Life in High Salt Concentrations with Changing Environmental Conditions: Insights from Genomic and Phenotypic Analysis of Salinivibrio sp.


Abstract

1. Introduction
2. Material and Methods
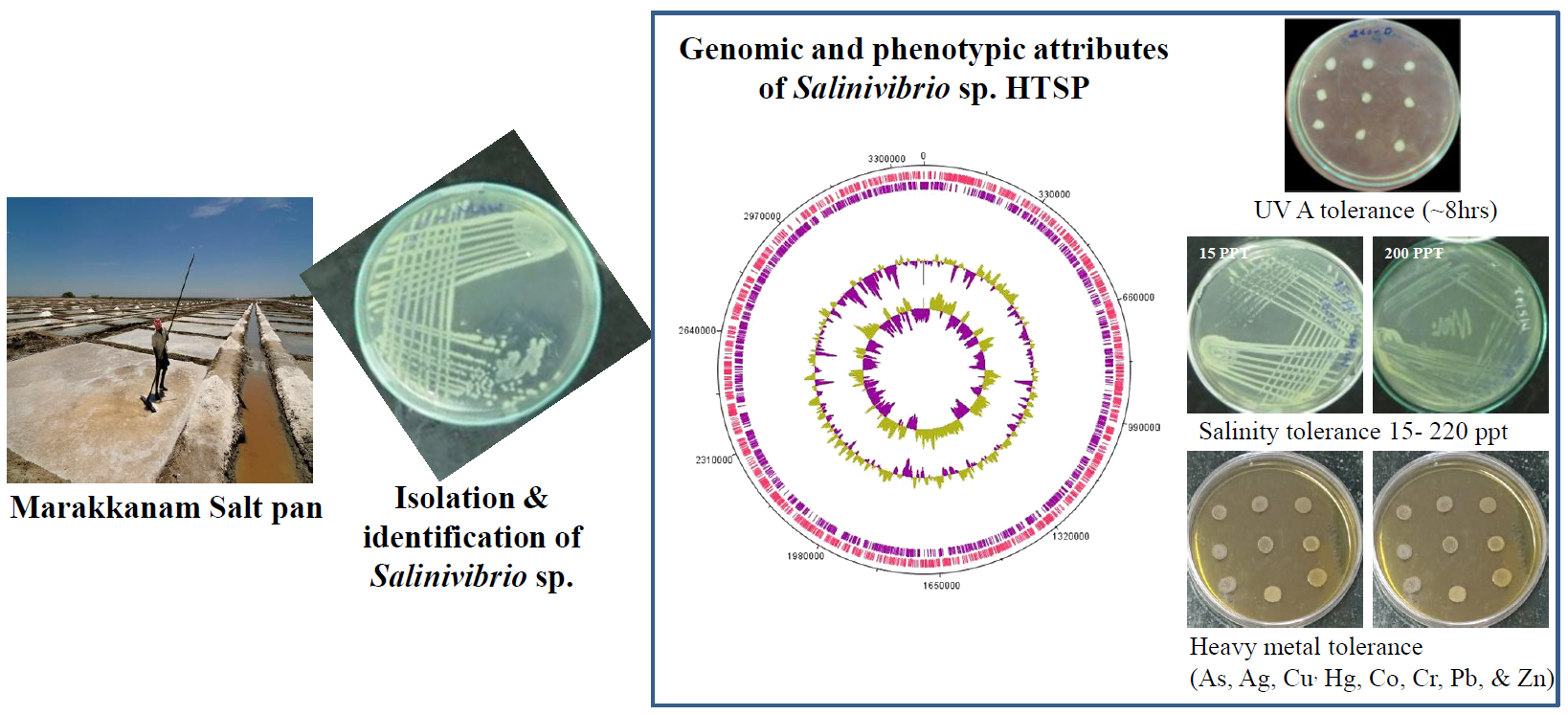
2.1. Isolation and Identification of the Strain
2.2. IlluminaNextSeq-Based Genome Sequencing
2.3. Genomic-Based Taxonomic Resolution
2.4. Salinity Tolerance
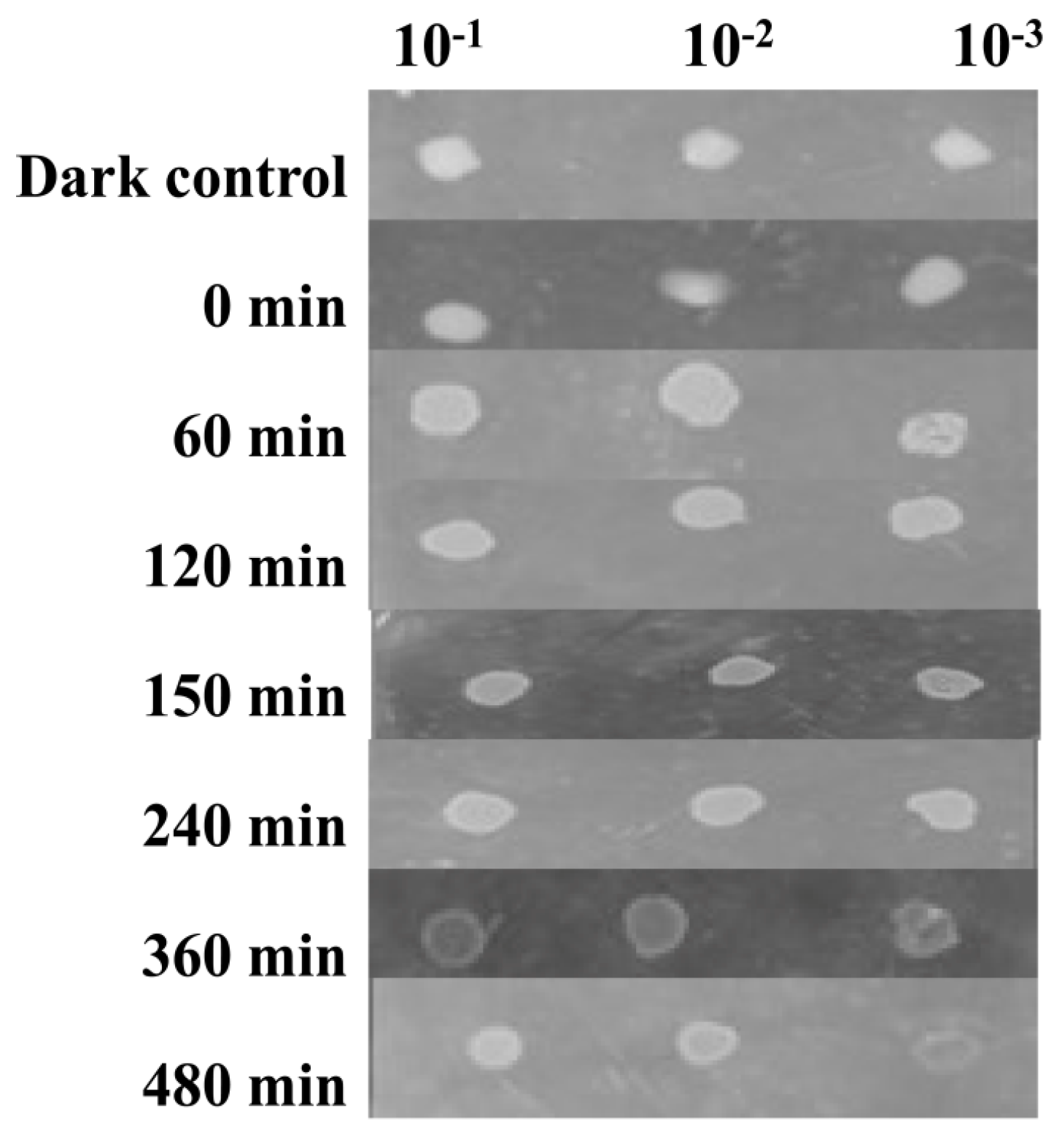
2.5. UV Tolerance
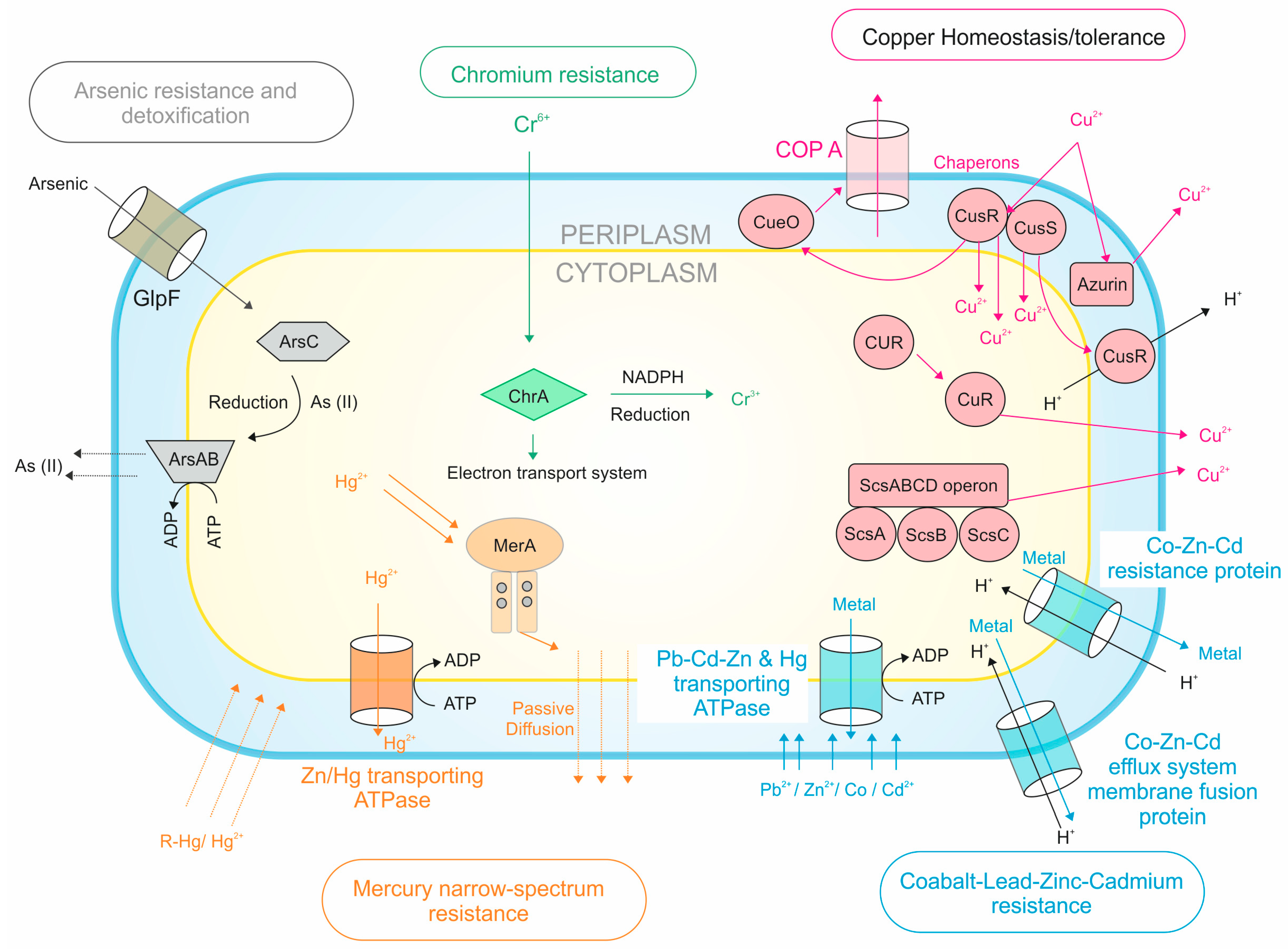
2.6. Heavy Metal Tolerance
3. Results
3.1. General Features of Salinivibrio sp. HTSP
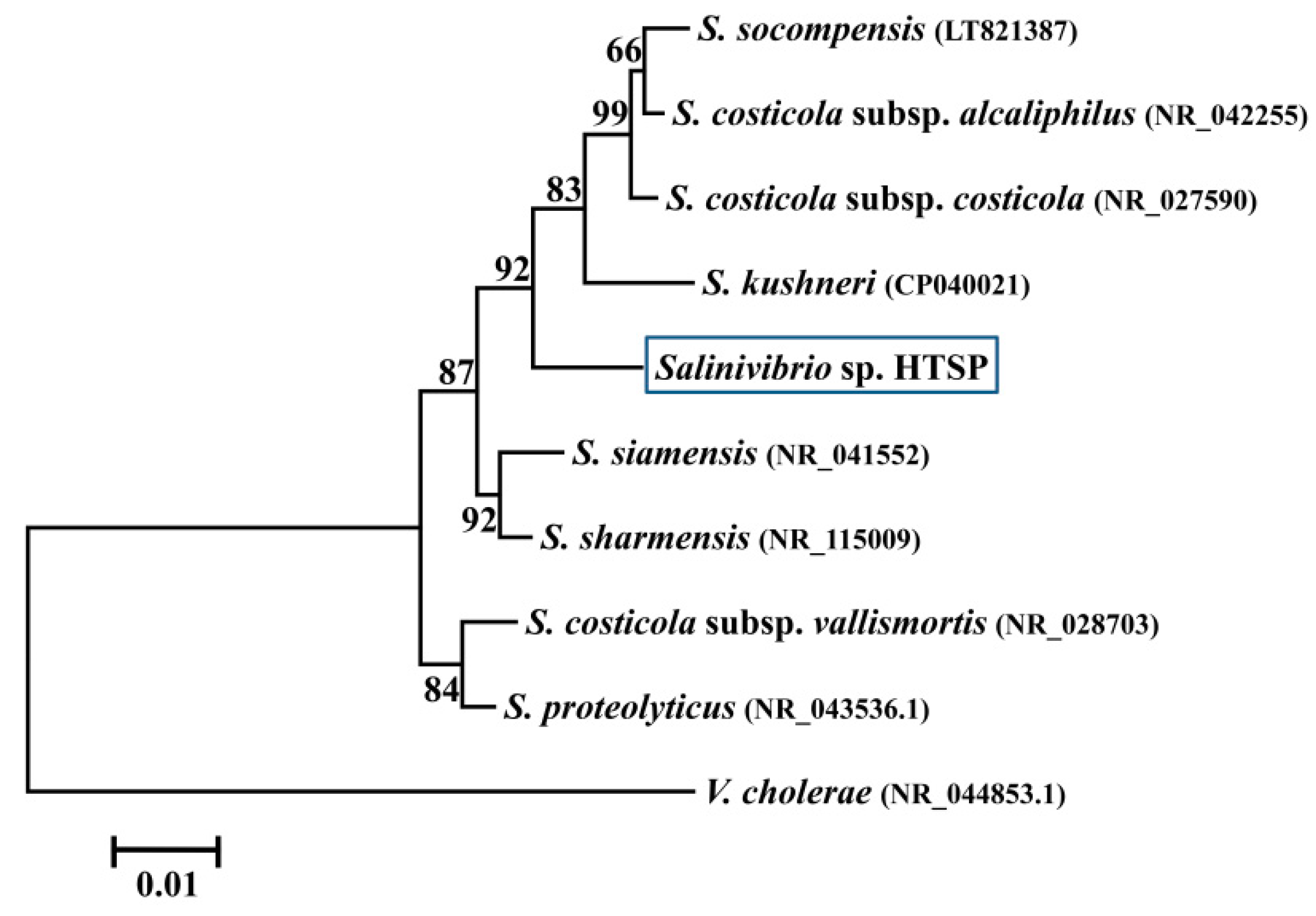
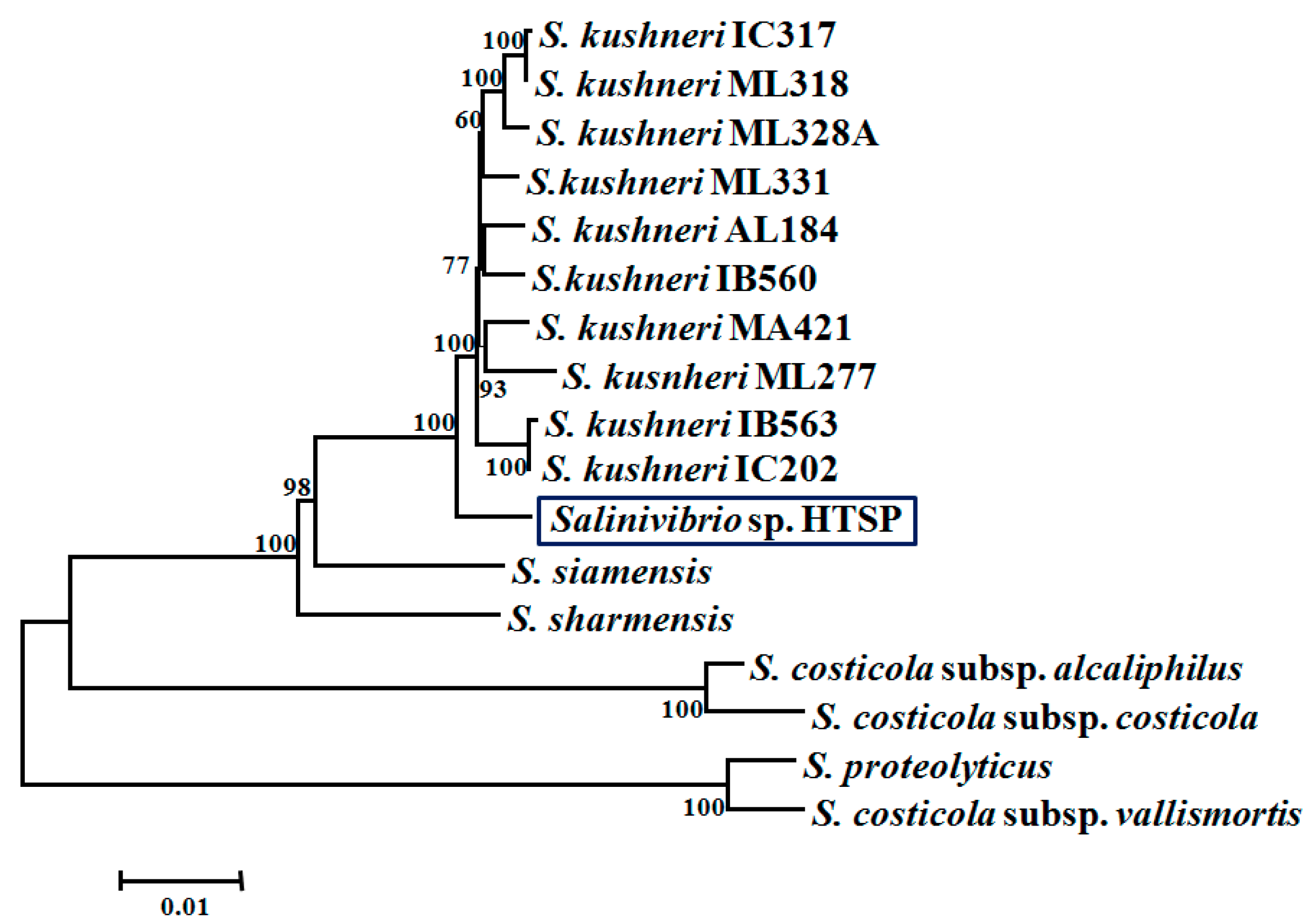
3.2. Taxonomic Assignment and Core Gene Phylogeny
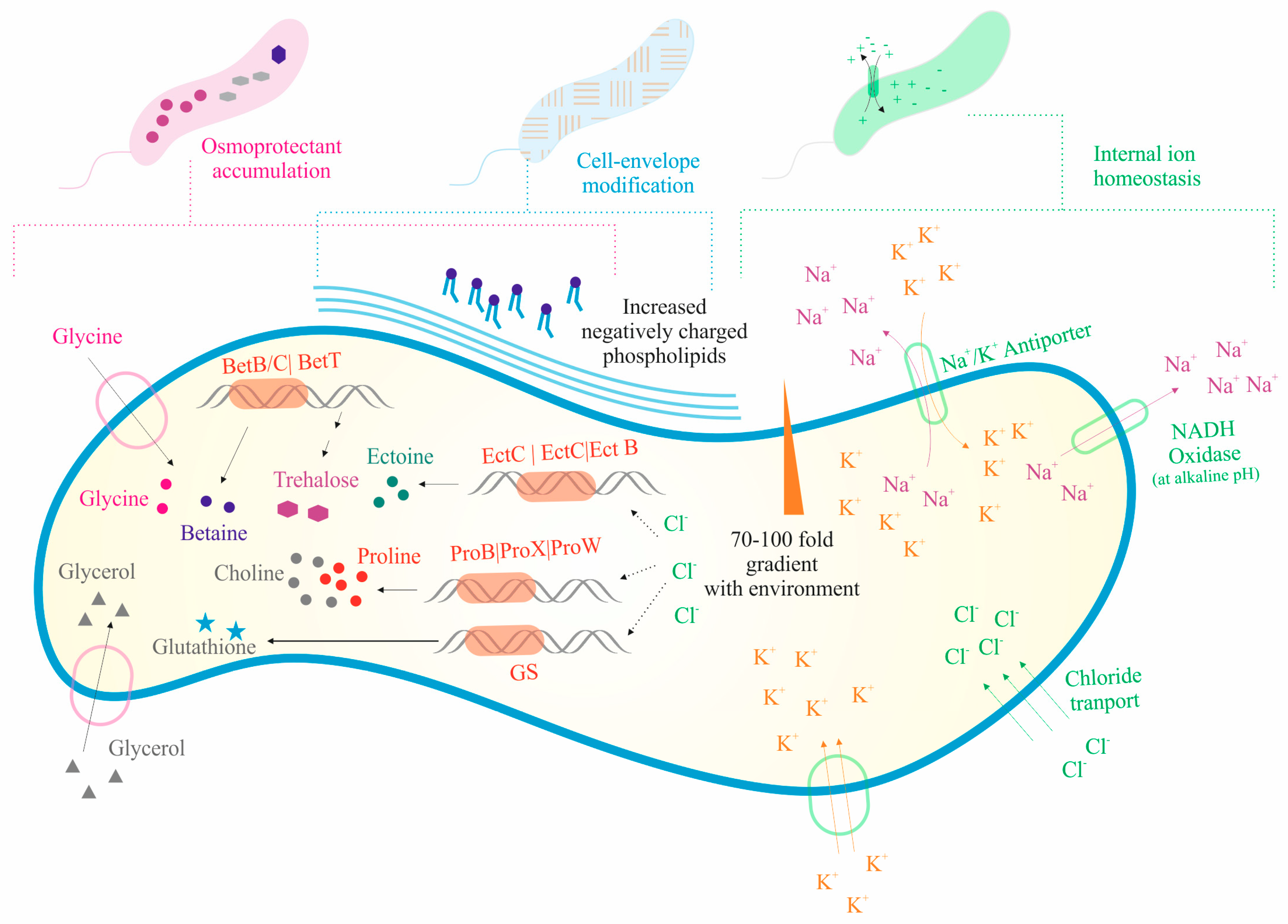
3.3. Salinity Range and Genes Related to Halotolerance
3.4. Resistance to UV-A Radiation
3.5. Heavy Metal Tolerance
3.6. General Stress-Related Genes
4. Discussion
4.1. Strategies for Halo Adaptation
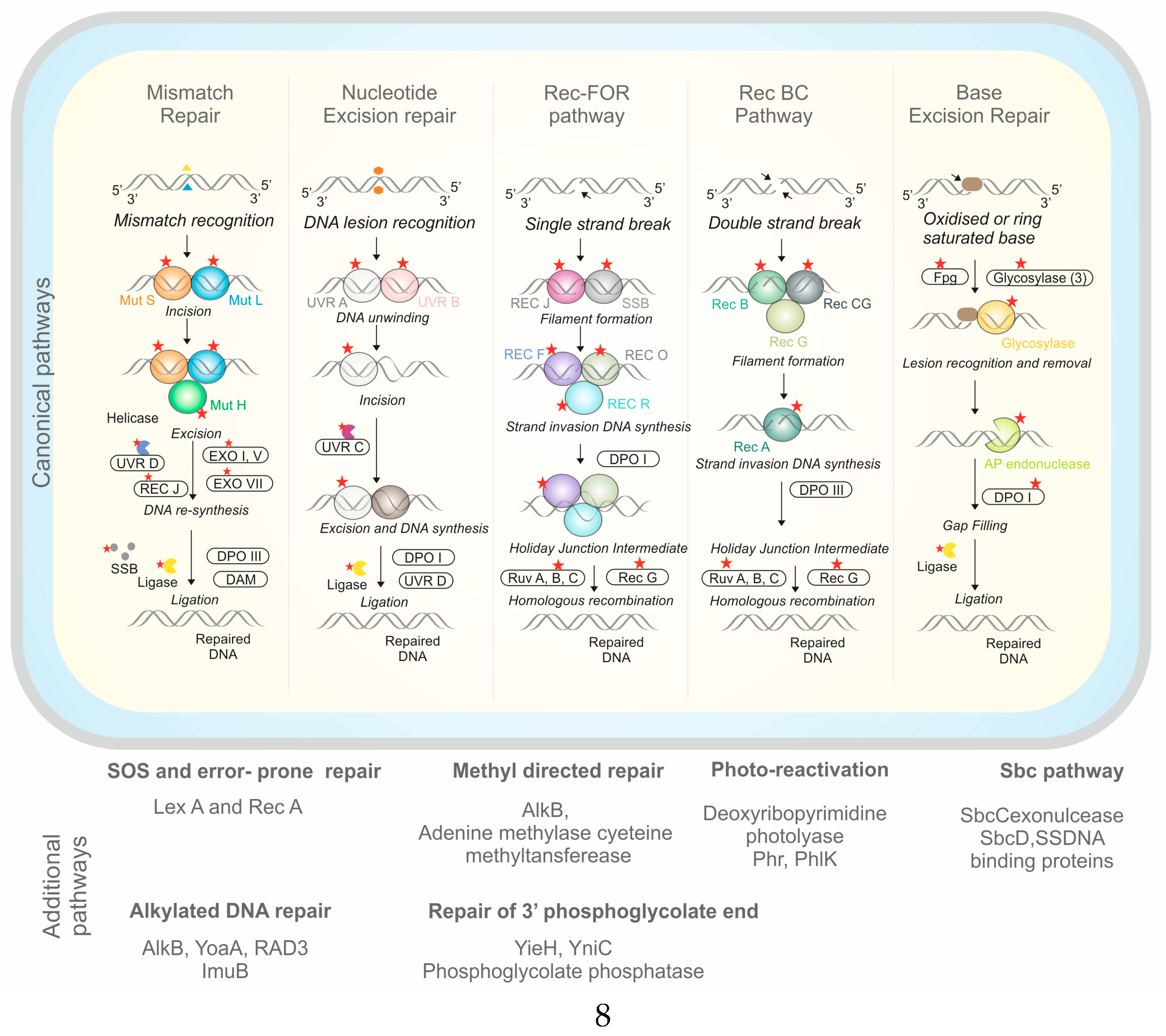
4.2. Genomic Basis of UV Tolerance
4.3. Heavy Metal Tolerance
4.4. General Stress-Responsive Genes
4.4.1. Periplasmic Stress Response
4.4.2. Oxidative Stress Response
5. Conclusion
Reference
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, D.L.; Baxter, B.K. DNA repair and photoprotection: Mechanisms of overcoming environmental ultraviolet radiation exposure in halophilic archaea. Front. Microbiol. 2017, 8, 1882. [Google Scholar] [CrossRef]
- Agogué, H.; Joux, F.; Obernosterer, I.; Lebaron, P. Resistance of marine bacterioneuston to solar radiation. Appl. Environ. Microbiol. 2005, 71, 5282–5289. [Google Scholar] [CrossRef] [PubMed]
- Flores, M.R.; Ordoñez, O.F.; Maldonado, M.J.; Farías, M.E. Isolation of UV-B resistant bacteria from two high altitude Andean lakes (4400 m) with saline and non saline conditions. J. Gen. Appl Microbiol. 2009, 55, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Krishna, P.S.; Sreenivas, A.; Singh, D.K.; Shivaji, S.; Prakash, J.S. Draft genome sequence of Bacillus okhensis Kh10-101T, a halo-alkali tolerant bacterium from Indian salt pan. Genom. Data 2015, 6, 283–284. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Paraneeiswaran, A.; Shukla, S.K.; Prashanth, K.; Rao, T.S. Microbial reduction of [Co (III)–EDTA]− by Bacillus licheniformis SPB-2 strain isolated from a solar salt pan. J. Hazard. Mater. 2015, 283, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Dey, R.; Pal, K.K.; Sherathia, D.; Dalsania, T.; Savsani, K.; Patel, I.; Sukhadiya, B.; Mandaliya, M.; Thomas, M.; Ghorai, S. Draft genome sequence of Bacillus sp. strain NSP2. 1, a nonhalophilic bacterium isolated from the salt marsh of the Great Rann of Kutch, India. Genome Announc. 2013, 1, e00909–e00913. [Google Scholar] [PubMed]
