Cut-Lengths of Perennial Ryegrass Leaf-Blades Influences In Vitro Fermentation by the Anaerobic Fungus Neocallimastix frontalis

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Maintenance and Pre-Growth of Neocallimastix frontalis Cultures
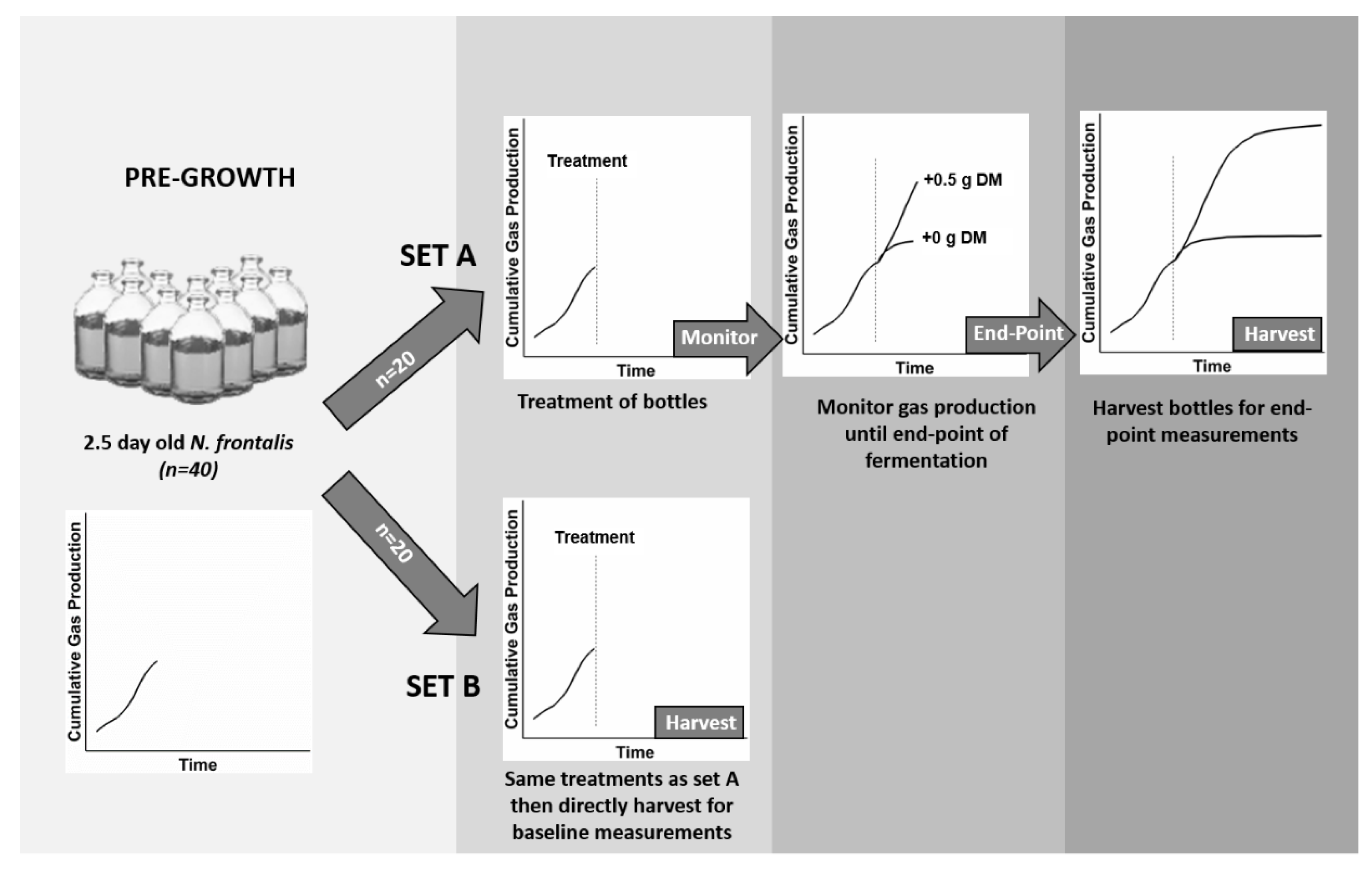
2.3. Modified In Vitro Gas Production Technique
2.4. In Vitro Fermentation of Different Cut-Lengths of Perennial Ryegrass Leaf-Blades by Cultures of N. frontalis Pre-Grown on Barley Straw
2.5. Determination of Perennial Ryegrass Fermentation End-Products, Initial Dry Matter Loss, and Apparent Dry Matter Loss
2.6. Statistical Analysis
3. Results
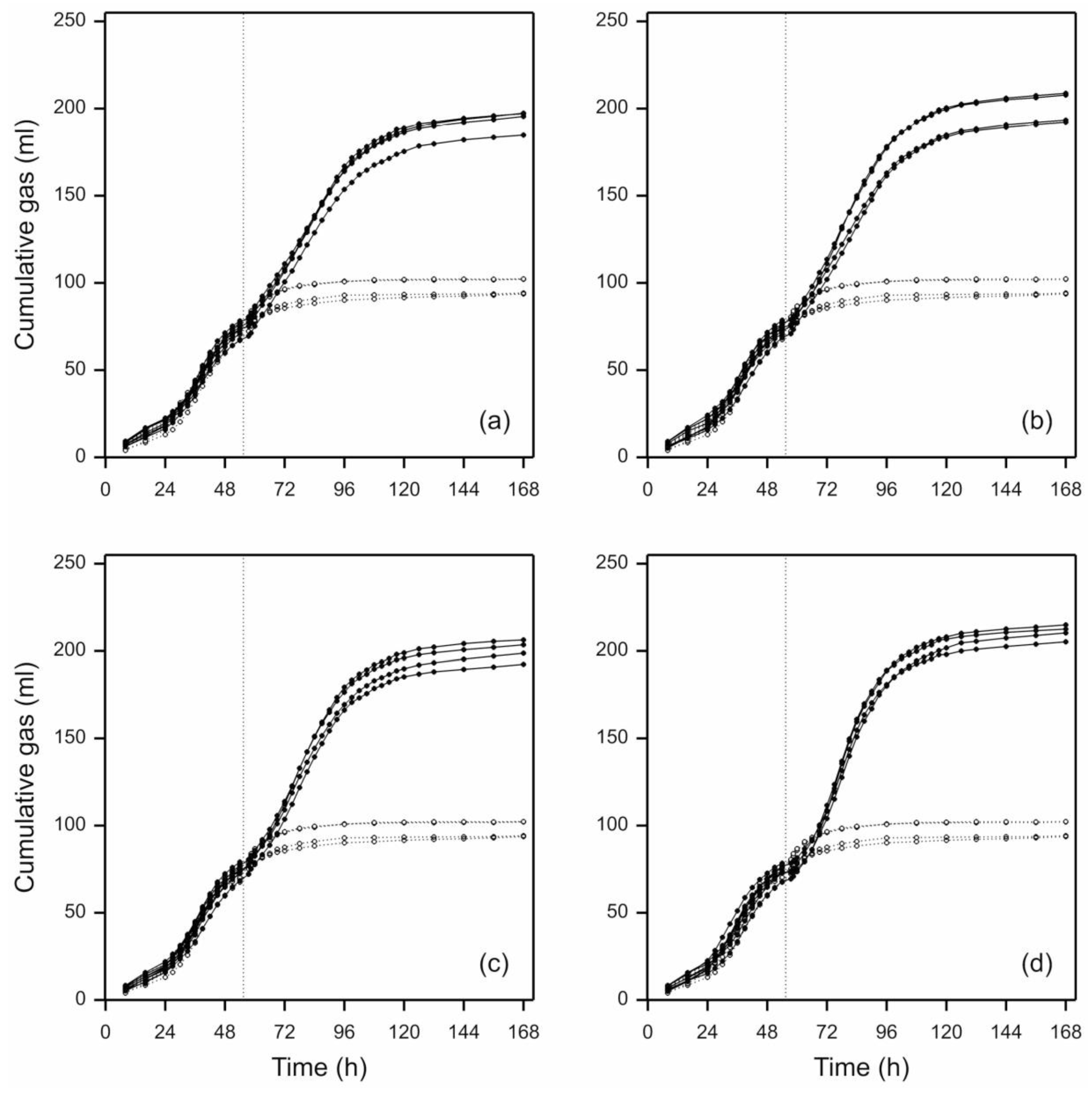
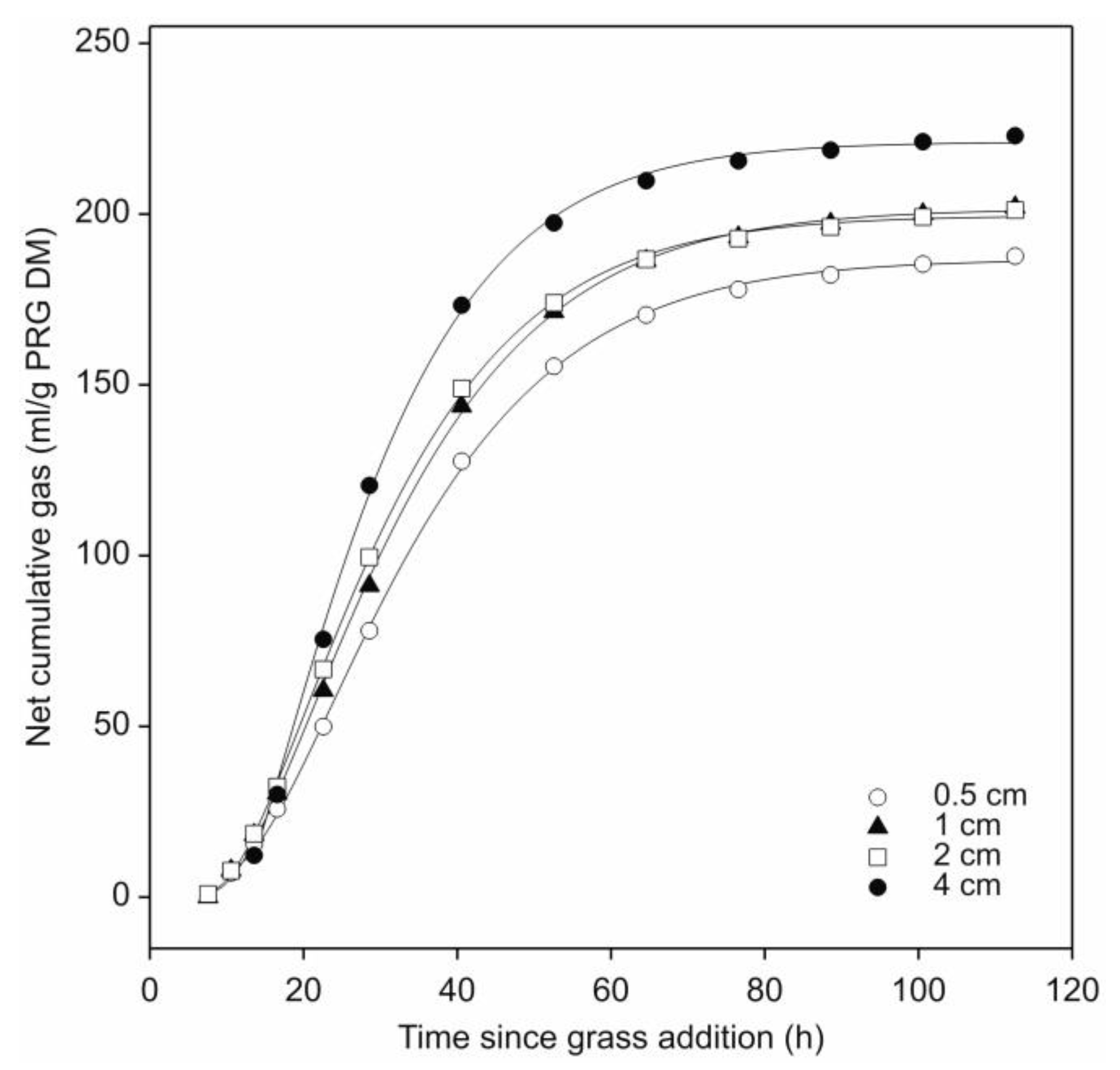
3.1. Effect of Cut-Length of Fresh Perennial Ryegrass Leaf-Blades on Their Fermentation by N. frontalis
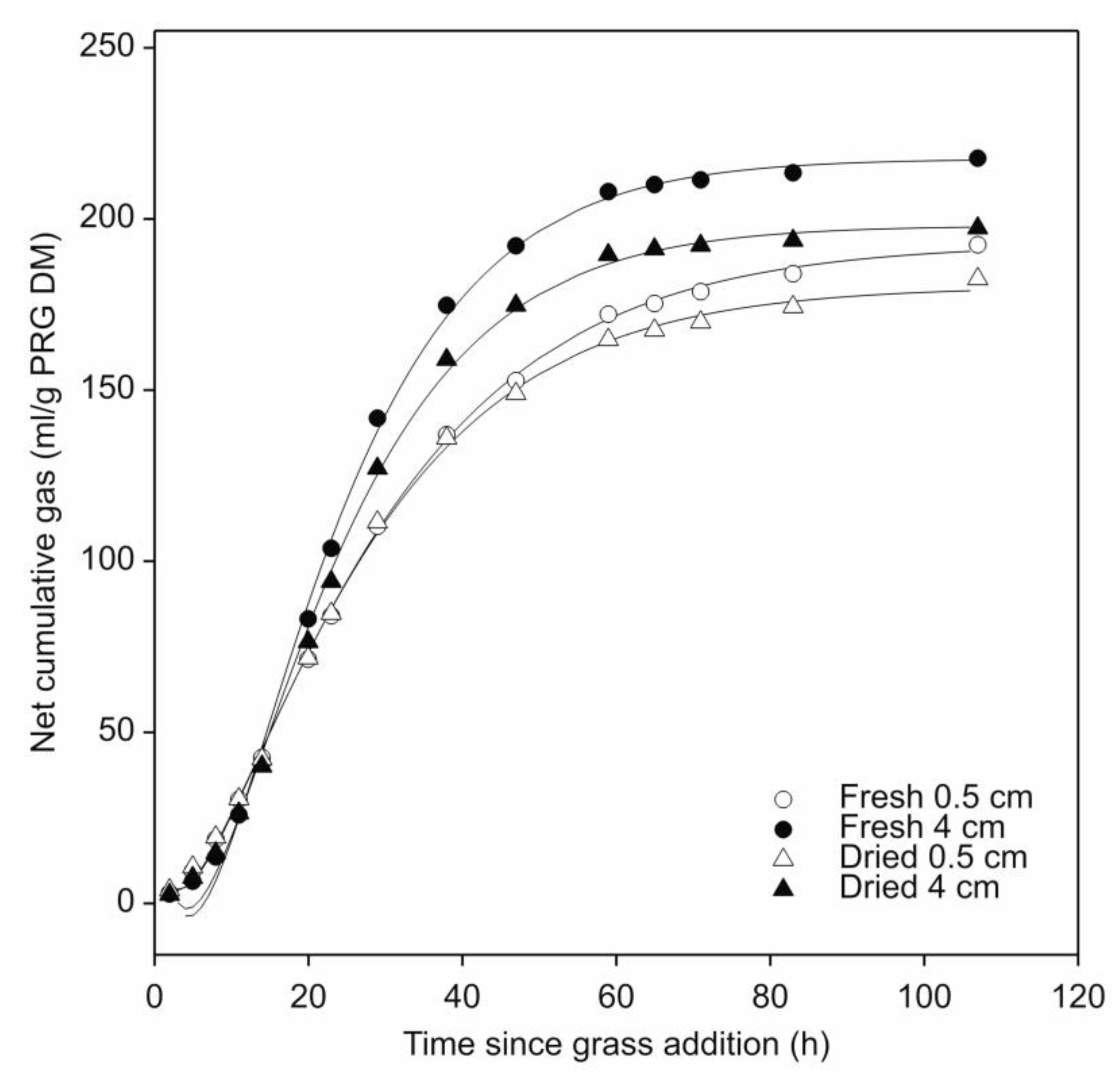
3.2. Effect of Two Different Cut-Lengths of Fresh and Air-Dried Leaf-Blades of Perennial Ryegrass on Their Fermentation by N. frontalis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gordon, G.L.R.; Phillips, M.W. The role of anaerobic gut fungi in ruminants. Nutr. Res. Rev. 1998, 11, 133–168. [Google Scholar] [CrossRef]
- Trinci, A.P.J.; Davies, D.R.; Gull, K.; Lawrence, M.I.; Bonde Nielsen, B.; Rickers, A.; Theodorou, M.K. Anaerobic fungi in herbivorous animals. Mycol. Res. 1994, 98, 129–152. [Google Scholar] [CrossRef]
- Solomon, K.V.; Haitjema, C.H.; Henske, J.K.; Gilmore, S.P.; Borges-Rivera, D.; Lipzen, A.; Brewer, H.M.; Purvine, S.O.; Wright, A.T.; Theodorou, M.K.; et al. Early-branching gut fungi possess large, comprehensive array of biomass-degrading enzymes. Science 2016, 351, 1192–1195. [Google Scholar] [CrossRef]
- Li, Y.; Jin, W.; Cheng, Y.; Zhu, W. Effect of the associated methanogen Methanobrevibacter thaueri on the dynamic profile of end and intermediate metabolites of anaerobic fungus Piromyces sp. F1. Curr. Microbiol. 2016, 73, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Dollhofer, V.; Callaghan, T.M.; Griffith, G.W.; Lebuhn, M.; Bauer, J. Presence and transcriptional activity of anaerobic fungi in agricultural biogas plants. Bioresour. Technol. 