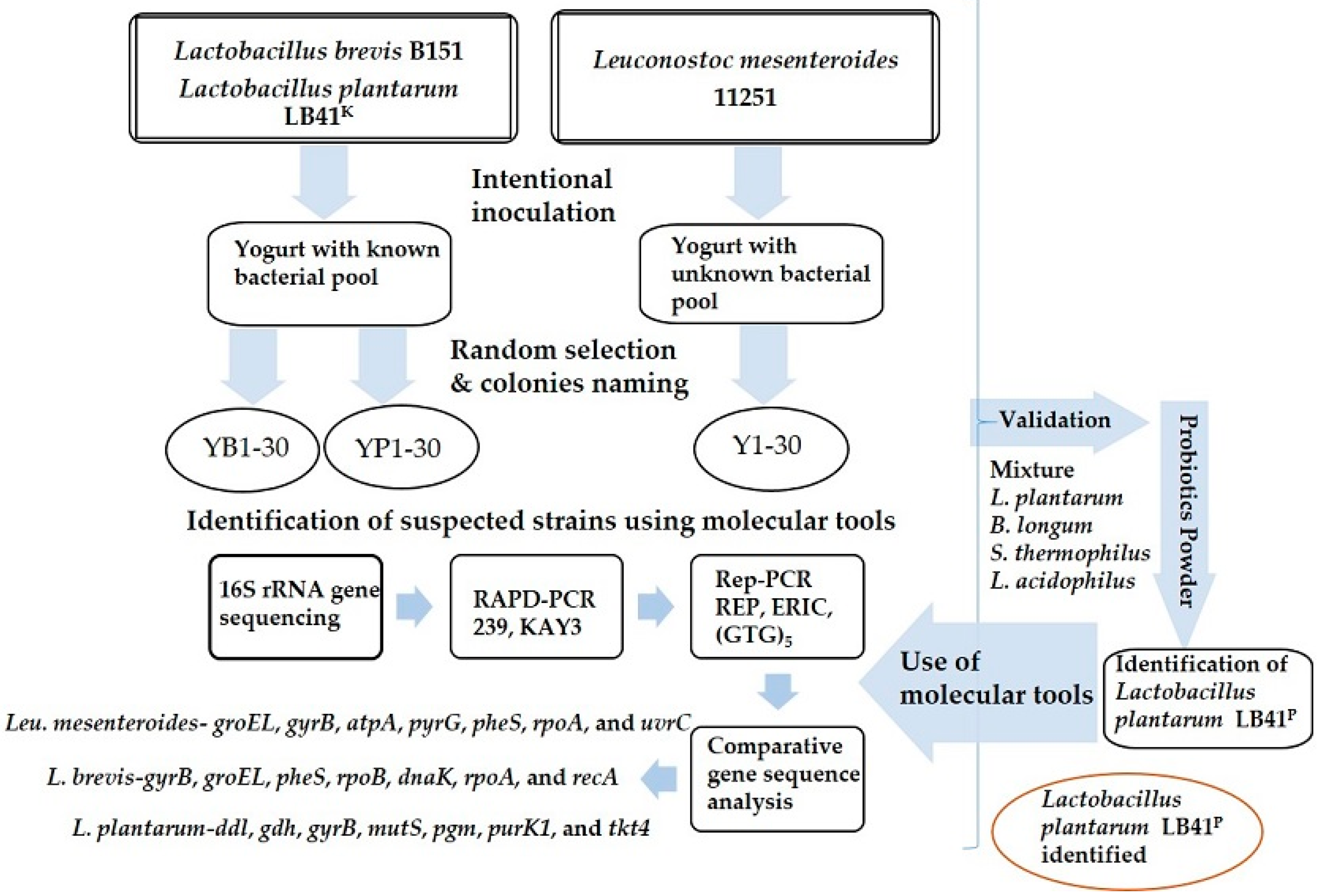
Tracking of Intentionally Inoculated Lactic Acid Bacteria Strains in Yogurt and Probiotic Powder


Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Yogurt and Probiotic Powder
2.3. DNA Isolation and PCR
2.4. Primer Synthesis and PCR
2.5. 16 S rRNA Gene Sequencing and RAPD Analysis
2.6. Rep-PCR Analysis
2.7. Comparative Sequence Analysis of the Housekeeping Genes
2.8. Tracing of the Intentionally Inoculated Strains
3. Results
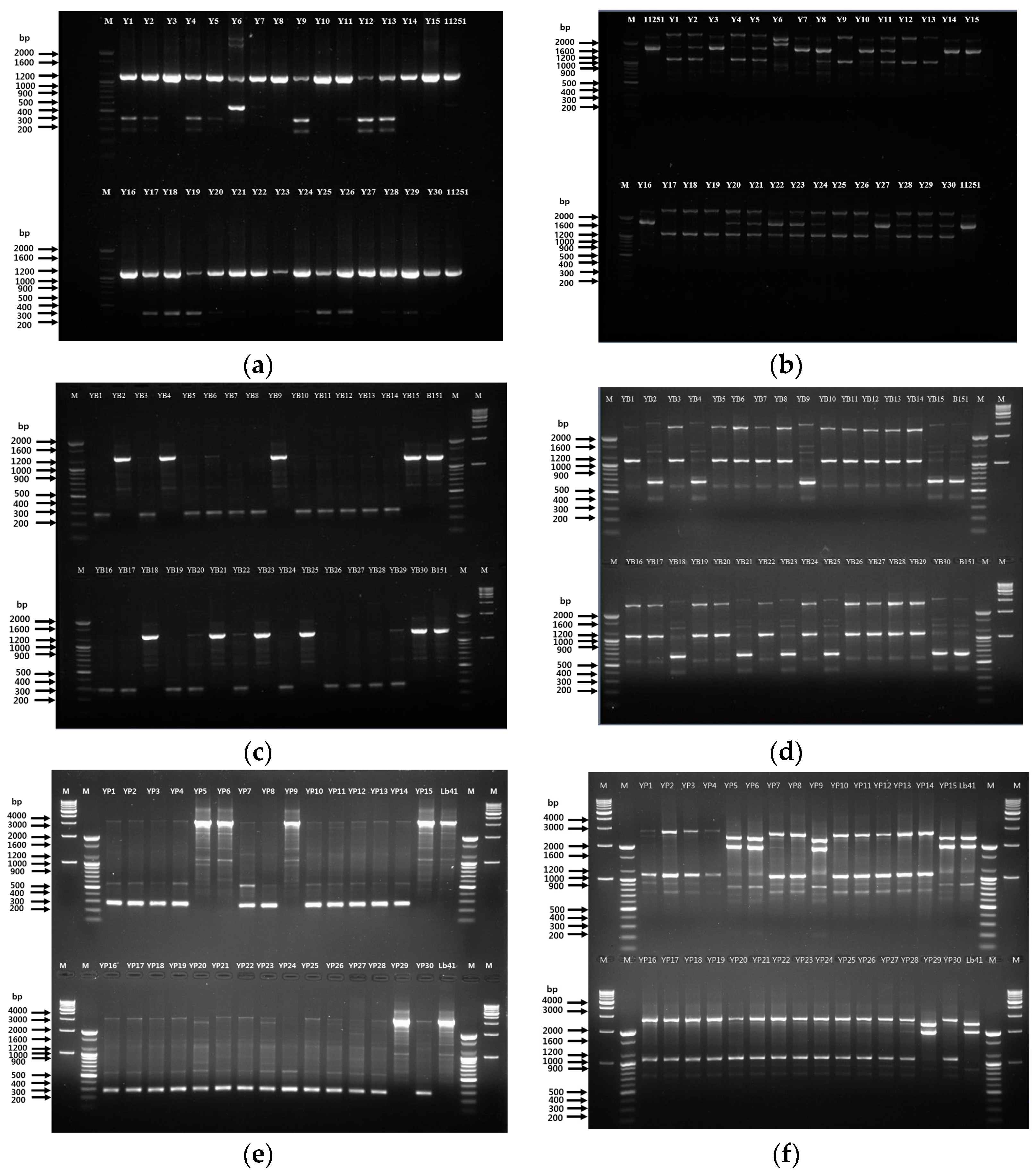
3.1. Analysis of 16S rRNA Sequences
3.2. Analysis of RAPD-PCR Fingerprints
3.3. Analysis of Rep-PCR Fingerprints
3.4. Comparative Housekeeping Gene Analysis
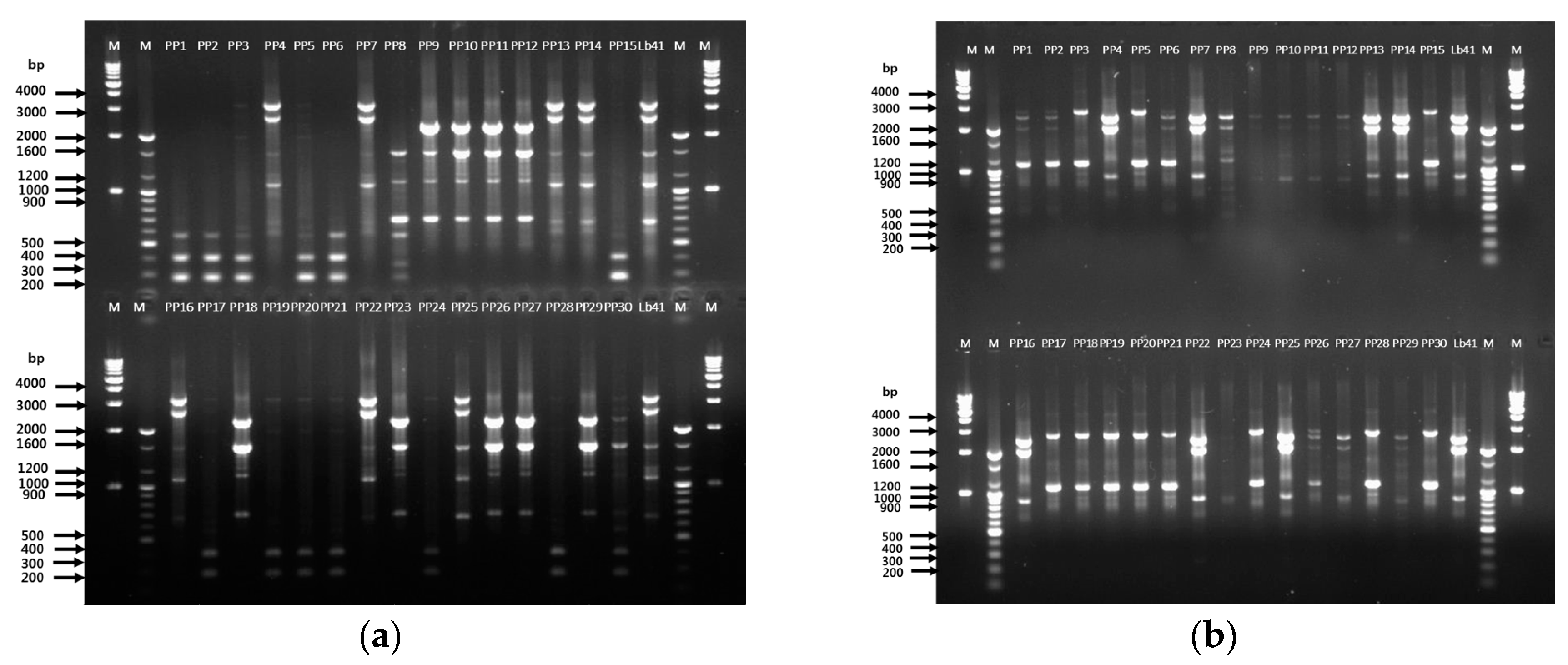
3.5. Validation Using Tracing in a Probiotic Powder
4. Discussion
5. Strengths and Limitations of the Study
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wassenaar, T.M.; Klein, G. Safety aspects and implications of regulation of probiotic bacteria in food and food supplements. J. Food Protect. 2008, 71, 1734–1741. [Google Scholar] [CrossRef] [PubMed]
- Marteau, P.R.; Vrese, M.D.; Cellier, C.J.; Schrezenmeir, J. Protection from gastrointestinal diseases with the use of probiotics. Am. J. Clin. Nutr. 2001, 73, 430s–436s. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.L.; Klaenhammer, T.R. Genomic evolution of domesticated microorganisms. Ann. Rev. Food Sci. Technol. 2010, 1, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.-H.; Han, S.H.; Kim, J.-S.; Kim, Y.; Jeong, Y.; Park, H.M.; Paek, N.-S. Inhibition of nitric oxide production, oxidative stress prevention, and probiotic activity of lactic acid bacteria isolated from the human vagina and fermented food. Microorganisms 2019, 7, 109. [Google Scholar] [CrossRef]
- Kamiya, T.; Watanabe, Y.; Makino, S.; Kano, H.; Tsuji, N. Improvement of intestinal immune cell function by lactic acid bacteria for dairy products. Microorganisms 2016, 5, 1. [Google Scholar] [CrossRef]
- Baradaran, A.; Foo, H.L.; Sieo, C.C.; Raha, A. Isolation, identification and characterization of lactic acid bacteria from Polygonum minus. Rom. Biotechnol. Lett. 2012, 17, 7245–7252. [Google Scholar]