- Galisteo, C.; Sánchez-Porro, C.; de la Haba, R.R.; López-Hermoso, C.; Fernández, A.B.; Farias, M.E.; Ventosa, A. Characterization of Salinivibrio socompensis sp. nov., A New Halophilic Bacterium Isolated from the High-Altitude Hypersaline Lake Socompa, Argentina. Microorganisms 2019, 7, 241. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-Y.; Garcia, J.-L.; Patel, B.; Cayol, J.-L.; Baresi, L.; Mah, R.A. Salinivibrio costicola subsp. vallismortis subsp. nov., a halotolerant facultative anaerobe from Death Valley, and emended description of Salinivibrio costicola. Int. J. Syst. Evol. Microbiol. 2000, 50, 615–622. [Google Scholar] [CrossRef]
- López-Hermoso, C.; Rafael, R.; Sánchez-Porro, C.; Bayliss, S.C.; Feil, E.J.; Ventosa, A. Draft Genome Sequences of Salinivibrio proteolyticus, Salinivibrio sharmensis, Salinivibrio siamensis, Salinivibrio costicola subsp. alcaliphilus, Salinivibrio costicola subsp. vallismortis, and 29 New Isolates Belonging to the Genus Salinivibrio. Genome Announc. 2017, 5, e00244–e00247. [Google Scholar]
- Romano, I.; Orlando, P.; Gambacorta, A.; Nicolaus, B.; Dipasquale, L.; Pascual, J.; Giordano, A.; Lama, L. Salinivibrio sharmensis sp. nov., a novel haloalkaliphilic bacterium from a saline lake in Ras Mohammed Park (Egypt). Extremophiles 2011, 15, 213–220. [Google Scholar] [CrossRef]
- Ventosa, A.; Nieto, J.J.; Oren, A. Biology of moderately halophilic aerobic bacteria. Microbiol. Mol. Biol. Rev. 1998, 62, 504–544. [Google Scholar] [PubMed]
- Mellado, E.; Moore, E.; Nieto, J.; Ventosa, A. Analysis of 16S rRNA gene sequences of Vibrio costicola strains: Description of Salinivibrio costicola gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 1996, 46, 817–821. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.; Nieto, J.; Ventosa, A.; Ruiz-Berraquero, F. The susceptibility of the moderate halophile Vibrio costicola to heavy metals. J. Appl. Bacteriol. 1987, 63, 63–66. [Google Scholar] [CrossRef]
- Amoozegar, M.A.; Schumann, P.; Hajighasemi, M.; Fatemi, A.Z.; Karbalaei-Heidari, H.R. Salinivibrio proteolyticus sp. nov., a moderately halophilic and proteolytic species from a hypersaline lake in Iran. Int. J. Syst. Evol. Microbiol. 2008, 58, 1159–1163. [Google Scholar] [CrossRef]
- Chamroensaksri, N.; Tanasupawat, S.; Akaracharanya, A.; Visessanguan, W.; Kudo, T.; Itoh, T. Salinivibrio siamensis sp. nov., from fermented fish (pla-ra) in Thailand. Int. J. Syst. Evol. Microbiol. 2009, 59, 880–885. [Google Scholar] [CrossRef] [PubMed]
- López-Hermoso, C.; de la Haba, R.R.; Sánchez-Porro, C.; Papke, R.T.; Ventosa, A. Assessment of MultiLocus Sequence Analysis as a valuable tool for the classification of the genus Salinivibrio. Front. Microbiol. 2017b, 8, 1107. [Google Scholar] [CrossRef]
