Efficient Epstein-Barr Virus Progeny Production Mediated by Cancer-Derived LMP1 and Virally-Encoded microRNAs

 , ,
, , 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Plasmids
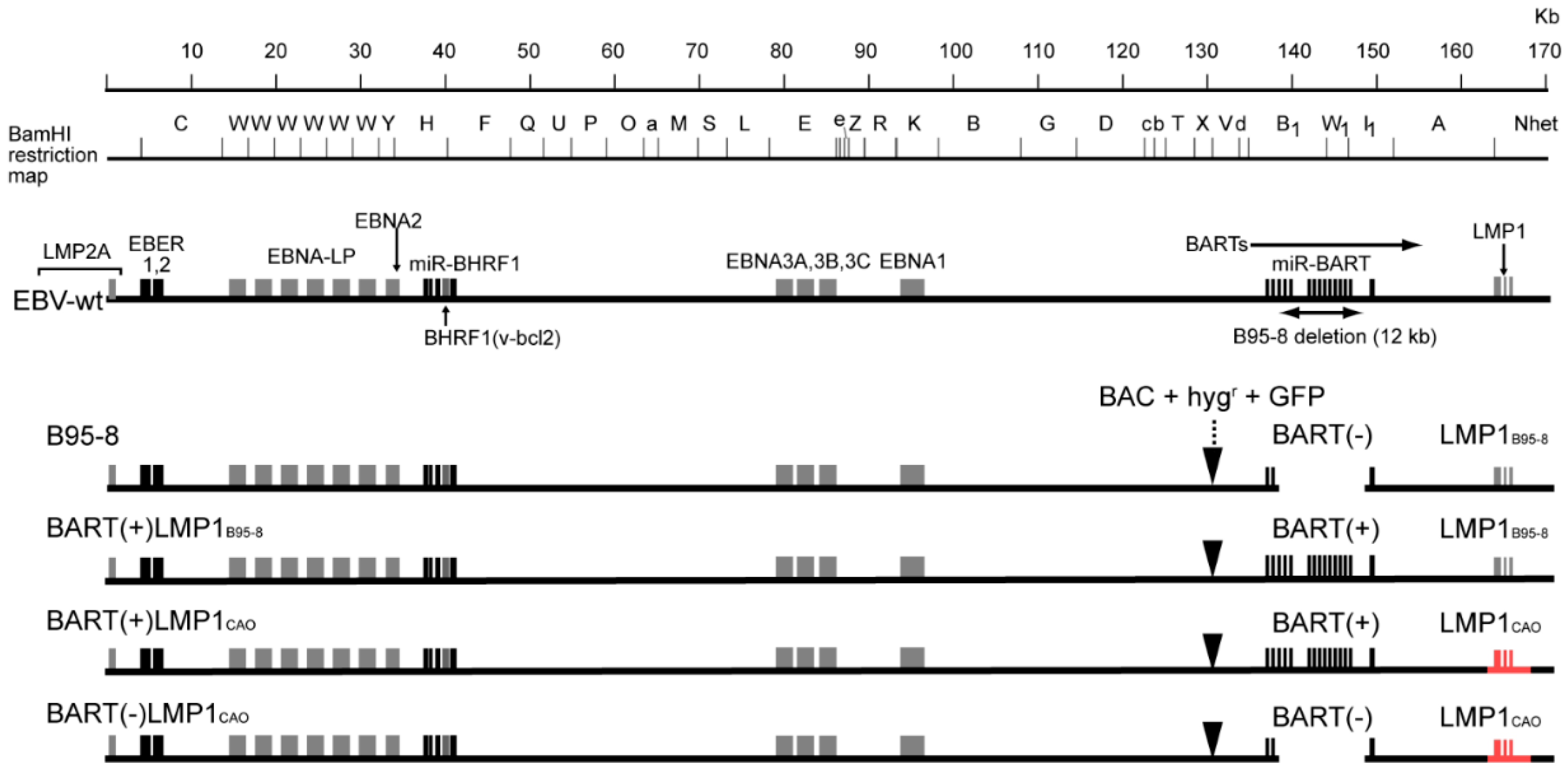
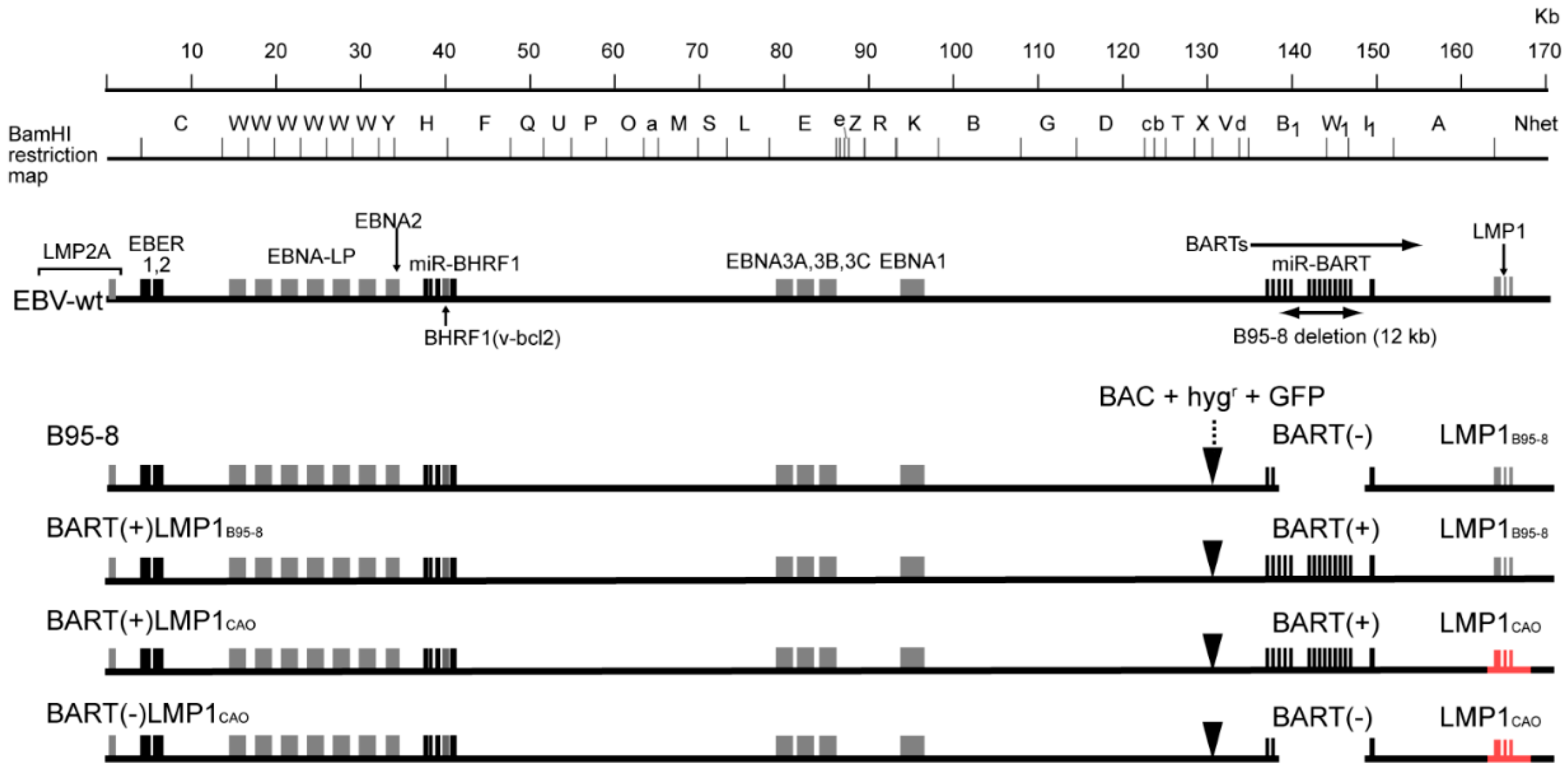
2.3. EBV-BAC Clones
2.4. Engineering of the Viral Genome in E. coli
2.5. Deep Sequencing
2.6. Recombinant Virus Production and Infection
2.7. Expression of Viral Protein and microRNA
2.8. Reporter Assay
2.9. LMP1 Sequences of Japanese NPC-Derived EBVs
2.10. Nucleotide Sequence Accession Numbers
3. Results
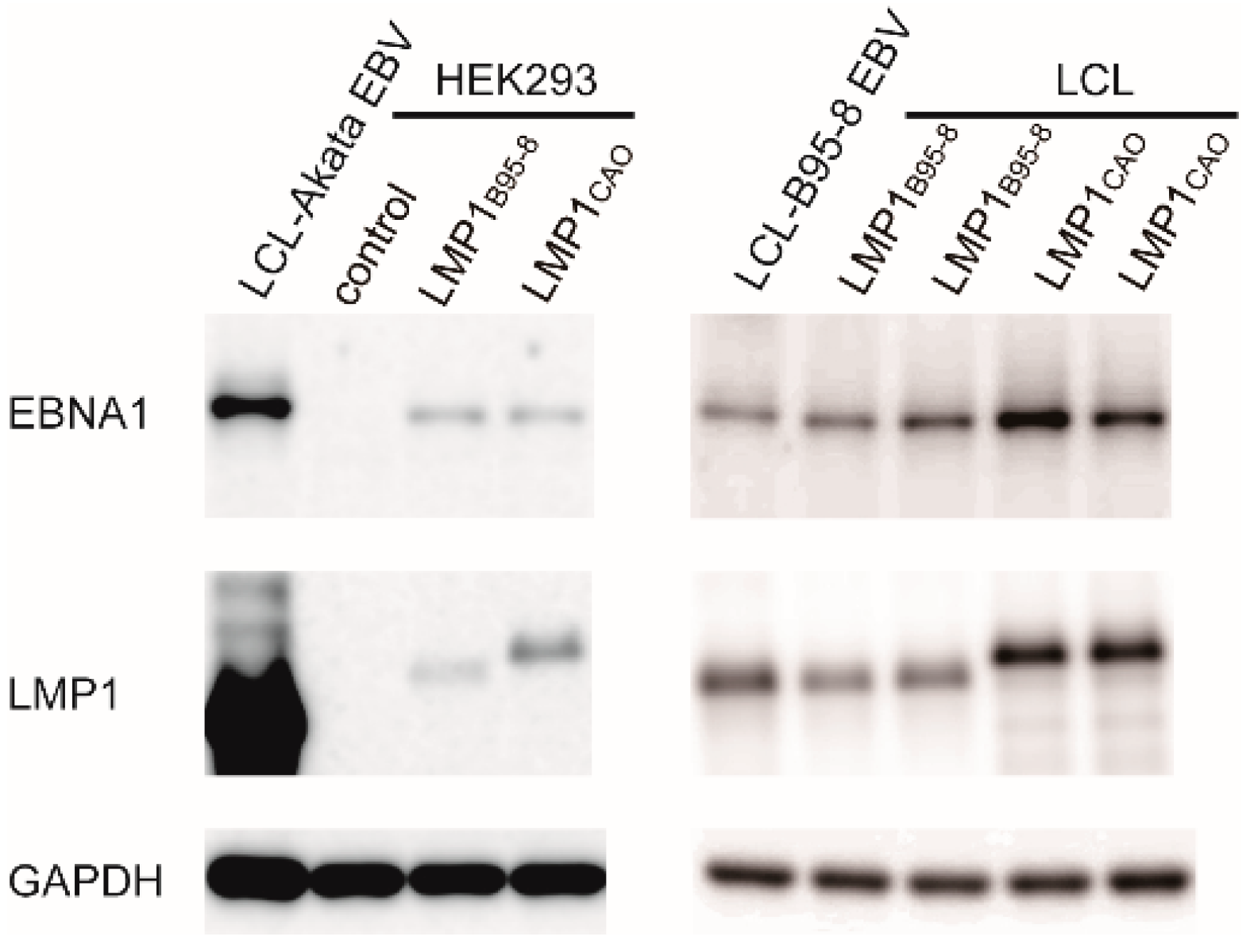
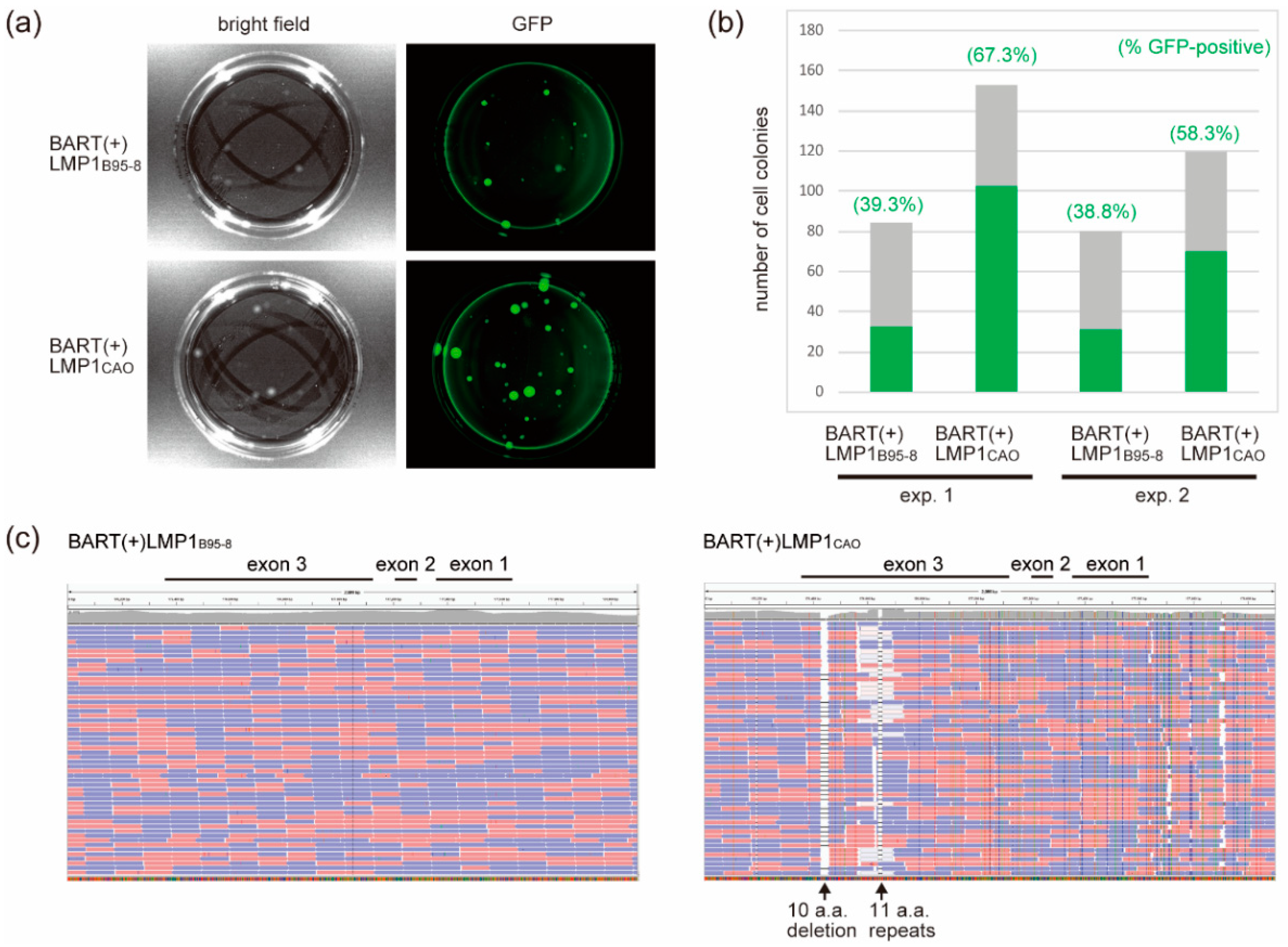
3.1. Stable Transfection of HEK293 Cells with an EBV-BAC Clone Harboring LMP1CAO and BART miRNA Genes
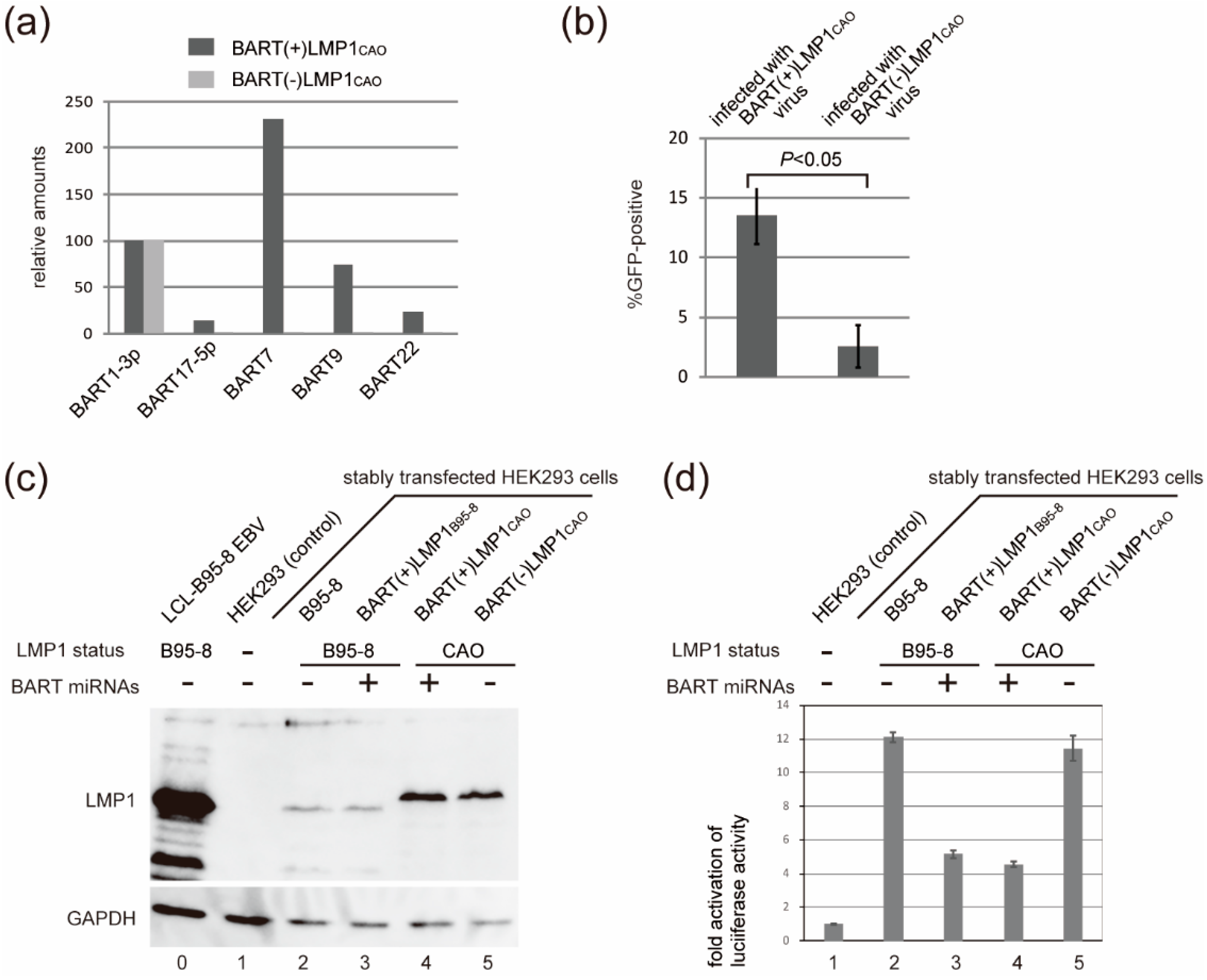
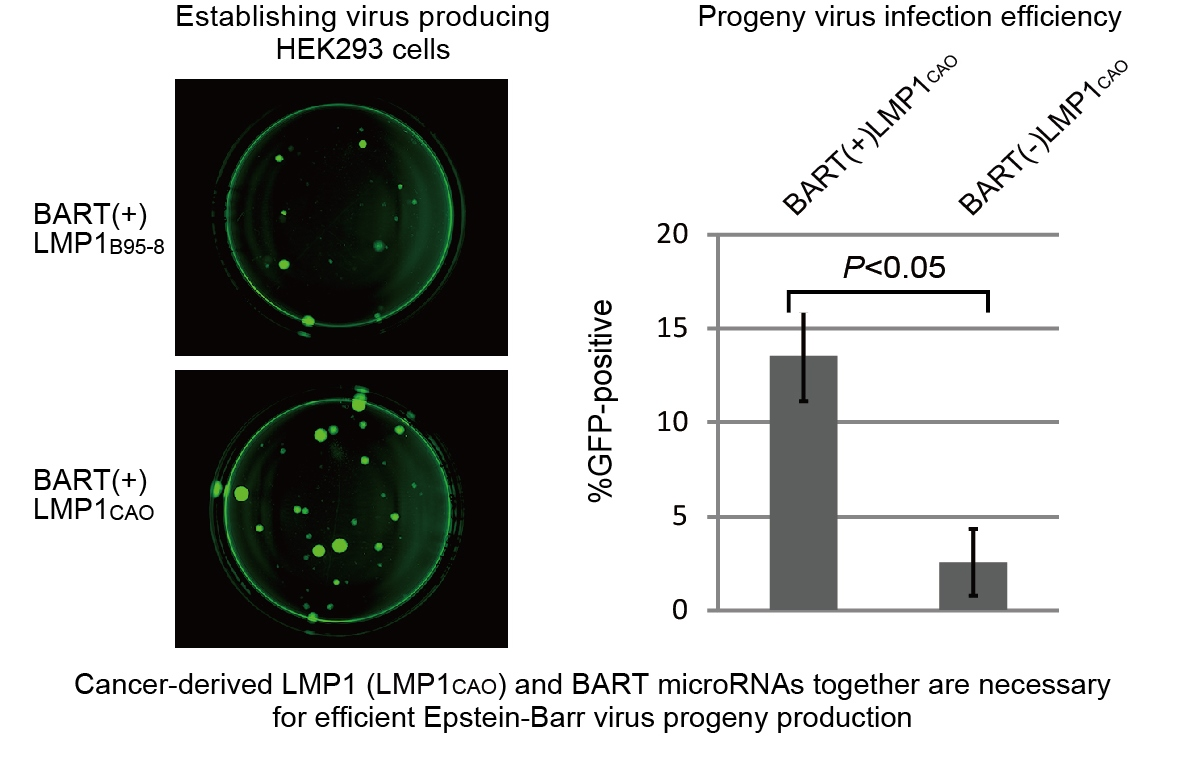
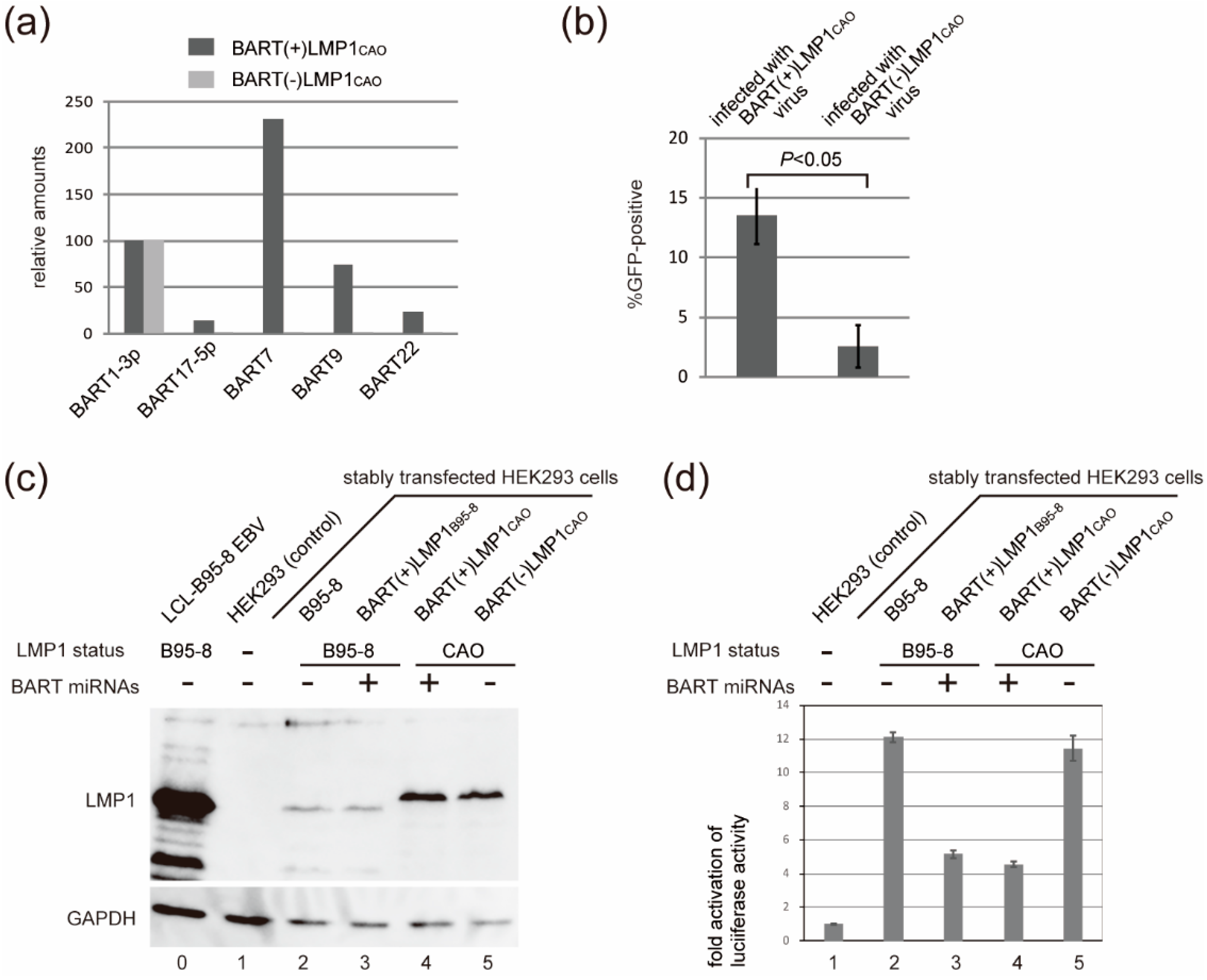
3.2. Efficient Production of Progeny Virus by HEK293 Cells Harboring BART(+)LMP1CAO
3.3. Deletion of BART miRNA Compromises the Ability to Generate Progeny Viruses
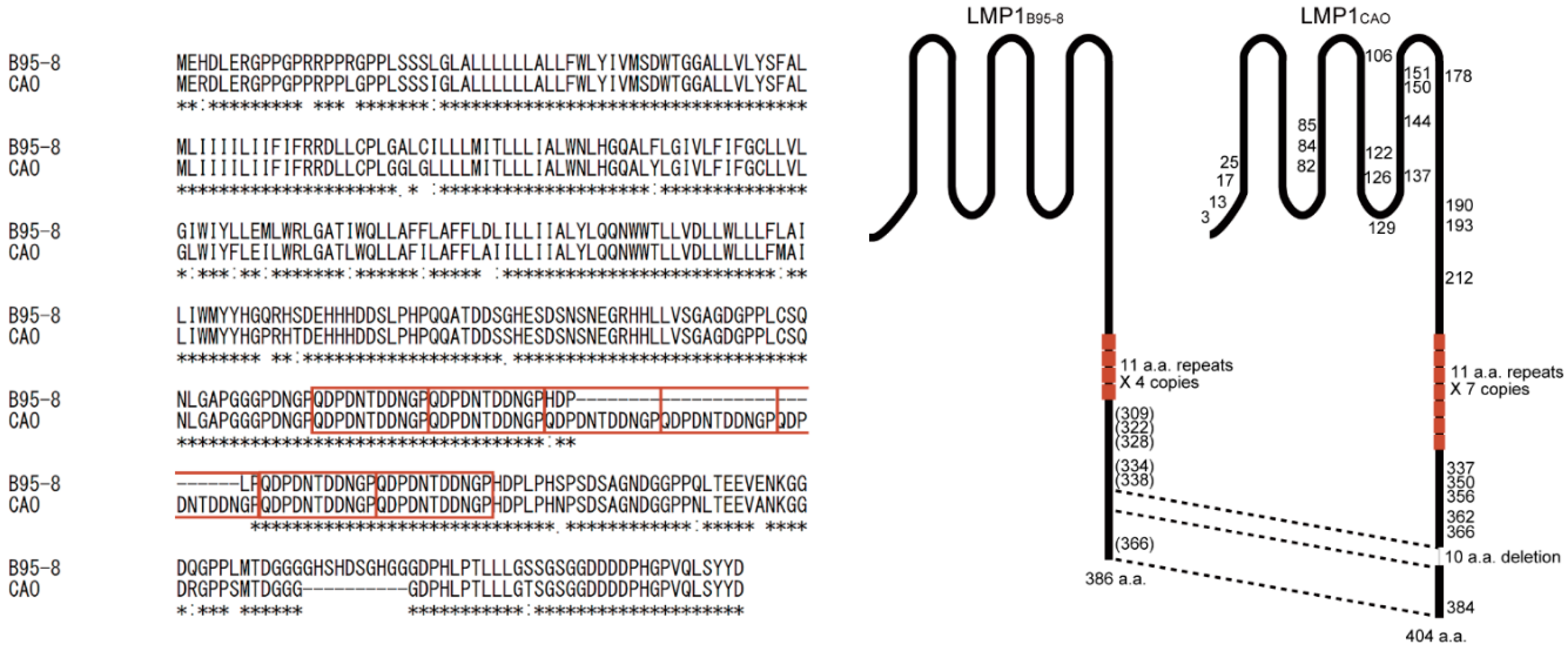
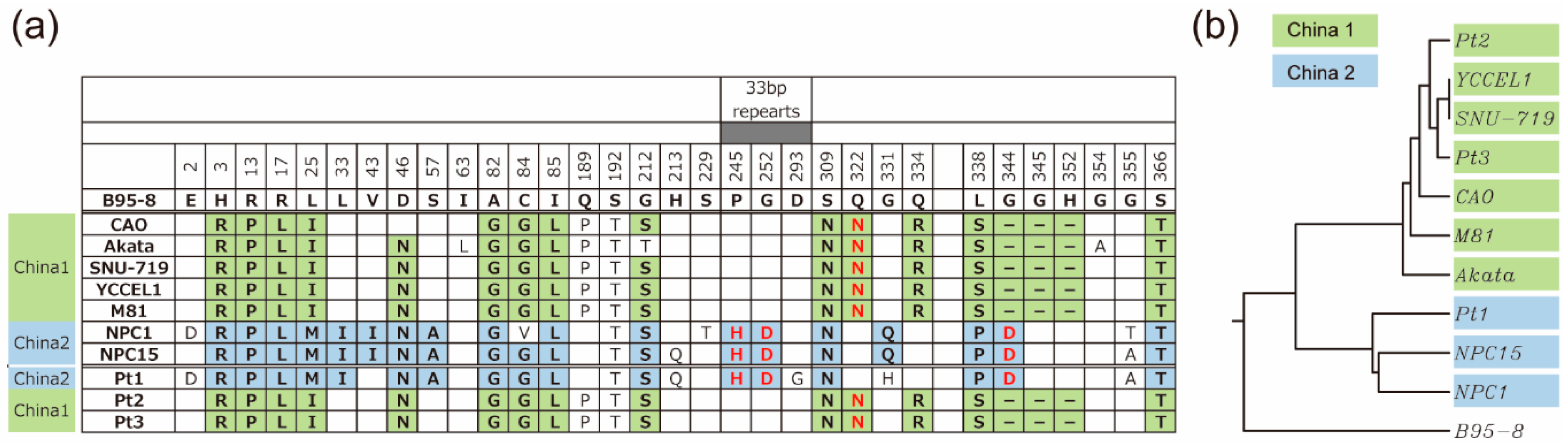
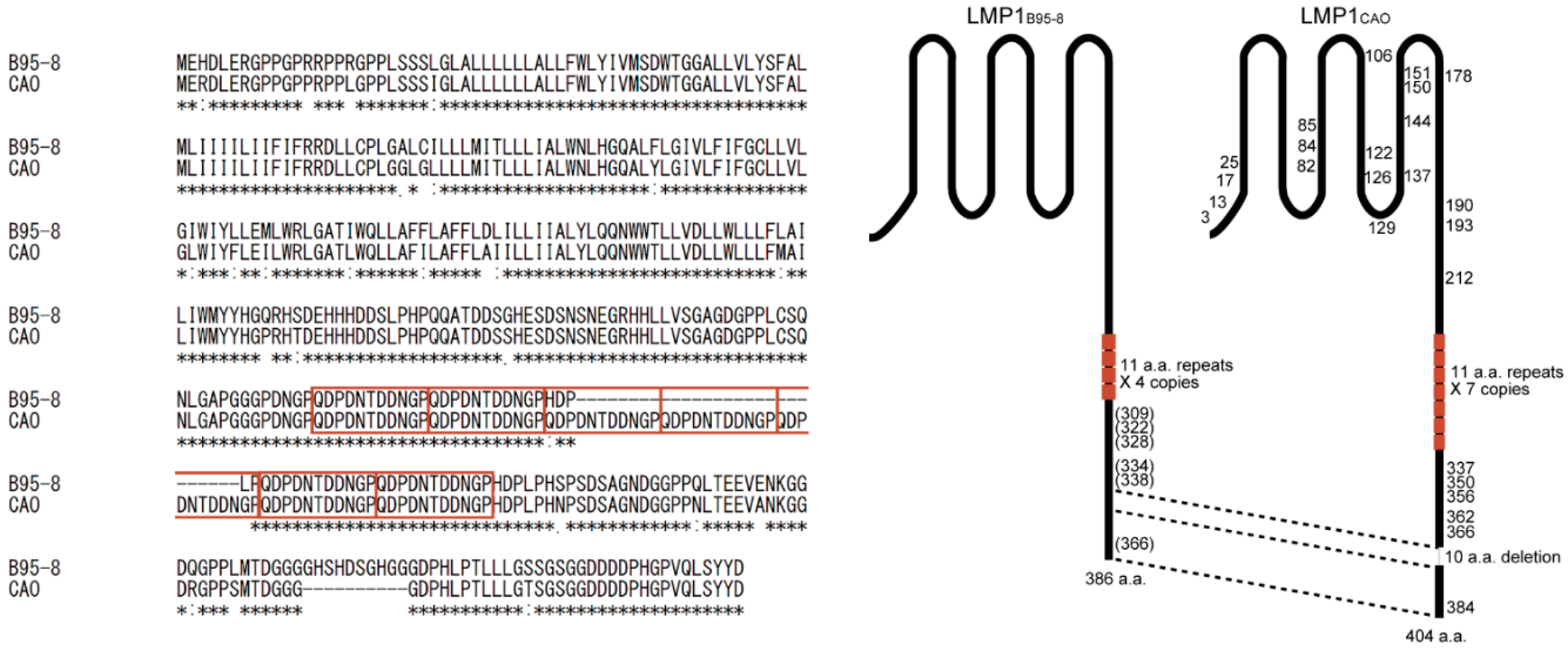
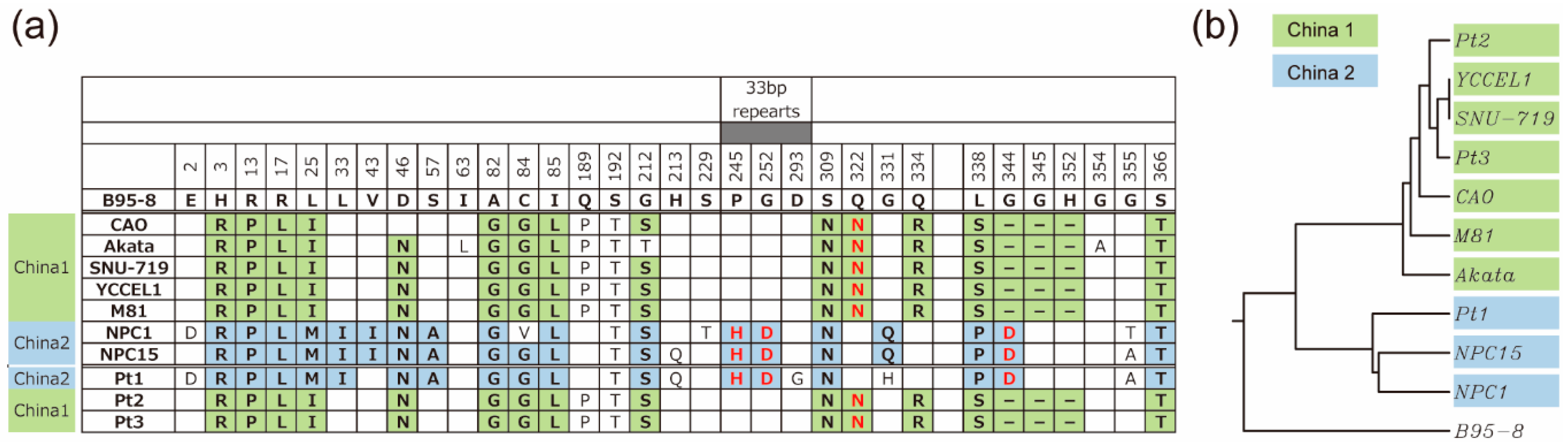
3.4. Amino Acid Sequences of LMP1 from Japanese NPC Patients
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Rickinson, A.B.; Kieff, E. Epstein-Barr virus. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 2, pp. 2655–2700. [Google Scholar]
- Tsao, S.W.; Tsang, C.M.; To, K.F.; Lo, K.W. The role of Epstein-Barr virus in epithelial malignancies. J. Pathol. 2015, 235, 323–333. [Google Scholar] [CrossRef]
