Effects of Acute and Chronic Exposure to Residual Level Erythromycin on Human Intestinal Epithelium Cell Permeability and Cytotoxicity

 ,
, 
Abstract
1. Introduction
2. Material and Method
2.1. Cell Culture
2.2. Transwell Model and Transepithelial Electrical Resistance (TER) Measurement
2.3. Acute Exposure of Erythromycin
2.4. Chronic Exposure of Erythromycin
2.5. Cytotoxicity Assay
2.6. qPCR Analysis of Genes Involved in the Cellular Integrity
3. Results
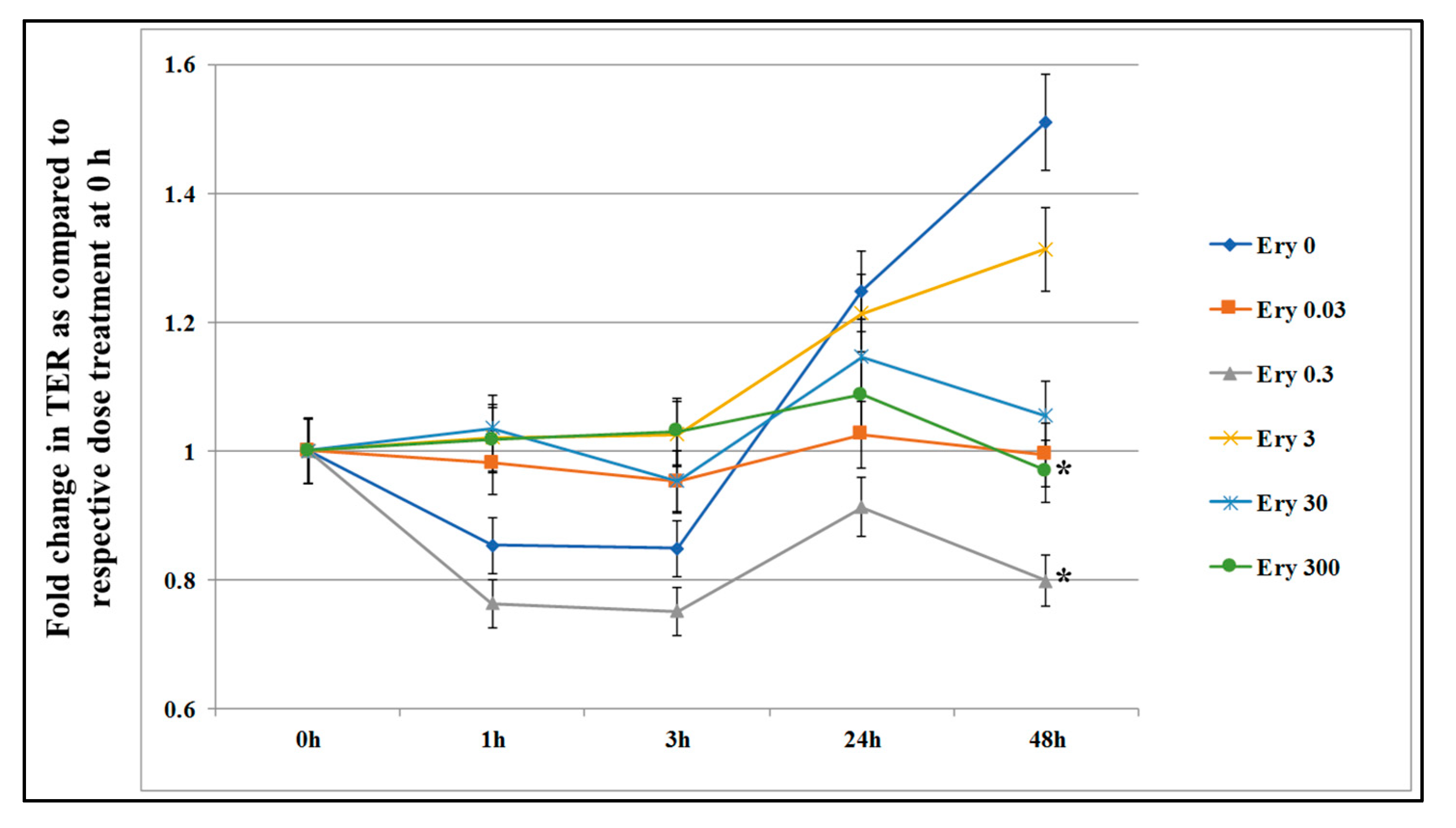
3.1. Effect of Acute Exposure on Epithelial Cells Permeability
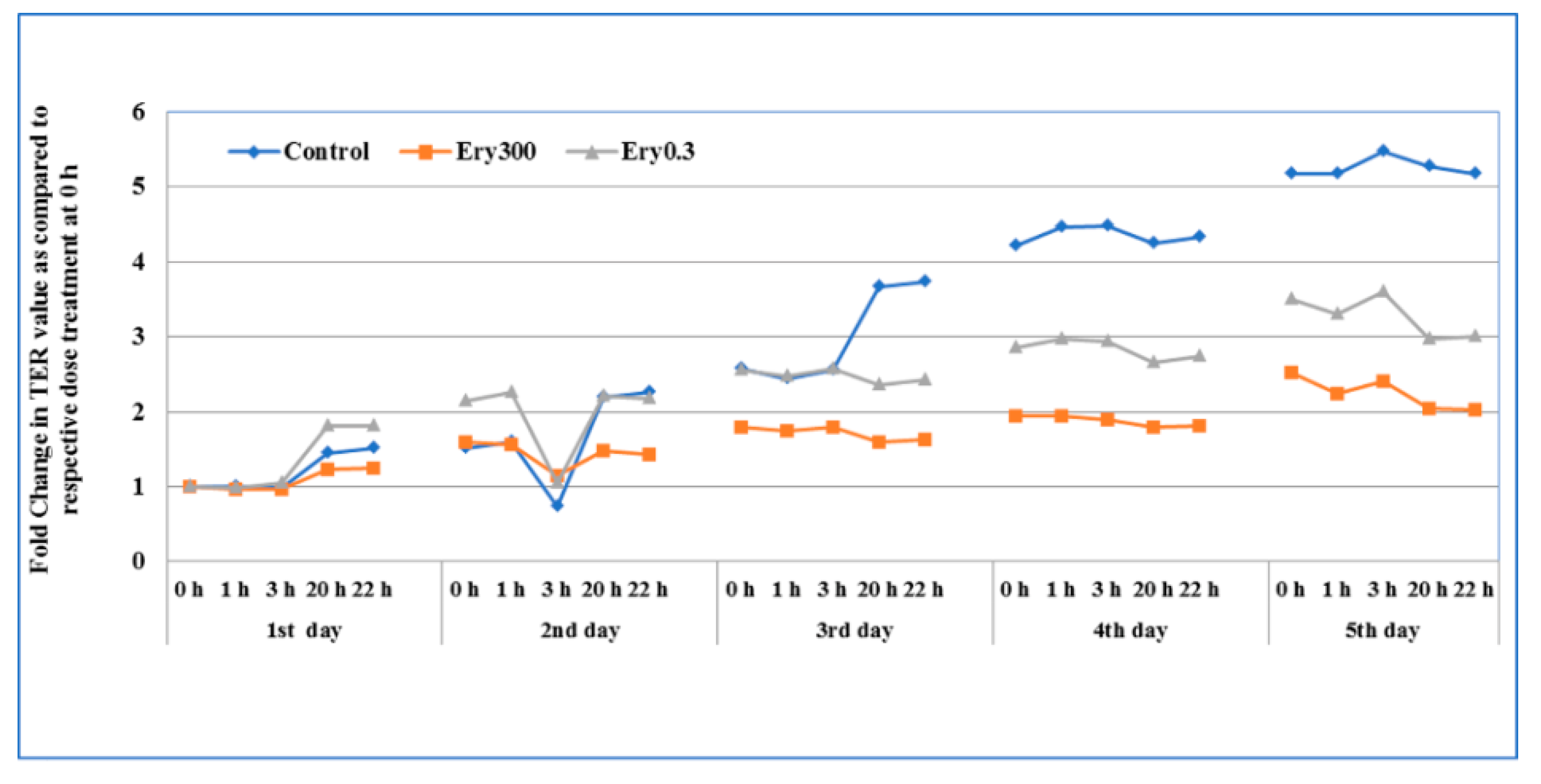
3.2. Effect of Chronic Exposure on Epithelial Permeability
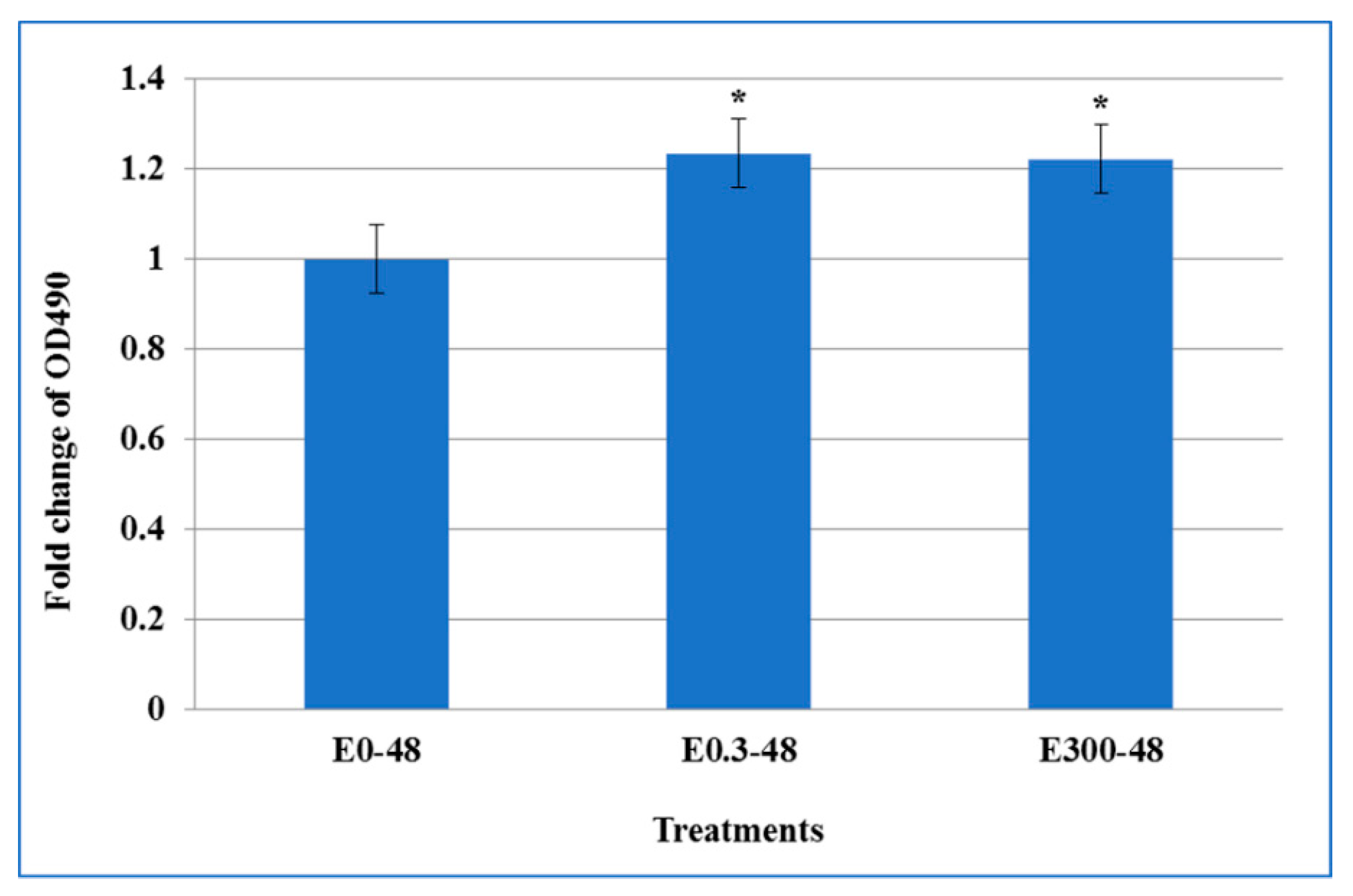
3.3. Effect of Acute Erythromycin Exposure on Cell Cytotoxicity
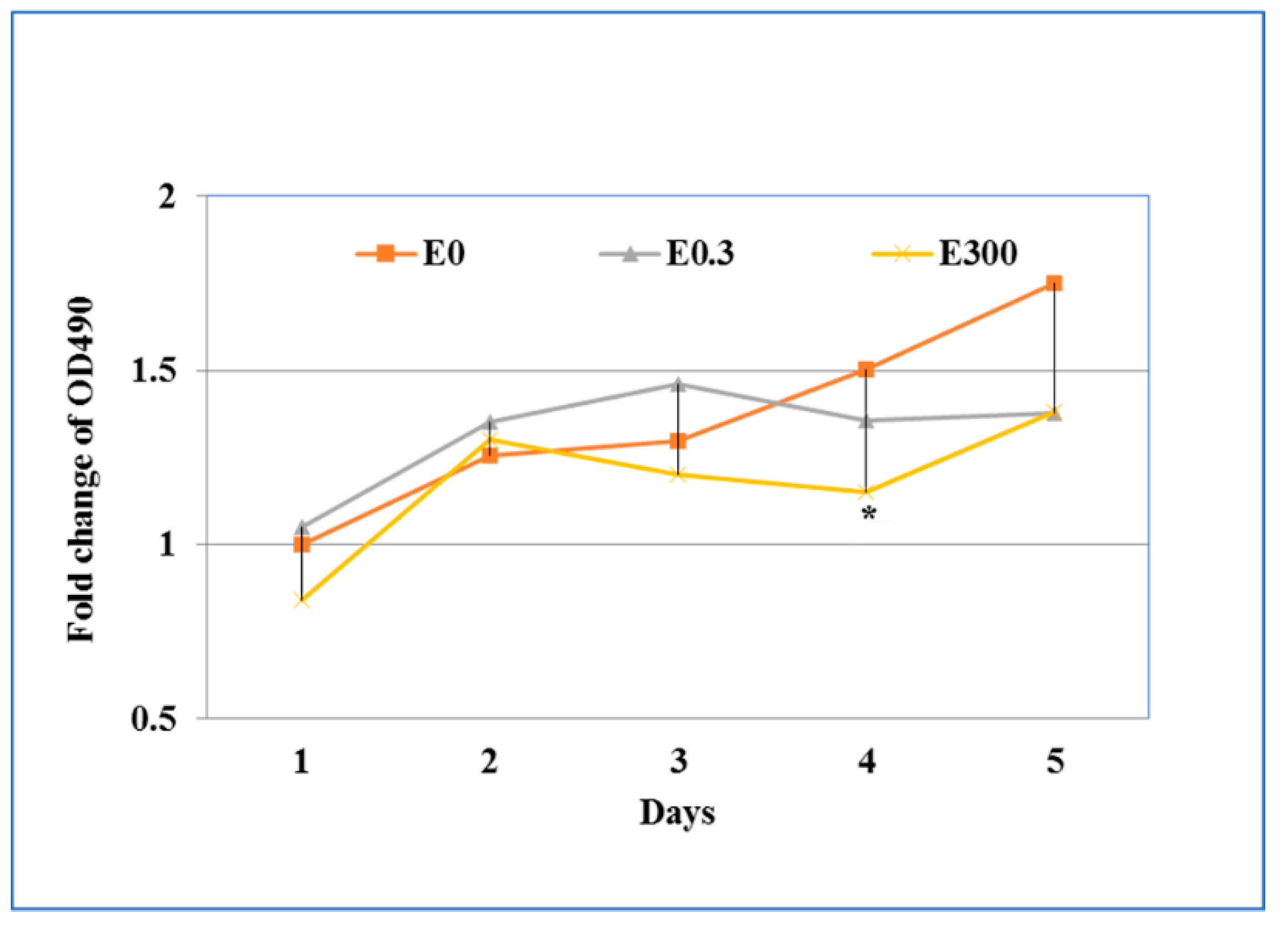
3.4. Effect of Chronic Erythromycin Exposure on Cytotoxicity
3.5. Impact of Acute Exposure on Expression of Human Cell Junction Genes
3.6. Impact of Chronic Exposure of Erythromycin on the Expression of Human Cell Junction Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kibwage, I.O.; Hoogmartens, J.; Roets, E.; Vanderhaeghe, H.; Verbist, L.; Dubost, M.; Pascal, C.; Petitjean, P.; Levol, G. Antibacterial activities of erythromycins a, b, c, and d and some of their derivatives. Antimicrob. Agents Chemother. 1985, 28, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, S.; Nagate, T.; Sugita, K.; Ono, T.; Numata, K.; Miyachi, J.; Misawa, Y.; Yamada, K.; Omura, S. Chemical modification of erythromycins. III. In vitro and in vivo antibacterial activities of new semisynthetic 6-o-methylerythromycins a, te-031 (clarithromycin) and te-032. J. Antibiot. 1990, 43, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Cramer, C.L.; Patterson, A.; Alchakaki, A.; Soubani, A.O. Immunomodulatory indications of azithromycin in respiratory disease: A concise review for the clinician. Postgrad. Med. 2017, 129, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Parnham, M.J.; Erakovic Haber, V.; Giamarellos-Bourboulis, E.J.; Perletti, G.; Verleden, G.M.; Vos, R. Azithromycin: Mechanisms of action and their relevance for clinical applications. Pharmacol. Ther. 2014, 143, 225–245. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Heinze, T.M.; Kim, S.J.; Cerniglia, C.E. Adsorption and clay-catalyzed degradation of erythromycin a on homoionic clays. J. Environ. Qual. 2004, 33, 257–264. [Google Scholar] [CrossRef] [PubMed]
- JECFA: Joint FAO/WHO Expert Committee On Food Additives. Evaluation of Certain Veterinary Drug Residues in Food: Sixty-Sixth Report of the Joint FAO/WHO Expert Committee on Food Additives; WHO Technical Report Series; WHO: Geneva, Switzerland, 2006; pp. 1–80. [Google Scholar]
- EMEA. Committee for veterinary medicinal products erythromycin, summary report (2). In The European Agency for the Evaluation of Medicinal Products Veterinary Medicines and Information Technology Unit; European Agency for the Evaluation of Medicinal Products: London, UK, 2000. [Google Scholar]
- Ge, B.; Domesle, K.J.; Yang, Q.; Young, S.R.; Rice-Trujillo, C.L.; Bodeis Jones, S.M.; Gaines, S.A.; Keller, M.W.; Li, X.; Pineiro, S.A.; et al. Effects of low concentrations of erythromycin, penicillin, and virginiamycin on bacterial resistance development in vitro. Sci. Rep. 2017, 7, 11017. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, M.; Parkman, H.P.; Shafi, M.A.; Abell, T.L.; Gerson, L.; American College of Gastroenterology. Clinical guideline: Management of gastroparesis. Am. J. Gastroenterol. 2013, 108, 18–37. [Google Scholar] [CrossRef]
