Parachlamydia acanthamoebae Detected during a Pneumonia Outbreak in Southeastern Finland, in 2017–2018

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Respiratory Tract Samples
2.2. DNA Extraction
2.3. Detection of Chlamydia pneumoniae
2.4. PanChlamydiales-PCR
2.5. Amoeba PCR
2.6. Sequencing and Phylogeny
3. Results
3.1. Detection of Chlamydia pneumoniae
3.2. Detection of Chlamydiales DNA
3.3. Amoeba PCR
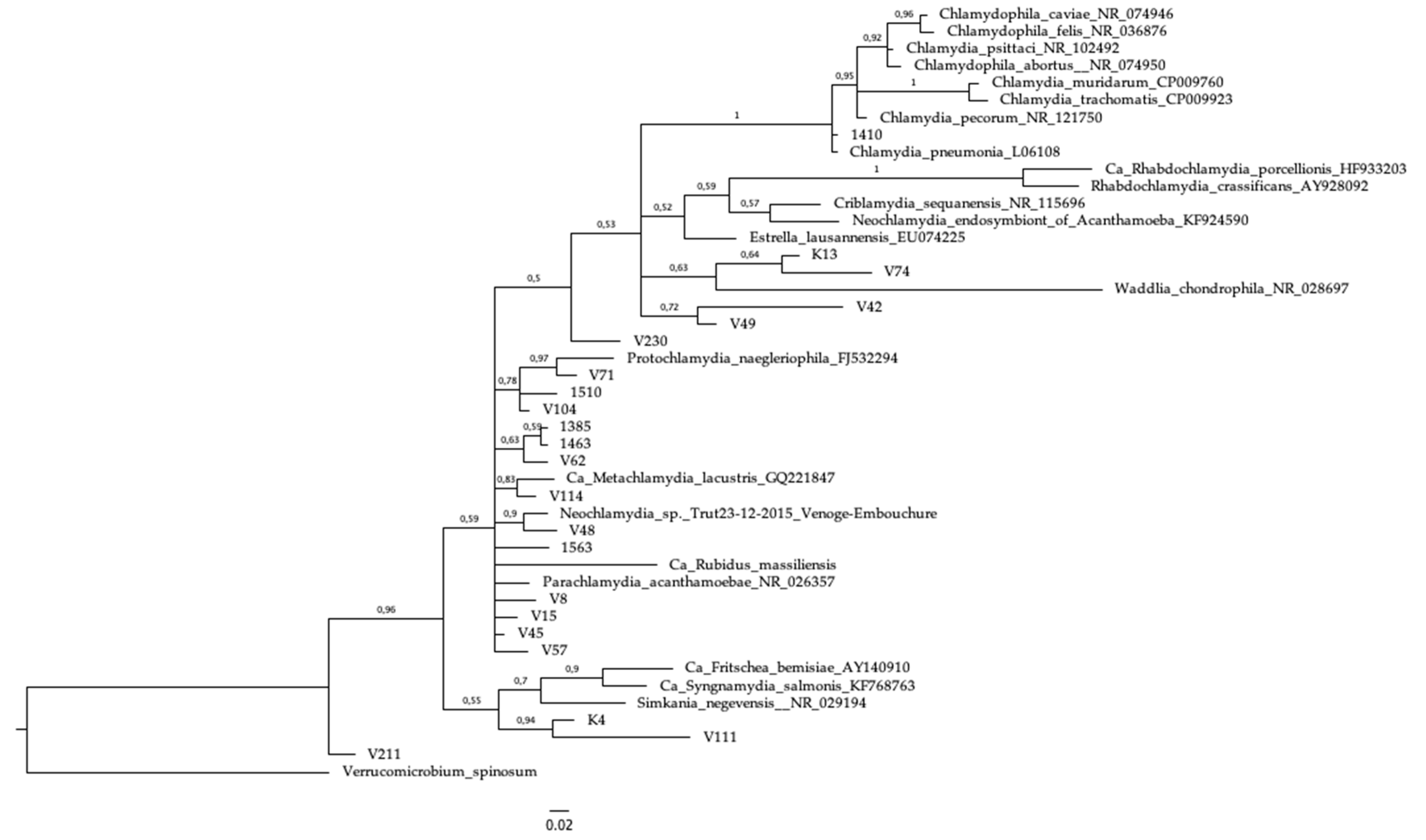
3.4. Sequencing and Phylogeny
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pillonel, T.; Bertelli, C.; Greub, G. Environmental Metagenomic Assemblies Reveal Seven New Highly Divergent Chlamydial Lineages and Hallmarks of a Conserved Intracellular Lifestyle. Front. Microbiol. 2018, 9, 79. [Google Scholar] [CrossRef] [PubMed]
- Corsaro, D.; Venditti, D. Diversity of the parachlamydiae in the environment. Crit. Rev. Microbiol. 2006, 32, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Brown, A.; Vaughan, L.; Greub, G.; Timms, P.; Polkinghorne, A. Twenty years of research into Chlamydia-like organisms: A revolution in our understanding of the biology and pathogenicity of members of the phylum Chlamydiae. Pathog. Dis. 2015, 73, 1–15. [Google Scholar] [CrossRef]
- Pilloux, L.; Casson, N.; Sommer, K.; Klos, A.; Stehle, J.C.; Pusztaszeri, M.; Greub, G. Severe pneumonia due to Parachlamydia acanthamoebae following intranasal inoculation: A mice model. Microbes Infect. 2015, 17, 755–760. [Google Scholar] [CrossRef]
- Greub, G.; Boyadjiev, I.; La Scola, B.; Raoult, D.; Martin, C. Serological hint suggesting that Parachlamydiaceae are agents of pneumonia in polytraumatized intensive care patients. Ann. N. Y. Acad. Sci. 2003, 990, 311–319. [Google Scholar] [CrossRef]
- Verweij, S.P.; Kebbi-Beghdadi, C.; Land, J.A.; Ouburg, S.; Morre, S.A.; Greub, G. Waddlia chondrophila and Chlamydia trachomatis antibodies in screening infertile women for tubal pathology. Microbes Infect. 2015, 17, 745–748. [Google Scholar] [CrossRef]
- Baud, D.; Regan, L.; Greub, G. Emerging role of Chlamydia and Chlamydia-like organisms in adverse pregnancy outcomes. Curr. Opin. Infect. Dis. 2008, 21, 70–76. [Google Scholar] [CrossRef]
- Haider, S.; Collingro, A.; Walochnik, J.; Wagner, M.; Horn, M. Chlamydia-like bacteria in respiratory samples of community-acquired pneumonia patients. FEMS Microbiol. Lett. 2008, 281, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Goy, G.; Croxatto, A.; Posfay-Barbe, K.M.; Gervaix, A.; Greub, G. Development of a real-time PCR for the specific detection of Waddlia chondrophila in clinical samples. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 1483–1486. [Google Scholar] [CrossRef] [PubMed]