- Kieff, E.; Rickinson, A.B. Epstein-Barr virus and its replication. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 2, pp. 2603–2654. [Google Scholar]
- Kaye, K.M.; Izumi, K.M.; Kieff, E. Epstein-Barr virus latent membrane protein 1 is essential for B-lymphocyte growth transformation. Proc. Natl. Acad. Sci. USA 1993, 90, 9150–9154. [Google Scholar] [CrossRef] [PubMed]
- Uchida, J.; Yasui, T.; Takaoka-Shichijo, Y.; Muraoka, M.; Kulwichit, W.; Raab-Traub, N.; Kikutani, H. Mimicry of CD40 signals by Epstein-Barr virus LMP1 in B lymphocyte responses. Science 1999, 286, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Tsao, S.W.; Tramoutanis, G.; Dawson, C.W.; Lo, A.K.; Huang, D.P. The significance of LMP1 expression in nasopharyngeal carcinoma. Semin. Cancer Biol. 2002, 12, 473–487. [Google Scholar] [CrossRef] [PubMed]
- Huen, D.S.; Henderson, S.A.; Croom-Carter, D.; Rowe, M. The Epstein-Barr virus latent membrane protein-1 (LMP1) mediates activation of NF-kappa B and cell surface phenotype via two effector regions in its carboxy-terminal cytoplasmic domain. Oncogene 1995, 10, 549–560. [Google Scholar] [PubMed]
- Gires, O.; Kohlhuber, F.; Kilger, E.; Baumann, M.; Kieser, A.; Kaiser, C.; Zeidler, R.; Scheffer, B.; Ueffing, M.; Hammerschmidt, W. Latent membrane protein 1 of Epstein-Barr virus interacts with JAK3 and activates STAT proteins. EMBO J. 1999, 18, 3064–3073. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liebowitz, D.; Kieff, E. An EBV membrane protein expressed in immortalized lymphocytes transforms established rodent cells. Cell 1985, 43, 831–840. [Google Scholar] [CrossRef]