- Lebel, M. Pharmacokinetic properties of clarithromycin: A comparison with erythromycin and azithromycin. Can. J. Infect. Dis. 1993, 4, 148–152. [Google Scholar] [CrossRef]
- Krasniqi, S.; Matzneller, P.; Kinzig, M.; Sorgel, F.; Huttner, S.; Lackner, E.; Muller, M.; Zeitlinger, M. Blood, tissue, and intracellular concentrations of erythromycin and its metabolite anhydroerythromycin during and after therapy. Antimicrob. Agents Chemother. 2012, 56, 1059–1064. [Google Scholar] [CrossRef]
- Stupans, I.; Sansom, L.N. The inhibition of drug oxidation by anhydroerythromycin, an acid degradation product of erythromycin. Biochem. Pharmacol. 1991, 42, 2085–2090. [Google Scholar] [CrossRef]
- Mate, L.; Virkel, G.; Lifschitz, A.; Sallovitz, J.; Ballent, M.; Lanusse, C. Phase 1 and phase 2 metabolic activities along the small intestine in adult male sheep. J. Vet. Pharmacol. Ther. 2010, 33, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Pineau, T.; Galtier, P.; Bonfils, C.; Derancourt, J.; Maurel, P. Purification of a sheep liver cytochrome p-450 from the p450iiia gene subfamily. Its contribution to the n-dealkylation of veterinary drugs. Biochem. Pharmacol. 1990, 39, 901–909. [Google Scholar] [CrossRef]
- Sato, H.; Zhang, L.S.; Martinez, K.; Chang, E.B.; Yang, Q.; Wang, F.; Howles, P.N.; Hokari, R.; Miura, S.; Tso, P. Antibiotics suppress activation of intestinal mucosal mast cells and reduce dietary lipid absorption in sprague-dawley rats. Gastroenterology 2016, 151, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Gokulan, K.; Cerniglia, C.E.; Thomas, C.; Pineiro, S.A.; Khare, S. Effects of residual levels of tetracycline on the barrier functions of human intestinal epithelial cells. Food Chem. Toxicol. 2017, 109, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Odenwald, M.A.; Turner, J.R. Intestinal permeability defects: Is it time to treat? Clin. Gastroenterol. Hepatol. 2013, 11, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Barreau, F.; Hugot, J.P. Intestinal barrier dysfunction triggered by invasive bacteria. Curr. Opin. Microbiol. 2014, 17, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Kerr, C.A.; Grice, D.M.; Tran, C.D.; Bauer, D.C.; Li, D.; Hendry, P.; Hannan, G.N. Early life events influence whole-of-life metabolic health via gut microflora and gut permeability. Crit. Rev. Microbiol. 2015, 41, 326–340. [Google Scholar] [CrossRef]
- Thevaranjan, N.; Puchta, A.; Schulz, C.; Naidoo, A.; Szamosi, J.C.; Verschoor, C.P.; Loukov, D.; Schenck, L.P.; Jury, J.; Foley, K.P.; et al. Age-Associated Microbial Dysbiosis Promotes Intestinal Permeability, Systemic Inflammation, and Macrophage Dysfunction. Cell Host Microbe 2017, 21, 455–466. [Google Scholar] [CrossRef]
- Odenwald, M.A.; Turner, J.R. The intestinal epithelial barrier: A therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef]
- Caruso, R.; Fina, D.; Paoluzi, O.A.; Del Vecchio Blanco, G.; Stolfi, C.; Rizzo, A.; Caprioli, F.; Sarra, M.; Andrei, F.; Fantini, M.C.; et al. Il-23-mediated regulation of il-17 production in helicobacter pylori-infected gastric mucosa. Eur. J. Immunol. 2008, 38, 470–478. [Google Scholar] [CrossRef]
