Anti-Biofilm Effect of Selected Essential Oils and Main Components on Mono- and Polymicrobic Bacterial Cultures


 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. EOs
2.3. Determination of Minimum Inhibitory Concentration (MIC)
2.4. Mono- and Dual-Species Biofilm Formation
2.5. Scanning Electron Microscopy (SEM) Observations
2.6. Statistical Analysis
3. Results
3.1. Composition of EOs and MICs
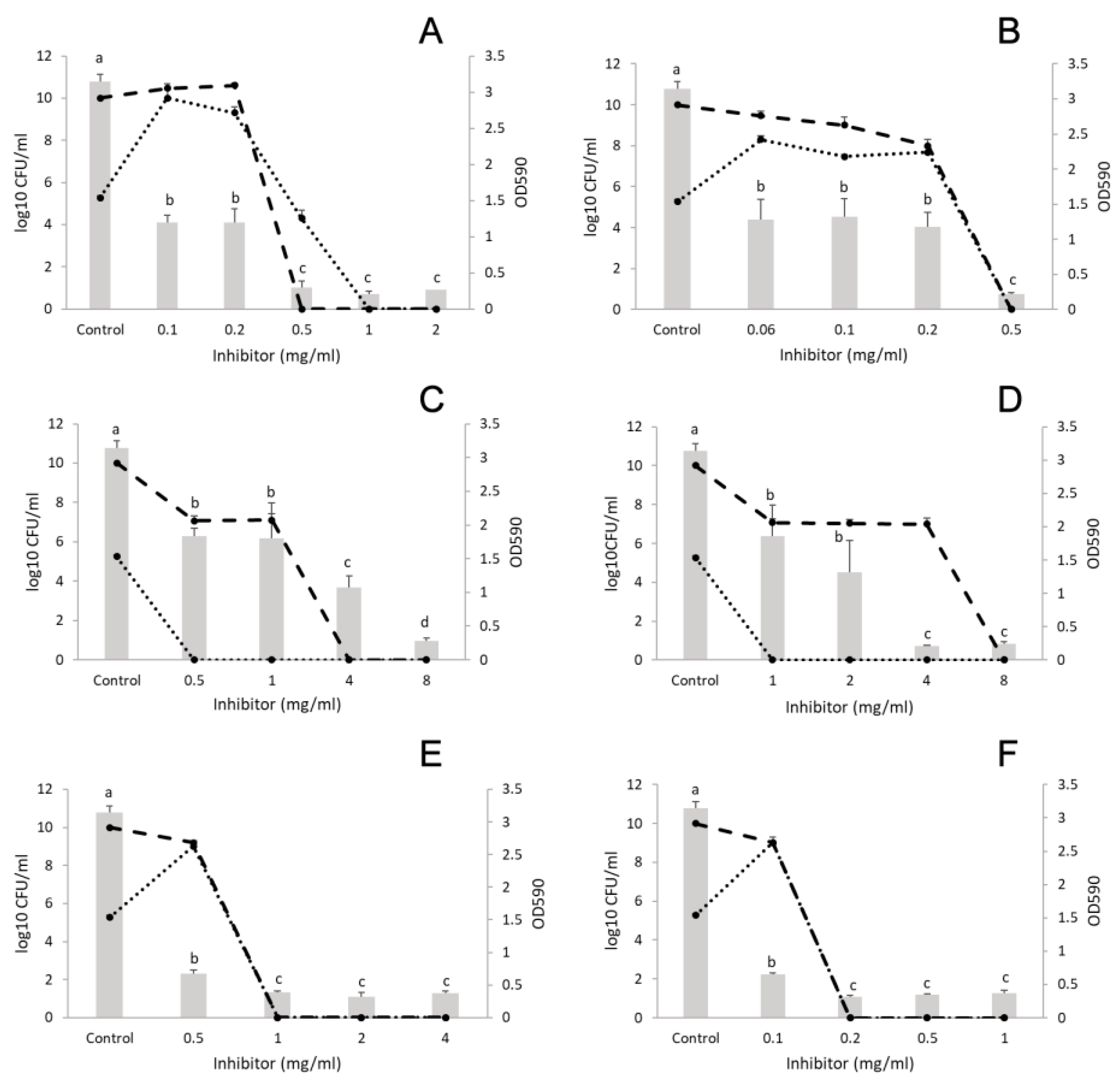
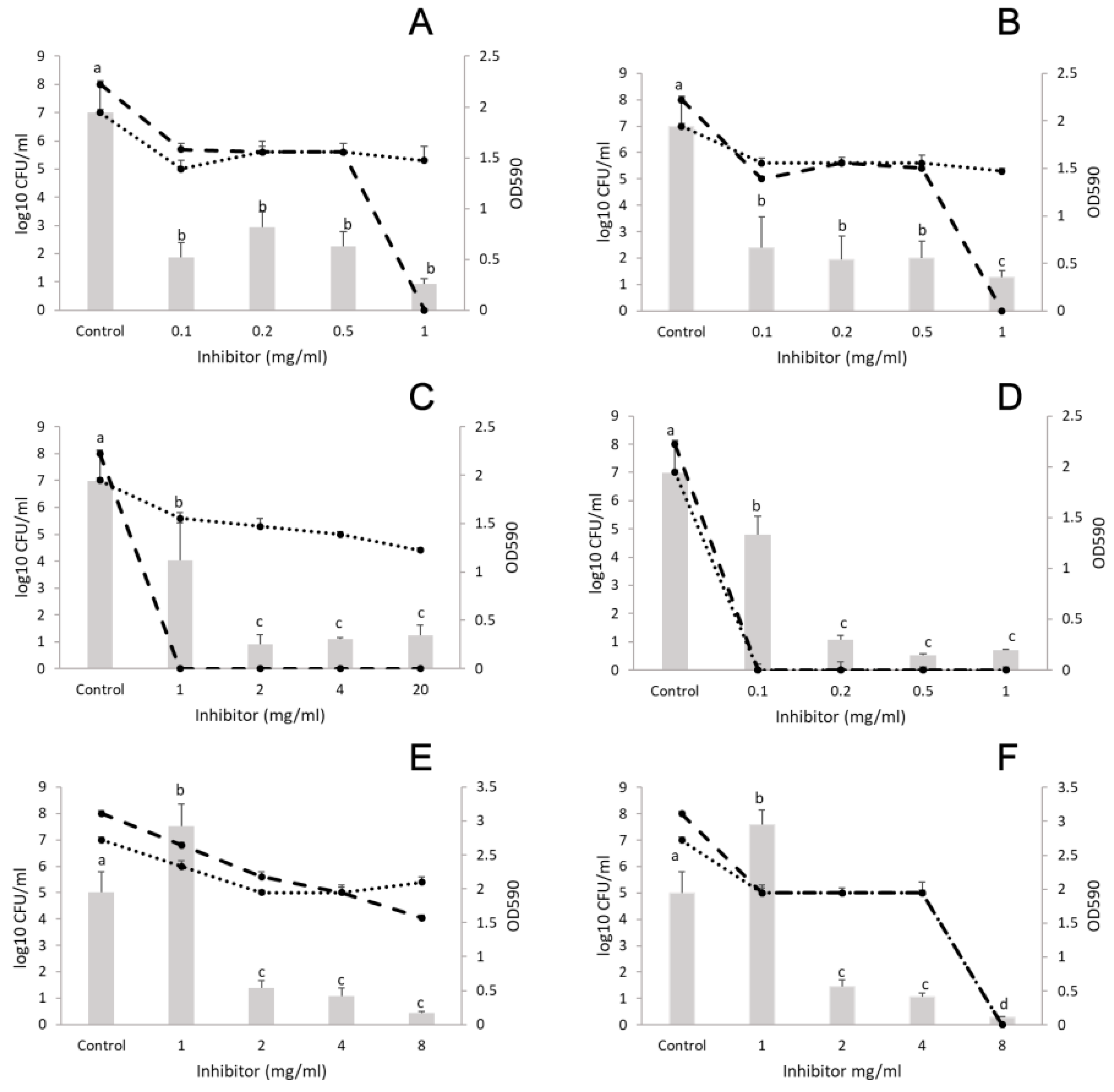
3.2. Anti-Biofilm-Forming Effect of EOs and Their Major Components
3.2.1. Monocultures
3.2.2. Polymicrobial Cultures
L. monocytogenes and E. coli
L. monocytogenes and S. aureus
L. monocytogenes and P. putida
3.3. SEM Observations
3.3.1. L. monocytogenes and E. coli
3.3.2. L. monocytogenes and S. aureus
3.3.3. L. monocytogenes and P. putida
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Shi, X.; Zhu, X. Biofilm formation and food safety in food industries. Trends Food Sci. Technol. 2009, 20, 407–413. [Google Scholar] [CrossRef]
- Midelet, G.; Carpentier, B. Transfer of microorganisms, including Listeria monocytogenes, from various materials to beef. Appl. Environ. Microbiol. 2002, 68, 4015–4024. [Google Scholar] [CrossRef]
- Johnson, L.R. Microcolony and biofilm formation as a survival strategy for bacteria. J. Theor. Biol. 2008, 251, 24–34. [Google Scholar] [CrossRef]
- Wong, H.S.; Townsend, K.M.; Fenwick, S.G.; Trengove, R.D.; O’Handley, R.M. Comparative susceptibility of planktonic and 3-day-old Salmonella Typhimurium biofilms to disinfectants. J. Appl. Microbiol. 2010, 108, 2222–2228. [Google Scholar] [CrossRef]
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Rendueles, O.; Ghigo, J.M. Multi-species biofilms: How to avoid unfriendly neighbours. FEMS Microbiol. Rev. 2012, 36, 972–989. [Google Scholar] [CrossRef]
- Ica, T.; Caner, V.; Istanbullu, O.; Nguyen, H.D.; Ahmed, B.; Call, D.R.; Beyenal, H. Characterization of mono- and mixed-culture Campylobacter jejuni biofilm. Appl. Environ. Microbiol. 2012, 78, 1033–1038. [Google Scholar] [CrossRef]
- Giaouris, E.; Chorianopoulos, N.; Doulgeraki, A.; Nychas, G.J. Co-culture with Listeria monocytogenes within a dual-species biofilm community strongly increases resistance of Pseudomonas putida to benzalkonium chloride. PLoS ONE 2013, 8, e77276. [Google Scholar] [CrossRef]