- Dawson, C.W.; Port, R.J.; Young, L.S. The role of the EBV-encoded latent membrane proteins LMP1 and LMP2 in the pathogenesis of nasopharyngeal carcinoma (NPC). Semin. Cancer Biol. 2012, 22, 144–153. [Google Scholar] [CrossRef]
- Farrell, P.J. Epstein-Barr Virus Strain Variation. Curr. Top. Microbiol. Immunol. 2015, 390, 45–69. [Google Scholar]
- Feederle, R.; Klinke, O.; Kutikhin, A.; Poirey, R.; Tsai, M.H.; Delecluse, H.J. Epstein-Barr Virus: From the Detection of Sequence Polymorphisms to the Recognition of Viral Types. Curr. Top. Microbiol. Immunol. 2015, 390, 119–148. [Google Scholar]
- Edwards, R.H.; Seillier-Moiseiwitsch, F.; Raab-Traub, N. Signature amino acid changes in latent membrane protein 1 distinguish Epstein-Barr virus strains. Virology 1999, 261, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.F.; Zabarovsky, E.R.; Chen, F.; Cao, S.L.; Ernberg, I.; Klein, G.; Winberg, G. Isolation and sequencing of the Epstein-Barr virus BNLF-1 gene (LMP1) from a Chinese nasopharyngeal carcinoma. J. Gen. Virol. 1991, 72, 2399–2409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blake, S.M.; Eliopoulos, A.G.; Dawson, C.W.; Young, L.S. The transmembrane domains of the EBV-encoded latent membrane protein 1 (LMP1) variant CAO regulate enhanced signalling activity. Virology 2001, 282, 278–287. [Google Scholar] [CrossRef]
- Johnson, R.J.; Stack, M.; Hazlewood, S.A.; Jones, M.; Blackmore, C.G.; Hu, L.F.; Rowe, M. The 30-base-pair deletion in Chinese variants of the Epstein-Barr virus LMP1 gene is not the major effector of functional differences between variant LMP1 genes in human lymphocytes. J. Virol. 1998, 72, 4038–4048. [Google Scholar] [PubMed]