- Carman, R.J.; Simon, M.A.; Petzold, H.E., 3rd; Wimmer, R.F.; Batra, M.R.; Fernandez, A.H.; Miller, M.A.; Bartholomew, M. Antibiotics in the human food chain: Establishing no effect levels of tetracycline, neomycin, and erythromycin using a chemostat model of the human colonic microflora. Regul. Toxicol. Pharmacol. 2005, 43, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Youakim, A.; Ahdieh, M. Interferon-gamma decreases barrier function in t84 cells by reducing zo-1 levels and disrupting apical actin. Am. J. Physiol. 1999, 276, G1279–G1288. [Google Scholar] [PubMed]
- Bruewer, M.; Luegering, A.; Kucharzik, T.; Parkos, C.A.; Madara, J.L.; Hopkins, A.M.; Nusrat, A. Proinflammatory cytokines disrupt epithelial barrier function by apoptosis-independent mechanisms. J. Immunol. 2003, 171, 6164–6172. [Google Scholar] [CrossRef] [PubMed]
- Khare, S.; Lawhon, S.D.; Drake, K.L.; Nunes, J.E.; Figueiredo, J.F.; Rossetti, C.A.; Gull, T.; Everts, R.E.; Lewin, H.A.; Galindo, C.L.; et al. Systems biology analysis of gene expression during in vivo mycobacterium avium paratuberculosis enteric colonization reveals role for immune tolerance. PLoS ONE 2012, 7, e42127. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.M.; Gokulan, K.; Cerniglia, C.E.; Khare, S. Size and dose dependent effects of silver nanoparticle exposure on intestinal permeability in an in vitro model of the human gut epithelium. J. Nanobiotechnol. 2016, 14, 62. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time pcr data by the comparative ct method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Selsted, M.E.; Miller, S.I.; Henschen, A.H.; Ouellette, A.J. Enteric defensins: antibiotic peptide components of intestinal host defense. J. Cell Biol. 1992, 118, 929–936. [Google Scholar] [CrossRef]
- Pastorelli, L.; De Salvo, C.; Mercado, J.R.; Vecchi, M.; Pizarro, T.T. Central role of the gut epithelial barrier in the pathogenesis of chronic intestinal inflammation: Lessons learned from animal models and human genetics. Front. Immunol. 2013, 4, 280. [Google Scholar] [CrossRef]
- Craft, K.M.; Gaddy, J.A.; Townsend, S.D. Human milk oligosaccharides sensitize group b streptococcus to clindamycin, erythromycin, gentamycin, and minocycline on a strain specific basis. ACS Chem. Biol. 2018, 13, 2020–2026. [Google Scholar] [CrossRef]
- Takahashi, E.; Indalao, I.L.; Sawabuchi, T.; Mizuno, K.; Sakai, S.; Kimoto, T.; Kim, H.; Kido, H. Clarithromycin suppresses induction of monocyte chemoattractant protein-1 and matrix metalloproteinase-9 and improves pathological changes in the lungs and heart of mice infected with influenza a virus. Comp. Immunol. Microbiol. Infect. Dis. 2018, 56, 6–13. [Google Scholar] [CrossRef]
- Zhou, P.; Chen, Y.; Lu, Q.; Qin, H.; Ou, H.; He, B.; Ye, J. Cellular metabolism network of bacillus thuringiensis related to erythromycin stress and degradation. Ecotoxicol. Environ. Saf. 2018, 160, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Liu, D.; Singh, A.K.; Drolia, R.; Bai, X.; Tenguria, S.; Bhunia, A.K. Tunicamycin mediated inhibition of wall teichoic acid affects staphylococcus aureus and listeria monocytogenes cell morphology, biofilm formation and virulence. Front. Microbiol. 2018, 9, 1352. [Google Scholar] [CrossRef] [PubMed]
- Otterson, M.F.; Sarna, S.K. Gastrointestinal motor effects of erythromycin. Am. J. Physiol. 1990, 259, G355–G363. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, R.; Miyayama, T.; Mitsui, T.; Akiba, Y.; Higashida, A.; Takata, S.; Kawanishi, T.; Aso, Y.; Itoh, Z.; Omura, S. Nonlinear intestinal pharmacokinetics of mitemcinal, the first acid-resistant non-peptide motilin receptor agonist, in rats. Xenobiotica 2007, 37, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Galligan, J.J.; Vanner, S. Basic and clinical pharmacology of new motility promoting agents. Neurogastroenterol. Motil. 2005, 17, 643–653. [Google Scholar] [CrossRef] [PubMed]