- Heiskanen-Kosma, T.; Paldanius, M.; Korppi, M. Simkania negevensis may be a true cause of community acquired pneumonia in children. Scand. J. Infect. Dis. 2008, 40, 127–130. [Google Scholar] [CrossRef]
- Niemi, S.; Greub, G.; Puolakkainen, M. Chlamydia-related bacteria in respiratory samples in Finland. Microbes Infect. 2011, 13, 824–827. [Google Scholar] [CrossRef]
- Lamoth, F.; Jaton, K.; Vaudaux, B.; Greub, G. Parachlamydia and Rhabdochlamydia: Emerging agents of community-acquired respiratory infections in children. Clin. Infect. Dis. 2011, 53, 500–501. [Google Scholar] [CrossRef]
- Kurkela, S.; Puolakkainen, M.; Hokynar, K.; Nieminen, T.; Saxen, H.; Mannonen, L.; Pietikäinen, R. Mycoplasma pneumoniae outbreak, Southeastern Finland, 2017–2018: Molecular epidemiology and laboratory diagnostic lessons. Eur. J. Clin. Microbiol. Infect. Dis. 2019. under review. [Google Scholar]
- Nummi, M.; Mannonen, L.; Puolakkainen, M. Development of a multiplex real-time PCR assay for detection of Mycoplasma pneumoniae, Chlamydia pneumoniae and mutations associated with macrolide resistance in Mycoplasma pneumoniae from respiratory clinical specimens. Springerplus 2015, 4, 684. [Google Scholar] [CrossRef]
- Mannonen, L.; Kamping, E.; Penttila, T.; Puolakkainen, M. IFN-gamma induced persistent Chlamydia pneumoniae infection in HL and Mono Mac 6 cells: characterization by real-time quantitative PCR and culture. Microb. Pathog. 2004, 36, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Lienard, J.; Croxatto, A.; Aeby, S.; Jaton, K.; Posfay-Barbe, K.; Gervaix, A.; Greub, G. Development of a new chlamydiales-specific real-time PCR and its application to respiratory clinical samples. J. Clin. Microbiol. 2011, 49, 2637–2642. [Google Scholar] [CrossRef]
- Hokynar, K.; Sormunen, J.J.; Vesterinen, E.J.; Partio, E.K.; Lilley, T.; Timonen, V.; Panelius, J.; Ranki, A.; Puolakkainen, M. Chlamydia-Like Organisms (CLOs) in Finnish Ixodes ricinus Ticks and Human Skin. Microorganisms 2016, 4, 28. [Google Scholar] [CrossRef]
- Everett, K.D.; Bush, R.M.; Andersen, A.A. Emended description of the order Chlamydiales, proposal of Parachlamydiaceae fam. nov. and Simkaniaceae fam. nov., each containing one monotypic genus, revised taxonomy of the family Chlamydiaceae, including a new genus and five new species, and standards for the identification of organisms. Int. J. Syst. Bacteriol. 1999, 49, 415–440. [Google Scholar]
- Qvarnstrom, Y.; Visvesvara, G.S.; Sriram, R.; da Silva, A.J. Multiplex real-time PCR assay for simultaneous detection of Acanthamoeba spp., Balamuthia mandrillaris, and Naegleria fowleri. J. Clin. Microbiol. 2006, 44, 3589–3595. [Google Scholar] [CrossRef] [PubMed]