- Baer, R.; Bankier, A.T.; Biggin, M.D.; Deininger, P.L.; Farrell, P.J.; Gibson, T.J.; Hatfull, G.; Hudson, G.S.; Satchwell, S.C.; Seguin, C.; et al. DNA sequence and expression of the B95-8 Epstein-Barr virus genome. Nature 1984, 310, 207–211. [Google Scholar] [CrossRef]
- Nitta, T.; Chiba, A.; Yamamoto, N.; Yamaoka, S. Lack of cytotoxic property in a variant of Epstein-Barr virus latent membrane protein-1 isolated from nasopharyngeal carcinoma. Cell. Signal. 2004, 16, 1071–1081. [Google Scholar] [CrossRef]
- Dawson, C.W.; Eliopoulos, A.G.; Blake, S.M.; Barker, R.; Young, L.S. Identification of functional differences between prototype Epstein-Barr virus-encoded LMP1 and a nasopharyngeal carcinoma-derived LMP1 in human epithelial cells. Virology 2000, 272, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Shair, K.H.Y.; Reddy, A.; Cooper, V.S. New Insights from Elucidating the Role of LMP1 in Nasopharyngeal Carcinoma. Cancers 2018, 10, 86. [Google Scholar] [CrossRef]
- Delecluse, H.J.; Hilsendegen, T.; Pich, D.; Zeidler, R.; Hammerschmidt, W. Propagation and recovery of intact, infectious Epstein-Barr virus from prokaryotic to human cells. Proc. Natl. Acad. Sci. USA 1998, 95, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Zuo, L.; Yu, H.; Liu, L.; Tang, Y.; Wu, H.; Yang, J.; Zhu, M.; Du, S.; Zhao, L.; Cao, L.; et al. The copy number of Epstein-Barr virus latent genome correlates with the oncogenicity by the activation level of LMP1 and NF-kappaB. Oncotarget 2015, 6, 41033–41044. [Google Scholar] [CrossRef]
