.; Li, W.-J.; Behrens, B.; Wittgens, A.; Rosenau, F.; Hayen, H.; Blank, L.M. Designer rhamnolipids by reduction of congener diversity: Production and characterization. Microb. Cell Fact. 2017, 16, 225. [Google Scholar] [CrossRef]
- Deziel, E.; Lepine, F.; Milot, S.; Villemur, R. rhlA is required for the production of a novel biosurfactant promoting swarming motility in Pseudomonas aeruginosa: 3-(3-hydroxyalkanoyloxy)alkanoic acids (HAAs), the precursors of rhamnolipids. Microbiology 2003, 149, 2005–2013. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Baker, S.C.; Darton, R.C. Batch production of biosurfactant with foam fractionation. J. Chem. Technol. Biotechnol. 2006, 81, 1923–1931. [Google Scholar] [CrossRef]
- Alonso, S.; Martin, P.J. Impact of foaming on surfactin production by Bacillus subtilis: Implications on the development of integrated in situ foam fractionation removal systems. Biochem. Eng. J. 2016, 110, 125–133. [Google Scholar] [CrossRef]
- Winterburn, J.B.; Russell, A.B.; Martin, P.J. Integrated recirculating foam fractionation for the continuous recovery of biosurfactant from fermenters. Biochem. Eng. J. 2011, 54, 132–139. [Google Scholar] [CrossRef]
- Díaz De Rienzo, M.A.; Kamalanathan, I.D.; Martin, P.J. Comparative study of the production of rhamnolipid biosurfactants by B. thailandensis E264 and P. aeruginosa ATCC 9027 using foam fractionation. Process Biochem. 2016, 51, 820–827. [Google Scholar] [CrossRef]
- Biniarz, P.; Henkel, M.; Hausmann, R.; Łukaszewicz, M. Development of a bioprocess for the production of cyclic lipopeptides pseudofactins with efficient purification from collected foam. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef]
- Siemann, M.; Wagner, F. Development of a process mode for the continuous production of rhamnolipids. In Biosurfactants; Kosaric, N., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 1993; Volume 48, pp. 115–131. [Google Scholar]
- Beuker, J.; Steier, A.; Wittgens, A.; Rosenau, F.; Henkel, M.; Hausmann, R. Integrated foam fractionation for heterologous rhamnolipid production with recombinant Pseudomonas putida in a bioreactor. AMB Express 2016, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Anic, I.; Apolonia, I.; Franco, P.; Wichmann, R. Production of rhamnolipids by integrated foam adsorption in a bioreactor system. AMB Express 2018, 8, 122. [Google Scholar] [CrossRef] [PubMed]
- Heyd, M.; Franzreb, M.; Berensmeier, S. Continuous rhamnolipid production with integrated product removal by foam fractionation and magnetic separation of immobilized Pseudomonas aeruginosa. Biotechnol. Prog. 2011, 27, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Blank, L.M.; Küpper, B.; del Amor Villa, E.M.; Wichmann, R. Foam Adsorption. World Patent WO 2013/087674 A1, 20 June 2013. [Google Scholar]
- Bator, I.; Wittgens, A.; Rosenau, F.; Tiso, T.; Blank, L.M. Comparison of three xylose pathways in Pseudomonas putida KT2440 for the synthesis of valuable products. Front. Bioeng. Biotechnol. 2020, 7. [Google Scholar] [CrossRef]
- Zobel, S.; Benedetti, I.; Eisenbach, L.; de Lorenzo, V.; Wierckx, N.; Blank, L.M. Tn7-based device for calibrated heterologous gene expression in Pseudomonas putida. ACS Synth. Biol. 2015, 4, 1341–1351. [Google Scholar] [CrossRef]
- Martinez-Garcia, E.; de Lorenzo, V. Engineering multiple genomic deletions in Gram-negative bacteria: Analysis of the multi-resistant antibiotic profile of Pseudomonas putida KT2440. Environ. Microbiol. 2011, 13, 2702–2716. [Google Scholar] [CrossRef]