- Fu, L.-L.; Li, J.-R. Microbial source tracking: A tool for identifying sources of microbial contamination in the food chain. Crit. Rev. Food Sci. Nutr. 2014, 54, 699–707. [Google Scholar] [CrossRef]
- Temmerman, R.; Huys, G.; Swings, J. Identification of lactic acid bacteria: Culture-dependent and culture-independent methods. Trends Food Sci. Technol. 2004, 15, 348–359. [Google Scholar] [CrossRef]
- Dan, T.; Liu, W.; Sun, Z.; Lv, Q.; Xu, H.; Song, Y.; Zhang, H. A novel multi-locus sequence typing (MLST) protocol for Leuconostoc lactis isolates from traditional dairy products in China and Mongolia. BMC Microbiol. 2014, 14, 150. [Google Scholar] [CrossRef]
- Ben Amor, K.; Vaughan, E.E.; de Vos, W.M. Advanced molecular tools for the identification of lactic acid bacteria. J. Nutr. 2007, 137, 741S–747S. [Google Scholar] [CrossRef]
- Davis, C. Enumeration of probiotic strains: Review of culture-dependent and alternative techniques to quantify viable bacteria. J. Microbiol. Methods 2014, 103, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Lee, S.; Park, Y.-S.; Sharma, A. RAPD analysis of Leuconostoc mesenteroides strains associated with vegetables and food products from Korea. LWT-Food Sci. Technol. 2017, 77, 383–388. [Google Scholar] [CrossRef]
- Abdollahniya, D.; Hosseini, S.M.; Baghbaderani, B.K.; Mordadi, A.; Reza, M. Identification of Lactobacillus species isolated from traditional dairy products using RAPD-PCR. Avicenna J. Clin. Microbiol. Infect. 2018, 5, 7–13. [Google Scholar]
- Gevers, D.; Huys, G.; Swings, J. Applicability of rep-PCR fingerprinting for identification of Lactobacillus species. FEMS Microbiol. Lett. 2001, 205, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Healy, M.; Huong, J.; Bittner, T.; Lising, M.; Frye, S.; Raza, S.; Schrock, R.; Manry, J.; Renwick, A.; Nieto, R. Microbial DNA typing by automated repetitive-sequence-based PCR. J. Clin. Microbiol. 2005, 43, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Gallego, J.; Arroyo-López, F.; Rantsiou, K.; Jiménez-Díaz, R.; Garrido-Fernández, A.; Cocolin, L. Screening of lactic acid bacteria isolated from fermented table olives with probiotic potential. Food Res. Int. 2013, 50, 135–142. [Google Scholar] [CrossRef]