2017, 235, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Vinzelj, J.; Joshi, A.; Insam, H.; Podmirseg, S.M. Employing anaerobic fungi in biogas production: Challenges & opportunities. Bioresour. Technol. 2020, 300, 122867. [Google Scholar] [CrossRef]
- Henske, J.K.; Wilken, S.E.; Solomon, K.V.; Smallwood, C.R.; Shutthanandan, V.; Evans, J.E.; Theodorou, M.K.; O’Malley, M.A. Metabolic characterization of anaerobic fungi provides a path forward for bioprocessing of crude lignocellulose. Biotechnol. Bioeng. 2018, 115, 874–884. [Google Scholar] [CrossRef] [PubMed]
- Dollhofer, V.; Young, D.; Seppälä, S.; Hooker, C.; Youssef, N.; Podmirseg, S.M.; Nagler, M.; Reilly, M.; Li, Y.; Fliegerová, K.; et al. The biotechnological potential of the anaerobic gut fungi. In The Mycota Vol. II: Genetics and Biotechnology, 3rd ed.; Benz, J.P., Schipper, K., Eds.; Springer International Publishing: New York, NY, USA, 2020. [Google Scholar]
- Edwards, J.J.E.; Kingston-Smith, A.H.; Jimenez, H.R.; Huws, S.A.; Skøt, K.P.; Griffith, G.W.; McEwan, N.R.; Theodorou, M.K. Dynamics of initial colonization of nonconserved perennial ryegrass by anaerobic fungi in the bovine rumen. Fems Microbiol. Ecol. 2008, 66, 537–545. [Google Scholar] [CrossRef]
- Bauchop, T. Rumen anaerobic fungi of cattle and sheep. Appl. Environ. Microbiol. 1979, 38, 148–158. [Google Scholar] [CrossRef]
- Orpin, C.G. The rumen flagellate Piromonas communis: Its life history and invasion of plant material in the rumen. J. Gen. Microbiol. 1977, 99, 107–117. [Google Scholar] [CrossRef]
- Orpin, C.G. Anaerobic fungi—Taxonomy, biology, and distribution in nature. In Anaerobic Fungi: Biology, Ecology and Function; Mountford, D.O., Orpin, C.G., Eds.; Marcel Dekker: New York, NY, USA, 1994; pp. 1–45. [Google Scholar]
- Haitjema, C.H.; Gilmore, S.P.; Henske, J.K.; Solomon, K.V.; De Groot, R.; Kuo, A.; Mondo, S.J.; Salamov, A.A.; LaButti, K.; Zhao, Z.; et al. A parts list for fungal cellulosomes revealed by comparative genomics. Nat. Microbiol. 2017, 2, 17087. [Google Scholar] [CrossRef] [PubMed]
