Epidemiology and Sequence-Based Evolutionary Analysis of Circulating Non-Polio Enteroviruses


Abstract
1. Introduction
2. EV Classification and Nomenclature
3. Enterovirus Association with Severe Disease
3.1. Enterovirus A
3.2. Enterovirus B
3.3. Enterovirus C
3.4. Enterovirus D
4. Enterovirus Evolutionary Relatedness

5. Enterovirus Surveillance and Sequence Coverage
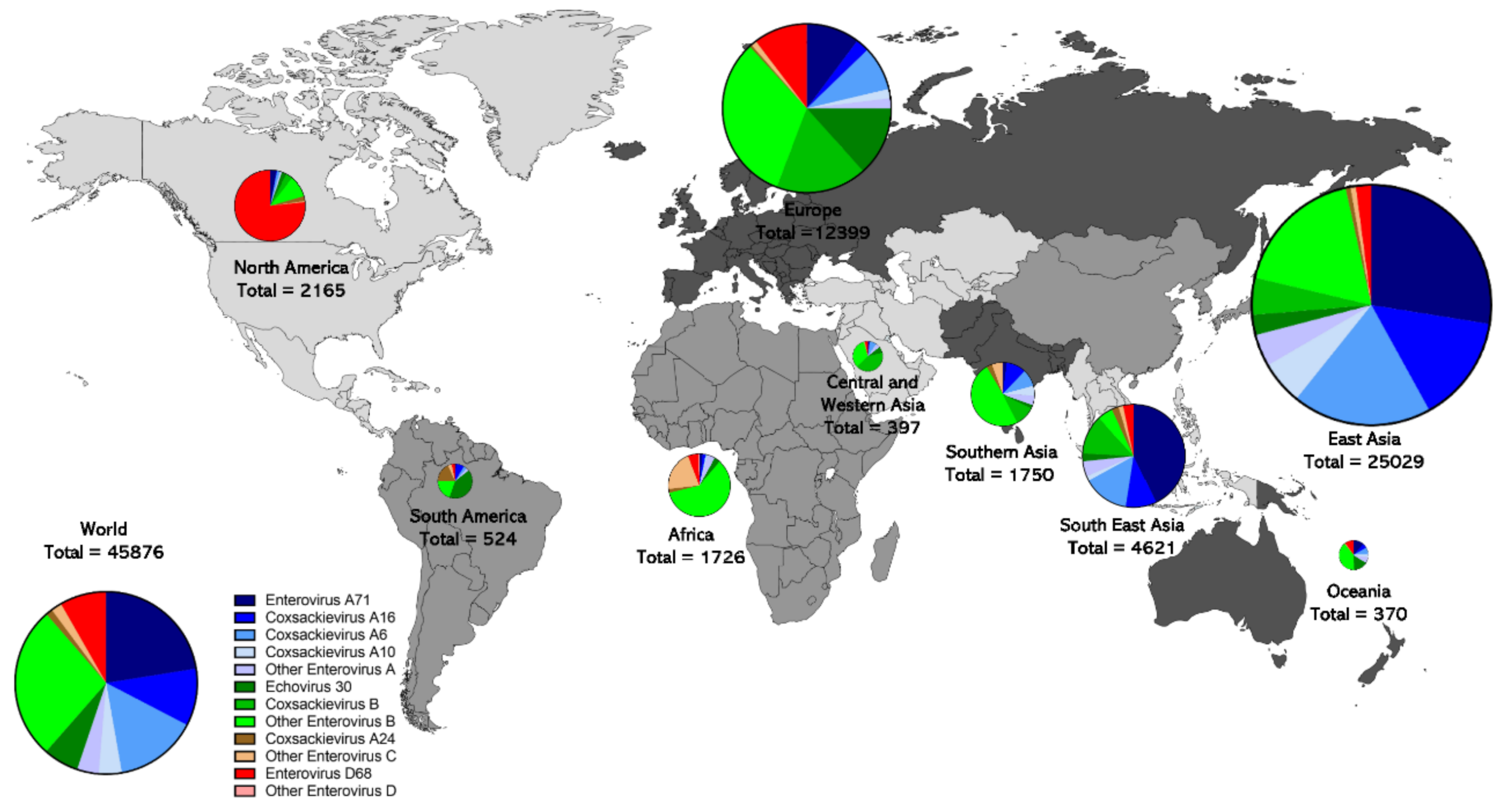
6. Geographic Distribution of Enterovirus Sequences
7. Enterovirus Circulation and Temporal Dynamics
8. Conclusions/Perspective
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pallansch, M.A.; Roos, R.P. Enteroviruses: Polioviruses, coxsackieviruses, echoviruses, and newer enteroviruses. In Fields Virology, 4th ed.; Knipe, D.M., Howley, P.M., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Stephen, E.S., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2001; pp. 723–775. [Google Scholar]
- Tangermann, R.H.; Aylward, R.; Birmingham, M.; Horner, R.; Olivé, J.M.; Nkowane, B.M.; Hull, H.F.; Burton, A. Current status of the global eradication of poliomyelitis. World Health Stat. Q. Rapp. Trimest. Stat. Sanit. Mond. 1997, 50, 188–194. [Google Scholar]
- Shaghaghi, M.; Soleyman-Jahi, S.; Abolhassani, H.; Yazdani, R.; Azizi, G.; Rezaei, N.; Barbouche, M.-R.; McKinlay, M.A.; Aghamohammadi, A. New insights into physiopathology of immunodeficiency-associated vaccine-derived poliovirus infection; systematic review of over 5 decades of data. Vaccine 2018, 36, 1711–1719. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Bolivar-Wagers, S.; Srinivas, N.; Holubar, M.; Maldonado, Y. Immunodeficiency-related vaccine-derived poliovirus (iVDPV) cases: A systematic review and implications for polio eradication. Vaccine 2015, 33, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Burns, C.C.; Diop, O.M.; Sutter, R.W.; Kew, O.M. Vaccine-Derived Polioviruses. J. Infect. Dis. 2014, 210, S283–S293. [Google Scholar] [CrossRef]
- Royston, L.; Tapparel, C. Rhinoviruses and Respiratory Enteroviruses: Not as Simple as ABC. Viruses 2016, 8, 16. [Google Scholar] [CrossRef]
- Drysdale, S.B.; Mejias, A.; Ramilo, O. Rhinovirus—Not just the common cold. J. Infect. 2017, 74, S41–S46. [Google Scholar] [CrossRef]
- (ICTV) ICoToV. Virus Taxonomy: 2019 Release. 28 March 2019. Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 11 April 2019).