- Vankerckhoven, V.; Huys, G.; Vancanneyt, M.; Vael, C.; Klare, I.; Romond, M.-B.; Entenza, J.M.; Moreillon, P.; Wind, R.D.; Knol, J. Biosafety assessment of probiotics used for human consumption: Recommendations from the EU-PROSAFE project. Trends Food Sci. Technol. 2008, 19, 102–114. [Google Scholar] [CrossRef]
- Laroute, V.; Tormo, H.; Couderc, C.; Mercier-Bonin, M.; Le Bourgeois, P.; Cocaign-Bousquet, M.; Daveran-Mingot, M.-L. From genome to phenotype: An integrative approach to evaluate the biodiversity of Lactococcus lactis. Microorganisms 2017, 5, 27. [Google Scholar] [CrossRef]
- Ruiz, P.; Celada, L.; Seseña, S.; Palop, M.L. Leuconostoc mesenteroides in the brewing process: A controversial role. Food Control 2018, 90, 415–421. [Google Scholar] [CrossRef]
- Salvetti, E.; Torriani, S.; Felis, G.E. The genus Lactobacillus: A taxonomic update. Probiotics Antimicrob. Proteins 2012, 4, 217–226. [Google Scholar] [CrossRef]
- Fusco, V.; Quero, G.M.; Chieffi, D.; Franz, C.M. Identification of Lactobacillus brevis using a species-specific AFLP-derived marker. Int. J. Food Microbiol. 2016, 232, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Xu, J.; Li, Q.; Xia, X.; Du, G. Characterization of a Lactobacillus brevis strain with potential oral probiotic properties. BMC Microbiol. 2018, 18, 221. [Google Scholar] [CrossRef] [PubMed]
- Bergsveinson, J.; Pittet, V.; Ewen, E.; Baecker, N.; Ziola, B. Genome sequence of rapid beer-spoiling isolate Lactobacillus brevis BSO 464. Genome Announc. 2015, 3, e01411–e01415. [Google Scholar] [CrossRef] [PubMed]
- Siezen, R.J.; Tzeneva, V.A.; Castioni, A.; Wels, M.; Phan, H.T.; Rademaker, J.L.; Starrenburg, M.J.; Kleerebezem, M.; Molenaar, D.; van Hylckama Vlieg, J.E. Phenotypic and genomic diversity of Lactobacillus plantarum strains isolated from various environmental niches. Environ. Microbiol. 2010, 12, 758–773. [Google Scholar] [CrossRef] [PubMed]