- Jahid, I.K.; Han, N.R.; Srey, S.; Ha, S.-D. Competitive interactions inside mixed-culture biofilms of Salmonella Typhimurium and cultivable indigenous microorganisms on lettuce enhance microbial resistance of their sessile cells to ultraviolet C (UV-C) irradiation. Food Res. Int. 2014, 55, 445–454. [Google Scholar] [CrossRef]
- Elias, S.; Banin, E. Multi-species biofilms: Living with friendly neighbors. FEMS Microbiol. Rev. 2012, 36, 990–1004. [Google Scholar] [CrossRef]
- Thakur, M.; Asrani, R.K.; Patial, V. Listeria monocytogenes: A Food-Borne Pathogen. In Foodborne Diseases. Handbook of Food Bioengineering; Holban, A.A., Grumezescu, A.M., Eds.; Academic Press: London, UK, 2018; Volume 15, pp. 157–192. [Google Scholar] [CrossRef]
- European Food Safety Authority. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, 5500. [Google Scholar] [CrossRef]
- Liu, D. Identification, subtyping and virulence determination of Listeria monocytogenes, an important foodborne pathogen. J. Med. Microbiol. 2006, 55, 645–659. [Google Scholar] [CrossRef]
- Puga, C.H.; SanJose, C.; Orgaz, B. Biofilm development at low temperatures enhances Listeria monocytogenes resistance to chitosan. Food Contr. 2016, 65, 143–151. [Google Scholar] [CrossRef]
- Fetsch, A.; Contzen, M.; Hartelt, K.; Kleiser, A.; Maassen, S.; Rau, J.; Kraushaar, B.; Layer, F.; Strommenger, B. Staphylococcus aureus food-poisoning outbreak associated with the consumption of ice-cream. Int. J. Food Microbiol. 2014, 187, 1–6. [Google Scholar] [CrossRef]
- Naimi, T.; Wicklund, J.; Olsen, S.J.; Krause, G.; Wells, J.G.; Bartkus, J.M.; Boxrud, D.J.; Sullivan, M.; Kassenborg, H.; Besser, J.M.; et al. Concurrent outbreaks of Shigella sonnei and enterotoxigenic Escherichia coli infections associated with parsley: Implications for surveillance and control of foodborne illness. J. Food Prot. 2003, 66, 535–541. [Google Scholar] [CrossRef]
- Leriche, V.; Carpentier, B. Limitation of adhesion and growth of Listeria monocytogenes on stainless steel surfaces by Staphylococcus sciuri biofilms. J. Appl. Microbiol. 2000, 88, 594–605. [Google Scholar] [CrossRef]