- Hartmans, S.; Smits, J.P.; van der Werf, M.J.; Volkering, F.; de Bont, J.A.M. Metabolism of Styrene Oxide and 2-Phenylethanol in the Styrene-Degrading Xanthobacter Strain 124X. Appl. Environ. Microbiol. 1989, 55, 2850–2855. [Google Scholar] [CrossRef]
- Hosseinpour Tehrani, H.; Becker, J.; Bator, I.; Saur, K.; Meyer, S.; Rodrigues Loia, A.C.; Blank, L.M.; Wierckx, N. Integrated strain- and process design enable production of 220 g/L itaconic acid with Ustilago maydis. Biotechnol. Biofuels 2019, 12, 263. [Google Scholar] [CrossRef]
- Behrens, B.; Baune, M.; Jungkeit, J.; Tiso, T.; Blank, L.M.; Hayen, H. High performance liquid chromatography-charged aerosol detection applying an inverse gradient for quantification of rhamnolipid biosurfactants. J. Chromatogr. A 2016, 1455, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Tiso, T.; Sabelhaus, P.; Behrens, B.; Wittgens, A.; Rosenau, F.; Hayen, H.; Blank, L.M. Creating metabolic demand as an engineering strategy in Pseudomonas putida– Rhamnolipid synthesis as an example. Metab. Eng. Commun. 2016, 3, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Anic, I.; Nath, A.; Franco, P.; Wichmann, R. Foam adsorption as an ex situ capture step for surfactants produced by fermentation. J. Biotechnol. 2017, 258, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Junker, B. Foam and its mitigation in fermentation systems. Biotechnol. Prog. 2007, 23, 767–784. [Google Scholar] [CrossRef] [PubMed]
- Delvigne, F.; Lecomte, J.-P. Foam formation and control in bioreactors. In Encyclopedia of Industrial Biotechnology: Bioprocess, Bioseparation, and Cell Technology; Flickinger, M.C., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; Volume 7. [Google Scholar]
- Küpper, B.; Mause, A.; Halka, L.; Imhoff, A.; Nowacki, C.; Wichmann, R. Fermentative Produktion von Monorhamnolipiden im Pilotmaßstab—Herausforderungen der Maßstabsvergrößerung. Chem. Ing. Tech. 2013, 85, 834–840. [Google Scholar] [CrossRef]
- Prins, A.; van’t Riet, K. Proteins and surface effects in fermentation: Foam, antifoam and mass transfer. Trends Biotechnol. 1987, 5, 296–301. [Google Scholar] [CrossRef]
- Burghoff, B. Foam fractionation applications. J. Biotechnol. 2012, 161, 126–137. [Google Scholar] [CrossRef]
- Linke, D.; Zorn, H.; Gerken, B.; Parlar, H.; Berger, R.G. Foam fractionation of exo-lipases from a growing fungus (Pleurotus sapidus). Lipids 2005, 40, 323–327. [Google Scholar] [CrossRef]
- Merz, J.; Burghoff, B.; Zorn, H.; Schembecker, G. Continuous foam fractionation: Performance as a function of operating variables. Sep. Purif. Technol. 2011, 82, 10–18. [Google Scholar] [CrossRef]
- Liu, W.; Zheng, H.; Wu, Z.; Wang, Y. Effects of pH profiles on nisin fermentation coupling with foam separation. Appl. Microbiol. Biotechnol. 2010, 85, 1401–1407. [Google Scholar] [CrossRef]
- Davis, D.A.; Lynch, H.C.; Varley, J. The application of foaming for the recovery of surfactin from B. subtilis ATCC 21332 cultures. Enzym. Microb. Technol. 2001, 28, 346–354. [Google Scholar] [CrossRef]
- Krasowska, A.; Sigler, K. How microorganisms use hydrophobicity and what does this mean for human needs? Front. Cell Infect. Microbiol. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Heipieper, H.J.; Neumann, G.; Cornelissen, S.; Meinhardt, F. Solvent-tolerant bacteria for biotransformations in two-phase fermentation systems. Appl. Microbiol. Biotechnol. 2007, 74, 961–973. [Google Scholar] [CrossRef]
- Eberlein, C.; Baumgarten, T.; Starke, S.; Heipieper, H.J. Immediate response mechanisms of Gram-negative solvent-tolerant bacteria to cope with environmental stress: Cis-trans isomerization of unsaturated fatty acids and outer membrane vesicle secretion. Appl. Microbiol. Biotechnol. 2018, 102, 2583–2593. [Google Scholar] [CrossRef] [PubMed]