- Oberste, M.S.; Maher, K.; Kilpatrick, D.R.; Pallansch, M.A. Molecular Evolution of the Human Enteroviruses: Correlation of Serotype with VP1 Sequence and Application to Picornavirus Classification. J. Virol. 1999, 73, 1941–1948. [Google Scholar] [CrossRef]
- Lukashev, A.N.; Vakulenko, Y.A. Molecular evolution of types in non-polio enteroviruses. J. Gen. Virol. 2017, 98, 2968–2981. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, J.; Zhang, S.; Lee, A.J.; Sun, G.; Larsen, C.N.; Zhao, H.; Gu, Z.; He, S.; Klem, E.B.; et al. Genetic changes found in a distinct clade of Enterovirus D68 associated with paralysis during the 2014 outbreak. Virus Evol. 2016, 2, vew015. [Google Scholar] [CrossRef]
- Pallansch, M.A.; Sandhu, H.S. The Eradication of Polio—Progress and Challenges. N. Engl. J. Med. 2006, 355, 2508–2511. [Google Scholar] [CrossRef]
- Witsø, E.; Palacios, G.; Cinek, O.; Stene, L.C.; Grinde, B.; Janowitz, D.; Lipkin, W.I.; Rønningen, K.S. High Prevalence of Human Enterovirus A Infections in Natural Circulation of Human Enteroviruses. J. Clin. Microbiol. 2006, 44, 4095–4100. [Google Scholar] [CrossRef] [PubMed]
- Ma, E.; Chan, K.C.; Cheng, P.; Wong, C.; Chuang, S.K. The enterovirus 71 epidemic in 2008—Public health implications for Hong Kong. Int. J. Infect. Dis. 2010, 14, e775–e780. [Google Scholar] [CrossRef] [PubMed]
- Tapparel, C.; Siegrist, F.; Petty, T.J.; Kaiser, L. Picornavirus and enterovirus diversity with associated human diseases. Infect. Genet. Evol. 2013, 14, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.; Chong, P. Is a multivalent hand, foot, and mouth disease vaccine feasible? Hum. Vaccines Immunother. 2015, 11, 2688–2704. [Google Scholar] [CrossRef]
- Suresh, S.; Forgie, S.; Robinson, J. Non-polio Enterovirus detection with acute flaccid paralysis: A systematic review. J. Med. Virol. 2018, 90, 3–7. [Google Scholar] [CrossRef]
- Liu, J.; Xiang, X.; Pu, Z.; Long, Y.; Xiao, D.; Zhang, W.; Li, Q.; Li, X.; Li, S.; Shao, Z.; et al. Epidemic pattern of hand-foot-and-mouth disease in Xi’an, China from 2008 through 2015. BMC Infect. Dis. 2019, 19, 19. [Google Scholar] [CrossRef]
- Xing, W.; Liao, Q.; Viboud, C.; Zhang, J.; Sun, J. Epidemiological characteristics of hand-foot-and-mouth disease in China, 2008–2012. Lancet Infect. Dis. 2014, 14, 308. [Google Scholar] [CrossRef]
- Shi, L.; Zhao, H.; Wu, D. Modelling and analysis of HFMD with the effects of vaccination, contaminated environments and quarantine in mainland China. Math. Biosci. Eng. MBE 2018, 16, 474–500. [Google Scholar] [CrossRef]
- Takahashi, S.; Metcalf, C.J.E.; Arima, Y.; Fujimoto, T.; Shimizu, H.; Van Doorn, H.R.; Le Van, T.; Chan, Y.-F.; Farrar, J.J.; Oishi, K.; et al. Epidemic dynamics, interactions and predictability of enteroviruses associated with hand, foot and mouth disease in Japan. J. R. Soc. Interface 2018, 15, 20180507. [Google Scholar] [CrossRef]
- Huang, W.; Huang, L.; Lu, C.; Cheng, A.-L.; Chang, L.-Y. Atypical hand-foot-mouth disease in children: A hospital-based prospective cohort study. Virol. J. 2013, 10, 209. [Google Scholar] [CrossRef]
- Chua, K.; Kasri, A. Hand foot and mouth disease due to enterovirus 71 in Malaysia. Virol. Sin. 2011, 26, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Ang, L.W.; Koh, B.K.; Chan, K.P.; Chua, L.T.; James, L.; Goh, K.T. Epidemiology and control of hand, foot and mouth disease in Singapore, 2001–2007. Ann. Acad. Med. Singap. 2009, 38, 106–112. [Google Scholar] [PubMed]
- Nhan, L.N.T.; Hong, N.T.T.; Nhu, L.N.T.; Nguyet, L.A.; Ny, N.T.H.; Thanh, T.T.; Han, D.D.K.; Van, H.M.T.; Thwaites, C.L.; Hien, T.T.; et al. Severe enterovirus A71 associated hand, foot and mouth disease, Vietnam, 2018: Preliminary report of an impending outbreak. Eurosurveillance 2018, 23, 1800590. [Google Scholar] [CrossRef] [PubMed]
- Anh, N.T.; Nhu, L.N.T.; Van, H.M.T.; Hong, N.T.T.; Thanh, T.T.; Hang, V.T.T.; Ny, N.T.H.; Nguyet, L.A.; Phuong, T.T.L.; Nhan, L.N.T.; et al. Emerging Coxsackievirus A6 Causing Hand, Foot and Mouth Disease, Vietnam. Emerg. Infect. Dis. 2018, 24, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Hoang, M.T.V.; Nguyen, T.A.; Tran, T.T.; Vu, T.T.H.; Le, N.T.N.; Nguyen, T.H.N.; Le, T.H.N.; Nguyen, H.T.T.; Nguyen, T.H.; Le, N.T.N.; et al. Clinical and aetiological study of hand, foot and mouth disease in southern Vietnam, 2013–2015: Inpatients and outpatients. Int. J. Infect. Dis. 2019, 80, 1–9. [Google Scholar] [CrossRef]
- Noisumdaeng, P.; Korkusol, A.; Prasertsopon, J.; Sangsiriwut, K.; Chokephaibulkit, K.; Mungaomklang, A.; Thitithanyanont, A.; Buathong, R.; Guntapong, R.; Puthavathana, P. Longitudinal study on enterovirus A71 and coxsackievirus A16 genotype/subgenotype replacements in hand, foot and mouth disease patients in Thailand, 2000–2017. Int. J. Infect. Dis. 2019, 80, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Puenpa, J.; Auphimai, C.; Korkong, S.; Vongpunsawad, S.; Poovorawan, Y. Enterovirus A71 Infection, Thailand, 2017. Emerg. Infect. Dis. 2018, 24, 1386–1387. [Google Scholar] [CrossRef] [PubMed]
- Duong, V.; Mey, C.; Eloit, M.; Zhu, H.; Danet, L.; Huang, Z.; Zou, G.; Tarantola, A.; Cheval, J.; Perot, P.; et al. Molecular epidemiology of human enterovirus 71 at the origin of an epidemic of fatal hand, foot and mouth disease cases in Cambodia. Emerg. Microbes Infect. 2016, 5, e104. [Google Scholar] [CrossRef] [PubMed]
- Pons-Salort, M.; Grassly, N.C. Serotype-specific immunity explains the incidence of diseases caused by human enteroviruses. Science 2018, 361, 800–803. [Google Scholar] [CrossRef]
- NikNadia, N.; Sam, I.C.; Rampal, S.; WanNorAmalina, W.; NurAtifah, G.; Verasahib, K.; Ong, C.C.; MohdAdib, M.; Chan, Y.F. Cyclical Patterns of Hand, Foot and Mouth Disease Caused by Enterovirus A71 in Malaysia. PLoS Negl. Trop. Dis. 2016, 10, e0004562. [Google Scholar] [CrossRef]
- Chen, L.; Yao, X.J.; Xu, S.J.; Yang, H.; Wu, C.L.; Lu, J.; Xu, W.-B.; Zhang, H.; Meng, J.; Zhang, Y.; et al. Molecular surveillance of coxsackievirus A16 reveals the emergence of a new clade in mainland China. Arch. Virol. 2019, 164, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Ji, T.; Guo, Y.; Huang, W.; Shi, Y.; Xu, Y.; Tong, W.; Yao, W.; Tan, Z.; Zeng, H.; Ma, J.; et al. The emerging sub-genotype C2 of CoxsackievirusA10 Associated with Hand, Foot and Mouth Disease extensively circulating in mainland of China. Sci. Rep. 2018, 8, 13357. [Google Scholar] [CrossRef] [PubMed]
