A Reporter System for Fast Quantitative Monitoring of Type 3 Protein Secretion in Enteropathogenic E. coli

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Generation of the sctA-phoA Construct
2.2. Optimization of Media
2.3. Bacterial Culture for In Vivo Secretion Assay
2.4. In Vivo Alkaline Phosphatase Assay
2.4.1. Treatment of the Spent Growth Medium Samples
2.4.2. Treatment of the Cell Samples
3. Results
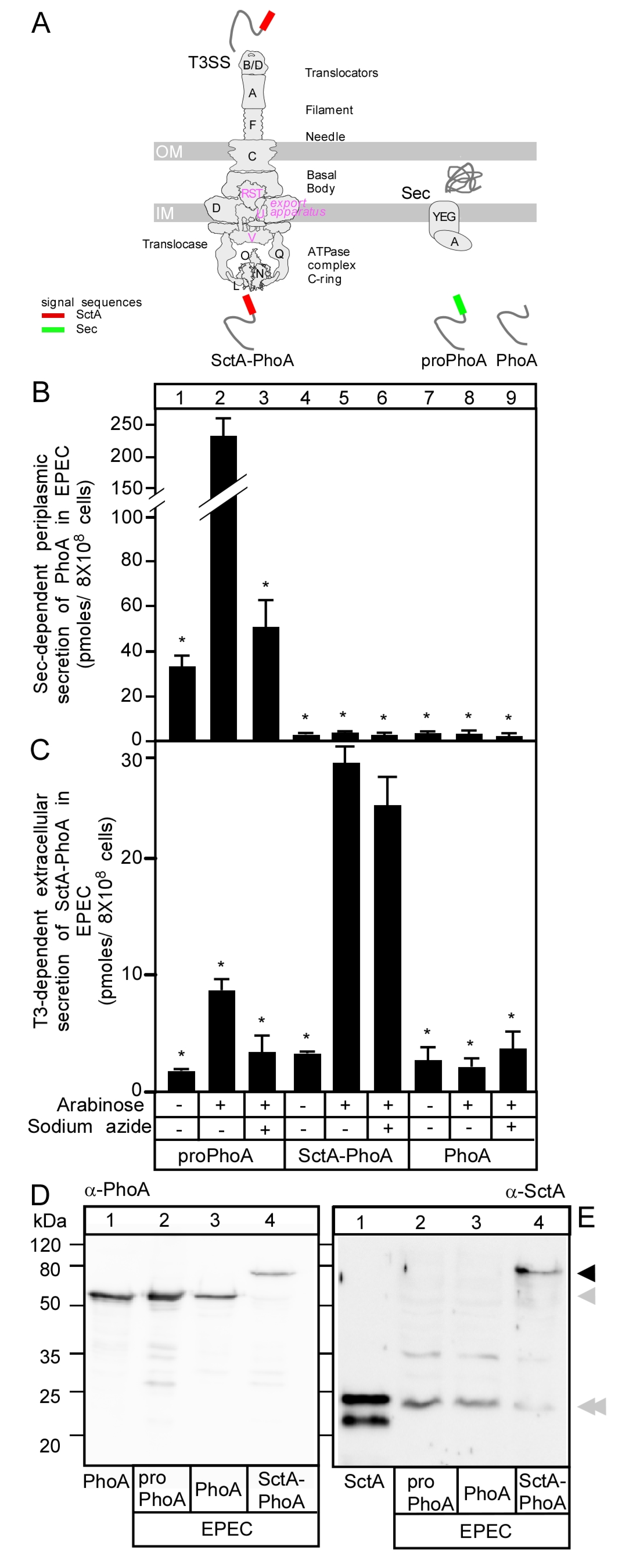
3.1. Secretion of SctA-PhoA Is T3SS-Dependent
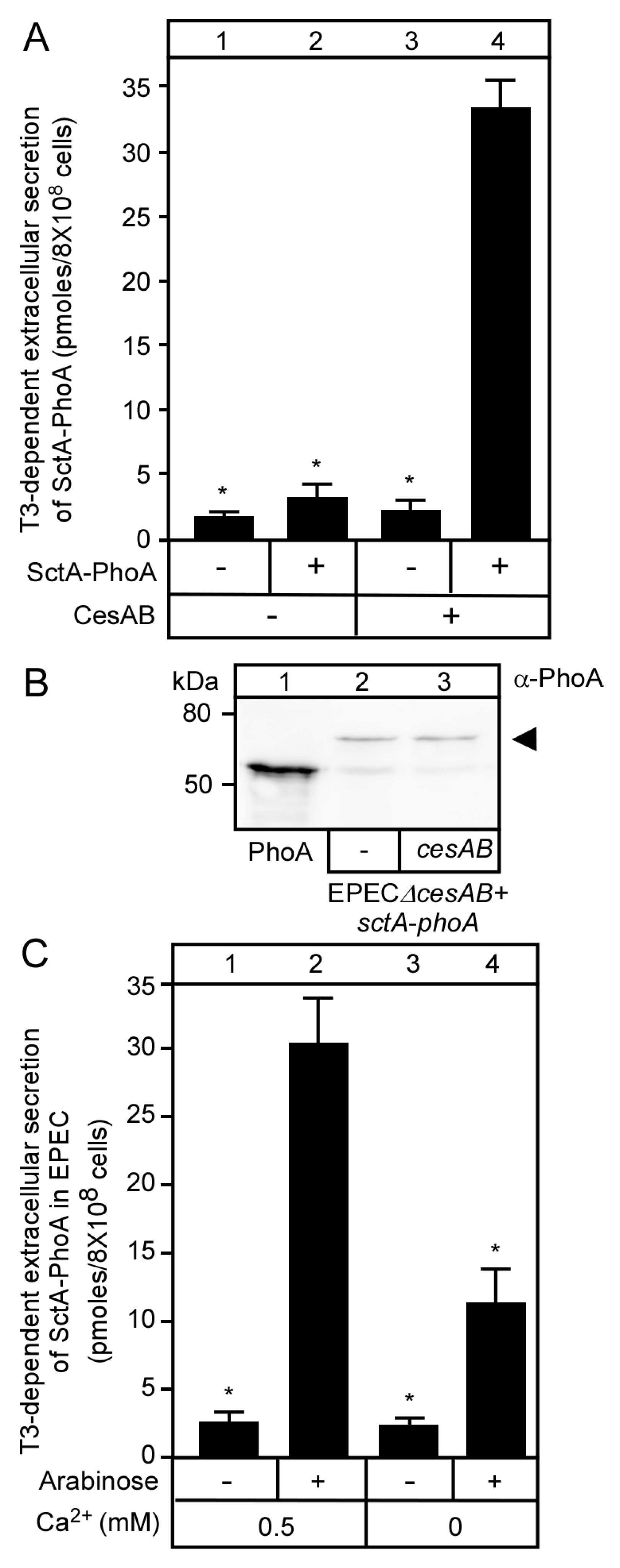
3.2. T3S-Dependent Secretion of SctA-PhoA and Native SctA Have the Same Requirements
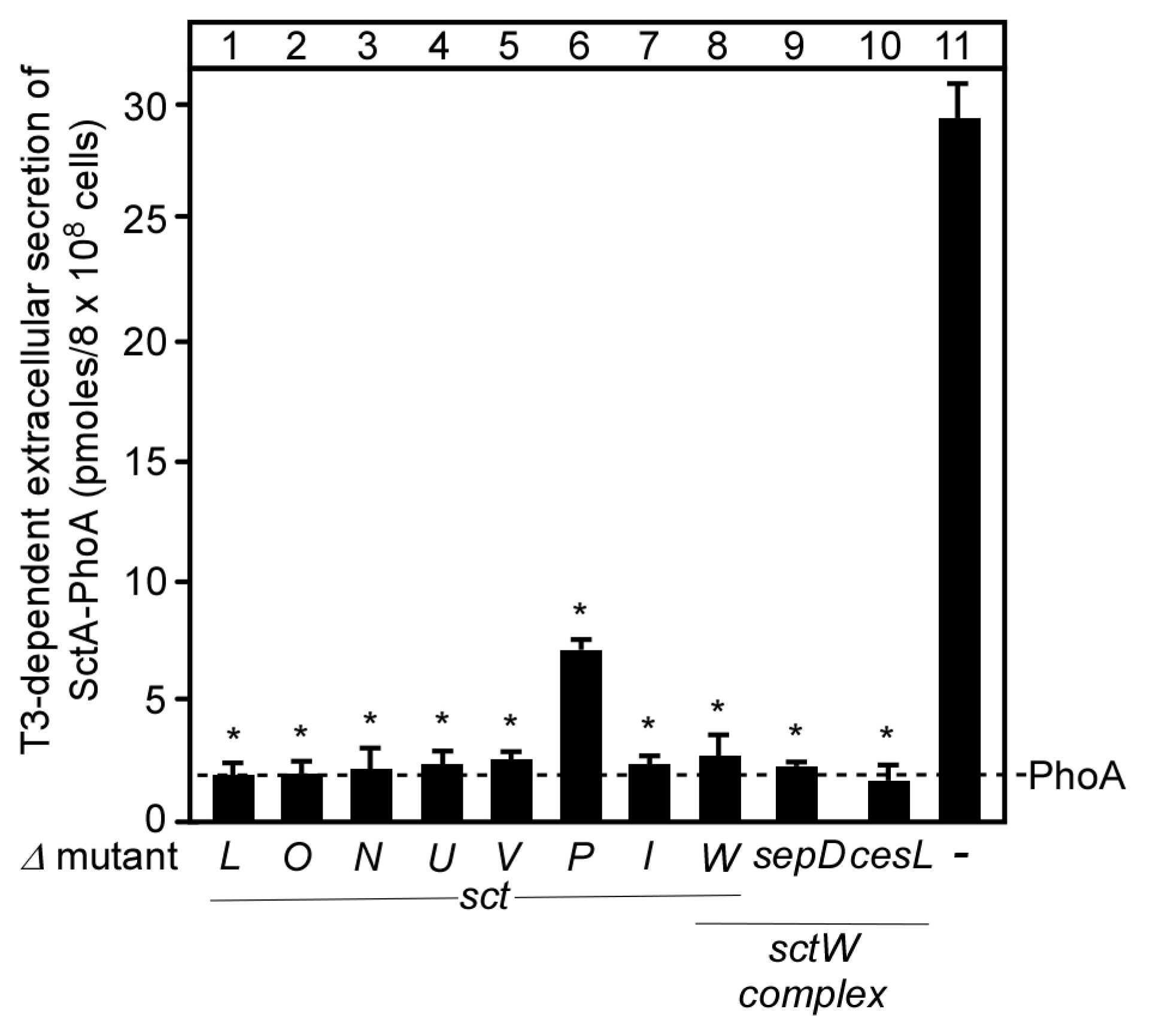
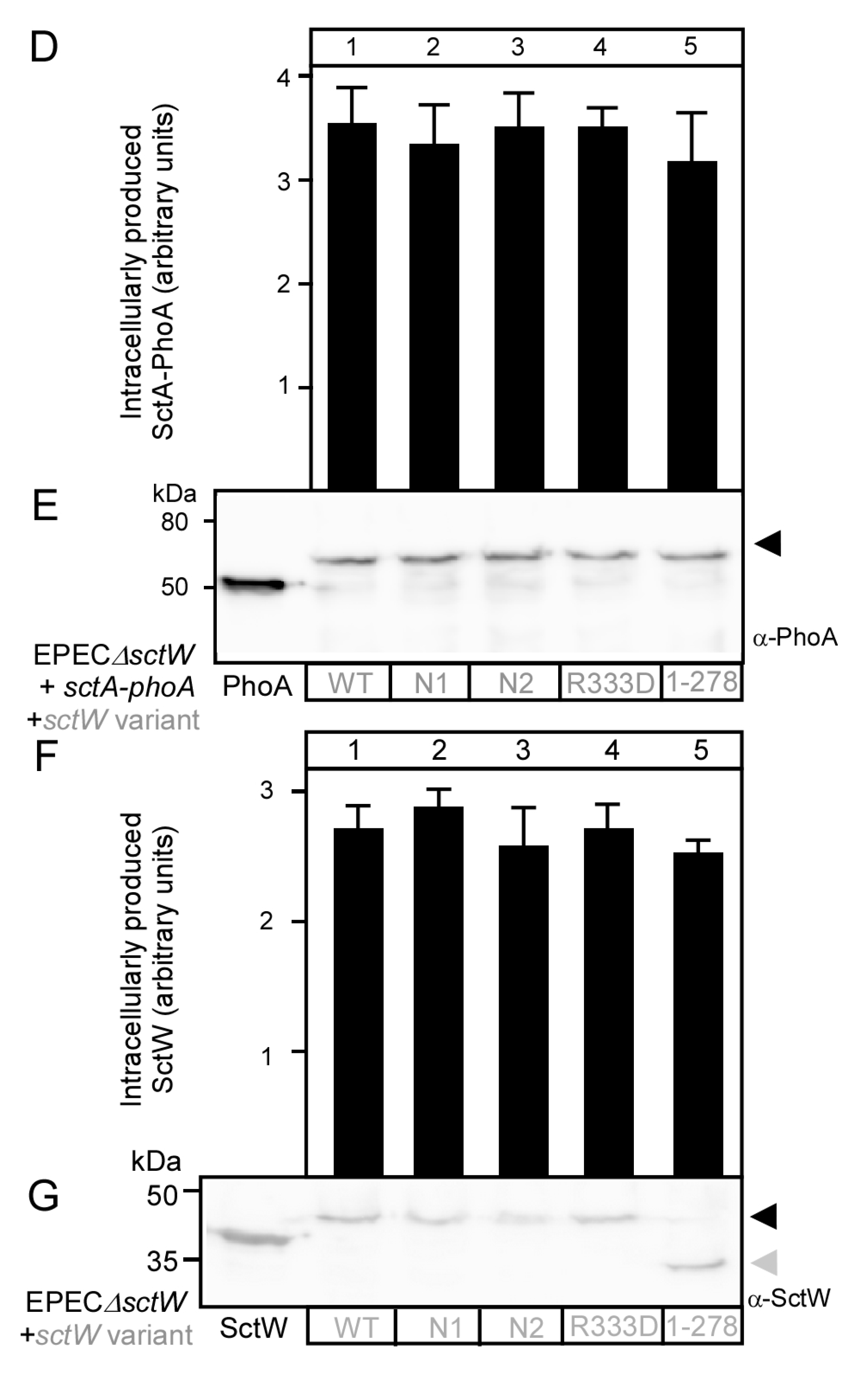
3.3. Functional Characterization of SctW Mutants Using the SctA-PhoA Reporter
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Deng, W.; Marshall, N.C.; Rowland, J.L.; McCoy, J.M.; Worrall, L.J.; Santos, A.S.; Strynadka, N.C.J.; Finlay, B.B. Assembly, structure, function and regulation of type III secretion systems. Nat. Rev. Microbiol. 2017, 15, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Portaliou, A.G.; Tsolis, K.C.; Loos, M.S.; Zorzini, V.; Economou, A. Type III Secretion: Building and Operating a Remarkable Nanomachine. Trends Biochem. Sci. 2016, 41, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Li, Y.; Hardwidge, P.R.; Frey, E.A.; Pfuetzner, R.A.; Lee, S.; Gruenheid, S.; Strynakda, N.C.; Puente, J.L.; Finlay, B.B. Regulation of Type Iii Secretion Hierarchy of Translocators and Effectors in Attaching and Effacing Bacterial Pathogens. Infect. Immun. 2005, 73, 2135–2146. [Google Scholar] [CrossRef] [PubMed]
- Burkinshaw, B.J.; Strynadka, N.C.J. Assembly and structure of the T3SS. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 1649–1663. [Google Scholar] [CrossRef]
- Biemans-Oldehinkel, E.; Sal-Man, N.; Deng, W.; Foster, L.J.; Finlay, B. Quantitative Proteomic Analysis Reveals Formation of an EscL-EscQ-EscN Type III Complex in Enteropathogenic Escherichia coli. J. Bacteriol. 2011, 193, 5514–5519. [Google Scholar] [CrossRef]