- Raab-Traub, N.; Dambaugh, T.; Kieff, E. DNA of Epstein-Barr virus VIII: B95-8, the previous prototype, is an unusual deletion derivative. Cell 1980, 22, 257–267. [Google Scholar] [CrossRef]
- Lo, A.K.; Dawson, C.W.; Jin, D.Y.; Lo, K.W. The pathological roles of BART miRNAs in nasopharyngeal carcinoma. J. Pathol. 2012, 227, 392–403. [Google Scholar] [CrossRef]
- Kanda, T.; Miyata, M.; Kano, M.; Kondo, S.; Yoshizaki, T.; Iizasa, H. Clustered microRNAs of the Epstein-Barr virus cooperatively downregulate an epithelial cell-specific metastasis suppressor. J. Virol. 2015, 89, 2684–2697. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.T.; Thorvaldsdottir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, N.; Yoshiyama, H.; Takada, K. Clonal propagation of Epstein-Barr virus (EBV) recombinants in EBV-negative Akata cells. J. Virol. 1996, 70, 7260–7263. [Google Scholar]
- Wang, D.; Liebowitz, D.; Wang, F.; Gregory, C.; Rickinson, A.; Larson, R.; Springer, T.; Kieff, E. Epstein-Barr virus latent infection membrane protein alters the human B-lymphocyte phenotype: Deletion of the amino terminus abolishes activity. J. Virol. 1988, 62, 4173–4184. [Google Scholar] [PubMed]
- Takada, K.; Horinouchi, K.; Ono, Y.; Aya, T.; Osato, T.; Takahashi, M.; Hayasaka, S. An Epstein-Barr virus-producer line Akata: Establishment of the cell line and analysis of viral DNA. Virus Genes 1991, 5, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Furuse, Y.; Oshitani, H.; Kiyono, T. Highly efficient CRISPR/Cas9-mediated cloning and functional characterization of gastric cancer-derived Epstein-Barr virus strains. J. Virol. 2016, 90, 4383–4393. [Google Scholar] [CrossRef] [PubMed]