- Marvin-Sikkema, F.D.; Richardson, A.J.; Stewart, C.S.; Gottschal, J.C.; Prins, R.A. Influence of hydrogen-consuming bacteria on cellulose degradation by anaerobic fungi. Appl. Environ. Microbiol. 1990, 56, 3793–3797. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.Y.; Theodorou, M.K.; Nielsen, B.B.; Trinci, A.P.J. Dilution rate increases production of plant cell-wall degrading enzymes by anaerobic fungi in continuous-flow culture. Anaerobe 1997, 3, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.F.; Edwards, J.E.; Allison, G.G.; Zhu, W.Y.; Theodorou, M.K. Diversity and activity of enriched ruminal cultures of anaerobic fungi and methanogens grown together on lignocellulose in consecutive batch culture. Bioresour. Technol. 2009, 100, 4821–4828. [Google Scholar] [CrossRef] [PubMed]
- Joblin, K.N.; Campbell, G.P.; Richardson, A.J.; Stewart, C.S. Fermentation of barley straw by anaerobic rumen bacteria and fungi in axenic culture and in co-culture with methanogens. Lett. Appl. Microbiol. 1989, 9, 195–197. [Google Scholar] [CrossRef]
- Orpin, C.G. The occurrence of chitin in the cell walls of the rumen organisms Neocallimastix frontalis, Piromonas communis and Sphaeromonas communis. J. Gen. Microbiol. 1977, 99, 215–218. [Google Scholar] [CrossRef]
- Rezaeian, M.; Beakes, G.W.; Chaudhry, A.S. Effect of feeding chopped and pelleted lucerne on rumen fungal mass, fermentation profiles and in sacco degradation of barley straw in sheep. Anim. Feed Sci. Technol. 2006, 128, 292–306. [Google Scholar] [CrossRef]
- France, J.; Dhanoa, M.S.; Theodorou, M.K.; Lister, S.J.; Davies, D.R.; Isac, D. A model to interpret gas accumulation profiles associated with in vitro degradation of ruminant feeds. J. Theor. Biol. 1993, 163, 99–111. [Google Scholar] [CrossRef]
- Theodorou, M.K.; Williams, B.A.; Dhanoa, M.S.; McAllan, A.B.; France, J. A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. Anim. Feed Sci. Technol. 1994, 48, 185–197. [Google Scholar] [CrossRef]