- Heard, J.; Harvey, E.; Johnson, B.B.; Wells, J.D.; Angove, M.J. The effect of filamentous bacteria on foam production and stability. Colloids Surf. B Biointerfaces 2008, 63, 21–26. [Google Scholar] [CrossRef]
- Brown, A.K.; Kaul, A.; Varley, J. Continuous foaming for protein recovery: Part I. Recovery of β-casein. Biotechnol. Bioeng. 1999, 62, 278–290. [Google Scholar] [CrossRef]
- Dickinson, J.E.; Laskovski, D.; Stevenson, P.; Galvin, K.P. Enhanced foam drainage using parallel inclined channels in a single-stage foam fractionation column. Chem. Eng. Sci. 2010, 65, 2481–2490. [Google Scholar] [CrossRef]
- Yang, Q.W.; Wu, Z.L.; Zhao, Y.L.; Wang, Y.; Li, R. Enhancing foam drainage using foam fractionation column with spiral internal for separation of sodium dodecyl sulfate. J. Hazard. Mater. 2011, 192, 1900–1904. [Google Scholar] [CrossRef]
- Li, Z.Q.; Zheng, H.J.; Wu, Z.L. Separation of bovine serum albumin by foam fractionation with wire gauze structured packing column. Appl. Biochem. Biotechnol. 2015, 175, 502–512. [Google Scholar] [CrossRef]
- Darton, R.C.; Supino, S.; Sweeting, K.J. Development of a multistaged foam fractionation column. Chem Eng. Process. 2004, 43, 477–482. [Google Scholar] [CrossRef]
- Boonyasuwat, S.; Chavadej, S.; Malakul, P.; Scamehorn, J.F. Surfactant recovery from water using a multistage foam fractionator: Part I Effects of air flow rate, foam height, feed flow rate and number of stages. Sep. Sci. Technol. 2005, 40, 1835–1853. [Google Scholar] [CrossRef]
- Rujirawanich, V.; Chuyingsakultip, N.; Triroj, M.; Malakul, P.; Chavadej, S. Recovery of surfactant from an aqueous solution using continuous multistage foam fractionation: Influence of design parameters. Chem. Eng. Process. 2012, 52, 41–46. [Google Scholar] [CrossRef]
- Li, X.; Evans, G.M.; Stevenson, P. Process intensification of foam fractionation by successive contraction and expansion. Chem. Eng. Res. Des. 2011, 89, 2298–2308. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | System 1 | Medium Volume [L] | Carbon Source | Space-Time Yield [gRL/L·h] | Produced RLs [gRL] | Reference |
|---|---|---|---|---|---|---|
| P. aeruginosa DSM 2874 | I | 18 | glycerol | 0.043 | 16 | [59] |
| P. aeruginosa DSM 2874 | I | 6 | glycerol | 0.023 | 70 | [62] |
| P. putida KT2440 2 | S | 1.5 | glucose | 0.038 | 1 | [60] |
| P. putida EM383 2 | S | 2.5 | glucose | 0.073 | 16 | [61] |
| P. putida KT2440 3 | S | 2 | glucose | 0.24 | 10 | [24] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blesken, C.C.; Strümpfler, T.; Tiso, T.; Blank, L.M. Uncoupling Foam Fractionation and Foam Adsorption for Enhanced Biosurfactant Synthesis and Recovery. Microorganisms 2020, 8, 2029. https://doi.org/10.3390/microorganisms8122029
Blesken CC, Strümpfler T, Tiso T, Blank LM. Uncoupling Foam Fractionation and Foam Adsorption for Enhanced Biosurfactant Synthesis and Recovery. Microorganisms. 2020; 8(12):2029. https://doi.org/10.3390/microorganisms8122029
Chicago/Turabian StyleBlesken, Christian C., Tessa Strümpfler, Till Tiso, and Lars M. Blank. 2020. "Uncoupling Foam Fractionation and Foam Adsorption for Enhanced Biosurfactant Synthesis and Recovery" Microorganisms 8, no. 12: 2029. https://doi.org/10.3390/microorganisms8122029
APA StyleBlesken, C. C., Strümpfler, T., Tiso, T., & Blank, L. M. (2020). Uncoupling Foam Fractionation and Foam Adsorption for Enhanced Biosurfactant Synthesis and Recovery. Microorganisms, 8(12), 2029. https://doi.org/10.3390/microorganisms8122029