- Radics, J.; Königsmaier, L.; Marlovits, T.C. Structure of a pathogenic type 3 secretion system in action. Nat. Struct. Mol. Biol. 2014, 21, 82–87. [Google Scholar] [CrossRef]
- Schraidt, O.; Marlovits, T.C. Three-Dimensional Model of Salmonella’s Needle Complex at Subnanometer Resolution. Science 2011, 331, 1192–1195. [Google Scholar] [CrossRef]
- Diepold, A.; Wagner, S. Assembly of the bacterial type III secretion machinery. FEMS Microbiol. Rev. 2014, 38, 802–822. [Google Scholar] [CrossRef]
- Portaliou, A.G.; Tsolis, K.C.; Loos, M.S.; Balabanidou, V.; Rayo, J.; Tsirigotaki, A.; Crepin, V.F.; Frankel, G.; Kalodimos, C.G.; Karamanou, S.; et al. Hierarchical protein targeting and secretion is controlled by an affinity switch in the type III secretion system of enteropathogenic Escherichia coli. EMBO J. 2017, 36, 3517–3531. [Google Scholar] [CrossRef]
- Tsirigotaki, A.; De Geyter, J.; Šoštarić, N.; Economou, A.; Karamanou, S. Protein export through the bacterial Sec pathway. Nat. Rev. Microbiol. 2017, 15, 21–36. [Google Scholar] [CrossRef]
- Orfanoudaki, G.; Economou, A. Proteome-wide subcellular topologies of E. coli polypeptides database (STEPdb). Mol. Cell. Proteom. 2014, 13, 3674–3687. [Google Scholar] [CrossRef] [PubMed]
- Sukhan, A.; Kubori, T.; Wilson, J.; Galán, J.E. Genetic Analysis of Assembly of the Salmonella enterica Serovar Typhimurium Type III Secretion-Associated Needle Complex. J. Bacteriol. 2001, 183, 1159–1167. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, A.; Puente, J.L.; Finlay, B.B. Secretin of the Enteropathogenic Escherichia Coli Type Iii Secretion System Requires Components of the Type Iii Apparatus for Assembly and Localization. Infect. Immun. 2003, 71, 3310–3319. [Google Scholar] [CrossRef] [PubMed]
- Takaya, A.; Takeda, H.; Tashiro, S.; Kawashima, H.; Yamamoto, T. Chaperone-mediated secretion switching from early to middle substrates in the type III secretion system encoded by Salmonella pathogenicity island 2. J. Biol. Chem. 2019, 294, 3783–3793. [Google Scholar] [CrossRef]