- Palser, A.L.; Grayson, N.E.; White, R.E.; Corton, C.; Correia, S.; Ba Abdullah, M.M.; Watson, S.J.; Cotten, M.; Arrand, J.R.; Murray, P.G.; et al. Genome diversity of Epstein-Barr virus from multiple tumor types and normal infection. J. Virol. 2015, 89, 5222–5237. [Google Scholar] [CrossRef] [PubMed]
- Kim do, N.; Seo, M.K.; Choi, H.; Kim, S.Y.; Shin, H.J.; Yoon, A.R.; Tao, Q.; Rha, S.Y.; Lee, S.K. Characterization of naturally Epstein-Barr virus-infected gastric carcinoma cell line YCCEL1. J. Gen. Virol. 2013, 94, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.T.; Seo, J.S.; Moon, U.Y.; Kang, K.H.; Shin, D.J.; Yoon, S.K.; Kim, W.H.; Park, J.G.; Lee, S.K. A naturally derived gastric cancer cell line shows latency I Epstein-Barr virus infection closely resembling EBV-associated gastric cancer. Virology 2004, 320, 330–336. [Google Scholar] [CrossRef]
- Tsai, M.H.; Raykova, A.; Klinke, O.; Bernhardt, K.; Gartner, K.; Leung, C.S.; Geletneky, K.; Sertel, S.; Munz, C.; Feederle, R.; et al. Spontaneous lytic replication and epitheliotropism define an Epstein-Barr virus strain found in carcinomas. Cell Rep. 2013, 5, 458–470. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Cherng, J.M.; Lin, H.J.; Tsang, C.W.; Liu, Y.X.; Lee, S.P. Amino acid changes in functional domains of latent membrane protein 1 of Epstein-Barr virus in nasopharyngeal carcinoma of southern China and Taiwan: Prevalence of an HLA A2-restricted ‘epitope-loss variant’. J. Gen. Virol. 2004, 85, 2023–2034. [Google Scholar] [CrossRef]