- Yamaura, Y.; Chapron, B.D.; Wang, Z.; Himmelfarb, J.; Thummel, K.E. Functional comparison of human colonic carcinoma cell lines and primary small intestinal epithelial cells for investigations of intestinal drug permeability and first-pass metabolism. Drug Metab. Dispos. 2016, 44, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Juuti-Uusitalo, K.M.; Kaukinen, K.; Maki, M.; Tuimala, J.; Kainulainen, H. Gene expression in tgfbeta-induced epithelial cell differentiation in a three-dimensional intestinal epithelial cell differentiation model. BMC Genom. 2006, 7, 279. [Google Scholar] [CrossRef] [PubMed]
- Bourgine, J.; Billaut-Laden, I.; Happillon, M.; Lo-Guidice, J.M.; Maunoury, V.; Imbenotte, M.; Broly, F. Gene expression profiling of systems involved in the metabolism and the disposition of xenobiotics: Comparison between human intestinal biopsy samples and colon cell lines. Drug Metab. Dispos. 2012, 40, 694–705. [Google Scholar] [CrossRef]
- Navarro, H.; Arruebo, M.P.; Sorribas, V.; Alcalde, A.I. Effect of erythromycin on l-threonine transport in rabbit jejunum in vitro. J. Vet. Pharmacol. Ther. 1992, 15, 188–193. [Google Scholar] [CrossRef]
- Alcalde, A.I.; Navarro, H.; Sorribas, V.; Marco, R. Study of the action of intramuscularly administered erythromycin on the l-threonine transport and the digestive enzymatic activity in rabbit jejunum. Life Sci. 1996, 59, 821–833. [Google Scholar] [CrossRef]
- Navarro, H.; Arruebo, M.P.; Alcalde, A.I.; Sorribas, V. Effect of erythromycin on d-galactose absorption and sucrase activity in rabbit jejunum. Can. J. Physiol. Pharmacol. 1993, 71, 191–194. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Liang, Q.; Lian, T.; Wu, Q.; Gaur, U.; Li, D.; Yang, D.; Mao, X.; Jin, Z.; Li, Y.; et al. Rapamycin preserves gut homeostasis during drosophila aging. Oncotarget 2015, 6, 35274–35283. [Google Scholar] [CrossRef] [PubMed]
- Volkert, G.; Jahn, A.; Dinkel, C.; Fahlbusch, F.; Zurn, C.; Hilgers, K.F.; Rascher, W.; Hartner, A.; Marek, I. Contribution of the alpha8 integrin chain to the expression of extracellular matrix components. Cell Commun. Adhes. 2014, 21, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Kaiserlian, D.; Rigal, D.; Abello, J.; Revillard, J.P. Expression, function and regulation of the intercellular adhesion molecule-1 (icam-1) on human intestinal epithelial cell lines. Eur. J. Immunol. 1991, 21, 2415–2421. [Google Scholar] [CrossRef] [PubMed]
- Sumagin, R.; Parkos, C.A. Epithelial adhesion molecules and the regulation of intestinal homeostasis during neutrophil transepithelial migration. Tissue Barriers 2015, 3, e969100. [Google Scholar] [CrossRef]
- Sumagin, R.; Robin, A.Z.; Nusrat, A.; Parkos, C.A. Transmigrated neutrophils in the intestinal lumen engage icam-1 to regulate the epithelial barrier and neutrophil recruitment. Mucosal Immunol. 2014, 7, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Sumagin, R.; Brazil, J.C.; Nava, P.; Nishio, H.; Alam, A.; Luissint, A.C.; Weber, D.A.; Neish, A.S.; Nusrat, A.; Parkos, C.A. Neutrophil interactions with epithelial-expressed icam-1 enhances intestinal mucosal wound healing. Mucosal Immunol. 2016, 9, 1151–1162. [Google Scholar] [CrossRef]
