The Degradative Capabilities of New Amycolatopsis Isolates on Polylactic Acid


 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacterial Strains and Media
2.2.1. Strains
2.2.2. Media
2.3. Actinomycete Isolation from Soils
2.4. Screening of Potential PLA-Degrading Actinomycetes
2.5. Identification of Potential PLA-Degrading Actinomycetes
2.6. Growth at Different Temperatures
2.7. PLA Biodegradation
2.7.1. Utilization of Lactic Acid as a Carbon Source
2.7.2. PLA Biofragmentation
2.7.3. PLA Biodeterioration
Weight Loss of PLA Films
Effect of Biodeterioration on the Physical Features of PLA Film: Visual Inspection and Environmental Scanning Electron Microscopy (ESEM) Analysis
2.7.4. Statistical Analysis
3. Results and Discussion
3.1. Isolation of Actinomycetes able to Degrade Emulsified PLA
3.2. Actinomycete Identification and Growth at Different Temperatures
3.3. Degradation of PLA
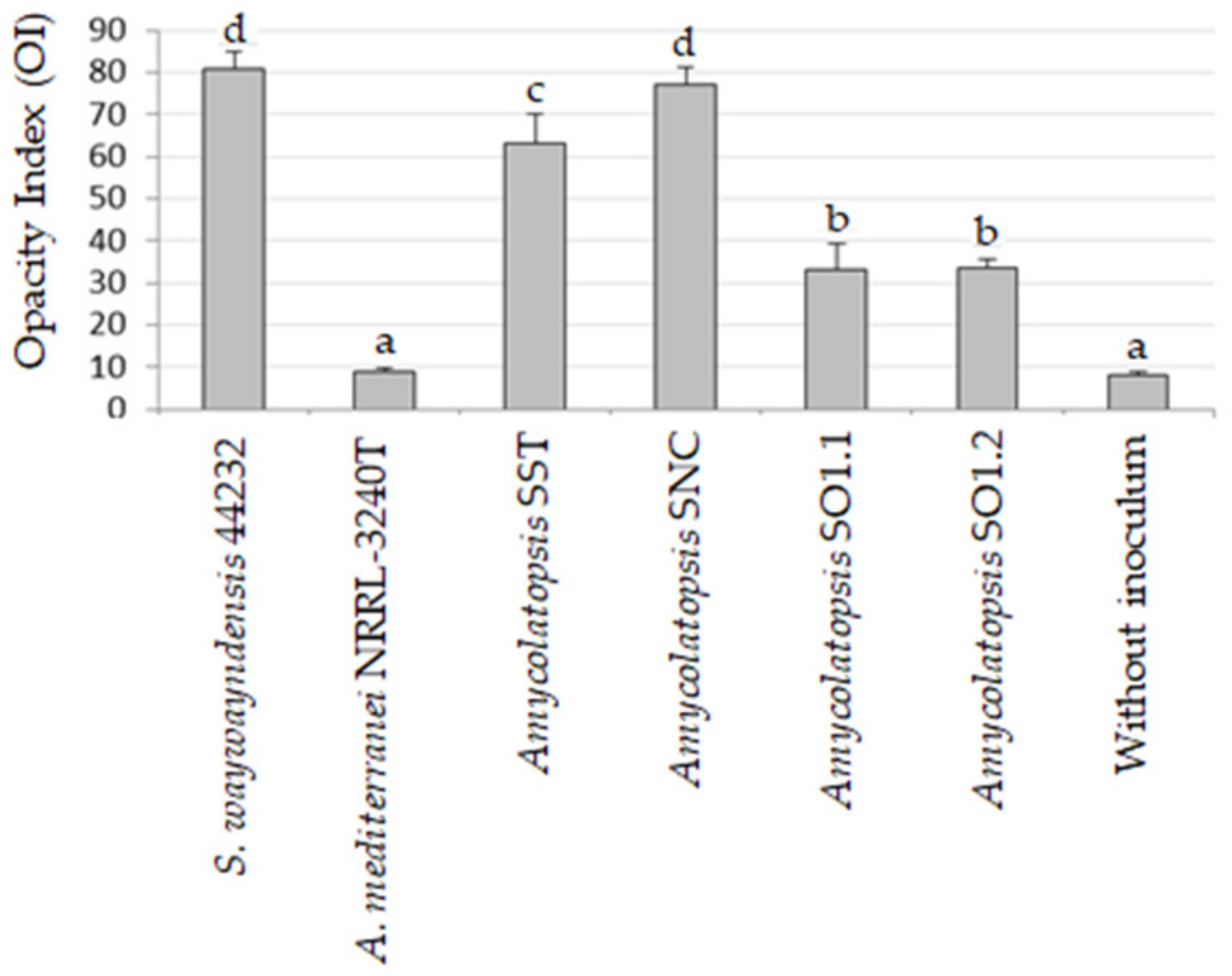
3.3.1. Biofragmentation
3.3.2. Assimilation
3.3.3. Biodeterioration
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Garlotta, D. Literature Review of Poly(Lactic Acid). J. Polym. Environ. 2002, 9, 63–84. [Google Scholar] [CrossRef]
- Karamanlioglu, M.; Preziosi, R.; Robson, G.D. Abiotic and biotic environmental degradation of the bioplastic polymer poly(lactic acid): A review. Polym. Degrad. Stab. 2017, 137, 122–130. [Google Scholar] [CrossRef]
- Kim, M.Y.; Kim, C.; Moon, J.; Heo, J.; Jung, S.P.; Kim, J.R. Polymer film-based screening and isolation of polylactic acid (PLA)-degrading microorganisms. J. Microbiol. Biotechnol. 2017, 27, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Emadian, S.M.; Onay, T.T.; Demirel, B. Biodegradation of bioplastics in natural environments. Waste Manag. 2017, 59, 526–536. [Google Scholar] [CrossRef]
- Tokiwa, Y.; Calabia, B.P. Biodegradability and biodegradation of poly(lactide). Appl. Microbiol. Biotechnol. 2006, 72, 244–251. [Google Scholar] [CrossRef]
- Jarerat, A.; Pranamuda, H.; Tokiwa, Y. Poly(L-lactide)-degrading activity in various actinomycetes. Macromol. Biosci. 2002, 2, 420–428. [Google Scholar] [CrossRef]
- Tiwari, K.; Gupta, R.K. Rare actinomycetes: a potential storehouse for novel antibiotics. Crit. Rev. Biotechnol. 2012, 32, 108–132. [Google Scholar] [CrossRef]
- Lazzarini, A.; Cavaletti, L.; Toppo, G.; Marinelli, F. Rare genera of actinomycetes as potential producers of new antibiotics. Anton. Leeuw. Int. J. Gen. Mol. Microbiol. 2000, 78, 399–405. [Google Scholar] [CrossRef]