- Theodorou, M.K.; Davies, D.R.; Nielsen, B.B.; Lawrence, M.I.G.; Trinci, A.P.J. Determination of growth of anaerobic fungi on soluble and cellulosic substrates using a pressure transducer. Microbiology 1995, 141, 671–678. [Google Scholar] [CrossRef]
- Stewart, C.S.; McPherson, C.A.; Cansunar, E. The effect of lasalocid on glucose uptake, hydrogen production and the solubilization of straw by the anaerobic rumen fungus Neocallimastix frontalis. Lett. Appl. Microbiol. 1987, 5, 5–7. [Google Scholar] [CrossRef]
- Davies, D.R.; Theodorou, M.K.; Lawrence, M.I.G.; Trinci, A.P.J. Distribution of anaerobic fungi in the digestive tract of cattle and their survival in faeces. J. Gen. Microbiol. 1993, 139, 1395–1400. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, H.R. Anaerobic Fungi in the Ruminant Digestive Tract. Ph.D. Thesis, Aberystwyth University, Aberystwyth, UK, 2008. [Google Scholar]
- Kim, E.J.; Parker, D.S.; Scollan, N.D. Fishmeal supplementation of steers fed on grass silage: Effects on rumen function, nutrient flow to and disappearance from the small intestine. Anim. Res. 2001, 50, 337–348. [Google Scholar] [CrossRef]
- Baird, D.B.; Murray, D.A.; Payne, R.W.; Soutar, D.M. Introduction to Genstat for Windows; VSN International: Hemel Hempstead, UK, 2019. [Google Scholar]
- Nielsen, B.B.; Zhu, W.Y.; Dhanoa, M.S.; Trinci, A.P.J.; Theodorou, M.K. Comparison of the growth kinetics of anaerobic gut fungi on wheat straw in batch culture. Anaerobe 2002, 8, 216–222. [Google Scholar] [CrossRef]
- Wilken, S.E.; Leggieri, P.A.; Kerdman-Andrade, C.; Reilly, M.; Theodorou, M.K.; O’Malley, M.A. An arduino based automatic pressure evaluation system (A-APES) to quantify growth of non-model anaerobes in culture. Aiche J. 2020. [Google Scholar] [CrossRef]
- Ren, Y.; Yu, M.; Wu, C.; Wang, Q.; Gao, M.; Huang, Q.; Liu, Y. A comprehensive review on food waste anaerobic digestion: Research updates and tendencies. Bioresour. Technol. 2018, 247, 1069–1076. [Google Scholar] [CrossRef]