- Scheikl, U.; Tsao, H.F.; Horn, M.; Indra, A.; Walochnik, J. Free-living amoebae and their associated bacteria in Austrian cooling towers: A 1-year routine screening. Parasitol. Res. 2016, 115, 3365–3374. [Google Scholar] [CrossRef]
- Kuiper, M.W.; Valster, R.M.; Wullings, B.A.; Boonstra, H.; Smidt, H.; van der Kooij, D. Quantitative detection of the free-living amoeba Hartmannella vermiformis in surface water by using real-time PCR. Appl. Environ. Microbiol. 2006, 72, 5750–5756. [Google Scholar] [CrossRef] [PubMed]
- Pillonel, T.; Bertelli, C.; Salamin, N.; Greub, G. Taxogenomics of the order Chlamydiales. Int. J. Syst. Evol. Microbiol. 2015, 65, 1381–1393. [Google Scholar] [CrossRef] [PubMed]
- Hokynar, K.; Vesterinen, E.J.; Lilley, T.M.; Pulliainen, A.T.; Korhonen, S.J.; Paavonen, J.; Puolakkainen, M. Molecular Evidence of Chlamydia-Like Organisms in the Feces of Myotis daubentonii Bats. Appl. Environ. Microbiol. 2017, 83, 2. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Polkinghorne, A.; Borel, N.; Heijne, M.; Pannekoek, Y. New evidence for domesticated animals as reservoirs of Chlamydia-associated community-acquired pneumonia. Clin. Microbiol. Infect. 2018, 25, 131–132. [Google Scholar] [CrossRef]
- Ramakers, B.P.; Heijne, M.; Lie, N.; Le, T.N.; van Vliet, M.; Claessen, V.P.J.; Tolsma, P.J.P.; De Rosa, M.; Roest, H.I.J.; Vanrompay, D.; et al. Zoonotic Chlamydia caviae Presenting as Community-Acquired Pneumonia. N. Engl. J. Med. 2017, 377, 992–994. [Google Scholar] [CrossRef]
- Van Grootveld, R.; Bilsen, M.P.; Boelsums, T.L.; Heddema, E.R.; Groeneveld, G.H.; Gooskens, J.; de Boer, M.G.J. Chlamydia caviae Causing Community-Acquired Pneumonia: An Emerging Zoonosis. Vector. Borne. Zoonotic. Dis. 2018, 18, 635–637. [Google Scholar] [CrossRef]
- Marrie, T.J.; Raoult, D.; La Scola, B.; Birtles, R.J.; de Carolis, E.; Canadian Community-Acquired Pneumonia Study Group. Legionella-like and other amoebal pathogens as agents of community-acquired pneumonia. Emerg. Infect. Dis. 2001, 7, 1026–1029. [Google Scholar] [CrossRef]
- Greub, G.; Berger, P.; Papazian, L.; Raoult, D. Parachlamydiaceae as rare agents of pneumonia. Emerg. Infect. Dis. 2003, 9, 755–756. [Google Scholar] [CrossRef]
- Greub, G.; Mege, J.L.; Raoult, D. Parachlamydia acanthamoebae enters and multiplies within human macrophages and induces their apoptosis [corrected]. Infect. Immun. 2003, 71, 5979–5985. [Google Scholar] [CrossRef]
- Casson, N.; Medico, N.; Bille, J.; Greub, G. Parachlamydia acanthamoebae enters and multiplies within pneumocytes and lung fibroblasts. Microbes Infect. 2006, 8, 1294–1300. [Google Scholar] [CrossRef] [PubMed]