- Gouridis, G.; Karamanou, S.; Koukaki, M.; Economou, A. In Vitro Assays to Analyze Translocation of the Model Secretory Preprotein Alkaline Phosphatase. Methods Mol. Biol. 2010, 619, 157–172. [Google Scholar]
- Knutton, S.; Rosenshine, I.; Pallen, M.J.; Nisan, I.; Neves, B.C.; Bain, C.; Wolff, C.; Dougan, G.; Frankel, G. A Novel Espa-Associated Surface Organelle of Enteropathogenic Escherichia Coli Involved in Protein Translocation into Epithelial Cells. EMBO J. 1998, 17, 2166–2176. [Google Scholar] [CrossRef]
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142. [Google Scholar] [CrossRef]
- Bray, J. Isolation of antigenically homogeneous strains of Bact. coli neapolitanum from summer diarrhœa of infants. J. Pathol. Bacteriol. 1945, 57, 239–247. [Google Scholar] [CrossRef]
- Taylor, K.A.; O’Connell, C.B.; Luther, P.W.; Donnenberg, M.S. The EspB Protein of Enteropathogenic Escherichia coli Is Targeted to the Cytoplasm of Infected HeLa Cells. Infect. Immun. 1998, 66, 5501–5507. [Google Scholar] [CrossRef]
- Wagner, S.; Grin, I.; Malmsheimer, S.; Singh, N.; Torres-Vargas, C.E.; Westerhausen, S. Bacterial type III secretion systems: A complex device for the delivery of bacterial effector proteins into eukaryotic host cells. FEMS Microbiol. Lett. 2018, 365, 1–13. [Google Scholar] [CrossRef]
- Shaulov, L.; Gershberg, J.; Deng, W.; Finlay, B.B.; Sal-Man, N. The Ruler Protein EscP of the Enteropathogenic Escherichia coli Type III Secretion System Is Involved in Calcium Sensing and Secretion Hierarchy Regulation by Interacting with the Gatekeeper Protein SepL. MBio 2017, 8, e01733-16. [Google Scholar] [CrossRef] [PubMed]
- Pendergrass, H.A.; May, A.E. Delivery of heterologous proteins, enzymes, and antigens via the bacterial type III secretion system. Microorganisms 2020, 8, 777. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, K.J.; Bevington, J.L.; Rosales, S.L.; Burdette, L.A.; Valdivia, E.; Tullman-Ercek, D. Proteins adopt functionally active conformations after type III secretion. Microb. Cell Factories 2016, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sory, M.P.; Cornelis, G.R. Translocation of a hybrid YopE-adenylate cyclase from Yersinia enterocolitica into HeLa cells. Mol. Microbiol. 1994, 14, 583–594. [Google Scholar] [CrossRef]