- Kirby, M.E.; Theodorou, M.K.; Brizuela, C.M.; Huntington, J.A.; Powles, J.; Wilkinson, R.G. The anaerobic digestion of pig carcase with and without sugar beet pulp, as a novel on-farm disposal method. Waste Manag. 2018, 75, 251–260. [Google Scholar] [CrossRef]
- Papadimitriou, E.K. Hydrolysis of organic matter during autoclaving of commingled houshold waste. Waste Manag. 2010, 30, 572–582. [Google Scholar] [CrossRef]
- Pandey, A.; Larroche, C.; Dussap, C.-G.; Gnansunou, E.; Khanal, S.K.; Ricke, S.C. Biofuels: Alternative Feedstocks and Conversion Processes for the Production of Liquid and Gaseous Biofuels, 2nd ed.; Academic Press: San Diego, CA, USA, 2019. [Google Scholar]
- Garrote, G.; Domínguez, H.; Parajó, J. Hydrothermal processing of lignocellulosic materials. Holz Als Roh Und Werkst. 1999, 57, 191–202. [Google Scholar] [CrossRef]
- Gay, L. Chitin content and chitin synthase activity as indicators of the growth of three different anaerobic rumen fungi. Fems Microbiol. Lett. 1991, 80, 99–102. [Google Scholar] [CrossRef]
- Costa-de-Oliveira, S.; Silva, A.P.; Miranda, I.M.; Salvador, A.; Azevedo, M.M.; Munro, C.A.; Rodrigues, A.G.; Pina-Vaz, C. Determination of Chitin content in fungal cell wall: An alternative flow cytometric method. Cytom. Part A 2013, 83, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Wallander, H.; Ekblad, A.; Godbold, D.L.; Johnson, D.; Bahr, A.; Baldrian, P.; Björk, R.G.; Kieliszewska-Rokicka, B.; Kjøller, R.; Kraigher, H.; et al. Evaluation of methods to estimate production, biomass and turnover of ectomycorrhizal mycelium in forests soils—A review. Soil Biol. Biochem. 2013, 57, 1034–1047. [Google Scholar] [CrossRef]
- Henske, J.K.; Gilmore, S.P.; Knop, D.; Cunningham, F.J.; Sexton, J.A.; Smallwood, C.R.; Shutthanandan, V.; Evans, J.E.; Theodorou, M.K.; O’Malley, M.A. Transcriptomic characterization of Caecomyces churrovis: A novel, non-rhizoid-forming lignocellulolytic anaerobic fungus. Biotechnol. Biofuels 2017, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Marano, A.V.; Gleason, F.H.; Bärlocher, F.; Pires-Zottarelli, C.L.A.; Lilje, O.; Schmidt, S.K.; Rasconi, S.; Kagami, M.; Barrera, M.D.; Sime-Ngando, T.; et al. Quantitative methods for the analysis of zoosporic fungi. J. Microbiol. Methods 2012, 89, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Lowe, S.E.; Griffith, G.G.; Milne, A.; Theodorou, M.K.; Trinci, A.P.J. The life cycle and growth kinetics of an anaerobic rumen fungus. J. Gen. Microbiol. 