- Casson, N.; Posfay-Barbe, K.M.; Gervaix, A.; Greub, G. New diagnostic real-time PCR for specific detection of Parachlamydia acanthamoebae DNA in clinical samples. J. Clin. Microbiol. 2008, 46, 1491–1493. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Garin, N.; Hugli, O.; Genne, D.; Greub, G. Lack of Chlamydia-related bacteria among patients with community-acquired pneumonia. New Microbes New Infect. 2015, 8, 164–165. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Garin, N.; Genne, D.; Carballo, S.; Chuard, C.; Eich, G.; Hugli, O.; Lamy, O.; Nendaz, M.; Petignat, P.A.; Perneger, T.; et al. beta-Lactam monotherapy vs beta-lactam-macrolide combination treatment in moderately severe community-acquired pneumonia: A randomized noninferiority trial. JAMA Intern. Med. 2014, 174, 1894–1901. [Google Scholar] [CrossRef]
- Vouga, M.; Diabi, H.; Boulos, A.; Baud, D.; Raoult, D.; Greub, G. Antibiotic susceptibility of Neochlamydia hartmanellae and Parachlamydia acanthamoebae in amoebae. Microbes Infect. 2015, 17, 761–765. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Target | Primer/Probe | Sequence | Reference |
|---|---|---|---|
| M. pneumoniae Hypothetical protein C985_0367 | MP Fw | ATGTACTATCAGCAAAAGCTCAGTATGG | [14] |
| MP Rev | CCACATACCGCTTTAAGTTAGCAA | ||
| MP probe | Cy5-CTAACCAAAACAGCCCTTCAACGGCA-Iowa black RQ-Sp | ||
| C. pneumoniae ompA | CP Fw | AAGGGCTATAAAGGCGTTGCT | [14,15] |
| CP Rev | TGGTCGCAGACTTTGTTCCA | ||
| CP probe | Tx Red-TCCCCTTGCCAACAGACGCTGG-Iowa black RQ-Sp | ||
| Homo sapiens beta-globin chain | BG Fw | GGTTGGGATAAGGCTGGATTATT | [14] |
| BG Rev | CAGGAGCTGTGGGAGGAAGA | ||
| BG Probe | JOE/ZEN-CAAGCTAGGCCCTTTTGCTAATCATGTTCA-Iowa black FQ | ||
| M. pneumoniae gene for 23S rRNA | MR–MP Fw | GCTATAGACTCGGTGAAATCCAGG | [14] |
| MR–MP Rev | GCTACAGTAAAGCTTCACTGGGTC | ||
| MR–MP SimpleProbe® | GCGCA XI ACGGGACGGAAAGAC | ||
| Chlamydiales 16S rRNA gene | panCh-Fwd | CCGCCAACACTGGGACT | * [16,17] |
| panCh-Rev | GGAGTTAGCCGGTGCTTCTTTAC | ||
| panCh-Probe | FAM-CTACGGGAGGCTGCAGTCGAGAATC-BHQ1 | ||
| Acanthamoeba spp. 18S rRNA gene | Acan Fw | CCAGATCGTTTACCGTGAA | * [19] |
| Acan Rv | TATTAATGCCCCCAACTATCC | ||
| Acan probe | FAM-TCTGCCACCGAATACATTAGCATGG-Iowa black FQ | ||
| Vahlkampfiidae spp. 18S rRNA gene | Vahl Fw | GTATAGTCGCAAGACCGAAAC | * [20] |
| Vahl Rv | CAAGACAGATCACTCCACGA | ||
| Vahl probe | Cy5-GGGAAACTCACCAGGTCAGGACAC-Iowa black FQ | ||
| Vermamoeba vermiformis 18S rRNA gene | Verm Fw | TACGAGGTCAGGACACTGTG | * [21] |
| Verm Rv | ACCATCCGGAGTTCTCG | ||
| Verm probe | JOE-AATTCCTAGTAGGCGCGAGTCATCAA-Iowa black FQ |
| Sample ID | Age (Years) | Gender | GROUP * | School ** | Symptoms | Chlamydiales Family | Other | |
|---|---|---|---|---|---|---|---|---|
| 1410 | 8 | male | 1 | yes | Chlamydiaceae | C. pneumoniae PCR+ | ||