- Puenpa, J.; Vongpunsawad, S.; Österback, R.; Waris, M.; Eriksson, E.; Albert, J.; Midgley, S.; Fischer, T.K.; Eis-Hübinger, A.M.; Cabrerizo, M.; et al. Molecular epidemiology and the evolution of human coxsackievirus A6. J. Gen. Virol. 2016, 97, 3225–3231. [Google Scholar] [CrossRef]
- Wang, S.H.; Wang, A.; Liu, P.P.; Zhang, W.Y.; Du, J.; Xu, S.; Liu, G.-C.; Zheng, B.-S.; Huan, C.; Zhao, K.; et al. Divergent Pathogenic Properties of Circulating Coxsackievirus A6 Associated with Emerging Hand, Foot, and Mouth Disease. J. Virol. 2018, 92, e00303-18. [Google Scholar] [CrossRef] [PubMed]
- Bian, L.; Wang, Y.; Yao, X.; Mao, Q.; Xu, M.; Liang, Z. Coxsackievirus A6: A new emerging pathogen causing hand, foot and mouth disease outbreaks worldwide. Expert Rev. Anti Infect. Ther. 2015, 13, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhang, Y.; Ji, T.; Gu, X.; Yang, Q.; Zhu, S.; Xu, W.; Xu, Y.; Shi, Y.; Huang, X.; et al. Persistent circulation of Coxsackievirus A6 of genotype D3 in mainland of China between 2008 and 2015. Sci. Rep. 2017, 7, 5491. [Google Scholar] [CrossRef]
- Huang, J.; Liao, Q.; Ooi, M.H.; Cowling, B.J.; Chang, Z.; Wu, P.; Liu, F.; Li, Y.; Luo, L.; Yu, S.; et al. Epidemiology of Recurrent Hand, Foot and Mouth Disease, China, 2008–2015. Emerg. Infect. Dis. 2018, 24, 432–442. [Google Scholar] [CrossRef]
- Bessaud, M.; Razafindratsimandresy, R.; Nougairède, A.; Joffret, M.L.; Deshpande, J.M.; Dubot-Pérès, A.; Héraud, J.-M.; De Lamballerie, X.; Delpeyroux, F.; Bailly, J.-L. Molecular Comparison and Evolutionary Analyses of VP1 Nucleotide Sequences of New African Human Enterovirus 71 Isolates Reveal a Wide Genetic Diversity. PLoS ONE 2014, 9, e90624. [Google Scholar] [CrossRef]
- Wang, J.; Teng, Z.; Cui, X.; Li, C.; Pan, H.; Zheng, Y.; Mao, S.; Yang, Y.; Wu, L.; Guo, X.; et al. Epidemiological and serological surveillance of hand-foot-and-mouth disease in Shanghai, China, 2012–2016. Emerg. Microbes Infect. 2018, 7, 8. [Google Scholar] [CrossRef]
- Li, J.; Pan, H.; Wang, X.; Zhu, Q.; Ge, Y.; Cai, J.; Li, Y.; Xia, A.; Hu, J.; Zeng, M. Epidemiological surveillance of hand, foot and mouth disease in Shanghai in 2014–2016, prior to the introduction of the enterovirus 71 vaccine. Emerg. Microbes Infect. 2018, 7, 37. [Google Scholar] [CrossRef]
- Wang, J.; Hu, T.; Sun, D.; Ding, S.; Carr, M.J.; Xing, W.; Li, S.; Wang, X.; Shi, W. Epidemiological characteristics of hand, foot, and mouth disease in Shandong, China, 2009–2016. Sci. Rep. 2017, 7, 8900. [Google Scholar] [CrossRef] [PubMed]
- Ooi, M.H.; Wong, S.C.; Lewthwaite, P.; Cardosa, M.J.; Solomon, T. Clinical features, diagnosis, and management of enterovirus 71. Lancet Neurol. 2010, 9, 1097–1105. [Google Scholar] [CrossRef]
- Teoh, H.L.; Mohammad, S.S.; Britton, P.N.; Kandula, T.; Lorentzos, M.S.; Booy, R.; Jones, C.A.; Rawlinson, W.; Ramachandran, V.; Rodriguez, M.L.; et al. Clinical Characteristics and Functional Motor Outcomes of Enterovirus 71 Neurological Disease in Children. JAMA Neurol. 2016, 73, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-M.; Liu, C.C.; Tseng, H.W.; Wang, J.-R.; Huang, C.-C.; Chen, Y.-J.; Yang, Y.; Lin, S.; Yeh, T. Clinical Spectrum of Enterovirus 71 Infection in Children in Southern Taiwan, with an Emphasis on Neurological Complications. Clin. Infect. Dis. 1999, 29, 184–190. [Google Scholar] [CrossRef] [PubMed]
- McMinn, P.; Stratov, I.; Nagarajan, L.; Davis, S. Neurological Manifestations of Enterovirus 71 Infection in Children during an Outbreak of Hand, Foot, and Mouth Disease in Western Australia. Clin. Infect. Dis. 2001, 32, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Vélez, C.M.; Anderson, M.S.; Robinson, C.C.; McFarland, E.J.; Nix, W.A.; Pallansch, M.A.; Oberste, M.S.; Glodé, M.P. Outbreak of Neurologic Enterovirus Type 71 Disease: A Diagnostic Challenge. Clin. Infect. Dis. 2007, 45, 950–957. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chang, Y.C.; Huang, C.C.; Lui, C.C.; Lee, K.W.; Huang, S.C. Acute flaccid paralysis in infants and young children with enterovirus 71 infection: MR imaging findings and clinical correlates. Am. J. Neuroradiol. 2001, 22, 200–205. [Google Scholar]
- Schubert, R.D.; Hawes, I.A.; Ramachandran, P.S.; Ramesh, A.; Crawford, E.D.; Pak, J.E.; Wu, W.; Cheung, C.K.; O’Donovan, B.D.; Tato, C.M.; et al. Pan-viral serology implicates enteroviruses in acute flaccid myelitis. Nat. Med. 2019, 25, 1748–1752. [Google Scholar] [CrossRef]
- Yu, P.; Bao, L.; Xu, L.; Li, F.; Lv, Q.; Deng, W.; Xu, Y.; Qin, C. Neurotropism In Vitro and Mouse Models of Severe and Mild Infection with Clinical Strains of Enterovirus 71. Viruses 2017, 9, 351. [Google Scholar] [CrossRef]
- Lin, J.; Shih, S. Cell and tissue tropism of enterovirus 71 and other enteroviruses infections. J. Biomed. Sci. 2014, 21, 18. [Google Scholar] [CrossRef]
- Mandary, M.B.; Poh, C.L. Changes in the EV-A71 Genome through Recombination and Spontaneous Mutations: Impact on Virulence. Viruses 2018, 10, 320. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.; Wang, Y.; Bian, L.; Xu, M.; Liang, Z. EV-A71 vaccine licensure: A first step for multivalent enterovirus vaccine to control HFMD and other severe diseases. Emerg. Microbes Infect. 2016, 5, e75. [Google Scholar] [CrossRef] [PubMed]
- Yee, P.T.I.; Poh, C.L. Impact of genetic changes, pathogenicity and antigenicity on Enterovirus- A71 vaccine development. Virology 2017, 506, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Wang, H.; Liu, Y.; Li, Y.; Jiang, P.; Liu, G.; Lin, X.; Li, M.; Wang, S.; Ji, F.; et al. Non-Polio Enteroviruses from Acute Flaccid Paralysis Surveillance in Shandong Province, China, 1988–2013. Sci. Rep. 2014, 4, 6167. [Google Scholar] [CrossRef] [PubMed]
- Masa-Calles, J.; Torner, N.; López-Perea, N.; Torres de Mier, M.V.; Fernández-Martínez, B.; Cabrerizo, M.; Gallardo-García, V.; Malo, C.; Margolles-Martins, M.J.; Portell, M.; et al. Acute flaccid paralysis (AFP) surveillance: Challenges and opportunities from 18 years’ experience, Spain, 1998 to 2015. Eurosurveillance 2018, 23, 1700423. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Garcia, M.D.; Kebe, O.; Fall, A.D.; Ndiaye, K. Identification and molecular characterization of non-polio enteroviruses from children with acute flaccid paralysis in West Africa, 2013–2014. Sci. Rep. 2017, 7, 3808. [Google Scholar] [CrossRef]