- De las Rivas, B.; Marcobal, Á.; Muñoz, R. Development of a multilocus sequence typing method for analysis of Lactobacillus plantarum strains. Microbiology 2006, 152, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Altay, F.; Karbancıoglu-Güler, F.; Daskaya-Dikmen, C.; Heperkan, D. A review on traditional Turkish fermented non-alcoholic beverages: Microbiota, fermentation process and quality characteristics. Int. J. Food Microbiol. 2013, 167, 44–56. [Google Scholar] [CrossRef]
- Guidone, A.; Zotta, T.; Ross, R.P.; Stanton, C.; Rea, M.C.; Parente, E.; Ricciardi, A. Functional properties of Lactobacillus plantarum strains: A multivariate screening study. LWT-Food Sci. Technol. 2014, 56, 69–76. [Google Scholar] [CrossRef]
- Versalovic, J.; Schneider, M.; De Bruijn, F.; Lupski, J.R. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol. Cell. Biol. 1994, 5, 25–40. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Franz, C.; Cho, G.-S.; Holzapfel, W.H.; Gálvez, A. Safety of Lactic Acid Bacteria; Wiley Online Library: Hoboken, NJ, USA, 2010; pp. 341–360. [Google Scholar]
- Fenster, K.; Freeburg, B.; Hollard, C.; Wong, C.; Rønhave Laursen, R.; Ouwehand, A.C. The production and delivery of probiotics: A review of a practical approach. Microorganisms 2019, 7, 83. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.A.; Schoeni, J.L.; Vegge, C.; Pane, M.; Stahl, B.; Bradley, M.; Goldman, V.S.; Burguière, P.; Atwater, J.B.; Sanders, M.E. Improving end-user trust in the quality of commercial probiotic products. Front. Microbiol. 2019, 10, 739. [Google Scholar] [CrossRef] [PubMed]
- De Vos, W.M. Systems solutions by lactic acid bacteria: From paradigms to practice. In Proceedings of the Microbial Cell Factories, Egmond aan Zee, The Netherlands, 28 August–1 September 2011; p. S2. [Google Scholar]
- Cibik, R.; Lepage, E.; Tailliez, P. Molecular diversity of Leuconostoc mesenteroides and Leuconostoc citreum isolated from traditional French cheeses as revealed by RAPD GTG printing, 16S rDNA sequencing and 16S rDNA fragment amplification. Syst. Appl. Microbiol. 2000, 23, 267–278. [Google Scholar] [CrossRef]
