Effect of Temperatures Used in Food Storage on Duration of Heat Stress Induced Invasiveness of L. monocytogenes

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. L. monocytogenes Strains
2.2. Growth of L. monocytogenes Strains
2.3. Cell Line and Culture Conditions
2.4. Plaque Forming Assay
2.5. Statistical Analysis
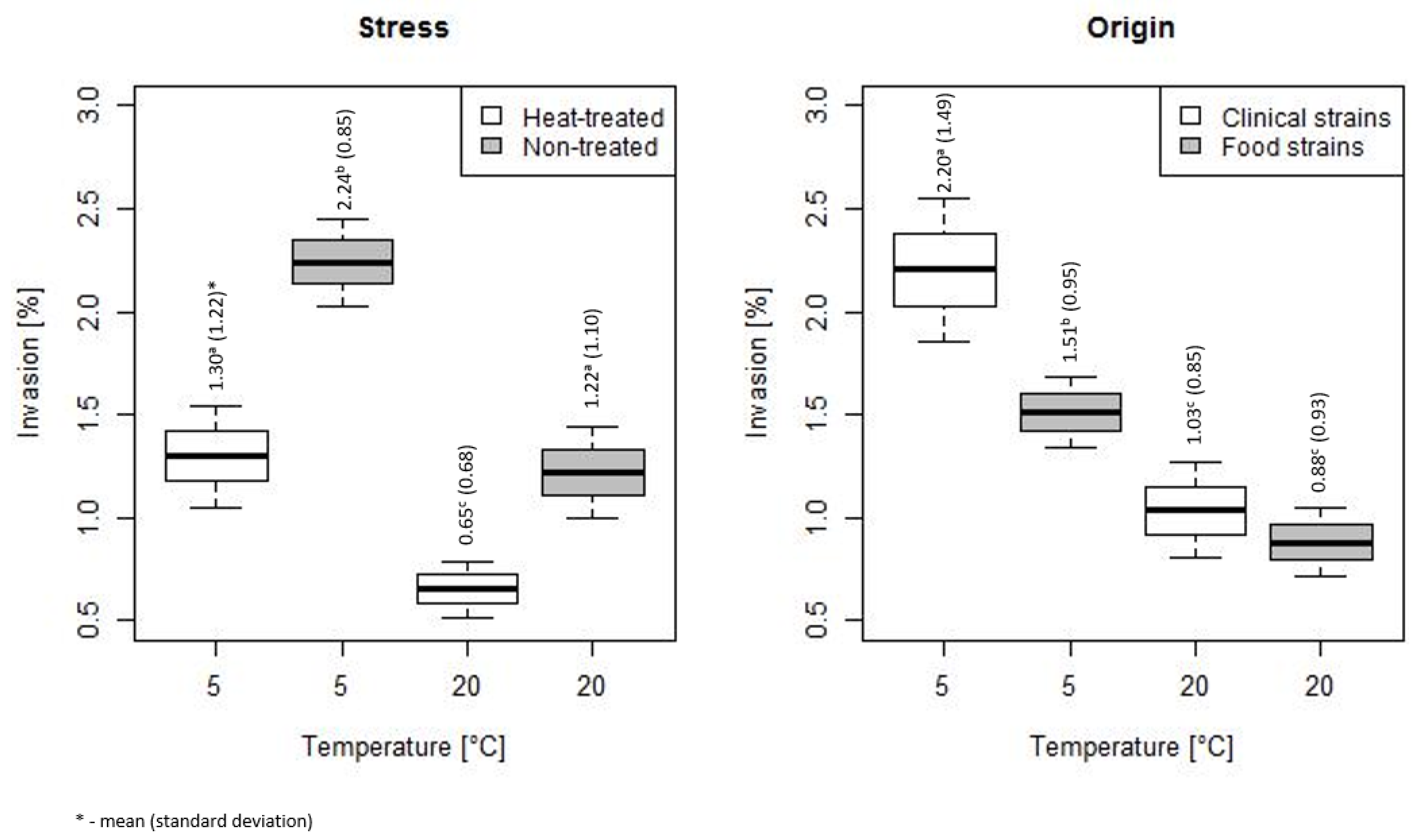
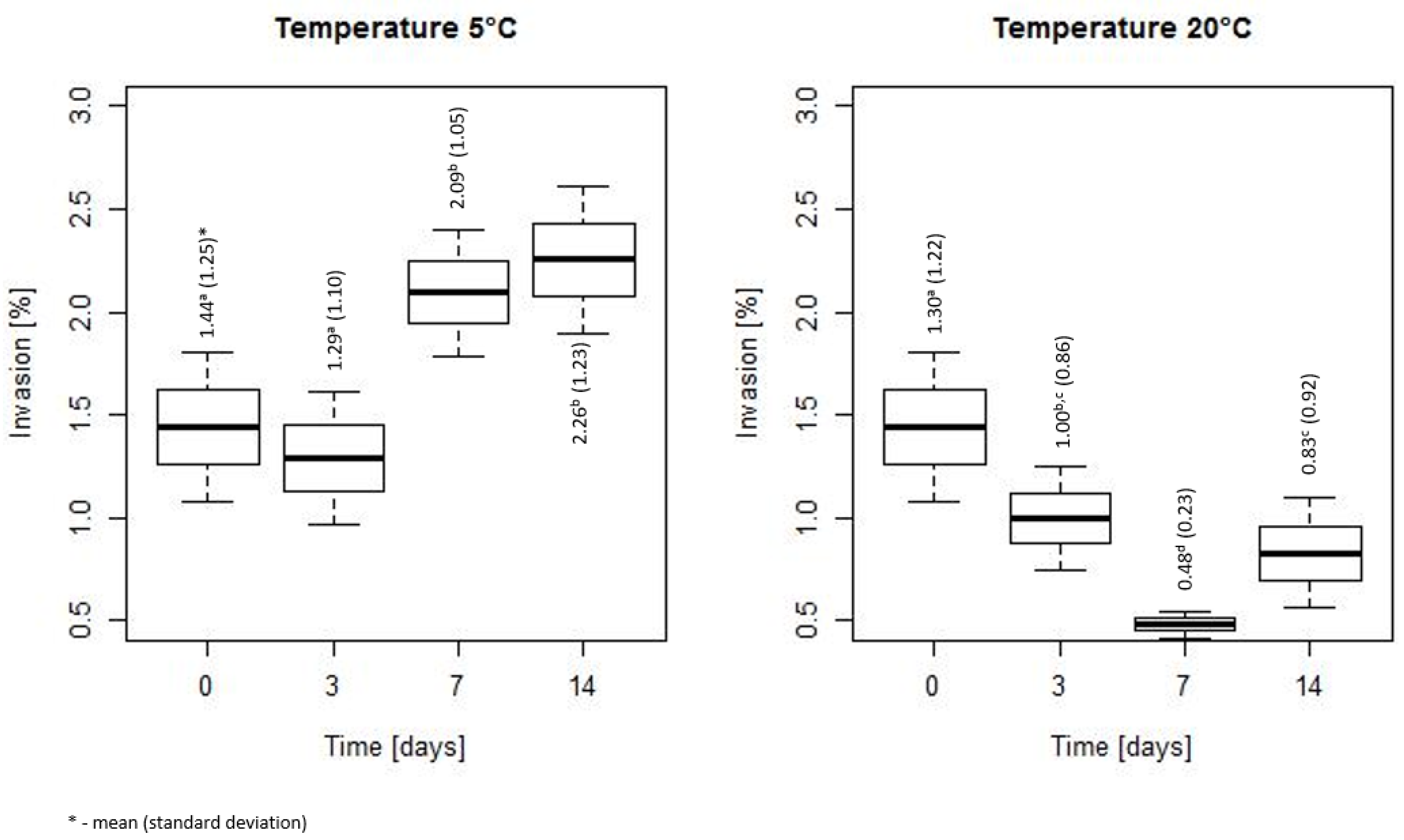
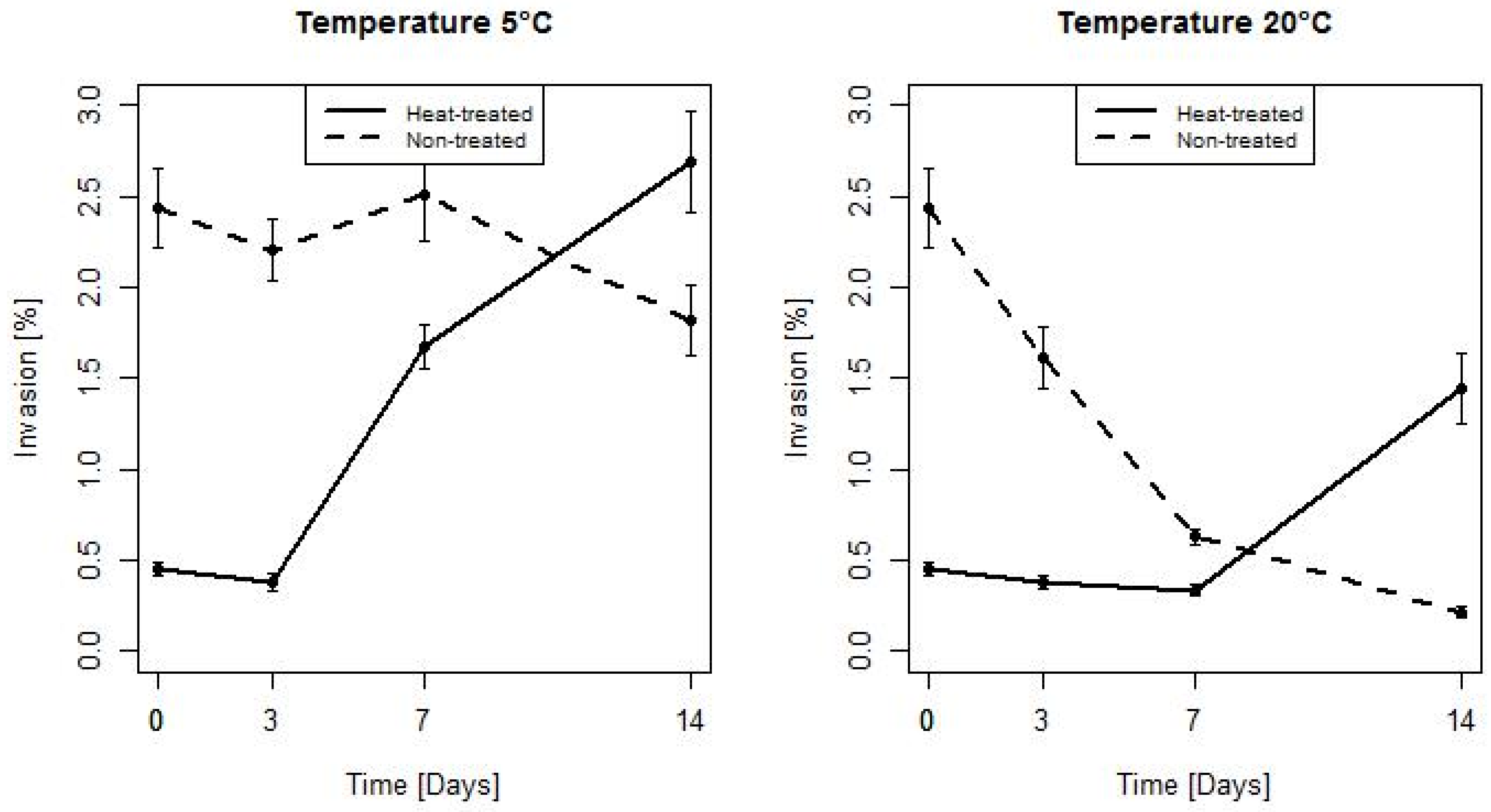
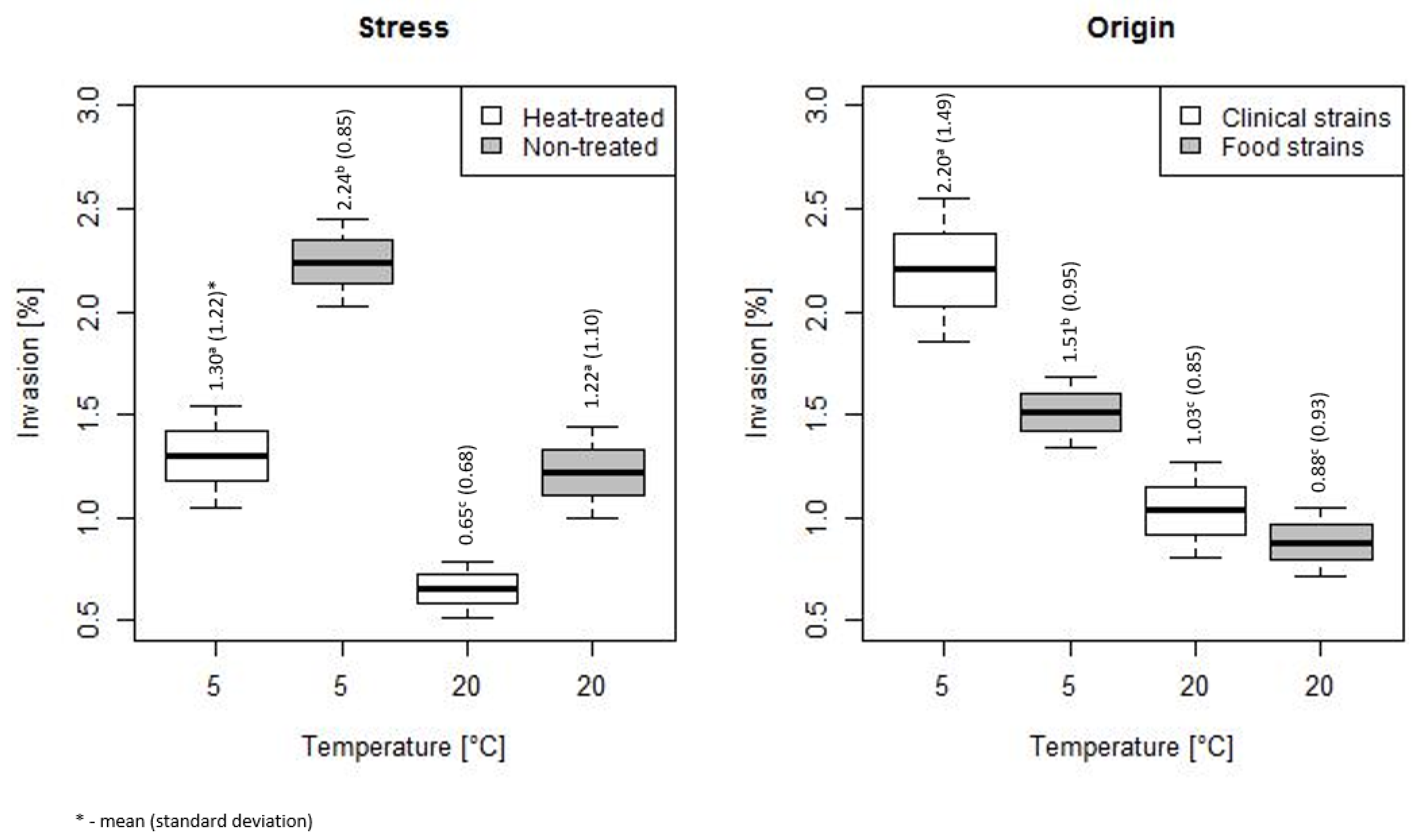
3. Results
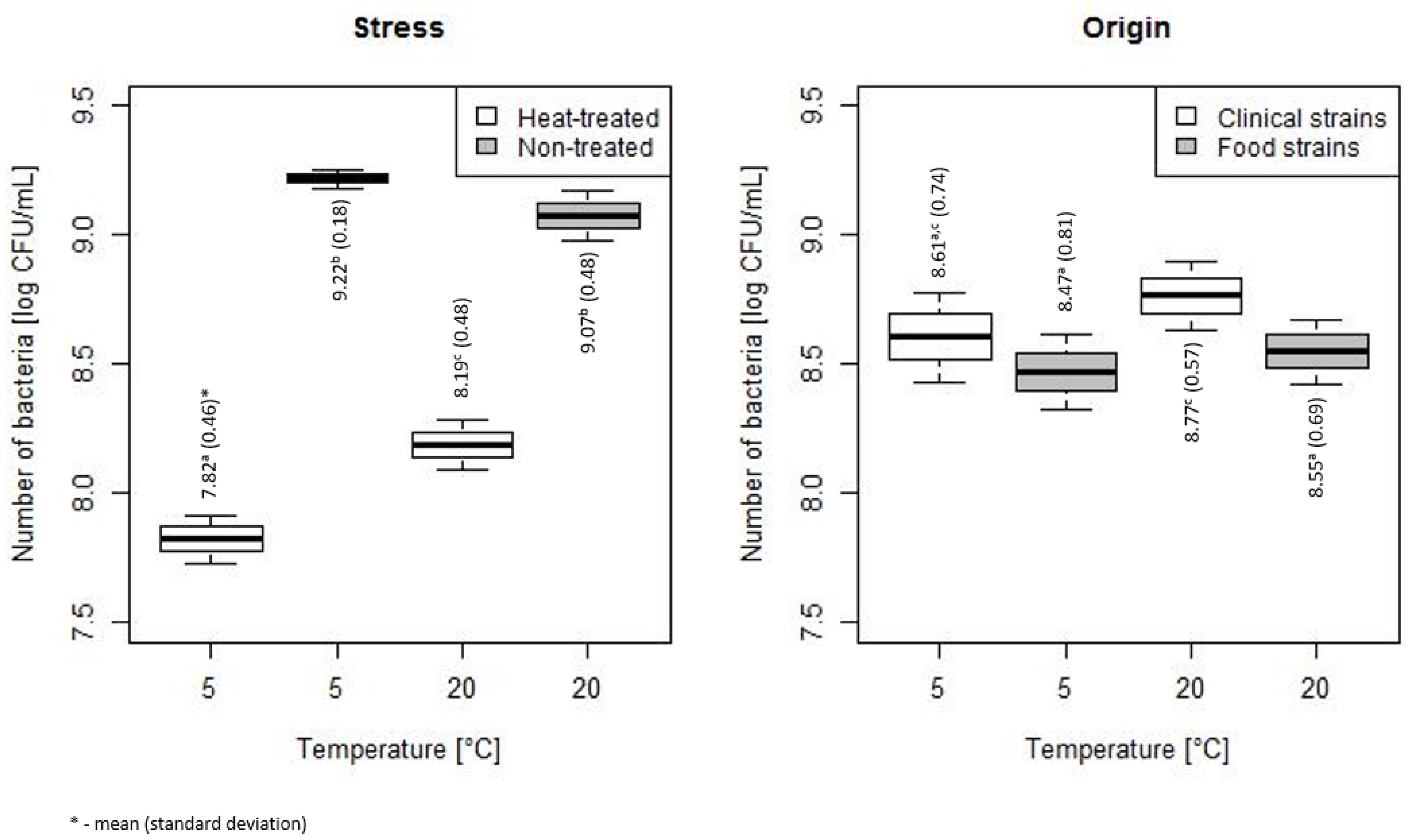
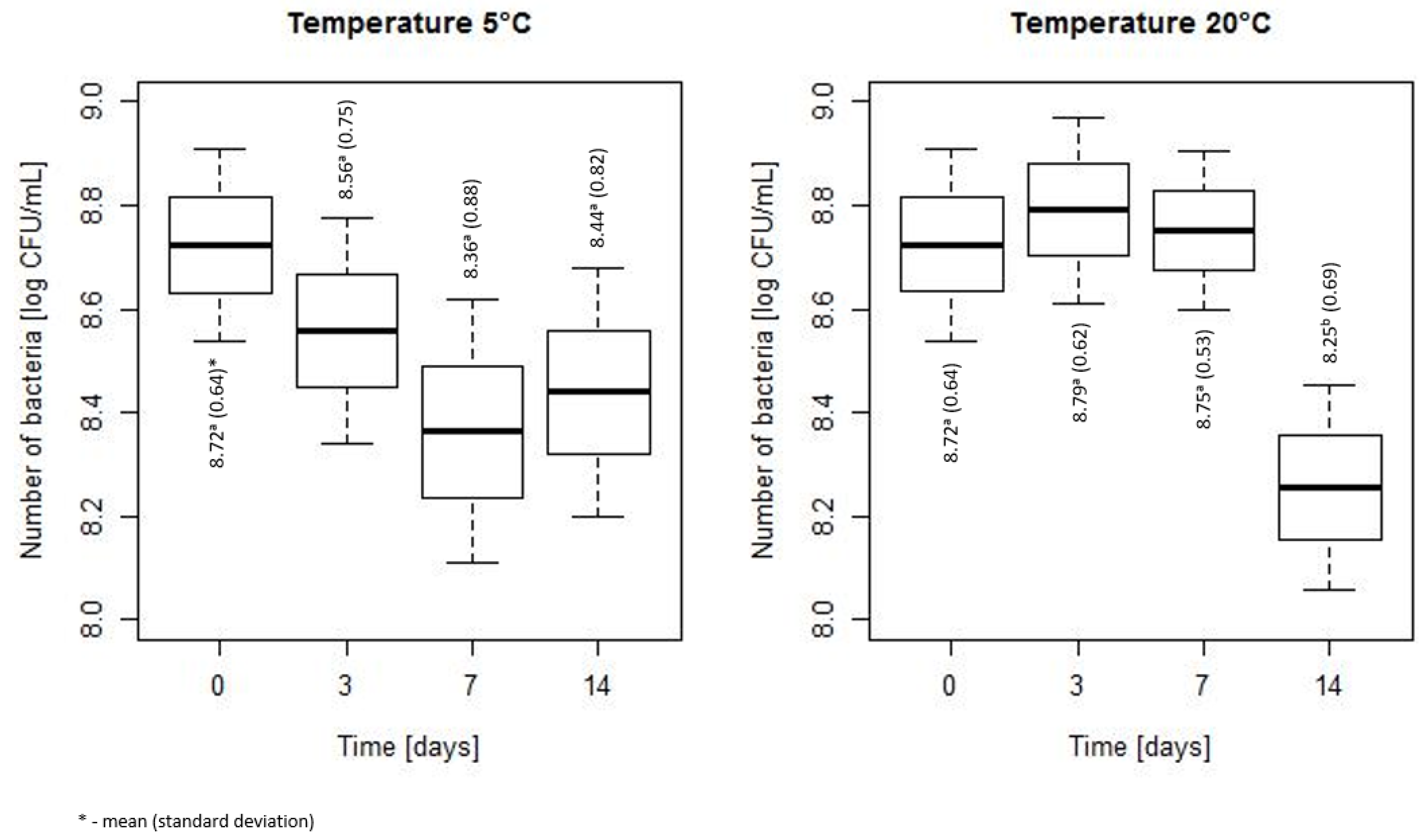
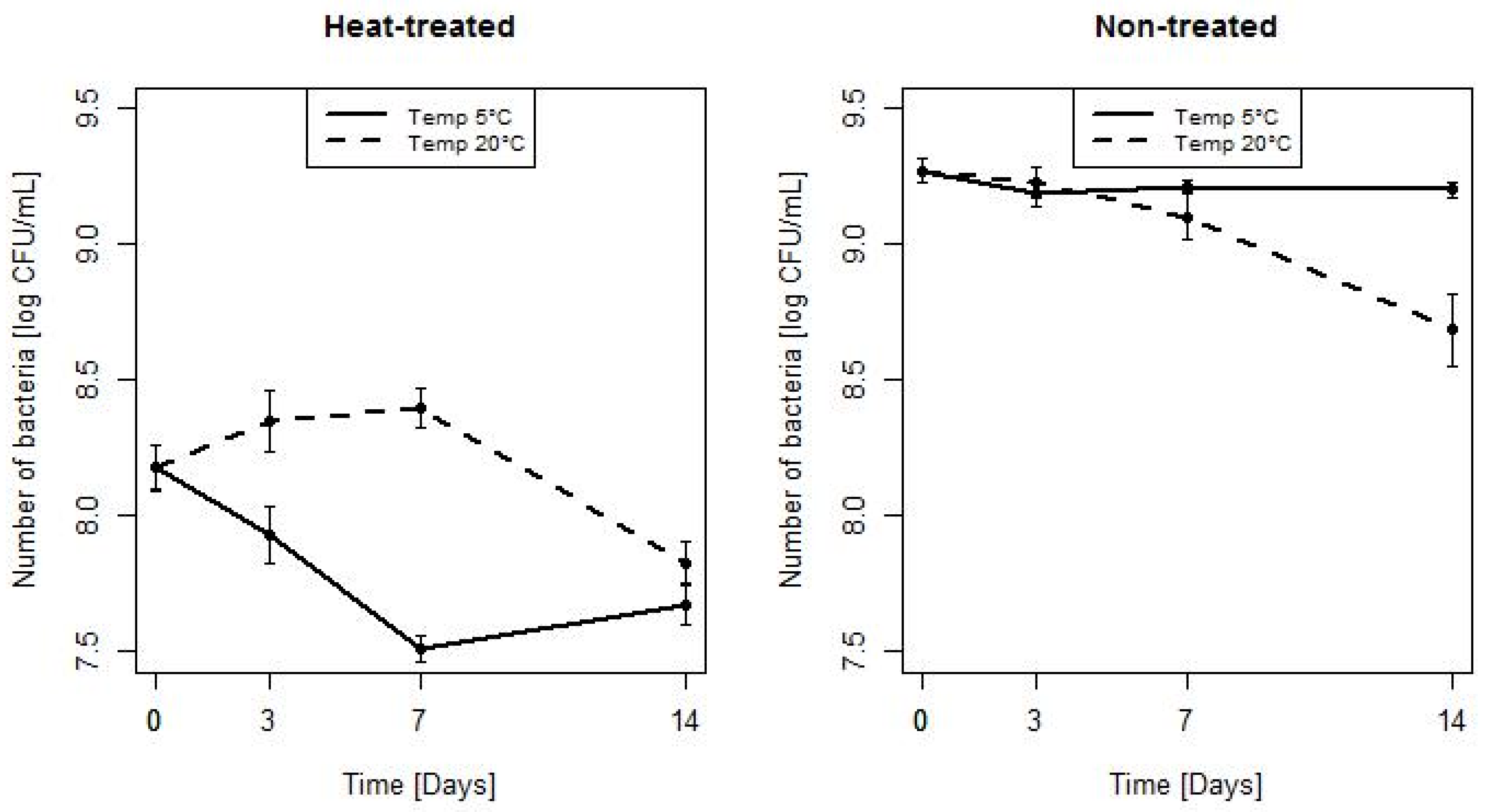