- Delogu, R.; Battistone, A.; Buttinelli, G.; Fiore, S.; Fontana, S.; Amato, C.; Cristiano, K.; Gamper, S.; Simeoni, J.; Frate, R.; et al. Poliovirus and Other Enteroviruses from Environmental Surveillance in Italy, 2009–2015. Food Environ. Virol. 2018, 10, 333–342. [Google Scholar] [CrossRef]
- Verma, N.A.; Zheng, X.T.; Harris, M.U.; Cadichon, S.B.; Melin-Aldana, H.; Khetsuriani, N.; Oberste, M.S.; Shulman, S.T. Outbreak of Life-Threatening Coxsackievirus B1 Myocarditis in Neonates. Clin. Infect. Dis. 2009, 49, 759–763. [Google Scholar] [CrossRef]
- Garmaroudi, F.S.; Marchant, D.; Hendry, R.; Luo, H.; Yang, D.; Ye, X.; Shi, J.; McManus, B.M. Coxsackievirus B3 replication and pathogenesis. Future Microbiol. 2015, 10, 629–653. [Google Scholar] [CrossRef]
- Schmidt, N.J.; Magoffin, R.L.; Lennette, E.H. Association of group B coxsackie viruses with cases of pericarditis, myocarditis, or pleurodynia by demonstration of immunoglobulin M antibody. Infect. Immun. 1973, 8, 341–348. [Google Scholar] [CrossRef]
- Yin-Murphy, M.; Lim, K.H.; Ho, Y.M. A coxsackievirus type A24 epidemic of acute conjunctivitis. Southeast Asian J. Trop. Med. Public Health 1976, 7, 1–5. [Google Scholar]
- Tavares, F.; Campos, R.D.M.; Burlandy, F.; Fontella, R.; De Melo, M.M.M.; Da Costa, E.V.; Da Silva, E.E. Molecular Characterization and Phylogenetic Study of Coxsackievirus A24v Causing Outbreaks of Acute Hemorrhagic Conjunctivitis (AHC) in Brazil. PLoS ONE 2011, 6, e23206. [Google Scholar] [CrossRef] [PubMed]
- Medina, N.H.; Haro-Munoz, E.; Pellini, A.C.; Machado, B.C.; Russo, D.H.; Timenetsky, M.D.; Carmona, R.D.C.C. Acute hemorrhagic conjunctivitis epidemic in Sao Paulo State, Brazil, 2011. Rev. Panam. Salud Publica 2016, 39, 137–141. [Google Scholar] [PubMed]
- Zhang, Y.; Sun, Q.; Cui, H.; Yan, D.; Fan, Q.; Song, Y.; Zhu, S.; Li, X.; Huang, G.; Ji, T.; et al. Circulation of multiple serotypes of highly divergent enterovirus C in the Xinjiang Uighur Autonomous Region of China. Sci. Rep. 2016, 6, 33595. [Google Scholar] [CrossRef] [PubMed]
- Enfissi, A.; Joffret, M.L.; Delaune, D.; Delpeyroux, F.; Rousset, D.; Bessaud, M. Coxsackievirus A24 Variant Associated with Acute Haemorrhagic Conjunctivitis Cases, French Guiana, 2017. Intervirology 2017, 60, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Shukla, D.; Kumar, A.; Srivastava, S.; Dhole, T.N. Molecular identification and phylogenetic study of coxsackievirus A24 variant isolated from an outbreak of acute hemorrhagic conjunctivitis in India in 2010. Arch. Virol. 2013, 158, 679–684. [Google Scholar] [CrossRef]
- Burr, S.E.; Sillah, A.; Joof, H.; Bailey, R.L.; Holland, M.J. An outbreak of acute haemorrhagic conjunctivitis associated with coxsackievirus A24 variant in The Gambia, West Africa. BMC Res. Notes 2017, 10, 692. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, N.; Huang, X.; Jin, X.; Geng, X.; Chan, T.C.; Liu, S.-L. Molecular epidemiology of acute hemorrhagic conjunctivitis caused by coxsackie A type 24 variant in China, 2004–2014. Sci. Rep. 2017, 7, 45202. [Google Scholar] [CrossRef]
- Schieble, J.H.; Fox, V.L.; Lennette, E.H. A probable new human picornavirus associated with respiratory diseases. Am. J. Epidemiol. 1967, 85, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T.; Oshitani, H. Global reemergence of enterovirus D68 as an important pathogen for acute respiratory infections. Rev. Med. Virol. 2015, 25, 102–114. [Google Scholar] [CrossRef]
- Holm-Hansen, C.C.; Midgley, S.E.; Fischer, T.K. Global emergence of enterovirus D68: A systematic review. Lancet Infect. Dis. 2016, 16, e64–e75. [Google Scholar] [CrossRef]
- Kramer, R.; Sabatier, M.; Wirth, T.; Pichon, M.; Lina, B.; Schuffenecker, I.; Josset, L. Molecular diversity and biennial circulation of enterovirus D68: A systematic screening study in Lyon, France, 2010 to 2016. Eurosurveillance 2018, 23, 1700711. [Google Scholar] [CrossRef] [PubMed]
- Bowers, J.R.; Valentine, M.; Harrison, V.; Fofanov, V.Y.; Gillece, J.; Delisle, J.; Patton, B.; Schupp, J.; Sheridan, K.; Lemmer, D.; et al. Genomic Analyses of Acute Flaccid Myelitis Cases among a Cluster in Arizona Provide Further Evidence of Enterovirus D68 Role. mBio 2019, 10, e02262-18. [Google Scholar] [CrossRef]
- Force TUKAFP. An increase in reports of acute flaccid paralysis (AFP) in the United Kingdom, 1 January 2018–21 January 2019: Early findings. Eurosurveillance 2019, 24, 1900093. [Google Scholar] [CrossRef]
- Cottrell, S.; Moore, C.; Perry, M.; Hilvers, E.; Williams, C.; Shankar, A.G. Prospective enterovirus D68 (EV-D68) surveillance from September 2015 to November 2018 indicates a current wave of activity in Wales. Eurosurveillance 2018, 23, 1800578. [Google Scholar] [CrossRef] [PubMed]
- Aliabadi, N.; Messacar, K.; Pastula, D.M.; Robinson, C.C.; Leshem, E.; Sejvar, J.J.; Nix, W.A.; Oberste, M.S.; Feikin, D.R.; Dominguez, S.R. Enterovirus D68 Infection in Children with Acute Flaccid Myelitis, Colorado, USA, 2014. Emerg. Infect. Dis. 2016, 22, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- McKay, S.L.; Lee, A.D.; Lopez, A.S.; Nix, W.A.; Dooling, K.L.; Keaton, A.A.; Spence-Davizon, E.; Herlihy, R.; Clark, T.A.; Hopkins, S.E.; et al. Increase in Acute Flaccid Myelitis—United States, 2018. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 1273–1275. [Google Scholar] [CrossRef]
- Harrison, C.J.; Weldon, W.C.; Pahud, B.A.; Jackson, M.A.; Oberste, M.S.; Selvarangan, R. Neutralizing Antibody against Enterovirus D68 in Children and Adults before 2014 Outbreak, Kansas City, Missouri, USA(1). Emerg. Infect. Dis. 2019, 25, 585–588. [Google Scholar] [CrossRef]
- Brown, D.M.; Hixon, A.M.; Oldfield, L.M.; Zhang, Y.; Novotny, M.; Wang, W.; Das, S.R.; Shabman, R.S.; Tyler, K.L.; Scheuermann, R.H. Contemporary Circulating Enterovirus D68 Strains Have Acquired the Capacity for Viral Entry and Replication in Human Neuronal Cells. mBio 2018, 9, e01954-18. [Google Scholar] [CrossRef]
- Carballo, C.M.; Erro, M.G.; Sordelli, N.; Vazquez, G.; Mistchenko, A.S.; Cejas, C.; Rodriguez, M.; Cisterna, D.M.; Freire, M.C.; Contrini, M.M.; et al. Acute Flaccid Myelitis Associated with Enterovirus D68 in Children, Argentina, 2016. Emerg. Infect. Dis. 2019, 25, 573. [Google Scholar] [CrossRef]