- Mukhtar, S.; Zaheer, A.; Aiysha, D.; Abdulla Malik, K.; Mehnaz, S. Actinomycetes: A Source of Industrially Important Enzymes. J. Proteomics Bioinform. 2017, 10, 12. [Google Scholar] [CrossRef]
- Bubpachat, T.; Sombatsompop, N.; Prapagdee, B. Isolation and role of polylactic acid-degrading bacteria on degrading enzymes productions and PLA biodegradability at mesophilic conditions. Polym. Degrad. Stab. 2018, 152, 75–85. [Google Scholar] [CrossRef]
- Jeon, H.J.; Kim, M.N. Biodegradation of poly(L-lactide) (PLA) exposed to UV irradiation by a mesophilic bacterium. Int. Biodeterior. Biodegradation 2013, 85, 289–293. [Google Scholar] [CrossRef]
- Tomita, K.; Kuroki, Y.; Nagai, K. Isolation of thermophiles degrading poly(L-lactic acid). J. Biosci. Bioeng. 1999, 87, 752–755. [Google Scholar] [CrossRef]
- Sakai, K.; Kawano, H.; Iwami, A.; Nakamura, M.; Moriguchi, M. Isolation of a thermophilic poly-L-lactide degrading bacterium from compost and its enzymatic characterization. J. Biosci. Bioeng. 2001, 92, 298–300. [Google Scholar] [CrossRef]
- Tomita, K.; Nakajima, T.; Kikuchi, Y.; Miwa, N. Degradation of poly(L-lactic acid) by a newly isolated thermophile. Polym. Degrad. Stab. 2004, 84, 433–438. [Google Scholar] [CrossRef]
- Kim, M.N.; Kim, W.G.; Weon, H.Y.; Lee, S.H. Poly(L-lactide)-degrading activity of a newly isolated bacterium. J. Appl. Polym. Sci. 2008, 109, 234–239. [Google Scholar] [CrossRef]
- Pranamuda, H.; Tokiwa, Y.; Tanaka, H. Polylactide degradation by an Amycolatopsis sp. Appl. Environ. Microbiol. 1997, 63, 1637–1640. [Google Scholar]
- Kim, M.N.; Park, S.T. Degradation of poly(L-lactide) by a mesophilic bacterium. J. Appl. Polym. Sci. 2010, 117, 67–74. [Google Scholar] [CrossRef]
- Nakamura, K.; Tomita, T.; Abe, N.; Kamio, Y. Purification and characterization of an extracellular poly(L-Lactic Acid) depolymerase from a soil isolate, Amycolatopsis sp. strain K104-1. Appl. Environ. Microbiol. 2001, 67, 345–353. [Google Scholar] [CrossRef]
- Shah, A.A.; Hasan, F.; Hameed, A.; Ahmed, S. Biological degradation of plastics: A comprehensive review. Biotechnol. Adv. 2008, 26, 246–265. [Google Scholar] [CrossRef]
- Sukkhum, S.; Tokuyama, S.; Tamura, T.; Kitpreechavanich, V. A novel poly (L-lactide) degrading actinomycetes isolated from Thai forest soil, phylogenic relationship and the enzyme characterization. J. Gen. Appl. Microbiol. 2009, 55, 459–467. [Google Scholar] [CrossRef]