3.1. Assessment of Bacterial Count During Storage at 5 and 20 °C
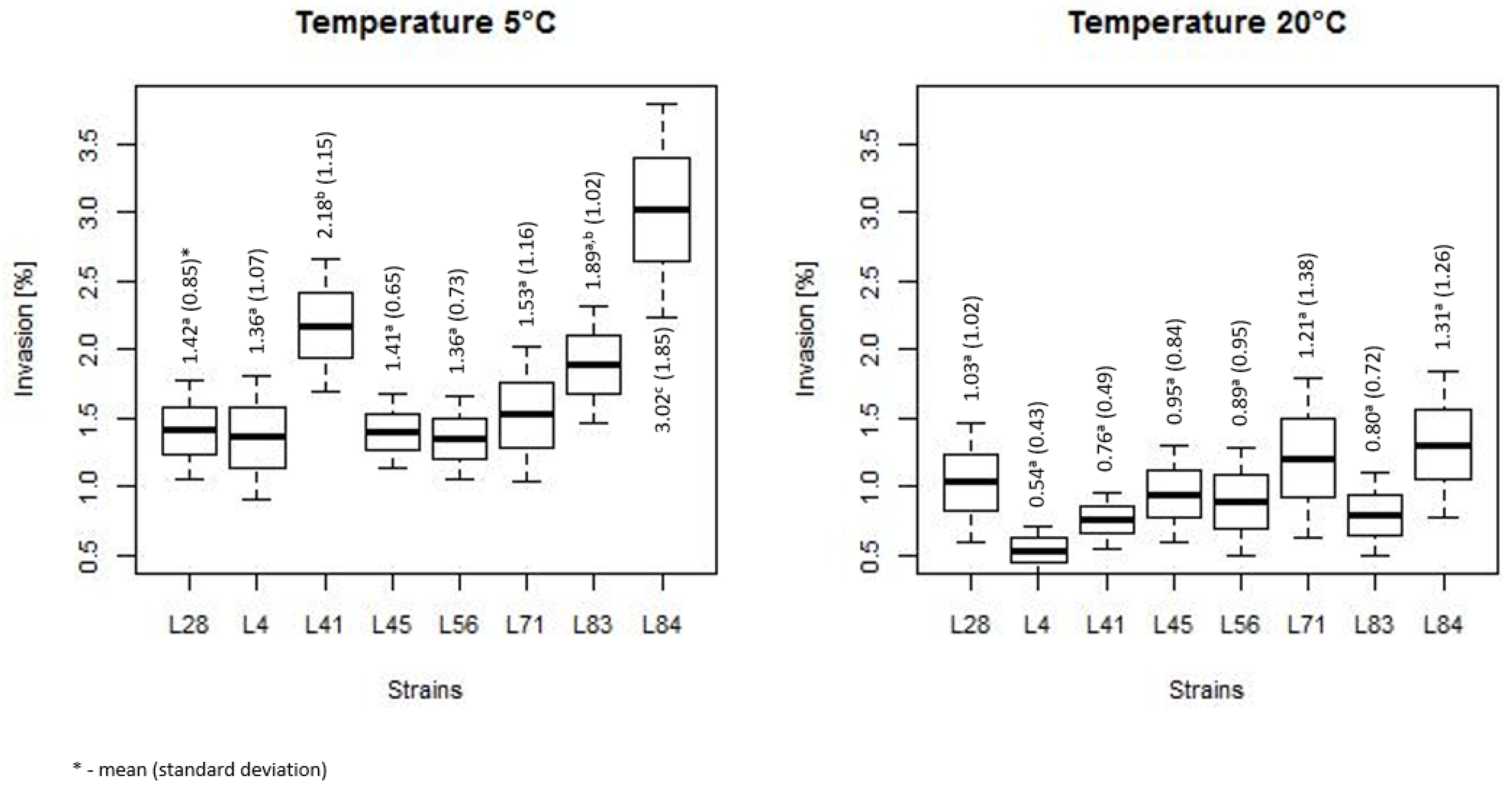
3.2. Duration of Heat Stress Effect on Invasiveness of L. monocytogenes
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Alonso-Hernando, A.; Prieto, M.; Garcı’a-Ferna´ndez, C.; Alonso-Calleja, C.; Capita, R. Increase over time in the prevalence of multiple antibiotic resistance among isolates of Listeria monocytogenes from poultry in Spain. Food Control 2012, 23, 37–41. [Google Scholar] [CrossRef]
- Melo, J.; Andrew, P.W.; Faleiro, M.L. Listeria monocytogenes in cheese and the dairy environment remains a food safety challenge: The role of stress responses. Food Res. Int. 2015, 64, 75–90. [Google Scholar] [CrossRef]
- Arslan, F.; Meynet, E.; Sunbul, M.; Sipahi, O.R.; Kurtaran, B.; Kaya, S.; Inkaya, A.C.; Pagliano, P.; Sengoz, G.; Batirel, A.; et al. The clinical features, diagnosis, treatment, and prognosis of neuroinvasive listeriosis: A multinational study. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- EFSA (European Food Safety Authority), ECDC (European Centre for Disease Prevention and Control): The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, 5500.