- Knoester, M.; Helfferich, J.; Poelman, R.; Van Leer-Buter, C.; Brouwer, O.F.; Niesters, H.G.M. Twenty-nine Cases of Enterovirus-D68–associated Acute Flaccid Myelitis in Europe 2016: A Case Series and Epidemiologic Overview. Pediatr. Infect. Dis. J. 2019, 38, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Bal, A.; Sabatier, M.; Wirth, T.; Coste-Burel, M.; Lazrek, M.; Stefic, K.; Brengel-Pesce, K.; Morfin, F.; Lina, B.; Schuffenecker, I.; et al. Emergence of enterovirus D68 clade D1, France, August to November 2018. Eurosurveillance 2019, 24, 1800699. [Google Scholar] [CrossRef] [PubMed]
- Pellegrinelli, L.; Giardina, F.; Lunghi, G.; Renteria, S.C.; Greco, L.; Fratini, A.; Galli, C.; Piralla, A.; Binda, S.; Pariani, E.; et al. Emergence of divergent enterovirus (EV) D68 sub-clade D1 strains, northern Italy, September to October 2018. Eurosurveillance 2019, 24, 1900090. [Google Scholar] [CrossRef]
- Kaida, A.; Iritani, N.; Yamamoto, S.P.; Kanbayashi, D.; Hirai, Y.; Togawa, M.; Amo, K.; Kohdera, U.; Nishigaki, T.; Shiomi, M.; et al. Distinct genetic clades of enterovirus D68 detected in 2010, 2013, and 2015 in Osaka City, Japan. PLoS ONE 2017, 12, e0184335. [Google Scholar] [CrossRef] [PubMed]
- Sejvar, J.J.; Lopez, A.S.; Cortese, M.M.; Leshem, E.; Pastula, D.M.; Miller, L.; Glaser, C.; Kambhampati, A.; Shioda, K.; Aliabadi, N.; et al. Acute Flaccid Myelitis in the United States, August–December 2014: Results of Nationwide Surveillance. Clin. Infect. Dis. 2016, 63, 737–745. [Google Scholar] [CrossRef]
- Messacar, K.; Schreiner, T.L.; Maloney, A.J.; Wallace, A.; Ludke, J.; Oberste, M.S.; Nix, W.A.; Robinson, C.C.; Glodé, M.P.; Abzug, M.J.; et al. A cluster of acute flaccid paralysis and cranial nerve dysfunction temporally associated with an outbreak of enterovirus D68 in children in Colorado, USA. Lancet 2015, 385, 1662–1671. [Google Scholar] [CrossRef]
- Van Haren, K.; Ayscue, P.; Waubant, E.; Clayton, A.; Sheriff, H.; Yagi, S.; Glenn-Finer, R.; Padilla, T.; Strober, J.B.; Aldrovandi, G.M.; et al. Acute Flaccid Myelitis of Unknown Etiology in California, 2012–2015. JAMA 2015, 314, 2663–2671. [Google Scholar] [CrossRef]
- Ayscue, P.; Van Haren, K.; Sheriff, H.; Waubant, E.; Waldron, P.; Yagi, S.; Yen, C.; Clayton, A.; Padilla, T.; Pan, C.; et al. Acute Flaccid Paralysis with Anterior Myelitis—California, June 2012–June 2014. MMWR Morb. Mortal. Wkly. Rep. 2014, 63, 903–906. [Google Scholar]
- Greninger, A.L.; Naccache, S.N.; Messacar, K.; Clayton, A.; Yu, G.; Somasekar, S.; Federman, S.; Stryke, D.; Anderson, C.; Yagi, S.; et al. A novel outbreak enterovirus D68 strain associated with acute flaccid myelitis cases in the USA (2012–2014): A retrospective cohort study. Lancet Infect. Dis. 2015, 15, 671–682. [Google Scholar] [CrossRef]
- Messacar, K.; Schreiner, T.L.; Van Haren, K.; Yang, M.; Glaser, C.A.; Tyler, K.L.; Dominguez, S.R. Acute flaccid myelitis: A clinical review of US cases 2012–2015. Ann. Neurol. 2016, 80, 326–338. [Google Scholar] [CrossRef]
- Messacar, K.; Abzug, M.J.; Dominguez, S.R. 2014 outbreak of enterovirus D68 in North America. J. Med. Virol. 2016, 88, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.F.; Montmayeur, A.; Castro, C.; Cone, M.; Stringer, J.; Lamson, D.M.; Rogers, S.L.; Chern, S.-W.W.; Magaña, L.; Marine, R.L.; et al. Detection and Genomic Characterization of Enterovirus D68 in Respiratory Samples Isolated in the United States in 2016. Genome Announc. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Lang, M.; Mirand, A.; Savy, N.; Henquell, C.; Maridet, S.; Perignon, R.; Labbé, A.; Peigue-Lafeuille, H. Acute flaccid paralysis following enterovirus D68 associated pneumonia, France, 2014. Eurosurveillance 2014, 19, 20952. [Google Scholar] [CrossRef] [PubMed]
- Bragstad, K.; Jakobsen, K.; Rojahn, A.E.; Skram, M.K.; Vainio, K.; Holberg-Petersen, M.; Hungnes, O.; Dudman, S.G.; Kran, A.-M.B. High frequency of enterovirus D68 in children hospitalised with respiratory illness in Norway, autumn 2014. Influ. Other Respir. Viruses 2014, 9, 59–63. [Google Scholar] [CrossRef]
- Levy, A.; Roberts, J.A.; Lang, J.; Tempone, S.; Kesson, A.; Dofai, A.; Daley, A.J.; Thorley, B.R.; Speers, D.J. Enterovirus D68 disease and molecular epidemiology in Australia. J. Clin. Virol. 2015, 69, 117–121. [Google Scholar] [CrossRef]
- Dyda, A.; Stelzer-Braid, S.; Adam, D.; Chughtai, A.A.; MacIntyre, C.R. The association between acute flaccid myelitis (AFM) and Enterovirus D68 (EV-D68)—What is the evidence for causation? Eurosurveillance 2018, 23. [Google Scholar] [CrossRef]
- Chong, P.F.; Kira, R.; Mori, H.; Okumura, A.; Torisu, H.; Yasumoto, S.; Shimizu, H.; Fujimoto, T.; Hanaoka, N.; Kusunoki, S.; et al. Clinical Features of Acute Flaccid Myelitis Temporally Associated With an Enterovirus D68 Outbreak: Results of a Nationwide Survey of Acute Flaccid Paralysis in Japan, August–December 2015. Clin. Infect. Dis. 2018, 66, 653–664. [Google Scholar] [CrossRef]
- Xiang, Z.; Xie, Z.; Liu, L.; Ren, L.; Xiao, Y.; Paranhos-Baccalà, G.; Wang, J. Genetic divergence of enterovirus D68 in China and the United States. Sci. Rep. 2016, 6, 27800. [Google Scholar] [CrossRef]
- Zhang, T.G.; Li, H.Q.; Li, A.H.; Chen, M.; Gong, C.; Luo, M.; Dong, M.; Huang, F. The Genomic Characterization of Enterovirus D68 from 2011 to 2015 in Beijing, China. Biomed. Environ. Sci. 2016, 29, 675–677. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, X.; Dai, W.; Liu, Q.; Xiong, P.; Wang, S.; Geng, L.; Gong, S.; Huang, Z. A Mouse Model of Enterovirus D68 Infection for Assessment of the Efficacy of Inactivated Vaccine. Viruses 2018, 10, 58. [Google Scholar] [CrossRef]
- Hixon, A.M.; Yu, G.; Leser, J.S.; Yagi, S.; Clarke, P.; Chiu, C.Y.; Tyler, K.L. A mouse model of paralytic myelitis caused by enterovirus D68. PLoS Pathog. 2017, 13, e1006199. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Hassan, F.; Schuster, J.E.; Simenauer, A.; Selvarangan, R.; Halpin, R.A.; Lin, X.; Fedorova, N.; Stockwell, T.B.; Lam, T.T.-Y.; et al. Molecular Evolution and Intraclade Recombination of Enterovirus D68 during the 2014 Outbreak in the United States. J. Virol. 2015, 90, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.N.; Yang, S.L.; Shih, S.R.; Huang, Y.C.; Chang, P.Y.; Huang, C.G.; Kao, K.-C.; Hu, H.-C.; Liu, Y.-C.; Tsao, K.-C. Molecular evolution and the global reemergence of enterovirus D68 by genome-wide analysis. Medicine (Baltim.) 2016, 95, e4416. [Google Scholar] [CrossRef] [PubMed]