- Kabilan, M. quot Microbial diversity of halophilic archaea and bacteria in solar salterns and studies on their production of antiarchaeal substancesquot. Ph.D. thesis, BITS, Pilani, India, 2016. [Google Scholar]
- Prescott, L.M.; Sherwood, L.; Woolverton, C. Prescott, Harley, and Klein’s Microbiology; McGraw-Hill Higher Education: New York, NY, USA, 2007. [Google Scholar]
- Ventosa, A.; Quesada, E.; Rodriguez-Valera, F.; Ruiz-Berraquero, F.; Ramos-Cormenzana, A. Numerical taxonomy of moderately halophilic Gram-negative rods. Microbiology 1982, 128, 1959–1968. [Google Scholar] [CrossRef]
- Ravindran, J.; Kannapiran, E.; Manikandan, B.; Francis, K.; Arora, S.; Karunya, E.; Kumar, A.; Singh, S.; Jose, J. UV-absorbing bacteria in coral mucus and their response to simulated temperature elevations. Coral Reefs 2013, 32, 1043–1050. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; pp. 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2013, 42, D206–D214. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Ha, S.-m.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Kim, Y.O.; Park, S.-C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, N.M.; Gupta, V.K.; Dutta, C. BPGA-an ultra-fast pan-genome analysis pipeline. Sci. Rep. 2016, 6, 24373. [Google Scholar] [CrossRef]
- Albarracín, V.H.; Pathak, G.P.; Douki, T.; Cadet, J.; Borsarelli, C.D.; Gärtner, W.; Farias, M.E. Extremophilic Acinetobacter strains from high-altitude lakes in Argentinean Puna: Remarkable UV-B resistance and efficient DNA damage repair. Orig. Life Evol. Biosph. 2012, 42, 201–221. [Google Scholar] [CrossRef]
- Kurth, D.; Belfiore, C.; Gorriti, M.F.; Cortez, N.; Farias, M.E.; Albarracín, V.H. Genomic and proteomic evidences unravel the UV-resistome of the poly-extremophile Acinetobacter sp. Ver3. Front. Microbiol. 2015, 6, 328. [Google Scholar] [CrossRef]
- Nieto, J.; Fernandez-Castillo, R.; Marquez, M.; Ventosa, A.; Quesada, E.; Ruiz-Berraquero, F. Survey of metal tolerance in moderately halophilic eubacteria. Appl. Environ. Microbiol. 1989, 55, 2385–2390. [Google Scholar]
- Nieto, J.; Ventosa, A.; Ruiz-Berraquero, F. Susceptibility of halobacteria to heavy metals. Appl. Environ. Microbiol. 1987, 53, 1199–1202. [Google Scholar]
- Petersen, T.N.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785. [Google Scholar] [CrossRef]
- López-Hermoso, C.; Rafael, R.; Sánchez-Porro, C.; Ventosa, A. Salinivibrio kushneri sp. nov., a moderately halophilic bacterium isolated from salterns. Syst. Appl. Microbiol. 2018, 41, 159–166. [Google Scholar]