- Kazimierczak, M.J.; Mithoe, S.C.; Boor, K.J.; Wiedmann, M. Listeria monocytogenes sigma B regulates stress response and virulence functions. J. Bacteriol. 2003, 185, 5722–5734. [Google Scholar] [CrossRef]
- Dorey, A.; Marinho, C.; Piveteau, P.; O’Byrne, C. Role and regulation of the stress activated sigma factor sigma B (σB) in the saprophytic and host-associated life stages of Listeria monocytogenes. Adv. Appl. Microbiol. 2019, 106, 1–48. [Google Scholar]
- Roche, S.M.; Gracieux, P.; Milohanic, E.; Albert, I.; Virlogeux-Payant, S.; Témoin, O.; Grépinet, A.; Kerouanton, C.; Jacquet, P.; Cossart, P.; et al. Investigation of specific substitutions in virulence genes characterizing phenotypic groups of low-virulence field strains of Listeria monocytogenes. Appl. Environ. Microbiol. 2005, 71, 6039–6048. [Google Scholar] [CrossRef]
- Zhou, X.; Jiao, X.; Wiedmann, M. Listeria monocytogenes in the Chinese food system: Strain characterization through partial actA sequencing and tissue-culture pathogenicity assays. J. Med. Microbiol. 2005, 54, 217–224. [Google Scholar] [CrossRef]
- Neuhaus, K.; Satorhelyi, P.; Schauer, K.; Scherer, S.; Fuchs, T.M. Acid shock of Listeria monocytogenes at low environmental temperatures induces prfA, epithelial cell invasion, and lethality towards Caenorhabditis elegans. BMC Genom. 2013, 14, 285. [Google Scholar] [CrossRef]
- Pricope, L.; Nicolau, A.; Wagner, M.; Rychli, K. The effect of sublethal concentrations of benzalkonium chloride on invasiveness and intracellular proliferation of Listeria monocytogenes. Food Control. 2013, 3, 1230–1235. [Google Scholar] [CrossRef]
- Wałecka-Zacharska, E.; Kosek-Paszkowska, K.; Bania, J.; Karpíšková, R. Salt stress-induced invasiveness of major Listeria monocytogenes serotypes. Lett. Appl. Microbiol. 2013, 56, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Larsen, N.; Jespersen, L. Expression of virulence-related genes in Listeria monocytogenes grown on Danish hard cheese as affected by NaCl content. Food. Pathog. Dis. 2015, 12, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Duodu, S.; Holst-Jensen, A.; Skjerdal, T.; Cappelier, J.M.; Pilet, M.F.; Loncarevic, S. Influence of storage temperature on gene expression and virulence potential of Listeria monocytogenes strains grown in a salmon matrix. Food Microbiol. 2010, 27, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Pricope-Ciolacu, L.; Nicolau, A.; Wagner, M.; Rychli, K. The effect of milk components and storage conditions on the virulence of Listeria monocytogenes as determined by a Caco-2 cell assay. Int. J. Food Microbiol. 2013, 166, 59–64. [Google Scholar] [CrossRef]
- Colás-Medà, P.; Viñasa, I.; Oliveira, M.; Anguera, M.; Serrano, J.C.E.; Abadias, M. Exposure to minimally processed pear and melon during shelf life could modify the pathogenic potential of Listeria monocytogenes. Food Microbiol. 2017, 62, 275–281. [Google Scholar] [CrossRef]
- Wałecka, E.; Molenda, J.; Karpišková, R.; Bania, J. Effect of heat exposure on invasiness of Listeria monocytogenes strains. Food. Pathog. Dis. 2011, 8, 839–841. [Google Scholar] [CrossRef]
- Wałecka Zacharska, E.; Gmyrek, R.; Skowron, K.; Kosek Paszkowska, K.; Bania, J. Duration of heat stress effect on invasiveness of L. monocytogenes strains. BioMed Res. Int. 2018, 2018, 1457480. [Google Scholar]
- Doumith, M.; Buchrieser, C.; Glaser, P.; Jacquet, C.; Martin, P. Differentiation of the major Listeria monocytogenes serovars by multiplex PCR. J. Clin. Microbiol. 2004, 42, 3819–3822. [Google Scholar] [CrossRef]
- Jarvis, N.A.; O’Bryan, C.A.; Dawoud, T.M.; Park, S.H.; Kwon, Y.M.; Crandall, P.G.; Ricke, S.C. An overview of Salmonella thermal destruction during food processing and preparation. Food Control 2016, 68, 280–290. [Google Scholar] [CrossRef]
- Pereira, A.; Alves, A.; Ferreira, V.; Teixeira, P.C.M. The Impact of environmental stresses in the virulence traits of Listeria monocytogenes relevant to Food Safety. IntechOpen 2018, 7, 90–108. [Google Scholar]
- Lado, B.H.; Yousef, A.E. Characteristics of Listeria monocytogenes important to food processors, in Listeria, Listeriosis, and Food Safety; Ryser, E.T., Marth, E.H., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 157–213. [Google Scholar]
- Skandamis, P.N.; Yoon, Y.; Stopforth, J.D.; Kendall, P.A.; Sofos, J.N. Heat and acid tolerance of Listeria monocytogenes after exposure to single and multiple sublethal stress. Food Microbiol. 2008, 25, 294–303. [Google Scholar] [CrossRef] [PubMed]
- McWilliams, G.; Furey, S.; Williamson, T.; O’Brolchain, M.; Anderson, W.; Quinn, G. Issuing temperature guidance to consumers on the cooking and storage of food. Safefood 2004, 1–22. [Google Scholar]
- Mortimore, S.; Wallace, C. HACCP: A Practical Approach; Sara, M., Carol, W., Eds.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; pp. 155–173. ISBN 978-1-4614-5027-6. [Google Scholar]