- Yip, C.C.Y.; Lo, J.Y.C.; Sridhar, S.; Lung, D.C.; Luk, S.; Chan, K.-H.; Chan, J.F.W.; Cheng, V.C.C.; Woo, P.C.Y.; Yuen, K.-Y.; et al. First Report of a Fatal Case Associated with EV-D68 Infection in Hong Kong and Emergence of an Interclade Recombinant in China Revealed by Genome Analysis. Int. J. Mol. Sci. 2017, 18, 1065. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.; Yip, C.C.; Zhao, P.S.; Chow, W.N.; To, K.K.; Wu, A.K.; Yuen, K.-Y.; Woo, P.C.Y. Enterovirus D68 Infections Associated with Severe Respiratory Illness in Elderly Patients and Emergence of a Novel Clade in Hong Kong. Sci. Rep. 2016, 6, 25147. [Google Scholar] [CrossRef]
- Dyrdak, R.; Grabbe, M.; Hammas, B.; Ekwall, J.; Hansson, E.K.; Luthander, J.; Naucler, P.; Reinius, H.; Rotzén-Östlund, M.; Albert, J. Outbreak of enterovirus D68 of the new B3 lineage in Stockholm, Sweden, August to September 2016. Eurosurveillance 2016, 21, 30403. [Google Scholar] [CrossRef]
- Knoester, M.; Schölvinck, E.H.; Poelman, R.; Smit, S.; Vermont, C.L.; Niesters, H.G.; Van Leer-Buter, C.C. Upsurge of Enterovirus D68, the Netherlands, 2016. Emerg. Infect. Dis. 2017, 23, 140–143. [Google Scholar] [CrossRef]
- Esposito, S.; Chidini, G.; Cinnante, C.; Napolitano, L.; Giannini, A.; Terranova, L.; Niesters, H.G.M.; Principi, N.; Calderini, E. Acute flaccid myelitis associated with enterovirus-D68 infection in an otherwise healthy child. Virol. J. 2017, 14, 4. [Google Scholar] [CrossRef]
- Wang, G.; Zhuge, J.; Huang, W.; Nolan, S.M.; Gilrane, V.L.; Yin, C.; Dimitrova, N.; Fallon, J.T. Enterovirus D68 Subclade B3 Strain Circulating and Causing an Outbreak in the United States in 2016. Sci. Rep. 2017, 7, 1242. [Google Scholar] [CrossRef]
- Huang, W.; Wang, G.; Zhuge, J.; Nolan, S.M.; Dimitrova, N.; Fallon, J.T. Whole-Genome Sequence Analysis Reveals the Enterovirus D68 Isolates during the United States 2014 Outbreak Mainly Belong to a Novel Clade. Sci. Rep. 2015, 5, 15223. [Google Scholar] [CrossRef]
- Bitnun, A.; Yeh, E.A. Acute Flaccid Paralysis and Enteroviral Infections. Curr. Infect. Dis. Rep. 2018, 20, 34. [Google Scholar] [CrossRef] [PubMed]
- Midgley, C.; Jackson, M.A.; Selvarangan, R.; Turabelidze, G.; Obringer, E.; Johnson, D.; Giles, B.L.; Patel, A.; Echols, F.; Oberste, M.S.; et al. Severe Respiratory Illness Associated with Enterovirus D68—Missouri and Illinois, 2014. MMWR Morb. Mortal. Wkly. Rep. 2014, 63, 798–799. [Google Scholar] [PubMed]
- Harvala, H.; Broberg, E.; Benschop, K.; Berginc, N.; Ladhani, S.N.; Susi, P.; Christiansen, C.; McKenna, J.; Allen, D.; Makiello, P.; et al. Recommendations for enterovirus diagnostics and characterisation within and beyond Europe. J. Clin. Virol. 2018, 101, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Savolainen-Kopra, C.; Blomqvist, S. Mechanisms of genetic variation in polioviruses. Rev. Med. Virol. 2010, 20, 358–371. [Google Scholar] [CrossRef] [PubMed]
- Sanjuán, R. From Molecular Genetics to Phylodynamics: Evolutionary Relevance of Mutation Rates Across Viruses. PLoS Pathog. 2012, 8, e1002685. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Guo, W.; Wang, H.; Zhu, S.; Wang, D.; Bai, R.; Li, X.; Yan, D.; Wang, H.; et al. Emergence and Transmission Pathways of Rapidly Evolving Evolutionary Branch C4a Strains of Human Enterovirus 71 in the Central Plain of China. PLoS ONE 2011, 6, e27895. [Google Scholar] [CrossRef]
- Liu, W.; Wu, S.; Xiong, Y.; Li, T.; Wen, Z.; Yan, M.; Qin, K.; Liu, Y.; Wu, J. Co-Circulation and Genomic Recombination of Coxsackievirus A16 and Enterovirus 71 during a Large Outbreak of Hand, Foot, and Mouth Disease in Central China. PLoS ONE 2014, 9, e96051. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, Z.; Yang, W.; Ren, J.; Tan, X.; Wang, Y.; Naiying, M.; Xu, S.; Zhu, S.; Cui, A.; et al. An emerging recombinant human enterovirus 71 responsible for the 2008 outbreak of Hand Foot and Mouth Disease in Fuyang city of China. Virol. J. 2010, 7, 94. [Google Scholar] [CrossRef]
- Chen, S.-P.; Huang, Y.-C.; Li, W.-C.; Chiu, C.-H.; Huang, C.-G.; Tsao, K.-C.; Lin, T.-Y. Comparison of Clinical Features Between Coxsackievirus A2 and Enterovirus 71 During the Enterovirus Outbreak in Taiwan, 2008: A Children’s Hospital Experience. J. Microbiol. Immunol. Infect. 2010, 43, 99–104. [Google Scholar] [CrossRef]
- Lu, Q.-B.; Zhang, X.-A.; Wo, Y.; Xu, H.-M.; Li, X.-J.; Wang, X.-J.; Ding, S.-J.; Chen, X.-D.; He, C.; Liu, L.-J.; et al. Circulation of Coxsackievirus A10 and A6 in Hand-Foot-Mouth Disease in China, 2009–2011. PLoS ONE 2012, 7, e52073. [Google Scholar] [CrossRef]
- Leitch, E.C.M.; Bendig, J.; Cabrerizo, M.; Cardosa, J.; Hyypiä, T.; Ivanova, O.E.; Kelly, A.; Kroes, A.C.M.; Lukashev, A.; Macadam, A.; et al. Transmission Networks and Population Turnover of Echovirus 30. J. Virol. 2009, 83, 2109–2118. [Google Scholar] [CrossRef] [PubMed]
- Machado, R.S.; Ferreira, J.L.; Alves, J.C.S.; Bandeira, R.S.; Lemos, P.S.; Filho, L.C.F.; Cardoso, J.F.; Vianez-Junior, J.L.S.G.; Tavares, F.N. Nearly Complete Genome Sequences of Enterovirus 96 and Enterovirus 99 Strains Isolated in the Northern Region of Brazil. Microbiol. Resour. Announc. 2019, 8, e00549-19. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, X.; Cui, A.; Mao, N.; Xu, S.; Zhu, Z.; Zhou, J.; Shi, J.; Zhao, Y.; Wang, X.; et al. Complete Genome Analysis of the C4 Subgenotype Strains of Enterovirus 71: Predominant Recombination C4 Viruses Persistently Circulating in China for 14 Years. PLoS ONE 2013, 8, e56341. [Google Scholar] [CrossRef] [PubMed]
- Leitch, E.C.M.; Cabrerizo, M.; Cardosa, J.; Harvala, H.; Ivanova, O.E.; Kroes, A.C.M.; Lukashev, A.; Muir, P.; Odoom, J.; Roivainen, M.; et al. Evolutionary Dynamics and Temporal/Geographical Correlates of Recombination in the Human Enterovirus Echovirus Types 9, 11, and 30. J. Virol. 2010, 84, 9292–9300. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lukashev, A.N.; Shumilina, E.Y.; Belalov, I.S.; Ivanova, O.E.; Eremeeva, T.P.; Reznik, V.I.; Trotsenko, O.E.; Drexler, J.F.; Drosten, C. Recombination strategies and evolutionary dynamics of the Human enterovirus A global gene pool. J. Gen. Virol. 2014, 95, 868–873. [Google Scholar] [CrossRef]