- Wolff, C.; Nisan, I.; Hanski, E.; Frankel, G.; Rosenshine, I. Protein translocation into host epithelial cells by infecting enteropathogenic Escherichia coli. Mol. Microbiol. 1998, 28, 143–155. [Google Scholar] [CrossRef]
- Crawford, J.A.; Kaper, J.B. The N-terminus of enteropathogenic Escherichia coli (EPEC) Tir mediates transport across bacterial and eukaryotic cell membranes. Mol. Microbiol. 2002, 46, 855–868. [Google Scholar] [CrossRef]
- Button, J.E.; Galán, J.E. Regulation of chaperone/effector complex synthesis in a bacterial type III secretion system. Mol. Microbiol. 2011, 81, 1474–1483. [Google Scholar] [CrossRef]
- Pan, N.J.; Brady, M.J.; Leong, J.M.; Goguen, J.D. Targeting Type III Secretion in Yersinia pestis. Antimicrob. Agents Chemother. 2009, 53, 385–392. [Google Scholar] [CrossRef]
- Schesser, K.; Frithz-Lindsten, E.; Wolf-Watz, H. Delineation and mutational analysis of the Yersinia pseudotuberculosis YopE domains which mediate translocation across bacterial and eukaryotic cellular membranes. J. Bacteriol. 1996, 178, 7227–7233. [Google Scholar] [CrossRef]
- Felise, H.B.; Nguyen, H.V.; Pfuetzner, R.A.; Barry, K.C.; Jackson, S.R.; Blanc, M.-P.; Bronstein, P.A.; Kline, T.; Miller, S.I. An Inhibitor of Gram-Negative Bacterial Virulence Protein Secretion. Cell Host Microbe 2008, 4, 325–336. [Google Scholar] [CrossRef]
- Lindner, F.; Milne-Davies, B.; Langenfeld, K.; Stiewe, T.; Diepold, A. LITESEC-T3SS—Light-controlled protein delivery into eukaryotic cells with high spatial and temporal resolution. Nat. Commun. 2020, 11, 2381. [Google Scholar] [CrossRef] [PubMed]
- Munera, D.; Crepin, V.F.; Marches, O.; Frankel, G. N-Terminal Type III Secretion Signal of Enteropathogenic Escherichia coli Translocator Proteins. J. Bacteriol. 2010, 192, 3534–3539. [Google Scholar] [CrossRef] [PubMed]