- Selvarajan, R.; Sibanda, T.; Tekere, M.; Nyoni, H.; Meddows-Taylor, S. Diversity analysis and bioresource characterization of halophilic bacteria isolated from a South African salt pan. Molecules 2017, 22, 657. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Zhang, W.; Zhang, Q.; Nagata, S. Accumulation and role of compatible solutes in fast-growing Salinivibrio costicola subsp. yaniae. Can. J. Microbiol. 2010, 56, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Gunde-Cimerman, N.; Plemenitaš, A.; Oren, A. Strategies of adaptation of microorganisms of the three domains of life to high salt concentrations. Fems Microbiol. Rev. 2018, 42, 353–375. [Google Scholar] [CrossRef] [PubMed]
- Galinski, E.A. Compatible solutes of halophilic eubacteria: Molecular principles, water-solute interaction, stress protection. Experientia 1993, 49, 487–496. [Google Scholar] [CrossRef]
- Gorriti, M.F.; Dias, G.M.; Chimetto, L.A.; Trindade-Silva, A.E.; Silva, B.S.; Mesquita, M.M.; Gregoracci, G.B.; Farias, M.E.; Thompson, C.C.; Thompson, F.L. Genomic and phenotypic attributes of novel salinivibrios from stromatolites, sediment and water from a high altitude lake. BMC Genom. 2014, 15, 473. [Google Scholar] [CrossRef]
- Udagawa, T.; Unemoto, T.; Tokuda, H. Generation of Na+ electrochemical potential by the Na+-motive NADH oxidase and Na+/H+ antiport system of a moderately halophilic Vibrio costicola. J. Biol. Chem. 1986, 261, 2616–2622. [Google Scholar]
- Hamaide, F.; Kushner, D.J.; Sprott, G.D. Proton motive force and Na+/H+ antiport in a moderate halophile. J. Bacteriol. 1983, 156, 537–544. [Google Scholar]
- Epstein, W. The roles and regulation of potassium in bacteria. Prog. Nucleic Acid Res. Mol. Biol. 2003, 75, 293–320. [Google Scholar]
- Russell, N.; Kogut, M.; Kates, M. Phospholipid Biosynthesis in the Moderately Halophilic Bacterium Vibvio costicola During Adaptation to Changing Salt Concentrations. Microbiology 1985, 131, 781–789. [Google Scholar] [CrossRef][Green Version]
- Ponder, M.A.; Gilmour, S.J.; Bergholz, P.W.; Mindock, C.A.; Hollingsworth, R.; Thomashow, M.F.; Tiedje, J.M. Characterization of potential stress responses in ancient Siberian permafrost psychroactive bacteria. FEMS Microbiol. Ecol. 2005, 53, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Ordoñez, O.F.; Flores, M.R.; Dib, J.R.; Paz, A.; Farías, M.E. Extremophile culture collection from Andean lakes: Extreme pristine environments that host a wide diversity of microorganisms with tolerance to UV radiation. Microb. Ecol. 2009, 58, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Montes, G.; Argüello, J.M.; Valderrama, B. Evolution and diversity of periplasmic proteins involved in copper homeostasis in gamma proteobacteria. BMC Microbiol. 2012, 12, 249. [Google Scholar] [CrossRef] [PubMed]
- Huovinen, P.; Penttilä, H.; Soimasuo, M. Spectral attenuation of solar ultraviolet radiation in humic lakes in Central Finland. Chemosphere 2003, 51, 205–214. [Google Scholar] [CrossRef]