- Carpentier, B.; Chassaing, D. Interactions in biofilms between Listeria monocytogenes and resident microorganisms from food industry premises. Int. J. Food Microbiol. 2004, 97, 111–122. [Google Scholar] [CrossRef]
- Schweinsberg, F.; Bürkle, V. Nitrite: A co-carcinogen? J. Cancer Res. Clin. Oncol. 1985, 109, 200–202. [Google Scholar] [CrossRef]
- Cammack, R.; Joannou, C.L.; Cui, X.Y.; Torres Martinez, C.; Maraj, S.R.; Hughes, M.N. Nitrite and nitrosyl compounds in food preservation. BBA-Bioenergetics 1999, 1411, 475–488. [Google Scholar] [CrossRef]
- Mariutti, L.R.B.; Nogueira, G.C.; Bragagnolo, N. Lipid and cholesterol oxidation in chicken meat are inhibited by sage but not by garlic. J. Food Sci. 2011, 76, C909–C915. [Google Scholar] [CrossRef]
- Schelz, Z.; Hohmann, J.; Molnar, J. Recent advances in research of antimicrobial effects of essential oils and plant derived compounds on bacteria. In Ethnomedicine: A Source of Complementary Therapeutics; Chattopadhyay, D., Ed.; Research Signpost: Kerala, India, 2010; pp. 179–201. [Google Scholar]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils–a review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Tajkarimi, M.M.; Ibrahim, S.A.; Cliver, D.O. Antimicrobial herb and spice compounds in food. Food Contr. 2010, 21, 1199–1218. [Google Scholar] [CrossRef]
- Sánchez, E.; García, S.; Heredia, N. Extracts of edible and medicinal plants damage membranes of Vibrio cholerae. Appl. Environ. Microbiol. 2010, 76, 6888–6894. [Google Scholar] [CrossRef]
- Chorianopoulos, N.G.; Giaouris, E.D.; Skandamis, P.N.; Haroutounian, S.A.; Nychas, G.J.E. Disinfectant test against monoculture and mixed-culture biofilms composed of technological, spoilage and pathogenic bacteria: Bactericidal effect of essential oil and hydrosol of Satureja thymbra and comparison with standard acid-base sanitizers. J. Appl. Microbiol. 2008, 104, 1586–1596. [Google Scholar] [CrossRef]
- Oliveira, M.M.M.; Brugnera, D.F.; Cardoso, M.G.; Alves, E.; Piccoli, R.H. Disinfectant action of Cymbopogon sp. essential oils in different phases of biofilm formation by Listeria monocytogenes on stainless steel surface. Food Contr. 2010, 21, 549–553. [Google Scholar] [CrossRef]