- Porter, J.C.; Hall, A. Epithelial icam-1 and icam-2 regulate the egression of human t cells across the bronchial epithelium. FASEB J. 2009, 23, 492–502. [Google Scholar] [CrossRef]
- Martin-Padura, I.; Lostaglio, S.; Schneemann, M.; Williams, L.; Romano, M.; Fruscella, P.; Panzeri, C.; Stoppacciaro, A.; Ruco, L.; Villa, A.; et al. Junctional adhesion molecule, a novel member of the immunoglobulin superfamily that distributes at intercellular junctions and modulates monocyte transmigration. J. Cell Biol. 1998, 142, 117–127. [Google Scholar] [CrossRef]
- Arrate, M.P.; Rodriguez, J.M.; Tran, T.M.; Brock, T.A.; Cunningham, S.A. Cloning of human junctional adhesion molecule 3 (jam3) and its identification as the jam2 counter-receptor. J. Biol. Chem. 2001, 276, 45826–45832. [Google Scholar] [CrossRef]
- Santoso, S.; Sachs, U.J.; Kroll, H.; Linder, M.; Ruf, A.; Preissner, K.T.; Chavakis, T. The junctional adhesion molecule 3 (jam-3) on human platelets is a counterreceptor for the leukocyte integrin mac-1. J. Exp. Med. 2002, 196, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Chavakis, T.; Keiper, T.; Matz-Westphal, R.; Hersemeyer, K.; Sachs, U.J.; Nawroth, P.P.; Preissner, K.T.; Santoso, S. The junctional adhesion molecule-c promotes neutrophil transendothelial migration in vitro and in vivo. J. Biol. Chem. 2004, 279, 55602–55608. [Google Scholar] [CrossRef] [PubMed]
- Zen, K.; Babbin, B.A.; Liu, Y.; Whelan, J.B.; Nusrat, A.; Parkos, C.A. Jam-c is a component of desmosomes and a ligand for cd11b/cd18-mediated neutrophil transepithelial migration. Mol. Biol. Cell 2004, 15, 3926–3937. [Google Scholar] [CrossRef] [PubMed]
- Aurrand-Lions, M.; Lamagna, C.; Dangerfield, J.P.; Wang, S.; Herrera, P.; Nourshargh, S.; Imhof, B.A. Junctional adhesion molecule-c regulates the early influx of leukocytes into tissues during inflammation. J. Immunol. 2005, 174, 6406–6415. [Google Scholar] [CrossRef] [PubMed]
- Mandicourt, G.; Iden, S.; Ebnet, K.; Aurrand-Lions, M.; Imhof, B.A. Jam-c regulates tight junctions and integrin-mediated cell adhesion and migration. J. Biol. Chem. 2007, 282, 1830–1837. [Google Scholar] [CrossRef] [PubMed]
- Ebnet, K.; Aurrand-Lions, M.; Kuhn, A.; Kiefer, F.; Butz, S.; Zander, K.; Meyer zu Brickwedde, M.K.; Suzuki, A.; Imhof, B.A.; Vestweber, D. The junctional adhesion molecule (jam) family members jam-2 and jam-3 associate with the cell polarity protein par-3: A possible role for jams in endothelial cell polarity. J. Cell Sci. 2003, 116, 3879–3891. [Google Scholar] [CrossRef]
- Muller, U.; Bossy, B.; Venstrom, K.; Reichardt, L.F. Integrin alpha 8 beta 1 promotes attachment, cell spreading, and neurite outgrowth on fibronectin. Mol. Biol. Cell 1995, 6, 433–448. [Google Scholar] [CrossRef]
- Denda, S.; Muller, U.; Crossin, K.L.; Erickson, H.P.; Reichardt, L.F. Utilization of a soluble integrin-alkaline phosphatase chimera to characterize integrin alpha 8 beta 1 receptor interactions with tenascin: Murine alpha 8 beta 1 binds to the rgd site in tenascin-c fragments, but not to native tenascin-c. Biochemistry 1998, 37, 5464–5474. [Google Scholar] [CrossRef][Green Version]
- Brandenberger, R.; Schmidt, A.; Linton, J.; Wang, D.; Backus, C.; Denda, S.; Muller, U.; Reichardt, L.F. Identification and characterization of a novel extracellular matrix protein nephronectin that is associated with integrin alpha8beta1 in the embryonic kidney. J. Cell Biol. 2001, 154, 447–458. [Google Scholar] [CrossRef]