- Teeraphatpornchai, T.; Nakajima-Kambe, T.; Shigeno-Akutsu, Y.; Nakayama, M.; Nomura, N.; Nakahara, T.; Uchiyama, H. Isolation and characterization of a bacterium that degrades various polyester-based biodegradable plastics. Biotechnol. Lett. 2003, 25, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Gorrasi, G.; Pantani, R. Effect of PLA grades and morphologies on hydrolytic degradation at composting temperature: Assessment of structural modification and kinetic parameters. Polym. Degrad. Stab. 2013, 98, 1006–1014. [Google Scholar] [CrossRef]
- Jarerat, A.; Tokiwa, Y. Poly(L-lactide) degradation by Saccharothrix waywayandensis. Biotechnol. Lett. 2003, 25, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Pranamuda, H.; Tokiwa, Y. Degradation of poly(L-lactide) by strains belonging to genus Amycolatopsis. Biotechnol. Lett. 1999, 21, 901–905. [Google Scholar] [CrossRef]
- Subramani, R.; Aalbersberg, W. Culturable rare actinomycetes: Diversity, isolation and marine natural product discovery. Appl. Microbiol. Biotechnol. 2013, 97, 9291–9321. [Google Scholar] [CrossRef]
- Wang, D.S.; Xue, Q.H.; Zhu, W.J.; Zhao, J.; Duan, J.L.; Shen, G.H. Microwave irradiation is a useful tool for improving isolation of actinomycetes from soil. Mikrobiologiia 2013, 82, 106–114. [Google Scholar]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution. 1985, 39, 783. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Zucchi, T.D.; Tan, G.Y.A.; Goodfellow, M. Amycolatopsis thermophila sp. nov. and Amycolatopsis viridis sp. nov., thermophilic actinomycetes isolated from arid soil. Int. J. Syst. Evol. Microbiol. 2012, 62, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, K.; Gupta, R.K. Diversity and isolation of rare actinomycetes: An overview. Crit. Rev. Microbiol. 2013, 39, 256–294. [Google Scholar] [CrossRef]
- Sacherer, P.; Défago, G.; Haas, D. Extracellular protease and phospholipase C are controlled by the global regulatory gene gacA in the biocontrol strain Pseudomonas fluorescens CHA0. FEMS Microbiol. Lett. 1994, 116, 155–160. [Google Scholar] [CrossRef]
- Qi, X.; Ren, Y.; Wang, X. New advances in the biodegradation of poly(lactic) acid. Int. Biodeterior. Biodegrad. 2017, 117, 215–223. [Google Scholar] [CrossRef]
- Li, F.; Wang, S.; Liu, W.; Chen, G. Purification and characterization of poly(L-lactic acid)-degrading enzymes from Amycolatopsis orientalis ssp. orientalis. FEMS Microbiol. Lett. 2008, 282, 52–58. [Google Scholar] [CrossRef]