1987, 133, 1815–1827. [Google Scholar] [CrossRef][Green Version]
- Theodorou, M.K.; Zhu, W.Y.; Rickers, A.; Nielsen, B.B.; Gull, K.; Trinci, A.P.J. Biochemistry and Ecology of Anaerobic Fungi. In Human and Animal Relationships. The Mycota (A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); Howard, D.H., Miller, J.D., Eds.; Springer: Berlin, Germany, 1996; Volume 6. [Google Scholar]
- Haitjema, C.H.; Solomon, K.V.; Henske, J.K.; Theodorou, M.K.; O’Malley, M.A. Anaerobic gut fungi: Advances in isolation, culture, and cellulolytic enzyme discovery for biofuel production. Biotechnol. Bioeng. 2014, 111, 1471–1482. [Google Scholar] [CrossRef] [PubMed]
- Eduard, W. Fungal spores: A critical review of the toxicological and epidemiological evidence as a basis for occupational exposure limit setting. Crit. Rev. Toxicol. 2009, 39, 799–864. [Google Scholar] [CrossRef]
- Shearer, C.A.; Descals, E.; Kohlmeyer, B.; Kohlmeyer, J.; Marvanová, L.; Padgett, D.; Porter, D.; Raja, H.A.; Schmit, J.P.; Thorton, H.A.; et al. Fungal biodiversity in aquatic habitats. Biodivers. Conserv. 2007, 16, 49–67. [Google Scholar] [CrossRef]
- Akin, D.E. Association of rumen fungi with various forage grasses. Anim. Feed Sci. Technol. 1987, 16, 273–285. [Google Scholar] [CrossRef]
- Windham, W.R.; Akin, D.E. Rumen fungi and forage fiber degradation. Appl. Environ. Microbiol. 1984, 48, 473–476. [Google Scholar] [CrossRef]
- France, J.; Theodorou, M.K.; Davies, D. Use of zoospore concentrations and life cycle parameters in determining the population of anaerobic fungi in the rumen ecosystem. J. Theor. Biol. 1990, 147, 413–422. [Google Scholar] [CrossRef]
- Kingston-Smith, A.H.; Davies, T.E.; Edwards, J.E.; Theodorou, M.K. From plants to animals; The role of plant cell death in ruminant herbivores. J. Exp. Bot. 2008, 59, 521–532. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Leaf-Blade Cut-Length (cm) * | p | |||