- Bieritz, B.; Spessotto, P.; Colombatti, A.; Jahn, A.; Prols, F.; Hartner, A. Role of alpha8 integrin in mesangial cell adhesion, migration, and proliferation. Kidney Int. 2003, 64, 119–127. [Google Scholar] [CrossRef]
- Hartner, A.; Cordasic, N.; Klanke, B.; Muller, U.; Sterzel, R.B.; Hilgers, K.F. The alpha8 integrin chain affords mechanical stability to the glomerular capillary tuft in hypertensive glomerular disease. Am. J. Pathol. 2002, 160, 861–867. [Google Scholar] [CrossRef]
- Zargham, R.; Thibault, G. Alpha8beta1 integrin expression in the rat carotid artery: Involvement in smooth muscle cell migration and neointima formation. Cardiovasc. Res. 2005, 65, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Farias, E.; Lu, M.; Li, X.; Schnapp, L.M. Integrin alpha8beta1-fibronectin interactions promote cell survival via pi3 kinase pathway. Biochem. Biophys. Res. Commun. 2005, 329, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Zargham, R.; Wamhoff, B.R.; Thibault, G. Rna interference targeting alpha8 integrin attenuates smooth muscle cell growth. FEBS Lett. 2007, 581, 939–943. [Google Scholar] [CrossRef] [PubMed]
- Hartner, A.; Menendez-Castro, C.; Cordasic, N.; Marek, I.; Volkert, G.; Klanke, B.; Rascher, W.; Hilgers, K.F. Tubulointerstitial de novo expression of the alpha8 integrin chain in a rodent model of renal fibrosis—A potential target for anti-fibrotic therapy? PLoS ONE 2012, 7, e48362. [Google Scholar] [CrossRef] [PubMed]
- Marek, I.; Lichtneger, T.; Cordasic, N.; Hilgers, K.F.; Volkert, G.; Fahlbusch, F.; Rascher, W.; Hartner, A.; Menendez-Castro, C. Alpha8 integrin (itga8) signalling attenuates chronic renal interstitial fibrosis by reducing fibroblast activation, not by interfering with regulation of cell turnover. PLoS ONE 2016, 11, e0150471. [Google Scholar] [CrossRef] [PubMed]
- Kanoh, S.; Rubin, B.K. Mechanisms of action and clinical application of macrolides as immunomodulatory medications. Clin. Microbiol. Rev. 2010, 23, 590–615. [Google Scholar] [CrossRef]
- Vestbo, J.; Anderson, W.; Coxson, H.O.; Crim, C.; Dawber, F.; Edwards, L.; Hagan, G.; Knobil, K.; Lomas, D.A.; MacNee, W.; et al. Evaluation of copd longitudinally to identify predictive surrogate end-points (eclipse). Eur. Respir. J. 2008, 31, 869–873. [Google Scholar] [CrossRef]
- Culic, O.; Erakovic, V.; Cepelak, I.; Barisic, K.; Brajsa, K.; Ferencic, Z.; Galovic, R.; Glojnaric, I.; Manojlovic, Z.; Munic, V.; et al. Azithromycin modulates neutrophil function and circulating inflammatory mediators in healthy human subjects. Eur. J. Pharmacol. 2002, 450, 277–289. [Google Scholar] [CrossRef]
- Lin, S.J.; Kuo, M.L.; Hsiao, H.S.; Lee, P.T. Azithromycin modulates immune response of human monocyte-derived dendritic cells and cd4+ t cells. Int. Immunopharmacol. 2016, 40, 318–326. [Google Scholar] [CrossRef]
- Ratzinger, F.; Haslacher, H.; Poeppl, W.; Hoermann, G.; Kovarik, J.J.; Jutz, S.; Steinberger, P.; Burgmann, H.; Pickl, W.F.; Schmetterer, K.G. Azithromycin suppresses cd4(+) t-cell activation by direct modulation of mtor activity. Sci. Rep. 2014, 4, 7438. [Google Scholar] [CrossRef] [PubMed]
- Uriarte, S.M.; Molestina, R.E.; Miller, R.D.; Bernabo, J.; Farinati, A.; Eiguchi, K.; Ramirez, J.A.; Summersgill, J.T. Effect of macrolide antibiotics on human endothelial cells activated by chlamydia pneumoniae infection and tumor necrosis factor-alpha. J. Infect. Dis. 2002, 185, 1631–1636. [Google Scholar] [CrossRef][Green Version]