- Liu, D. Identification, subtyping and virulence determination of Listeria monocytogenes, an important foodborne pathogen. J. Med. Microbiol. 2006, 55, 645–659. [Google Scholar] [CrossRef] [PubMed]
- Rakic Martinez, M.; Wiedmann, M.; Ferguson, M.; Datta, A.R. Assessment of Listeria monocytogenes virulence in the Galleria mellonella insect larvae model. PLoS ONE 2017, 12, e0184557. [Google Scholar] [CrossRef] [PubMed]
- Law, R.J.; Gur-Arie, L.; Rosenshine, I.; Finlay, B.B. In vitro and in vivo model systems for studying enteropathogenic Escherichia coli infections. Cold Spring Harb. Perspect. Med. 2013, 3, a009977. [Google Scholar] [CrossRef] [PubMed]
- Roche, S.M.; Velge, P.; Bottreau, E.; Durier, C.; Marquet-van der Mee, N.; Pardon, P. Assesment of the virulence of Listeria monocytogenes: Agreement between a plaque-forming assay with HT-29 cells and infection of immonocompetent mice. Int. J. Food Microbiol. 2001, 68, 33–44. [Google Scholar] [CrossRef]
- Velge, P.; Bottreau, E.; Van-Langendonck, N.; Kaeffer, B. Cell proliferation enhances entry of Listeria monocytogenes into intestinal epithelial cells by two proliferation-dependent entry pathways. J. Med. Microbiol. 1997, 46, 681–692. [Google Scholar] [CrossRef]
- Stollewerk, K.; Cruz, C.D.; Fletcher, G.; Garriga, M.; Jofr´e, A. The effect of mild preservation treatments on the invasiveness of different Listeria monocytogenes strains on Greenshell mussels. Food Control 2017, 71, 322–328. [Google Scholar] [CrossRef]
- Ress, C.E.; Dodd, C.E.; Gibson, P.T.; Booth, I.R.; Stweart, G.S. Significance of bacteria in stationary phase to food microbiology. Int. J. Food Microbiol. 1955, 28, 263–275. [Google Scholar] [CrossRef]
- Kazmierczak, M.J.; Wiedmann, M.; Boor, K.J. Alternative sigma factors and their roles in bacterial virulence. J. Bacteriol. 2005, 185, 5722–5734. [Google Scholar] [CrossRef]
- Olesen, I.; Vogensen, F.K.; Jespersen, L. Gene transcription and virulence potential of Listeria monocytogenes strains after exposure to acidic and NaCl stress. Food. Pathog. Dis. 2009, 6, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Wałecka, E.; Molenda, J.; Karpišková, R.; Bania, J. Effect of osmotic stress and culture density on invasiveness of Listeria monocytogenes strains. Int. J. Foood Microbiol. 2011, 144, 440–445. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Source | Lineage | Serotype |
|---|---|---|---|
| L28 | Human | I | 4b |
| L84 | Human | I | 4b |
| L41 | Human | I | 1/2b |
| L45 | Food | I | 1/2b |
| L56 | Food | II | 1/2a |
| L71 | Food | II | 1/2a |
| L83 | Food | II | 1/2a |
| L4 | Food | III | 4c |
| Parameters | Value | Std. Error | t-Value | p-Value |
|---|---|---|---|---|
| (Intercept) | 9.22 | 0.07 | 133.98 | <0.001 |
| Stress HT vs. NT | −0.88 | 0.05 | −16.75 | <0.001 |
| Temperature 5 °C vs. 20 °C | 0.15 | 0.05 | 2.78 | 0.006 |
| Storage (Days) 3 vs. 0 | −0.05 | 0.05 | −0.93 | 0.353 |
| Storage (Days) 7 vs. 0 | −0.17 | 0.06 | −2.83 | 0.005 |
| Storage (Days) 14 vs. 0 | −0.38 | 0.07 | −5.1 | <0.001 |
| Stress HT: Temperature 5 °C | −0.51 | 0.07 | −6.88 | <0.001 |
| Parameters | Value | Std. Error | t-Value | p-Value |
|---|---|---|---|---|
| (Intercept) | 1.93 | 0.19 | 10.2 | <0.001 |
| Stress HT vs. NT | −1.8 | 0.16 | −11.06 | <0.001 |
| Temperature 5 vs. 20 | 1.02 | 0.1 | 9.9 | <0.001 |
| Storage (Days) 3 vs. 0 | −0.53 | 0.16 | −3.33 | 0.001 |
| Storage (Days) 7 vs. 0 | −0.87 | 0.2 | −4.24 | <0.001 |
| Storage (Days) 14 vs. 0 | −1.42 | 0.32 | −4.39 | <0.001 |
| Stress HT: Temperature 5 | −0.37 | 0.15 | −2.55 | 0.011 |
| Stress HT: Storage (Days) 3 | 0.46 | 0.21 | 2.22 | 0.027 |
| Stress HT: Storage (Days) 7 | 1.43 | 0.21 | 6.92 | <0.001 |
| Stress HT: Storage (Days) 14 | 3.04 | 0.21 | 14.74 | <0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wałecka-Zacharska, E.; Korkus, J.; Skowron, K.; Wietlicka-Piszcz, M.; Kosek-Paszkowska, K.; Bania, J. Effect of Temperatures Used in Food Storage on Duration of Heat Stress Induced Invasiveness of L. monocytogenes. Microorganisms 2019, 7, 467. https://doi.org/10.3390/microorganisms7100467
Wałecka-Zacharska E, Korkus J, Skowron K, Wietlicka-Piszcz M, Kosek-Paszkowska K, Bania J. Effect of Temperatures Used in Food Storage on Duration of Heat Stress Induced Invasiveness of L. monocytogenes. Microorganisms. 2019; 7(10):467. https://doi.org/10.3390/microorganisms7100467
Chicago/Turabian StyleWałecka-Zacharska, Ewa, Jakub Korkus, Krzysztof Skowron, Magdalena Wietlicka-Piszcz, Katarzyna Kosek-Paszkowska, and Jacek Bania. 2019. "Effect of Temperatures Used in Food Storage on Duration of Heat Stress Induced Invasiveness of L. monocytogenes" Microorganisms 7, no. 10: 467. https://doi.org/10.3390/microorganisms7100467
APA StyleWałecka-Zacharska, E., Korkus, J., Skowron, K., Wietlicka-Piszcz, M., Kosek-Paszkowska, K., & Bania, J. (2019). Effect of Temperatures Used in Food Storage on Duration of Heat Stress Induced Invasiveness of L. monocytogenes. Microorganisms, 7(10), 467. https://doi.org/10.3390/microorganisms7100467