- Wang, Y.; Kanai, K.; Satoh, Y.; Luo, B.; Sairenji, T. Carboxyl-terminal sequence variation of latent membrane protein 1 gene in Epstein-Barr virus-associated gastric carcinomas from Eastern China and Japan. Intervirology 2007, 50, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Lo, A.K.; To, K.F.; Lo, K.W.; Lung, R.W.; Hui, J.W.; Liao, G.; Hayward, S.D. Modulation of LMP1 protein expression by EBV-encoded microRNAs. Proc. Natl. Acad. Sci. USA 2007, 104, 16164–16169. [Google Scholar] [CrossRef] [Green Version]
- Verhoeven, R.J.; Tong, S.; Zhang, G.; Zong, J.; Chen, Y.; Jin, D.Y.; Chen, M.R.; Pan, J.; Chen, H. NF-kappaB Signaling Regulates Expression of Epstein-Barr Virus BART MicroRNAs and Long Noncoding RNAs in Nasopharyngeal Carcinoma. J. Virol. 2016, 90, 6475–6488. [Google Scholar] [CrossRef]
- Lin, X.; Tsai, M.H.; Shumilov, A.; Poirey, R.; Bannert, H.; Middeldorp, J.M.; Feederle, R.; Delecluse, H.J. The Epstein-Barr Virus BART miRNA Cluster of the M81 Strain Modulates Multiple Functions in Primary B Cells. PLoS Pathog. 2015, 11, e1005344. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.J.; Zhou, R.; Zong, J.F.; Lin, W.S.; Tong, S.; Guo, Q.J.; Lin, C.; Lin, S.J.; Chen, Y.X.; Chen, M.R.; et al. Epstein-Barr virus-coded miR-BART13 promotes nasopharyngeal carcinoma cell growth and metastasis via targeting of the NKIRAS2/NF-kappaB pathway. Cancer Lett. 2019, 447, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Feederle, R.; Bartlett, E.J.; Delecluse, H.J. Epstein-Barr virus genetics: Talking about the BAC generation. Herpesviridae 2010, 1, 6. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.H.; Lin, X.; Shumilov, A.; Bernhardt, K.; Feederle, R.; Poirey, R.; Kopp-Schneider, A.; Pereira, B.; Almeida, R.; Delecluse, H.J. The biological properties of different Epstein-Barr virus strains explain their association with various types of cancers. Oncotarget 2017, 8, 10238–10254. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HEK293 Cells Transfected with | BART(+)LMP1B95-8 | BART(+)LMP1CAO |
|---|---|---|
| No. of cell clones screened | 16 | 28 |
| No. of virus-producing cell clones obtained | 2 | 5 |
| infection efficiencies of progeny viruses to B cell lines | 10.0% 15.6% | 10.5% 11.5% 15.3% 16.5% 29.2% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yajima, M.; Miyata, M.; Ikuta, K.; Hasegawa, Y.; Oneyama, C.; Kanda, T. Efficient Epstein-Barr Virus Progeny Production Mediated by Cancer-Derived LMP1 and Virally-Encoded microRNAs. Microorganisms 2019, 7, 119. https://doi.org/10.3390/microorganisms7050119
Yajima M, Miyata M, Ikuta K, Hasegawa Y, Oneyama C, Kanda T. Efficient Epstein-Barr Virus Progeny Production Mediated by Cancer-Derived LMP1 and Virally-Encoded microRNAs. Microorganisms. 2019; 7(5):119. https://doi.org/10.3390/microorganisms7050119
Chicago/Turabian StyleYajima, Misako, Mamiko Miyata, Kazufumi Ikuta, Yasuhisa Hasegawa, Chitose Oneyama, and Teru Kanda. 2019. "Efficient Epstein-Barr Virus Progeny Production Mediated by Cancer-Derived LMP1 and Virally-Encoded microRNAs" Microorganisms 7, no. 5: 119. https://doi.org/10.3390/microorganisms7050119
APA StyleYajima, M., Miyata, M., Ikuta, K., Hasegawa, Y., Oneyama, C., & Kanda, T. (2019). Efficient Epstein-Barr Virus Progeny Production Mediated by Cancer-Derived LMP1 and Virally-Encoded microRNAs. Microorganisms, 7(5), 119. https://doi.org/10.3390/microorganisms7050119