- Stutz, J.; Ackermann, R.; Fast, J.D.; Barrie, L. Atmospheric reactive chlorine and bromine at the Great Salt Lake, Utah. Geophys. Res. Lett. 2002, 29, 18-11–18-14. [Google Scholar] [CrossRef]
- Wilson, C.; Caton, T.; Buchheim, J.; Buchheim, M.; Schneegurt, M.; Miller, R. DNA-repair potential of Halomonas spp. from the Salt Plains Microbial Observatory of Oklahoma. Microb. Ecol. 2004, 48, 541–549. [Google Scholar] [CrossRef]
- Sancar, G.B. Enzymatic photoreactivation: 50 years and counting. Mutat. Res. Fund. Mol. Mech. 2000, 451, 25–37. [Google Scholar] [CrossRef]
- Baliga, N.S.; Bjork, S.J.; Bonneau, R.; Pan, M.; Iloanusi, C.; Kottemann, M.C.; Hood, L.; DiRuggiero, J. Systems level insights into the stress response to UV radiation in the halophilic archaeon Halobacterium NRC-1. Genome Res. 2004, 14, 1025–1035. [Google Scholar] [CrossRef]
- McCready, S.; Marcello, L. Repair of UV damage in Halobacterium salinarum. Biochem. Soc. Trans. 2003, 31 Pt 3, 694–698. [Google Scholar] [CrossRef]
- Capes, M.D.; Coker, J.A.; Gessler, R.; Grinblat-Huse, V.; DasSarma, S.L.; Jacob, C.G.; Kim, J.-M.; DasSarma, P.; DasSarma, S. The information transfer system of halophilic archaea. Plasmid 2011, 65, 77–101. [Google Scholar] [CrossRef]
- Zhao, A.; Gray, F.C.; MacNeill, S.A. ATP-and NAD+-dependent DNA ligases share an essential function in the halophilic archaeon Haloferax volcanii. Mol. Microbiol. 2006, 59, 743–752. [Google Scholar] [CrossRef]
- Krokan, H.E.; Bjørås, M. Base excision repair. Cold Spring Harb. Perspect. Biol. 2013, 5, a012583. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.M. The RecA protein as a recombinational repair system. Mol. Microbiol. 1991, 5, 1295–1299. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhang, X.; Xu, H.; Xu, B.; Hua, Y. RadA: A protein involved in DNA damage repair processes of Deinococcus radiodurans R1. Chin. Sci. Bull. 2006, 51, 2993–2999. [Google Scholar] [CrossRef]
- Pagès, V.; Koffel-Schwartz, N.; Fuchs, R.P. recX, a new SOS gene that is co-transcribed with the recA gene in Escherichia coli. DNA Repair 2003, 2, 273–284. [Google Scholar] [CrossRef]
- Nies, D.H. Microbial heavy-metal resistance. Appl. Microbiol. Biotechnol. 1999, 51, 730–750. [Google Scholar] [CrossRef]
- Kaur, A.; Pan, M.; Meislin, M.; Facciotti, M.T.; El-Gewely, R.; Baliga, N.S. A systems view of haloarchaeal strategies to withstand stress from transition metals. Genome Res. 2006, 16, 841–854. [Google Scholar] [CrossRef]
- Das, D.; Salgaonkar, B.B.; Mani, K.; Braganca, J.M. Cadmium resistance in extremely halophilic archaeon Haloferax strain BBK2. Chemosphere 2014, 112, 385–392. [Google Scholar] [CrossRef]
- Ladomersky, E.; Petris, M.J. Copper tolerance and virulence in bacteria. Metallomics 2015, 7, 957–964. [Google Scholar] [CrossRef]