- Chomchoei, A.; Pathom-aree, W.; Yokota, A.; Kanongnuch, C.; Lumyong, S. Amycolatopsis thailandensis sp. nov., a poly(L-lactic acid)-degrading actinomycete, isolated from soil. Int. J. Syst. Evol. Microbiol. 2011, 61, 839–843. [Google Scholar] [CrossRef]
- Tokiwa, Y.; Jarerat, A. Biodegradation of poly(L-lactide). Biotechnol. Lett. 2004, 26, 771–777. [Google Scholar] [CrossRef]
- Tang, B.; Xie, F.; Zhao, W.; Wang, J.; Dai, S.; Zheng, H.; Ding, X.; Cen, X.; Liu, H.; Yu, Y.; et al. A systematic study of the whole genome sequence of Amycolatopsis methanolica strain 239 T provides an insight into its physiological and taxonomic properties which correlate with its position in the genus. Synth. Syst. Biotechnol. 2016, 1, 169–186. [Google Scholar] [CrossRef]
- Sánchez-Hidalgo, M.; González, I.; Díaz-Muñoz, C.; Martínez, G.; Genilloud, O. Comparative genomics and biosynthetic potential analysis of two lichen-isolated Amycolatopsis strains. Front. Microbiol. 2018, 9, 369. [Google Scholar] [CrossRef] [PubMed]
- Everest, G.J.; Meyers, P.R. The use of gyrB sequence analysis in the phylogeny of the genus Amycolatopsis. Antonie Van Leeuw. J. Microb. 2009, 95, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jarerat, A.; Tokiwa, Y.; Tanaka, H. Microbial poly(L-lactide)-degrading enzyme induced by amino acids, peptides, and poly(L-amino acids). J. Polym. Environ. 2004, 12, 139–146. [Google Scholar] [CrossRef]
- Lucas, N.; Bienaime, C.; Belloy, C.; Queneudec, M.; Silvestre, F.; Nava-Saucedo, J.-E. Polymer biodegradation: Mechanisms and estimation techniques—A review. Chemosphere 2008, 73, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Satti, S.M.; Shah, A.A.; Auras, R.; Marsh, T.L. Isolation and characterization of bacteria capable of degrading poly(lactic acid) at ambient temperature. Polym. Degrad. Stab. 2017, 144, 392–400. [Google Scholar] [CrossRef]
- Tokiwa, Y.; Calabia, B.; Ugwu, C.; Aiba, S. Biodegradability of plastics. Int. J. Mol. Sci. 2009, 10, 3722–3742. [Google Scholar] [CrossRef]
- Penkhrue, W.; Khanongnuch, C.; Masaki, K.; Pathom-aree, W.; Punyodom, W.; Lumyong, S. Isolation and screening of biopolymer-degrading microorganisms from northern Thailand. World J. Microbiol. Biotechnol. 2015, 31, 1431–1442. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Number of Soils Used | Number of Bacteria Positive for the Clear Zone Test |
|---|---|---|
| Tomita et al. 2004 [14] | 153 | 1 |