- Egger, D.; Bienz, K. Recombination of Poliovirus RNA Proceeds in Mixed Replication Complexes Originating from Distinct Replication Start Sites. J. Virol. 2002, 76, 10960–10971. [Google Scholar] [CrossRef]
- Kirkegaard, K.; Baltimore, D. The mechanism of RNA recombination in poliovirus. Cell 1986, 47, 433–443. [Google Scholar] [CrossRef]
- Chetverin, A.B. The puzzle of RNA recombination. FEBS Lett. 1999, 460, 1–5. [Google Scholar] [CrossRef]
- Gmyl, A.P.; Belousov, E.V.; Maslova, S.V.; Khitrina, E.V.; Chetverin, A.B.; Agol, V.I. Nonreplicative RNA Recombination in Poliovirus. J. Virol. 1999, 73, 8958–8965. [Google Scholar] [CrossRef]
- Kyriakopoulou, Z.; Pliaka, V.; Amoutzias, G.D.; Markoulatos, P. Recombination among human non-polio enteroviruses: Implications for epidemiology and evolution. Virus Genes 2015, 50, 177–188. [Google Scholar] [CrossRef]
- Muslin, C.; Joffret, M.-L.; Pelletier, I.; Blondel, B.; Delpeyroux, F. Evolution and Emergence of Enteroviruses through Intra- and Inter-species Recombination: Plasticity and Phenotypic Impact of Modular Genetic Exchanges in the 5′ Untranslated Region. PLoS Pathog. 2015, 11, e1005266. [Google Scholar] [CrossRef] [PubMed]
- Yozwiak, N.L.; Skewes-Cox, P.; Gordon, A.; Saborio, S.; Kuan, G.; Balmaseda, A.; Ganem, D.; Harris, E.; DeRisi, J.L. Human Enterovirus 109: A Novel Interspecies Recombinant Enterovirus Isolated from a Case of Acute Pediatric Respiratory Illness in Nicaragua. J. Virol. 2010, 84, 9047–9058. [Google Scholar] [CrossRef] [PubMed]
- Santti, J.; Hyypia, T.; Kinnunen, L.; Salminen, M. Evidence of Recombination among Enteroviruses. J. Virol. 1999, 73, 8741–8749. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Yoke-Fun, C.; Abubakar, S. Phylogenetic evidence for inter-typic recombination in the emergence of human enterovirus 71 subgenotypes. BMC Microbiol. 2006, 6, 74. [Google Scholar] [CrossRef]
- Majumdar, M.; Sharif, S.; Klapsa, D.; Wilton, T.; Alam, M.M.; Fernandez-Garcia, M.D.; Rehman, L.; Mujtaba, G.; McAllister, G.; Harvala, H.; et al. Environmental Surveillance Reveals Complex Enterovirus Circulation Patterns in Human Populations. Open Forum Infect. Dis. 2018, 5, ofy250. [Google Scholar] [CrossRef]
- Pickett, B.E.; Sadat, E.L.; Zhang, Y.; Noronha, J.M.; Squires, R.B.; Hunt, V.; Liu, M.; Kumar, S.; Zaremba, S.; Gu, Z.; et al. ViPR: An open bioinformatics database and analysis resource for virology research. Nucleic Acids Res. 2012, 40, D593–D598. [Google Scholar] [CrossRef]
- Bo, Y.-C.; Song, C.; Wang, J.-F.; Li, X.-W. Using an autologistic regression model to identify spatial risk factors and spatial risk patterns of hand, foot and mouth disease (HFMD) in Mainland China. BMC Public Health 2014, 14, 358. [Google Scholar] [CrossRef]
- Khetsuriani, N.; Lamonte-Fowlkes, A.; Oberst, S.; Pallansch, M.A. Enterovirus surveillance—United States, 1970–2005. MMWR Surveil. Summ. 2006, 55, 1–20. [Google Scholar]
- Tee, K.K.; Lam, T.T.; Chan, Y.F.; Bible, J.M.; Kamarulzaman, A.; Tong, C.Y.; Takebe, Y.; Pybus, O.G. Evolutionary Genetics of Human Enterovirus 71: Origin, Population Dynamics, Natural Selection, and Seasonal Periodicity of the VP1 Gene. J. Virol. 2010, 84, 3339–3350. [Google Scholar] [CrossRef]
- Tokarz, R.; Firth, C.; Madhi, S.A.; Howie, S.R.; Wu, W.; Sall, A.A.; Haq, S.; Briese, T.; Lipkin, W.I. Worldwide emergence of multiple clades of enterovirus 68. J. Gen. Virol. 2012, 93, 1952–1958. [Google Scholar] [CrossRef] [PubMed]
- Yarmolskaya, M.S.; Shumilina, E.Y.; Ivanova, O.E.; Drexler, J.F.; Lukashev, A. Molecular epidemiology of echoviruses 11 and 30 in Russia: Different properties of genotypes within an enterovirus serotype. Infect. Genet. Evol. 2015, 30, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Oberste, M.S.; Maher, K.; Kennett, M.L.; Campbell, J.J.; Carpenter, M.S.; Schnurr, D.; Pallansch, M.A. Molecular Epidemiology and Genetic Diversity of Echovirus Type 30 (E30): Genotypes Correlate with Temporal Dynamics of E30 Isolation. J. Clin. Microbiol. 1999, 37, 3928–3933. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, C.; Hovi, T.; Mulders, M.N. Molecular epidemiology of echovirus 30 in Europe: Succession of dominant sublineages within a single major genotype. Arch. Virol. 2001, 146, 521–537. [Google Scholar] [CrossRef]
- Brown, B.A.; Nix, W.A.; Sheth, M.; Frace, M.; Oberste, M.S. Seven Strains of Enterovirus D68 Detected in the United States during the 2014 Severe Respiratory Disease Outbreak. Genome Announc. 2014, 2. [Google Scholar] [CrossRef]
- Poelman, R.; Schuffenecker, I.; Van Leer-Buter, C.; Josset, L.; Niesters, H.G.; Lina, B. European surveillance for enterovirus D68 during the emerging North-American outbreak in 2014. J. Clin. Virol. 2015, 71, 1–9. [Google Scholar] [CrossRef]
- Midgley, C.M.; Watson, J.T.; Nix, W.A.; Curns, A.T.; Rogers, S.L.; Brown, B.A.; Conover, C.; Dominguez, S.R.; Feikin, D.R.; Gray, S.; et al. Severe respiratory illness associated with a nationwide outbreak of enterovirus D68 in the USA (2014): A descriptive epidemiological investigation. Lancet Respir. Med. 2015, 3, 879–887. [Google Scholar] [CrossRef]
- Oh, M.-D.; Park, S.; Choi, Y.; Kim, H.; Lee, K.; Park, W.; Yoo, Y.; Kim, E.-C.; Choe, K. Acute Hemorrhagic Conjunctivitis Caused by Coxsackievirus A24 Variant, South Korea, 2002. Emerg. Infect. Dis. 2003, 9, 1010–1012. [Google Scholar] [CrossRef]
- Pons-Salort, M.; Oberste, M.S.; Pallansch, M.A.; Abedi, G.R.; Takahashi, S.; Grenfell, B.T.; Grassly, N.C. The seasonality of nonpolio enteroviruses in the United States: Patterns and drivers. Proc. Natl. Acad. Sci. USA 2018, 115, 3078–3083. [Google Scholar] [CrossRef]
- Yang, B.; Liu, F.; Liao, Q.; Wu, P.; Chang, Z.; Huang, J.; Long, L.; Luo, L.; Li, Y.; Leung, G.M.; et al. Epidemiology of hand, foot and mouth disease in China, 2008 to 2015 prior to the introduction of EV-A71 vaccine. Eurosurveillance 2017, 22, 16-00824. [Google Scholar] [CrossRef]
- Podin, Y.; Gias, E.L.; Ong, F.; Leong, Y.W.; Yee, S.F.; Yusof, M.A.; Perera, D.; Teo, B.; Wee, T.-Y.; Yao, S.-K.; et al. Sentinel surveillance for human enterovirus 71 in Sarawak, Malaysia: Lessons from the first 7 years. BMC Public Health 2006, 6, 180. [Google Scholar] [CrossRef] [PubMed]
- National Institute of Infectious Diseases. Hand, Foot and Mouth Disease in Japan, 2002–2011. 2012. Available online: https://www.niid.go.jp/niid/en/basic-science/865-iasr/1735-tpc385.html (accessed on 28 March 2019).