- Munson, G.P.; Lam, D.L.; Outten, F.W.; O’Halloran, T.V. Identification of a copper-responsive two-component system on the chromosome of Escherichia coli K-12. J. Bacteriol. 2000, 182, 5864–5871. [Google Scholar] [CrossRef]
- Rosen, B.P. Transport and detoxification systems for transition metals, heavy metals and metalloids in eukaryotic and prokaryotic microbes. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 133, 689–693. [Google Scholar] [CrossRef]
- Lin, Y.-F.; Walmsley, A.R.; Rosen, B.P. An arsenic metallochaperone for an arsenic detoxification pump. Proc. Natl. Acad. Sci. USA 2006, 103, 15617–15622. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, H.; Shao, Z.; Liao, S.; Johnstone, L.; Rensing, C.; Wang, G. Genome sequence of deep-sea manganese-oxidizing bacterium Marinobacter manganoxydans MnI7-9. Am. Soc. Microbiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Hynninen, A. Zinc, Cadmium and Lead Resistance Mechanisms in Bacteria and Their Contribution to Biosensing. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2010. [Google Scholar]
- Nies, D.H. Efflux-mediated heavy metal resistance in prokaryotes. FEMS Microbiol. Rev. 2003, 27, 313–339. [Google Scholar] [CrossRef]
- Borremans, B.; Hobman, J.; Provoost, A.; Brown, N.; van Der Lelie, D. Cloning and functional analysis of thepbr lead resistance determinant of Ralstonia metallidurans CH34. J. Bacteriol. 2001, 183, 5651–5658. [Google Scholar] [CrossRef]
- Xu, L.; Xu, X.-W.; Meng, F.-X.; Huo, Y.-Y.; Oren, A.; Yang, J.-Y.; Wang, C.-S. Halomonas zincidurans sp. nov., a heavy-metal-tolerant bacterium isolated from the deep-sea environment. Int. J. Syst. Evol. Microbiol. 2013, 63, 4230–4236. [Google Scholar] [CrossRef]
- Freedman, Z.; Zhu, C.; Barkay, T. Mercury resistance and mercuric reductase activities and expression among chemotrophic thermophilic Aquificae. Appl. Environ. Microbiol. 2012, 78, 6568–6575. [Google Scholar] [CrossRef]
- Silver, S.; Phung, L.T. Bacterial Mercury resistance proteins. Encycl. Met. 2013, 27, 209–217. [Google Scholar]
- Zhou, P.; Huo, Y.-Y.; Xu, L.; Wu, Y.-H.; Meng, F.-X.; Wang, C.-S.; Xu, X.-W. Investigation of mercury tolerance in Chromohalobacter israelensis DSM 6768T and Halomonas zincidurans B6T by comparative genomics with Halomonas xinjiangensis TRM 0175T. Mar. Genom. 2015, 19, 15–16. [Google Scholar] [CrossRef]
- Boyd, E.; Barkay, T. The mercury resistance operon: From an origin in a geothermal environment to an efficient detoxification machine. Front. Microbiol. 2012, 3, 349. [Google Scholar] [CrossRef]
- Viti, C.; Marchi, E.; Decorosi, F.; Giovannetti, L. Molecular mechanisms of Cr (VI) resistance in bacteria and fungi. FEMS Microbiol. Rev. 2014, 38, 633–659. [Google Scholar] [CrossRef] [PubMed]