- Chun, B.H.; Kim, K.H.; Jeon, H.H.; Lee, S.H.; Jeon, C.O. Pan-genomic and transcriptomic analyses of Leuconostoc mesenteroides provide insights into its genomic and metabolic features and roles in kimchi fermentation. Sci. Rep. 2017, 7, 11504. [Google Scholar] [CrossRef]
- Fox, G.E.; Wisotzkey, J.D.; Jurtshuk, P., Jr. How close is close: 16S rRNA sequence identity may not be sufficient to guarantee species identity. Int. J. Syst. Evol. Microbiol. 1992, 42, 166–170. [Google Scholar] [CrossRef]
- Ashraf, R.; Shah, N.P. Selective and differential enumerations of Lactobacillus delbrueckii subsp. bulgaricus, Streptococcus thermophilus, Lactobacillus acidophilus, Lactobacillus casei and Bifidobacterium spp. in yoghurt—A review. Int. J. Food Microbiol. 2011, 149, 194–208. [Google Scholar] [CrossRef]
- Kaur, J.; Lee, S.; Sharma, A.; Park, Y.-S. DNA profiling of Leuconostoc mesenteroides strains isolated from fermented foods and farm produce in Korea by repetitive-element PCR. Food Sci. Biotechnol. 2017, 26, 1667–1673. [Google Scholar] [CrossRef]
- Williams, J.G.; Hanafey, M.K.; Rafalski, J.A.; Tingey, S.V. Genetic analysis using random amplified polymorphic DNA markers. In Recombinant DNA Methodology II; Elsevier: Amsterdam, The Netherlands, 1995; pp. 849–884. [Google Scholar]
- Gevers, D.; Cohan, F.M.; Lawrence, J.G.; Spratt, B.G.; Coenye, T.; Feil, E.J.; Stackebrandt, E.; Van de Peer, Y.; Vandamme, P.; Thompson, F.L. Re-evaluating prokaryotic species. Nat. Rev. Microbiol. 2005, 3, 733. [Google Scholar] [CrossRef]
- Konstantinidis, K.T.; Tiedje, J.M. Towards a genome-based taxonomy for prokaryotes. J. Bacteriol. 2005, 187, 6258–6264. [Google Scholar] [CrossRef]
- Shevtsov, A.; Kushugulova, A.; Tynybaeva, I.; Kozhakhmetov, S.; Abzhalelov, A.; Momynaliev, K.; Stoyanova, L. Identification of phenotypically and genotypically related Lactobacillus strains based on nucleotide sequence analysis of the groEL, rpoB, rplB, and 16S rRNA genes. Microbiology 2011, 80, 672. [Google Scholar] [CrossRef]