|---|---|---|---|---|---|
| 0.5 | 1.0 | 2.0 | 4.0 | ||
| Gas production (mL/g PRG DM #) | 186.7 a (184.1–188.8) | 201.6 b (199.4–203.4) | 199.6 b (197.1–201.3) | 221.1 c (217.5–224.7) | <0.05 |
| GP Half-life (t50) (h) | 31.8 a (30.9–32.7) | 30.1 b (29.3–30.8) | 28.2 c (27.8–28.9) | 26.4 d (24.6–27.6) | <0.05 |
| Fractional rate of GP at t50 (h−1) | 0.0450 a (0.0417–0.0479) | 0.0462 a (0.0427–0.0489) | 0.0494 a (0.0462–0.0516) | 0.0568 b (0.0517–0.0646) | <0.05 |
| Parameter | Leaf-Blade Cut-Length (cm) | s.e.m. | p * | p lin * | p dev * | |||
|---|---|---|---|---|---|---|---|---|
| 0.5 | 1.0 | 2.0 | 4.0 | |||||
| Initial DM Loss (g/g PRG DM) | 0.014 | 0.006 | 0.036 | 0.048 | 0.0175 | 0.332 | 0.102 | 0.832 |
| Apparent DM loss (g/g PRG DM) | 0.685 | 0.657 | 0.637 | 0.608 | 0.0147 | 0.019 | 0.003 | 0.576 |
| Formate (mmoL/g Apparent DM loss) | 2.30 | 3.30 | 2.89 | 4.00 | 0.408 | 0.068 | 0.024 | 0.311 |
| Acetate (mmoL/g Apparent DM loss) | 2.61 | 2.82 | 2.59 | 3.03 | 0.129 | 0.106 | 0.063 | 0.222 |
| Lactate (mmoL/g Apparent DM loss) | 1.27 | 1.39 | 1.25 | 1.10 | 0.082 | 0.165 | 0.061 | 0.437 |
| Parameter | Fresh Leaf-Blades * | Air-Dried Leaf-Blades * | p | ||
|---|---|---|---|---|---|
| 0.5 cm | 4.0 cm | 0.5 cm | 4.0 cm | ||
| Gas production (mL/g PRG DM #) | 192.7 b (187.9–195.7) | 227.2 c (223.4–233.6) | 180.2 a (174.3–184.1) | 198.0 b (194.9–204.7) | <0.05 |
| GP half-life (t50) (h) | 25.7 a (24.9–26.5) | 23.5 b (22.7–24.6) | 23.4 ab (23.1–25.1) | 23.4 b (22.6–24.5) | <0.05 |
| Fractional rate of GP at t50 (h−1) | 0.0397 a (0.0371–0.0436) | 0.0539 b (0.0471–0.0612) | 0.0437 ab (0.0397–0.0494) | 0.0526 b (0.0455–0.0592) | <0.05 |
| Variate | Length (L) | Condition (C) | Mean | C or L s.e.m. * | C × L s.e.m. | p | |||
|---|---|---|---|---|---|---|---|---|---|
| Fresh | Air-Dried | C | L | C × L | |||||
| Initial DM loss | 0.5 cm | 0.054 | 0.026 | 0.040 | 0.092 | 0.0131 | 0.180 | 0.245 | 0.137 |
| (g/g PRG DM) | 4.0 cm | 0.028 | 0.029 | 0.028 | |||||