- Voica, D.M.; Bartha, L.; Banciu, H.L.; Oren, A. Heavy metal resistance in halophilic Bacteria and Archaea. FEMS Microbiol. Lett. 2016, 363. [Google Scholar]
- Branco, R.; Chung, A.P.; Johnston, T.; Gurel, V.; Morais, P.; Zhitkovich, A. The chromate-inducible chrBACF operon from the transposable element TnOtChr confers resistance to chromium (VI) and superoxide. J. Bacteriol. 2008, 190, 6996–7003. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.L.; Gallegos, M.T.; Marqués, S.; Ramos-González, M.I.; Espinosa-Urgel, M.; Segura, A. Responses of Gram-negative bacteria to certain environmental stressors. Curr. Opin. Microbiol. 2001, 4, 166–171. [Google Scholar] [CrossRef]
- Raivio, T.L.; Popkin, D.L.; Silhavy, T.J. The Cpx envelope stress response is controlled by amplification and feedback inhibition. J. Bacteriol. 1999, 181, 5263–5272. [Google Scholar]
- Alba, B.M.; Gross, C.A. Regulation of the Escherichia coli σ E-dependent envelope stress response. Mol. Microbiol. 2004, 52, 613–619. [Google Scholar] [CrossRef]
- Cezairliyan, B.O.; Sauer, R.T. Inhibition of regulated proteolysis by RseB. Proc. Natl. Acad. Sci. USA 2007, 104, 3771–3776. [Google Scholar] [CrossRef]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef]
- Chiang, S.M.; Schellhorn, H.E. Regulators of oxidative stress response genes in Escherichia coli and their functional conservation in bacteria. Arch. Biochem. Biophys. 2012, 525, 161–169. [Google Scholar] [CrossRef]
- Shiu, C.-T.; Lee, T.-M. Ultraviolet-B-induced oxidative stress and responses of the ascorbate–glutathione cycle in a marine macroalga Ulva Fasciata. J. Exp. Bot. 2005, 56, 2851–2865. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Verlecar, X.; Jena, K.; Chainy, G. Biochemical markers of oxidative stress in Perna viridis exposed to mercury and temperature. Chem. Biol. Interact. 2007, 167, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Maleki, F.; Afra Khosravi, A.N.; Taghinejad, H.; Azizian, M. Bacterial heat shock protein activity. Clin. Diagn Res. JCDR 2016, 10, BE01. [Google Scholar] [CrossRef] [PubMed]
- Moat, A.G.; Foster, J.W.; Spector, M.P. Microbial Physiology; John Wiley & Sons: Hoboken, NJ, USA, 2003. [Google Scholar]
- Masip, L.; Veeravalli, K.; Georgiou, G. The many faces of glutathione in bacteria. Antioxid. Redox Signal. 2006, 8, 753–762. [Google Scholar] [CrossRef]
- Zhang, H.; Li, X.; Xie, Y.; Jin, J.; Liu, H.; Gao, X.; Xiong, L. Carbon starvation-induced lipoprotein Slp directs the synthesis of catalase and expression of OxyR regulator to protect against hydrogen peroxide stress in Escherichia coli. BioRxiv 2018, 386003. [Google Scholar] [CrossRef]