- Sharma, A.; Kaur, J.; Lee, S.; Park, Y.-S. Analysis of Leuconostoc citreum strains using multilocus sequence typing. Food Sci. Biotechnol. 2018, 27, 1755–1760. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kaur, J.; Lee, S.; Park, Y.-S. Genetic diversity analysis of Leuconostoc mesenteroides from Korean vegetables and food products by multilocus sequence typing. Appl. Microbiol. Biotechnol. 2018, 102, 4853–4861. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kaur, J.; Lee, S.; Park, Y.-S. Molecular discrimination of Lactobacillus brevis strains isolated from food products in South Korea using multilocus sequence typing. LWT-Food Sci. Technol. 2017, 86, 337–343. [Google Scholar] [CrossRef]
- Behera, S.S.; Ray, R.C.; Zdolec, N. Lactobacillus plantarum with functional properties: An approach to increase safety and shelf-life of fermented foods. Biol. Med. Res. Int. 2018, 2018. [Google Scholar] [CrossRef]
- Van Nevel, S.; Koetzsch, S.; Proctor, C.R.; Besmer, M.D.; Prest, E.I.; Vrouwenvelder, J.S.; Knezev, A.; Boon, N.; Hammes, F. Flow cytometric bacterial cell counts challenge conventional heterotrophic plate counts for routine microbiological drinking water monitoring. Water Res. 2017, 113, 191–206. [Google Scholar] [CrossRef]
- Chiron, C.; Tompkins, T.A.; Burguière, P. Flow cytometry: A versatile technology for specific quantification and viability assessment of micro-organisms in multistrain probiotic products. J. Appl. Microbiol. 2018, 124, 572–584. [Google Scholar] [CrossRef]
- Rodríguez-Sánchez, B.; Marín, M.; Sánchez-Carrillo, C.; Cercenado, E.; Ruiz, A.; Rodríguez-Créixems, M.; Bouza, E. Improvement of matrix-assisted laser desorption/ionization time-of-flight mass spectrometry identification of difficult-to-identify bacteria and its impact in the workflow of a clinical microbiology laboratory. Diagn. Microbiol. Infect. Dis. 2014, 79, 1–6. [Google Scholar] [CrossRef]
- Sloan, A.; Wang, G.; Cheng, K. Traditional approaches versus mass spectrometry in bacterial identification and typing. Clin. Chim. Acta 2017, 473, 180–185. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial No. | Colony ID | BLAST * Result | Colony ID | BLAST Result | Colony ID | BLAST Result |
|---|---|---|---|---|---|---|
| 1. | Y1 | S. thermophilus | YB1 | S. thermophilus | YP1 | S. thermophilus |