- Choi, Y.; Koh, S.J.; Lee, H.S.; Kim, J.W.; Gwan Kim, B.; Lee, K.L.; Kim, J.S. Roxithromycin inhibits nuclear factor kappab signaling and endoplasmic reticulum stress in intestinal epithelial cells and ameliorates experimental colitis in mice. Exp. Biol. Med. 2015, 240, 1664–1671. [Google Scholar] [CrossRef] [PubMed]
- Amsden, G.W. Anti-inflammatory effects of macrolides—An underappreciated benefit in the treatment of community-acquired respiratory tract infections and chronic inflammatory pulmonary conditions? J. Antimicrob. Chemother. 2005, 55, 10–21. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Desaki, M.; Okazaki, H.; Sunazuka, T.; Omura, S.; Yamamoto, K.; Takizawa, H. Molecular mechanisms of anti-inflammatory action of erythromycin in human bronchial epithelial cells: Possible role in the signaling pathway that regulates nuclear factor-kappab activation. Antimicrob. Agents Chemother. 2004, 48, 1581–1585. [Google Scholar] [CrossRef] [PubMed]
- Sevilla-Sanchez, D.; Soy-Muner, D.; Soler-Porcar, N. Usefulness of macrolides as anti-inflammatories in respiratory diseases. Arch. Bronconeumol. 2010, 46, 244–254. [Google Scholar] [CrossRef]
- Erythromycin. In Drugs and Lactation Database (Lactmed); Bethesda: Rockville, MD, USA, 2006.
- Knowles, J.A. Effects on the infant of drug therapy in nursing mothers. Drug Ther. N. Y. 1973, 3, 57–59. [Google Scholar]
- Matsuda, S. Transfer of antibiotics into maternal milk. Biol. Res. Pregnancy Perinatol. 1984, 5, 57–60. [Google Scholar]
- Zhang, Y.; Zhang, Q.; Xu, Z. Tissue and body fluid distribution of antibacterial agents in pregnant and lactating women. Zhonghua Fu Chan Ke Za Zhi 1997, 32, 288–292. [Google Scholar]
- Wilson, I.D.; Nicholson, J.K. The role of gut microbiota in drug response. Curr. Pharm. Des. 2009, 15, 1519–1523. [Google Scholar] [CrossRef]
- Sorensen, H.T.; Skriver, M.V.; Pedersen, L.; Larsen, H.; Ebbesen, F.; Schonheyder, H.C. Risk of infantile hypertrophic pyloric stenosis after maternal postnatal use of macrolides. Scand. J. Infect. Dis. 2003, 35, 104–106. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Single Exposure for 48 h | |||||
|---|---|---|---|---|---|
| Gene | Function | Ery 0.3/Ery 0 | Ery 0.3/Ery 0 | Ery 300/Ery 0 | Ery 300/Ery0 |
| Fold Change | p Value | Fold Change | p Value | ||
| Tight junction | |||||
| ICAM1 | Intercellular adhesion molecule 1 | 2.37 | 0.000636 | 2.05 | 0.016607 |
| JAM3 | Junctional adhesion molecule 3 | 2.13 | 0.062148 | 2.65 | 0.040630 |
| CLDN10 | Claudin 10 | –7.4 | 0.373085 | –5 | 0.378114 |
| CLDN11 | Claudin 11 | 3.68 | 0.944793 | 2.85 | 0.999572 |
| CLDN14 | Claudin 14 | –1.55 | 0.592628 | 3.83 | 0.299305 |
| CLDN17 | Claudin 17 | –2.74 | 0.452451 | 6.44 | 0.369806 |
| CLDN18 | Claudin 18 | 2.02 | 0.402447 | –1.57 | 0.633391 |
| CLDN19 | Claudin 19 | –2.32 | 0.728604 | –3.51 | 0.727239 |
| CLDN6 | Claudin 6 | 1.27 | 0.465959 | 2.22 | 0.264438 |
| Focal adhesion | |||||
| CAV1 | Caveolin 1, caveolae protein | –2.94 | 0.164653 | –1.32 | 0.390713 |
| CAV3 | Caveolin 3 | 14.52 | 0.120646 | 1.15 | 0.556335 |
| ITGA4 | Integrin, alpha 4 | 5.07 | 0.385157 | –1.13 | 0.841795 |
| ITGA7 | Integrin, alpha 7 | 1.65 | 0.618012 | 2.2 | 0.367128 |
| ITGA8 | Integrin, alpha 8 | 2.76 | 0.382339 | 175.02 | 0.024344 |
| ITGAM | Integrin, alpha M | –1.06 | 0.902768 | 3.21 | 0.308094 |
| Gap junction | |||||
| GJA1 | Gap junction protein, alpha 1 | –1.2 | 0.795080 | –5.32 | 0.055363 |
| GJA3 | Gap junction protein, alpha 3 | –1.33 | 0.417673 | 3.53 | 0.368254 |