- Busche, T.; Tsolis, K.C.; Koepff, J.; Rebets, Y.; Rückert, C.; Hamed, M.B.; Bleidt, A.; Wiechert, W.; Lopatniuk, M.; Yousra, A.; et al. Multi-Omics and Targeted Approaches to Determine the Role of Cellular Proteases in Streptomyces Protein Secretion. Front. Microbiol. 2018, 9, 1174. [Google Scholar] [CrossRef] [PubMed]
- Diepold, A.; Wiesand, U.; Amstutz, M.; Cornelis, G.R. Assembly of the Yersinia injectisome: The missing pieces. Mol. Microbiol. 2012, 85, 878–892. [Google Scholar] [CrossRef] [PubMed]
- Sone, M.; Kishigami, S.; Yoshihisa, T.; Ito, K. Roles of Disulfide Bonds in Bacterial Alkaline Phosphatase. J. Biol. Chem. 1997, 272, 6174–6178. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.; Kong, A.; Lam, J.; Young, L. Periplasmic Alkaline Phosphatase Activity and Abundance in Escherichia coli B23 and C29 during Exponential and Stationary Phase. J. Exp. Microbiol. Immunol. 2007, 11, 8–13. [Google Scholar]
- Chiu, H.J.; Lin, W.S.; Syu, W., Jr. Type III secretion of EspB in enterohemorrhagic Escherichia coli O157:H7. Arch. Microbiol. 2003, 180, 218–226. [Google Scholar] [CrossRef]
- Mills, E.; Baruch, K.; Charpentier, X.; Kobi, S.; Rosenshine, I. Real-Time Analysis of Effector Translocation by the Type III Secretion System of Enteropathogenic Escherichia coli. Cell Host Microbe 2008, 3, 104–113. [Google Scholar] [CrossRef]
- Yip, C.K.; Finlay, B.B.; Strynadka, N.C. Structural characterization of a type III secretion system filament protein in complex with its chaperone. Nat. Struct. Mol. Biol. 2005, 12, 75–81. [Google Scholar] [CrossRef]
- Crepin, V.F.; Martinez, E.; Shaw, R.K.; Frankel, G.; Daniell, S.J. Structural and Functional Properties of Chimeric EspA-FliCi Filaments of EPEC. J. Mol. Biol. 2008, 378, 243–250. [Google Scholar] [CrossRef]
- Michiels, T.; Cornelis, G.R. Secretion of hybrid proteins by the Yersinia Yop export system. J. Bacteriol. 1991, 173, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Chatzi, K.E.; Sardis, M.F.; Tsirigotaki, A.; Koukaki, M.; Šoštarić, N.; Konijnenberg, A.; Sobott, F.; Kalodimos, C.G.; Karamanou, S.; Economou, A. Preprotein mature domains contain translocase targeting signals that are essential for secretion. J. Cell Biol. 2017, 216, 1357–1369. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Economou, A.; Karamanou, S. Optimization of type 3 protein secretion in enteropathogenic Escherichia coli. FEMS Microbiol. Lett. 2018, 365, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Siegele, D.A.; Hu, J.C. Gene expression from plasmids containing the araBAD promoter at subsaturating inducer concentrations represents mixed populations. Proc. Natl. Acad. Sci. USA 1997, 94, 816–8172. [Google Scholar] [CrossRef] [PubMed]
- Mandel, M.; Higa, A. Calcium-dependent bacteriophage DNA infection. J. Mol. Biol. 1970, 53, 159–162. [Google Scholar] [CrossRef]
- Lombardi, C.; Tolchard, J.; Bouillot, S.; Signor, L.; Gebus, C.; Liebl, D.; Fenel, D.; Teulon, J.-M.; Brock, J.; Habenstein, B.; et al. Structural and Functional Characterization of the Type Three Secretion System (T3SS) Needle of Pseudomonas aeruginosa. Front. Microbiol. 2019, 10, 573. [Google Scholar] [CrossRef]
- Creasey, E.A.; Friedberg, D.; Shaw, R.K.; Umanski, T.; Knutton, S.; Rosenshine, I.; Frankel, G. CesAB is an enteropathogenic Escherichia coli chaperone for the type-III translocator proteins EspA and EspB. Microbiology 2003, 149, 3639–3647. [Google Scholar] [CrossRef]
- Burkinshaw, B.J.; Souza, S.A.; Strynadka, N.C.J. Structural analysis of SepL, an enteropathogenic Escherichia coli type III secretion-system gatekeeper protein. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2015, 71, 1300–1308. [Google Scholar] [CrossRef]
- Rohl, C.A.; Fiori, W.; Baldwin, R.L. Alanine is helix-stabilizing in both template-nucleated and standard peptide helices. Proc. Natl. Acad. Sci. USA 1999, 96, 3682–3687. [Google Scholar] [CrossRef]
- Guzman, L.M.; Belin, D.; Carson, M.J.; Beckwith, J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 1995, 177, 4121–4130. [Google Scholar] [CrossRef]