| 2. | Y2 | S. thermophilus | YB2 | L. brevis | YP2 | S. thermophilus |
| 3. | Y3 | Leu. mesenteroides | YB3 | S. thermophilus | YP3 | S. thermophilus |
| 4. | Y4 | S. thermophilus | YB4 | L. brevis | YP4 | S. thermophilus |
| 5. | Y5 | S. thermophilus | YB5 | S. thermophilus | YP5 | L. plantarum |
| 6. | Y6 | L. plantarum | YB6 | S. thermophilus | YP6 | L. plantarum |
| 7. | Y7 | Leu. mesenteroides | YB7 | S. thermophilus | YP7 | S. thermophilus |
| 8. | Y8 | S. thermophilus | YB8 | S. thermophilus | YP8 | S. thermophilus |
| 9. | Y9 | S. thermophilus | YB9 | L. brevis | YP9 | L. plantarum |
| 10. | Y10 | Leu. mesenteroides | YB10 | S. thermophilus | YP10 | S. thermophilus |
| 11. | Y11 | S. thermophilus | YB11 | S. thermophilus | YP11 | S. thermophilus |
| 12. | Y12 | S. thermophilus | YB12 | S. thermophilus | YP12 | S. thermophilus |
| 13. | Y13 | S. thermophilus | YB13 | S. thermophilus | YP13 | S. thermophilus |
| 14. | Y14 | Leu. mesenteroides | YB14 | S. thermophilus | YP14 | S. thermophilus |
| 15. | Y15 | Leu. mesenteroides | YB15 | L. brevis | YP15 | L. plantarum |
| 16. | Y16 | Leu. mesenteroides | YB16 | S. thermophilus | YP16 | S. thermophilus |
| 17. | Y17 | S. thermophilus | YB17 | S. thermophilus | YP17 | S. thermophilus |
| 18. | Y18 | S. thermophilus | YB18 | L. brevis | YP18 | S. thermophilus |
| 19. | Y19 | S. thermophilus | YB19 | S. thermophilus | YP19 | S. thermophilus |
| 20. | Y20 | S. thermophilus | YB20 | S. thermophilus | YP20 | S. thermophilus |
| 21. | Y21 | S. thermophilus | YB21 | L. brevis | YP21 | S. thermophilus |
| 22. | Y22 | S. thermophilus | YB22 | S. thermophilus | YP22 | S. thermophilus |
| 23. | Y23 | S. thermophilus | YB23 | L. brevis | YP23 | S. thermophilus |
| 24. | Y24 | S. thermophilus | YB24 | S. thermophilus | YP24 | S. thermophilus |
| 25. | Y25 | S. thermophilus | YB25 | L. brevis | YP25 | S. thermophilus |
| 26. | Y26 | S. thermophilus | YB26 | S. thermophilus | YP26 | S. thermophilus |
| 27. | Y27 | Leu. mesenteroides | YB27 | S. thermophilus | YP27 | S. thermophilus |
| 28. | Y28 | S. thermophilus | YB28 | S. thermophilus | YP28 | S. thermophilus |