| GJA5 | Gap junction protein, alpha 5 | 2.93 | 0.364299 | 12.55 | 0.216151 |
| GJB4 | Gap junction protein, beta 4 | 1.65 | 0.342500 | 2.01 | 0.286874 |
| GJB5 | Gap junction protein, beta 5 | 1.32 | 0.365400 | 2.28 | 0.286658 |
| GJB6 | Gap junction protein, beta 6 | –1.28 | 0.092314 | 1.67 | 0.420076 |
| GJC3 | Gap junction protein, epsilon 1 | 1.95 | 0.367178 | –25.08 | 0.263848 |
| Adherens junction and desmosomes genes | |||||
| DSC1 | Desmocollin 1 | –46.06 | 0.144780 | –31.11 | 0.146747 |
| DSG3 | Desmoglein 3 | 2.18 | 0.307182 | 3 | 0.088211 |
| CDH2 | Cadherin 2, type 1, N-cadherin | –1.28 | 0.092314 | 4.81 | 0.336937 |
| NOTCH4 | Notch 4 | 1.71 | 0.217780 | 2.75 | 0.225017 |
| PVRL1 | Poliovirus receptor-related 1 | 1.42 | 0.011171 | 1.57 | 0.264911 |
| Multi Treatment for 5 Days | |||||
|---|---|---|---|---|---|
| Gene | Function | Ery 0.3/Ery 0 | Ery 0.3/Ery 0 | Ery 300/Ery 0 | Ery 300/Ery 0 |
| Fold Change | p Value | Fold Change | p Value | ||
| Tight junction | |||||
| ICAM2 | Intercellular adhesion molecule 2 | –1.09 | 0.308601 | 1.2 | 0.000850 |
| CLDN10 | Claudin 10 | –1.09 | 0.649218 | 4.34 | 0.364360 |
| CLDN14 | Claudin 14 | –14.64 | 0.121784 | –2.79 | 0.217719 |
| CLDN17 | Claudin 17 | –2.14 | 0.392266 | –2.6 | 0.371088 |
| CLDN18 | Claudin 18 | 3.18 | 0.298743 | 8.38 | 0.132072 |
| CLDN19 | Claudin 19 | 8.2 | 0.099657 | 1.41 | 0.596306 |
| CLDN8 | Claudin 8 | –2.05 | 0.235259 | 1.97 | 0.191047 |
| Focal adhesion | |||||
| CAV3 | Caveolin 3 | –14.02 | 0.158030 | –3.76 | 0.284148 |
| ITGA4 | Integrin, alpha 4 | –3.86 | 0.372310 | –3.91 | 0.367691 |
| ITGAM | Integrin, alpha M | –3.77 | 0.596918 | 1.22 | 0.803690 |
| ITGB3 | Integrin, beta 3 | –2.93 | 0.201419 | –1.26 | 0.410330 |
| Adherens junction and desmosomes genes | |||||
| DSC1 | Desmocollin 1 | –1.07 | 0.969266 | –2.69 | 0.136576 |
| DSG3 | Desmoglein 3 | 8.14 | 0.293908 | 9.94 | 0.165064 |
| Gap junction | |||||
| GJA5 | Gap junction protein, alpha 5 | 1.35 | 0.454641 | 6.77 | 0.330339 |
| GJB6 | Gap junction protein, beta 6 | 4.39 | 0.375203 | 1.12 | 0.596642 |
| GJC2 | Gap junction protein, gamma 2 | 1.204 | 0.78586 | –2.78 | 0.688153 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, H.; Gokulan, K.; Piñeiro, S.A.; Williams, K.M.; Yuan, Z.; Cerniglia, C.E.; Khare, S. Effects of Acute and Chronic Exposure to Residual Level Erythromycin on Human Intestinal Epithelium Cell Permeability and Cytotoxicity. Microorganisms 2019, 7, 325. https://doi.org/10.3390/microorganisms7090325
Hao H, Gokulan K, Piñeiro SA, Williams KM, Yuan Z, Cerniglia CE, Khare S. Effects of Acute and Chronic Exposure to Residual Level Erythromycin on Human Intestinal Epithelium Cell Permeability and Cytotoxicity. Microorganisms. 2019; 7(9):325. https://doi.org/10.3390/microorganisms7090325
Chicago/Turabian StyleHao, Haihong, Kuppan Gokulan, Silvia A. Piñeiro, Katherine M. Williams, Zonghui Yuan, Carl E. Cerniglia, and Sangeeta Khare. 2019. "Effects of Acute and Chronic Exposure to Residual Level Erythromycin on Human Intestinal Epithelium Cell Permeability and Cytotoxicity" Microorganisms 7, no. 9: 325. https://doi.org/10.3390/microorganisms7090325
APA StyleHao, H., Gokulan, K., Piñeiro, S. A., Williams, K. M., Yuan, Z., Cerniglia, C. E., & Khare, S. (2019). Effects of Acute and Chronic Exposure to Residual Level Erythromycin on Human Intestinal Epithelium Cell Permeability and Cytotoxicity. Microorganisms, 7(9), 325. https://doi.org/10.3390/microorganisms7090325