| Mean | 0.041 | 0.028 | |||||||
| Apparent DM loss | 0.5 cm | 0.662 | 0.655 | 0.658 | 0.0036 | 0.0050 | 0.895 | < 0.001 | 0.281 |
| (g/g PRG DM) | 4.0 cm | 0.629 | 0.634 | 0.631 | |||||
| Mean | 0.645 | 0.644 | |||||||
| Formate | 0.5 cm | 2.79 | 3.41 | 3.10 | 0.204 | 0.288 | 0.179 | 0.023 | 0.476 |
| (mmoL/g PRG DM loss) | 4.0 cm | 3.76 | 3.97 | 3.86 | |||||
| Mean | 3.28 | 3.69 | |||||||
| Acetate | 0.5 cm | 2.50 | 2.65 | 2.57 | 0.108 | 0.153 | 0.882 | < 0.001 | 0.280 |
| (mmoL/g PRG DM loss) | 4.0 cm | 3.47 | 3.27 | 3.37 | |||||
| Mean | 2.99 | 2.96 | |||||||
| Lactate | 0.5 cm | 2.26 | 2.08 | 2.17 | 0.072 | 0.102 | 0.054 | 0.644 | 0.714 |
| (mmoL/g PRG DM loss) | 4.0 cm | 2.35 | 2.09 | 2.22 | |||||
| Mean | 2.31 | 2.09 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jimenez, H.R.; Edwards, J.E.; Sanderson, R.; Kingston-Smith, A.H.; McEwan, N.R.; Theodorou, M.K. Cut-Lengths of Perennial Ryegrass Leaf-Blades Influences In Vitro Fermentation by the Anaerobic Fungus Neocallimastix frontalis. Microorganisms 2020, 8, 1774. https://doi.org/10.3390/microorganisms8111774
Jimenez HR, Edwards JE, Sanderson R, Kingston-Smith AH, McEwan NR, Theodorou MK. Cut-Lengths of Perennial Ryegrass Leaf-Blades Influences In Vitro Fermentation by the Anaerobic Fungus Neocallimastix frontalis. Microorganisms. 2020; 8(11):1774. https://doi.org/10.3390/microorganisms8111774
Chicago/Turabian StyleJimenez, Hugo R., Joan E. Edwards, Ruth Sanderson, Alison H. Kingston-Smith, Neil R. McEwan, and Michael K. Theodorou. 2020. "Cut-Lengths of Perennial Ryegrass Leaf-Blades Influences In Vitro Fermentation by the Anaerobic Fungus Neocallimastix frontalis" Microorganisms 8, no. 11: 1774. https://doi.org/10.3390/microorganisms8111774
APA StyleJimenez, H. R., Edwards, J. E., Sanderson, R., Kingston-Smith, A. H., McEwan, N. R., & Theodorou, M. K. (2020). Cut-Lengths of Perennial Ryegrass Leaf-Blades Influences In Vitro Fermentation by the Anaerobic Fungus Neocallimastix frontalis. Microorganisms, 8(11), 1774. https://doi.org/10.3390/microorganisms8111774