- Adukwu, E.C.; Allen, S.C.H.; Phillips, C.A. The anti-biofilm activity of lemongrass (Cymbopogon flexuosus) and grapefruit (Citrus paradisi) essential oils against five strains of Staphylococcus aureus. J. Appl. Microbiol. 2012, 113, 1217–1227. [Google Scholar] [CrossRef]
- Kerekes, E.B.; Deák, É.; Takó, M.; Tserennadmid, R.; Petkovits, T.; Vágvölgyi, C.; Krisch, J. Anti-biofilm forming and anti-quorum sensing activity of selected essential oils and their main components on food-related micro-organisms. J. Appl. Microbiol. 2013, 115, 933–942. [Google Scholar] [CrossRef]
- Kerekes, E.B.; Vidács, A.; Jenei Török, J.; Gömöri, C.; Petkovits, T.; Chandrashekaran, M.; Kadaikunnan, S.; Alharbi, N.S.; Vágvölgyi, C.; Krisch, J. Anti-listerial effect of selected essential oils and thymol. Acta Biol. Hung. 2016, 67, 333–343. [Google Scholar] [CrossRef]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Meth. 2008, 72, 157–165. [Google Scholar] [CrossRef]
- Gömöri, C.; Vidács, A.; Kerekes, E.B.; Nacsa-Farkas, E.; Böszörményi, A.; Vágvölgyi, C.; Krisch, J. Altered antimicrobial and anti-biofilm forming effect of thyme essential oil due to changes in composition. Nat. Prod. Commun. 2018, 13, 483–487. [Google Scholar] [CrossRef]
- Dorman, H.J.D.; Deans, S.G. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Almeida, C.; Azevedo, N.F.; Santos, S.; Keevil, C.W.; Vieira, M.J. Discriminating multi-species populations in biofilms with peptide nucleic acid fluorescence in situ hybridization (PNA FISH). PLoS ONE 2011, 6, e14786. [Google Scholar] [CrossRef]
- Millezi, F.M.; Pereira, M.O.; Batista, N.N.; Camargos, N.; Auad, I.; Cardoso, M.D.G.; Piccoli, R.H. Susceptibility of monospecies and dual-species biofilms of Staphylococcus aureus and Escherichia coli to essential oils. J. Food Saf. 2012, 32, 351–359. [Google Scholar] [CrossRef]
- Szczepanski, S.; Lipski, A. Essential oils show specific inhibiting effects on bacterial biofilm formation. Food Contr. 2014, 36, 224–229. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Wang, Y.; Jiang, P.; Quek, S. Antibacterial activity and mechanism of cinnamon essential oil against Escherichia coli and Staphylococcus aureus. Food Contr. 2016, 59, 282–289. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EOs and Components | MIC (mg/mL) | |||
|---|---|---|---|---|
| Escherichia coli | Listeria monocytogenes | Pseudomonas putida | Staphylococcus aureus | |