| Teeraphatpornchai et al. 2003 [21] | 400 | 4 |
| Sukkhum et al. 2009 [20] | 80 | 13 |
| Nakamura et al. 2001 [18] | 300 | 2 |
| Pranamuda et al. 1997 [16] | 45 | 1 |
| Kim and Park 2010 [17] | 60 | 1 |
| BM | ||
|---|---|---|
| Strain | without Gelatin | with 0.1% Gelatin |
| Lentzea waywayandensis DSM 44232 | NS a | 7.2 ± 7.2 * |
| Amycolatopsis sp. SST | NS | 27.9 ± 9.9 * |
| Amycolatopsis sp. SNC | 6.7 ± 2.7 * | 11.8 ± 6.7 * |
| Amycolatopsis sp. SO1.2 | NS | 11.0 ± 13.5 * |
| Amycolatopsis sp. SO1.1 | NS | 40.4 ± 21.8 * |
| Strain | Carbon Source | |
|---|---|---|
| d-Glucose | l-Lactic Acid | |
| L. waywayandensis DSM 44232 | +++ | + |
| Amycolatopsis mediterranei NRRL B-3240T | +++ | - |
| Amycolatopsis sp. SNC | +++ | ++ |
| Amycolatopsis sp. SST | +++ | + |
| Amycolatopsis sp. SO1.1 | +++ | - |
| Amycolatopsis sp. SO1.2 | +++ | + |
| BM | SEM | |||
|---|---|---|---|---|
| Strain | without Gelatin | with 0.1% Gelatin | without Gelatin | with 0.1% Gelatin |
| L. waywayandensis DSM 44232 | NS a | 37.3 ± 19.4 * | NS | NS |
| A. mediterranei NRRL B-3240T | NS | NS | NS | NS |
| Amycolatopsis sp. SST | NS | NS | NS | NS |
| Amycolatopsis sp. SNC | NS | NS | NS | 36.0 ± 7.3 * |
| Amycolatopsis sp. SO1.2 | NS | NS | NS | NS |
| Amycolatopsis sp. SO1.1 | NS | NS | NS | NS |
| Without inoculum | NS | NS | NS | NS |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Decorosi, F.; Exana, M.L.; Pini, F.; Adessi, A.; Messini, A.; Giovannetti, L.; Viti, C. The Degradative Capabilities of New Amycolatopsis Isolates on Polylactic Acid. Microorganisms 2019, 7, 590. https://doi.org/10.3390/microorganisms7120590
Decorosi F, Exana ML, Pini F, Adessi A, Messini A, Giovannetti L, Viti C. The Degradative Capabilities of New Amycolatopsis Isolates on Polylactic Acid. Microorganisms. 2019; 7(12):590. https://doi.org/10.3390/microorganisms7120590
Chicago/Turabian StyleDecorosi, Francesca, Maria Luna Exana, Francesco Pini, Alessandra Adessi, Anna Messini, Luciana Giovannetti, and Carlo Viti. 2019. "The Degradative Capabilities of New Amycolatopsis Isolates on Polylactic Acid" Microorganisms 7, no. 12: 590. https://doi.org/10.3390/microorganisms7120590
APA StyleDecorosi, F., Exana, M. L., Pini, F., Adessi, A., Messini, A., Giovannetti, L., & Viti, C. (2019). The Degradative Capabilities of New Amycolatopsis Isolates on Polylactic Acid. Microorganisms, 7(12), 590. https://doi.org/10.3390/microorganisms7120590