- Mladenova, Z.; Buttinelli, G.; Dikova, A.; Stoyanova, A.; Troyancheva, M.; Komitova, R.; Stoycheva, M.; Pekova, L.; Parmakova, K.; Fiore, L. Aseptic meningitis outbreak caused by echovirus 30 in two regions in Bulgaria, May–August 2012. Epidemiol. Infect. 2014, 142, 2159–2165. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kang, B.; Hwang, S.; Hong, J. Epidemics of viral meningitis caused by echovirus 6 and 30 in Korea in 2008. Virol. J. 2012, 9, 38. [Google Scholar] [CrossRef]
- Rudolph, H.; Dernbach, R.P.; Walka, M.; Rey-Hinterkopf, P.; Melichar, V.; Muschiol, E.; Schweitzer-Krantz, S.; Richter, J.W.; Weiss, C.; Böttcher, S.; et al. Comparison of clinical and laboratory characteristics during two major paediatric meningitis outbreaks of echovirus 30 and other non-polio enteroviruses in Germany in 2008 and 2013. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1651–1660. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, N.E.; Fout, G.S.; Keely, S.P. Retrospective Surveillance of Wastewater To Examine Seasonal Dynamics of Enterovirus Infections. mSphere 2017, 2, e00099-17. [Google Scholar] [CrossRef] [PubMed]
- Faleye, T.O.C.; Adewumi, M.O.; Japhet, M.O.; David, O.M.; Oluyege, A.O.; Adeniji, J.A.; Famurewa, O. Non-polio enteroviruses in faeces of children diagnosed with acute flaccid paralysis in Nigeria. Virol. J. 2017, 14, 175. [Google Scholar] [CrossRef]
- Zhang, Y.; Hong, M.; Sun, Q.; Zhu, S.; Tsewang; Li, X.; Yan, D.; Wang, D.; Xu, W. Molecular typing and characterization of a new serotype of human enterovirus (EV-B111) identified in China. Virus Res. 2014, 183, 75–80. [Google Scholar] [CrossRef]
- Adeniji, J.A.; Ayeni, F.A.; Ibrahim, A.; Tijani, K.A.; Faleye, T.O.C.; Adewumi, M.O. Comparison of Algorithms for the Detection of Enteroviruses in Stool Specimens from Children Diagnosed with Acute Flaccid Paralysis. J. Pathog. 2017, 2017, 9256056. [Google Scholar] [CrossRef]
- Horner, L.M.; Poulter, M.D.; Brenton, J.N.; Turner, R.B. Acute Flaccid Paralysis Associated with Novel Enterovirus C105. Emerg. Infect. Dis. 2015, 21, 1858–1860. [Google Scholar] [CrossRef]
- Maan, H.S.; Dhole, T.N.; Chowdhary, R. Identification and characterization of nonpolio enterovirus associated with nonpolio-acute flaccid paralysis in polio endemic state of Uttar Pradesh, Northern India. PLoS ONE 2019, 14, e0208902. [Google Scholar] [CrossRef] [PubMed]
- Barnadas, C.; Midgley, S.E.; Skov, M.N.; Jensen, L.; Poulsen, M.W.; Fischer, T.K. An enhanced Enterovirus surveillance system allows identification and characterization of rare and emerging respiratory enteroviruses in Denmark, 2015–2016. J. Clin. Virol. 2017, 93, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Donbraye, E.; Olasunkanmi, O.I.; Opabode, B.A.; Ishola, T.R.; Faleye, T.O.C.; Adewumi, O.M.; Adeniji, J.A. Abundance of enterovirus C in RD-L20B cell culture-negative stool samples from acute flaccid paralysis cases in Nigeria is geographically defined. J. Med. Microbiol. 2018, 67, 854–865. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Sequences * | |||
|---|---|---|---|
| All Time | 2014 to 2019 | Species | Type |
| 7174 | 2890 | A | Coxsackievirus A6 |
| 12720 | 2413 | A | Enterovirus A71 |
| 5318 | 1627 | A | Coxsackievirus A16 |
| 2068 | 742 | A | Coxsackievirus A10 |
| 974 | 435 | A | Coxsackievirus A4 |
| 418 | 139 | A | Coxsackievirus A2 |
| 275 | 111 | A | Coxsackievirus A5 |
| 2211 | 653 | B | Echovirus 11 |
| 1850 | 612 | B | Coxsackievirus B5 |
| 4402 | 516 | B | Echovirus 30 |
| 2139 | 497 | B | Echovirus 6 |
| 569 | 211 | B | Echovirus 18 |
| 1244 | 194 | B | Coxsackievirus B3 |
| 551 | 178 | B | Echovirus 25 |
| 238 | 166 | B | Echovirus 16 |
| 1100 | 129 | B | Coxsackievirus B4 |
| 803 | 106 | B | Echovirus 7 |
| 778 | 97 | B | Echovirus 9 |
| 430 | 90 | B | Echovirus 3 |
| 549 | 75 | B | Coxsackievirus B2 |
| 539 | 69 | B | Coxsackievirus B1 |
| 620 | 68 | B | Coxsackievirus A9 |
| 105 | 2 | B | Coxsackievirus B6 |
| 974 | 157 | C | Coxsackievirus A24 |
| 258 | 33 | C | Coxsackievirus A13 |
| 154 | 27 | C | Coxsackievirus A21 |
| 133 | 27 | C | Enterovirus C99 |
| 3841 | 2895 | D | Enterovirus D68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brown, D.M.; Zhang, Y.; Scheuermann, R.H. Epidemiology and Sequence-Based Evolutionary Analysis of Circulating Non-Polio Enteroviruses. Microorganisms 2020, 8, 1856. https://doi.org/10.3390/microorganisms8121856
Brown DM, Zhang Y, Scheuermann RH. Epidemiology and Sequence-Based Evolutionary Analysis of Circulating Non-Polio Enteroviruses. Microorganisms. 2020; 8(12):1856. https://doi.org/10.3390/microorganisms8121856
Chicago/Turabian StyleBrown, David M., Yun Zhang, and Richard H. Scheuermann. 2020. "Epidemiology and Sequence-Based Evolutionary Analysis of Circulating Non-Polio Enteroviruses" Microorganisms 8, no. 12: 1856. https://doi.org/10.3390/microorganisms8121856
APA StyleBrown, D. M., Zhang, Y., & Scheuermann, R. H. (2020). Epidemiology and Sequence-Based Evolutionary Analysis of Circulating Non-Polio Enteroviruses. Microorganisms, 8(12), 1856. https://doi.org/10.3390/microorganisms8121856