- O’Connell, C.B.; Creasey, E.A.; Knutton, S.; Elliott, S.; Crowther, L.J.; Luo, W.; Albert, M.J.; Kaper, J.B.; Frankel, G.; Donnenberg, M.S. SepL, a protein required for enteropathogenic Escherichia coli type III translocation, interacts with secretion component SepD. Mol. Microbiol. 2004, 52, 1613–1625. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Vallance, B.A.; Li, Y.; Puente, J.L.; Finlay, B.B. Citrobacter rodentium translocated intimin receptor (Tir) is an essential virulence factor needed for actin condensation, intestinal colonization and colonic hyperplasia in mice. Mol. Microbiol. 2003, 48, 95–115. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Puente, J.L.; Gruenheid, S.; Li, Y.; Vallance, B.A.; Vázquez, A.; Barba, J.; Ibarra, J.A.; O’Donnell, P.; Metalnikov, P.; et al. Dissecting virulence: Systematic and functional analyses of a pathogenicity island. Proc. Natl. Acad. Sci. USA 2004, 101, 3597–3602. [Google Scholar] [CrossRef] [PubMed]
- Kenny, B.; Lai, L.-C.; Finlay, B.B.; Donnenberg, M.S. EspA, a protein secreted by enteropathogenic Escherichia coli, is required to induce signals in epithelial cells. Mol. Microbiol. 1996, 20, 313–323. [Google Scholar] [CrossRef]
- Abe, A.; Kenny, B.; Stein, M.; Finlay, B.B. Characterization of two virulence proteins secreted by rabbit enteropathogenic Escherichia coli, EspA and EspB, whose maximal expression is sensitive to host body temperature. Infect. Immun. 1997, 65, 3547–3555. [Google Scholar] [CrossRef]
- Shaw, R.K.; Daniell, S.; Ebel, F.; Frankel, G.; Knutton, S. EspA filament-mediated protein translocation into red blood cells. Cell Microbiol. 2001, 3, 213–222. [Google Scholar] [CrossRef]
- Chen, L.; Balabanidou, V.; Remeta, D.P.; Minetti, C.A.; Portaliou, A.G.; Economou, A.; Kalodimos, C.G. Structural Instability Tuning as a Regulatory Mechanism in Protein-Protein Interactions. Mol. Cell 2011, 44, 734–744. [Google Scholar] [CrossRef]
- Sardis, M.F.; Tsirigotaki, A.; Chatzi, K.E.; Portaliou, A.G.; Gouridis, G.; Karamanou, S.; Economou, A. Preprotein Conformational Dynamics Drive Bivalent Translocase Docking and Secretion. Structure 2017, 25, 1056–1067. [Google Scholar] [CrossRef]