| 1563 | 10 | female | 1 | yes | Parachlamydiaceae | M. pneumoniae PCR+, IgM+ | ||
| 1385 | 6 | male | 1 | yes | Parachlamydiaceae | |||
| 1463 | 7 | female | 1 | yes | Parachlamydiaceae | M. pneumoniae IgM+ | ||
| 1510 | 7 | male | 1 | yes | Parachlamydiaceae | |||
| V15 | 13 | female | 2 | 1 | NA | Parachlamydiaceae | ||
| V45 | 14 | female | 2 | 1 | yes | fever, cough, rhinitis | Parachlamydiaceae | |
| V8 | 13 | male | 2 | 1 | NA | Parachlamydiaceae | M. pneumoniae PCR+ | |
| V48 | 14 | female | 2 | 1 | yes | fever, cough, rhinitis | Parachlamydiaceae | |
| V57 | 14 | male | 2 | 1 | NA | Parachlamydiaceae | ||
| V62 | 9 | male | 2 | 2 | yes | cough, rhinitis | Parachlamydiaceae | |
| V71 | 9 | male | 2 | 2 | no | Parachlamydiaceae | ||
| V104 | 12 | male | 2 | 2 | yes | cough, rhinitis | Parachlamydiaceae | |
| V111 | 17 | female | 2 | 3 | yes | fever, cough, rhinitis | Parachlamydiaceae | |
| V114 | 14 | female | 2 | 1 | NA | Parachlamydiaceae | ||
| K4 | 9 | female | 2 | 4 | yes | fever, cough | Parachlamydiaceae | |
| V230 | 17 | female | 2 | 3 | NA | Parachlamydiaceae | ||
| V74 | 9 | male | 2 | 2 | yes | fever | Criblamydiaceae | |
| K13 | 10 | male | 2 | 4 | yes | fever, cough, rhinitis | Criblamydiaceae | |
| V211 | 11 | female | 2 | 2 | yes | rhinitis | Simkaniaceae | |
| V42 | 14 | female | 2 | 1 | yes | cough, rhinitis | Unclassified | |
| V49 | 14 | male | 2 | 1 | yes | fever | Unclassified | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hokynar, K.; Kurkela, S.; Nieminen, T.; Saxen, H.; Vesterinen, E.J.; Mannonen, L.; Pietikäinen, R.; Puolakkainen, M. Parachlamydia acanthamoebae Detected during a Pneumonia Outbreak in Southeastern Finland, in 2017–2018. Microorganisms 2019, 7, 141. https://doi.org/10.3390/microorganisms7050141
Hokynar K, Kurkela S, Nieminen T, Saxen H, Vesterinen EJ, Mannonen L, Pietikäinen R, Puolakkainen M. Parachlamydia acanthamoebae Detected during a Pneumonia Outbreak in Southeastern Finland, in 2017–2018. Microorganisms. 2019; 7(5):141. https://doi.org/10.3390/microorganisms7050141
Chicago/Turabian StyleHokynar, Kati, Satu Kurkela, Tea Nieminen, Harri Saxen, Eero J. Vesterinen, Laura Mannonen, Risto Pietikäinen, and Mirja Puolakkainen. 2019. "Parachlamydia acanthamoebae Detected during a Pneumonia Outbreak in Southeastern Finland, in 2017–2018" Microorganisms 7, no. 5: 141. https://doi.org/10.3390/microorganisms7050141
APA StyleHokynar, K., Kurkela, S., Nieminen, T., Saxen, H., Vesterinen, E. J., Mannonen, L., Pietikäinen, R., & Puolakkainen, M. (2019). Parachlamydia acanthamoebae Detected during a Pneumonia Outbreak in Southeastern Finland, in 2017–2018. Microorganisms, 7(5), 141. https://doi.org/10.3390/microorganisms7050141