| 29. | Y29 | S. thermophilus | YB29 | S. thermophilus | YP29 | L. plantarum |
| 30. | Y30 | S. thermophilus | YB30 | L. brevis | YP30 | S. thermophilus |
| Serial No. | Colony ID | BLAST Result | Serial No. | Colony ID | BLAST Result |
|---|---|---|---|---|---|
| 1. | PP1 | S. thermophilus | 16. | PP16 | L. plantarum |
| 2. | PP2 | S. thermophilus | 17. | PP17 | S. thermophiles |
| 3. | PP3 | S. thermophilus | 18. | PP18 | L. acidophilus |
| 4. | PP4 | L. plantarum | 19. | PP19 | S. thermophilus |
| 5. | PP5 | S. thermophilus | 20. | PP20 | S. thermophilus |
| 6. | PP6 | S. thermophilus | 21. | PP21 | S. thermophilus |
| 7. | PP7 | L. plantarum | 22. | PP22 | L. plantarum |
| 8. | PP8 * | ------------- | 23. | PP23 | L. acidophilus |
| 9. | PP9 | L. acidophilus | 24. | PP24 | S. thermophilus |
| 10. | PP10 | L. acidophilus | 25. | PP25 | L. plantarum |
| 11. | PP11 | L. acidophilus | 26. | PP26 | L. acidophilus |
| 12. | PP12 | L. acidophilus | 27. | PP27 | L. acidophilus |
| 13. | PP13 | L. plantarum | 28. | PP28 | S. thermophilus |
| 14. | PP14 | L. plantarum | 29. | PP29 | L. acidophilus |
| 15. | PP15 | S. thermophilus | 30. | PP30 | S. thermophilus |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, A.; Kaur, J.; Lee, S.; Park, Y.-S. Tracking of Intentionally Inoculated Lactic Acid Bacteria Strains in Yogurt and Probiotic Powder. Microorganisms 2020, 8, 5. https://doi.org/10.3390/microorganisms8010005
Sharma A, Kaur J, Lee S, Park Y-S. Tracking of Intentionally Inoculated Lactic Acid Bacteria Strains in Yogurt and Probiotic Powder. Microorganisms. 2020; 8(1):5. https://doi.org/10.3390/microorganisms8010005
Chicago/Turabian StyleSharma, Anshul, Jasmine Kaur, Sulhee Lee, and Young-Seo Park. 2020. "Tracking of Intentionally Inoculated Lactic Acid Bacteria Strains in Yogurt and Probiotic Powder" Microorganisms 8, no. 1: 5. https://doi.org/10.3390/microorganisms8010005
APA StyleSharma, A., Kaur, J., Lee, S., & Park, Y.-S. (2020). Tracking of Intentionally Inoculated Lactic Acid Bacteria Strains in Yogurt and Probiotic Powder. Microorganisms, 8(1), 5. https://doi.org/10.3390/microorganisms8010005