- Rüssmann, H.; Shams, H.; Poblete, F.; Fu, Y.; Galán, J.E.; Donis, R.O. Delivery of Epitopes by the Salmonella Type III Secretion System for Vaccine Development. Science 1998, 281, 565. [Google Scholar]
- Gaytán, M.O.; Feria, J.M.; Soto, E.; Espinosa, N.; Benítez, J.M.; Georgellis, D.; González-Pedrajo, B. Novel insights into the mechanism of SepL-mediated control of effector secretion in enteropathogenic Escherichia coli. MicrobiologyOpen 2018, 7, 1–17. [Google Scholar] [CrossRef]
- Manoil, C.; Mekalanos, J.J.; Beckwith, J. Alkaline phosphatase fusions: Sensors of subcellular location. J. Bacteriol. 1990, 172, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Hamed, M.B.; Burchacka, E.; Angus, L.; Marchand, A.; De Geyter, J.; Loos, M.S.; Anné, J.; Klaassen, H.; Chaltin, P.; Karamanou, S.; et al. Small Molecule Antibiotic Inhibitors or Post-Translational Protein Secretion. Front. Microbiol. 2020. submitted. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barkalita, L.; Portaliou, A.G.; Loos, M.S.; Yuan, B.; Karamanou, S.; Economou, A. A Reporter System for Fast Quantitative Monitoring of Type 3 Protein Secretion in Enteropathogenic E. coli. Microorganisms 2020, 8, 1786. https://doi.org/10.3390/microorganisms8111786
Barkalita L, Portaliou AG, Loos MS, Yuan B, Karamanou S, Economou A. A Reporter System for Fast Quantitative Monitoring of Type 3 Protein Secretion in Enteropathogenic E. coli. Microorganisms. 2020; 8(11):1786. https://doi.org/10.3390/microorganisms8111786
Chicago/Turabian StyleBarkalita, Luit, Athina G. Portaliou, Maria S. Loos, Biao Yuan, Spyridoula Karamanou, and Anastassios Economou. 2020. "A Reporter System for Fast Quantitative Monitoring of Type 3 Protein Secretion in Enteropathogenic E. coli" Microorganisms 8, no. 11: 1786. https://doi.org/10.3390/microorganisms8111786
APA StyleBarkalita, L., Portaliou, A. G., Loos, M. S., Yuan, B., Karamanou, S., & Economou, A. (2020). A Reporter System for Fast Quantitative Monitoring of Type 3 Protein Secretion in Enteropathogenic E. coli. Microorganisms, 8(11), 1786. https://doi.org/10.3390/microorganisms8111786