| Cinnamon | 0.25 | 1 | 0.25 | 0.4 |
| Marjoram | 0.5 1 | 4 2 | 2 1 | 3.2 |
| Thyme | 1.0 | 2 2 | 20 | 0.8 |
| Trans-cinnamaldehyde | 0.25 | 0.25 | 0.25 | 0.4 |
| Terpinene-4-ol | 2 1 | 4 | 4 1 | 3.2 |
| Thymol | 0.5 | 0.5 2 | 0.5 | 0.8 |
| Bacteria | Biofilm Formation (OD 590) 1 | ||||||
|---|---|---|---|---|---|---|---|
| Positive Control | Cinnamon | Marjoram | Thyme | Trans-cinnamaldehyde | Terpinene-4-ol | Thymol | |
| Escherichia coli | 0.68 ± 0.17a | 0.77 ± 0.13a | 0.72 ± 0.10a 2 | 0.34 ± 0.02b | 0.32 ± 0.08b | 0.44 ± 0.11b 2 | 0.14 ± 0.02c |
| Listeria monocytogenes | 1.34 ± 0.14a | 0.14 ± 0.02c | 0.28 ± 0.04b | 0.30 ± 0.04b | 0.17 ± 0.02c | 0.34 ± 0.34b | 0.53 ± 0.08b |
| Pseudomonas putida | 1.52 ± 0.4a | 0.48 ± 0.05d | 0.08 ± 0.00b 2 | 0.51 ± 0.05d | 0.70 ± 0.05d | 1.09 ± 0.10b 2 | 0.43 ± 0.07d |
| Staphylococcus aureus | 2.36 ± 0.07a | 1.31 ± 0.30b | 1.18 ± 0.20b | 1.22 ± 0.16b | 1.19 ± 0.17b | 1.60 ± 0.30b | 1.37 ± 0.41b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerekes, E.B.; Vidács, A.; Takó, M.; Petkovits, T.; Vágvölgyi, C.; Horváth, G.; Balázs, V.L.; Krisch, J. Anti-Biofilm Effect of Selected Essential Oils and Main Components on Mono- and Polymicrobic Bacterial Cultures. Microorganisms 2019, 7, 345. https://doi.org/10.3390/microorganisms7090345
Kerekes EB, Vidács A, Takó M, Petkovits T, Vágvölgyi C, Horváth G, Balázs VL, Krisch J. Anti-Biofilm Effect of Selected Essential Oils and Main Components on Mono- and Polymicrobic Bacterial Cultures. Microorganisms. 2019; 7(9):345. https://doi.org/10.3390/microorganisms7090345
Chicago/Turabian StyleKerekes, Erika Beáta, Anita Vidács, Miklós Takó, Tamás Petkovits, Csaba Vágvölgyi, Györgyi Horváth, Viktória Lilla Balázs, and Judit Krisch. 2019. "Anti-Biofilm Effect of Selected Essential Oils and Main Components on Mono- and Polymicrobic Bacterial Cultures" Microorganisms 7, no. 9: 345. https://doi.org/10.3390/microorganisms7090345
APA StyleKerekes, E. B., Vidács, A., Takó, M., Petkovits, T., Vágvölgyi, C., Horváth, G., Balázs, V. L., & Krisch, J. (2019). Anti-Biofilm Effect of Selected Essential Oils and Main Components on Mono- and Polymicrobic Bacterial Cultures. Microorganisms, 7(9), 345. https://doi.org/10.3390/microorganisms7090345