- Koga, T.; Takumi, K. Nutrient starvation induces cross protection against heat, osmotic, or H2O2 challenge in Vibrio parahaemolyticus. Microbiol. Immunol. 1995, 39, 213–215. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GGDH | OrthoANI | |
|---|---|---|
| S. kushneri IB560 | 79.90% | 97.79% |
| S. kushneri IB563 | 78.10% | 97.61% |
| S. kushneri ML338A | 80.50% | 97.79% |
| S. kushneri IC317 | 80.30% | 97.84% |
| S. kushneri MA421 | 78.40% | 97.62% |
| S. kushneri ML318 | 80.40% | 97.94% |
| S. kushneri AL184 | 81.10% | 97.79% |
| S. kushneri ML331 | 80.10% | 97.76% |
| S. kushneri IC202 | 78% | 97.64% |
| S. kushneri ML277 | 79.90% | 97.75% |
| Salinity (ppt) | Growth | Time Taken for Growth in Days at ~30 °C |
|---|---|---|
| 0 | Absent | |
| 15 | Present | Overnight incubation |
| 30 | Present | Overnight incubation |
| 60 | Present | Overnight incubation |
| 120 | Present | 2 days |
| 150 | Present | 2 days |
| 200 | Present | 3 days |
| Salinity | 30 ppt | 60 ppt | 120 ppt | 150 ppt | 200 ppt | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Concentrations | 1 mM | 5 mM | 10 mM | 1 mM | 5 mM | 10 mM | 1 mM | 5 mM | 10 mM | 1 mM | 5 mM | 10 mM | 1 mM | 5 mM | 10 mM |
| Arsenic | +++ | +++ | - | +++ | +++ | - | +++ | +++ | - | +++ | +++ | + | +++ | +++ | + |
| Copper | +++ | + | - | +++ | + | - | +++ | ++ | - | +++ | +++ | + | +++ | +++ | + |
| Cobalt | +++ | +++ | + | +++ | +++ | ++ | +++ | +++ | ++ | +++ | +++ | ++ | +++ | +++ | ++ |
| Zinc | +++ | +++ | ++ | +++ | +++ | ++ | +++ | + | - | +++ | - | - | +++ | - | - |
| Chromium | +++ | - | - | +++ | - | - | +++ | - | - | +++ | - | - | ++ | - | - |
| Lead | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
John, J.; Siva, V.; Richa, K.; Arya, A.; Kumar, A. Life in High Salt Concentrations with Changing Environmental Conditions: Insights from Genomic and Phenotypic Analysis of Salinivibrio sp. Microorganisms 2019, 7, 577. https://doi.org/10.3390/microorganisms7110577
John J, Siva V, Richa K, Arya A, Kumar A. Life in High Salt Concentrations with Changing Environmental Conditions: Insights from Genomic and Phenotypic Analysis of Salinivibrio sp. Microorganisms. 2019; 7(11):577. https://doi.org/10.3390/microorganisms7110577
Chicago/Turabian StyleJohn, Jojy, Vinu Siva, Kumari Richa, Aditya Arya, and Amit Kumar. 2019. "Life in High Salt Concentrations with Changing Environmental Conditions: Insights from Genomic and Phenotypic Analysis of Salinivibrio sp." Microorganisms 7, no. 11: 577. https://doi.org/10.3390/microorganisms7110577
APA StyleJohn, J., Siva, V., Richa, K., Arya, A., & Kumar, A. (2019). Life in High Salt Concentrations with Changing Environmental Conditions: Insights from Genomic and Phenotypic Analysis of Salinivibrio sp. Microorganisms, 7(11), 577. https://doi.org/10.3390/microorganisms7110577