Differential Distribution and Determinants of Ammonia Oxidizing Archaea Sublineages in the Oxygen Minimum Zone off Costa Rica



Abstract
1. Introduction
2. Materials and methods
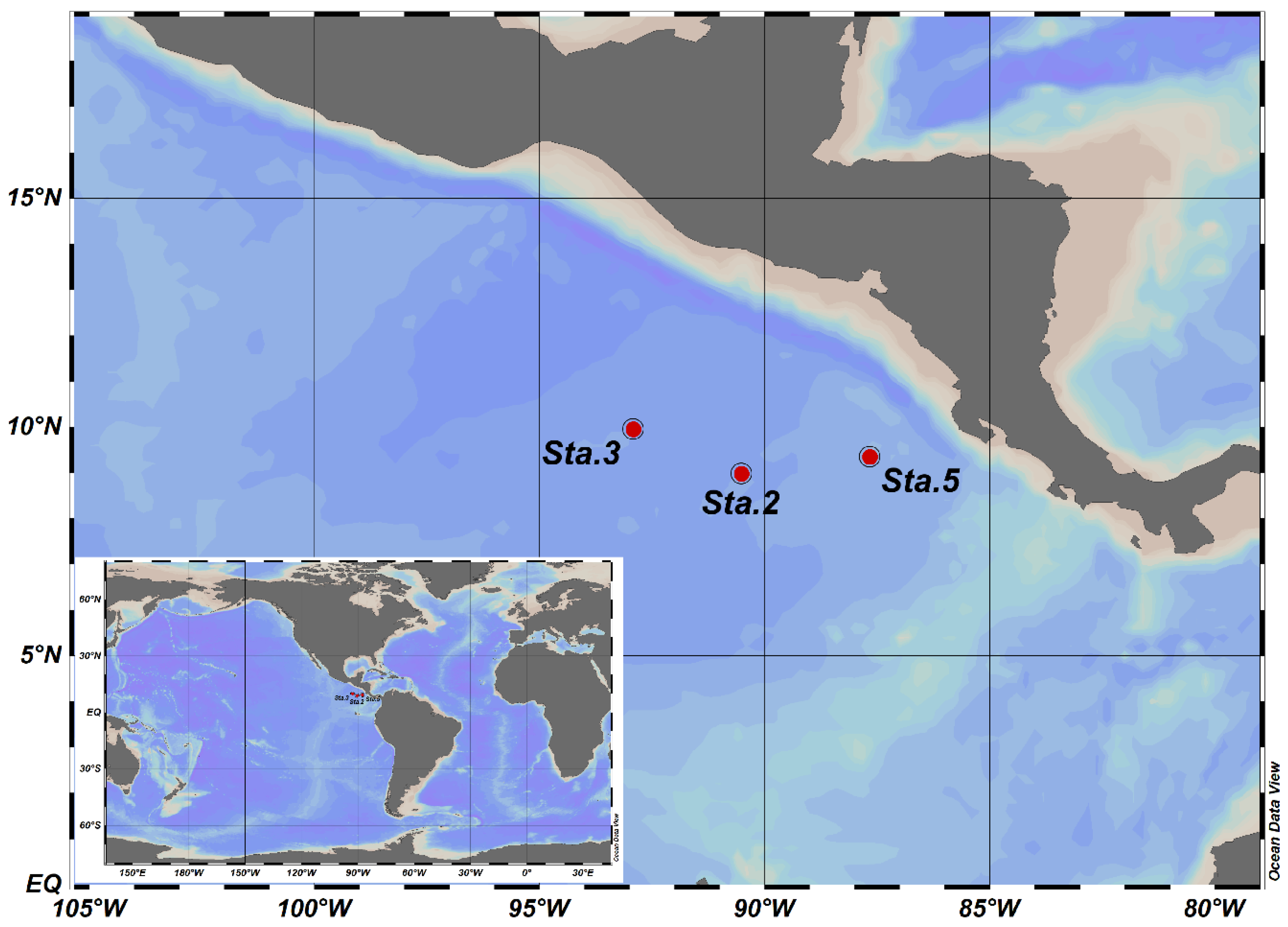
2.1. Sample Collection and Environmental Factor Measurement
2.2. DNA Extraction, PCR Amplification and High-Throughput Sequencing
2.3. Sequence Processing
2.4. Analyzing the Relationship between AOA and Environmental Parameters
3. Results and Discussion
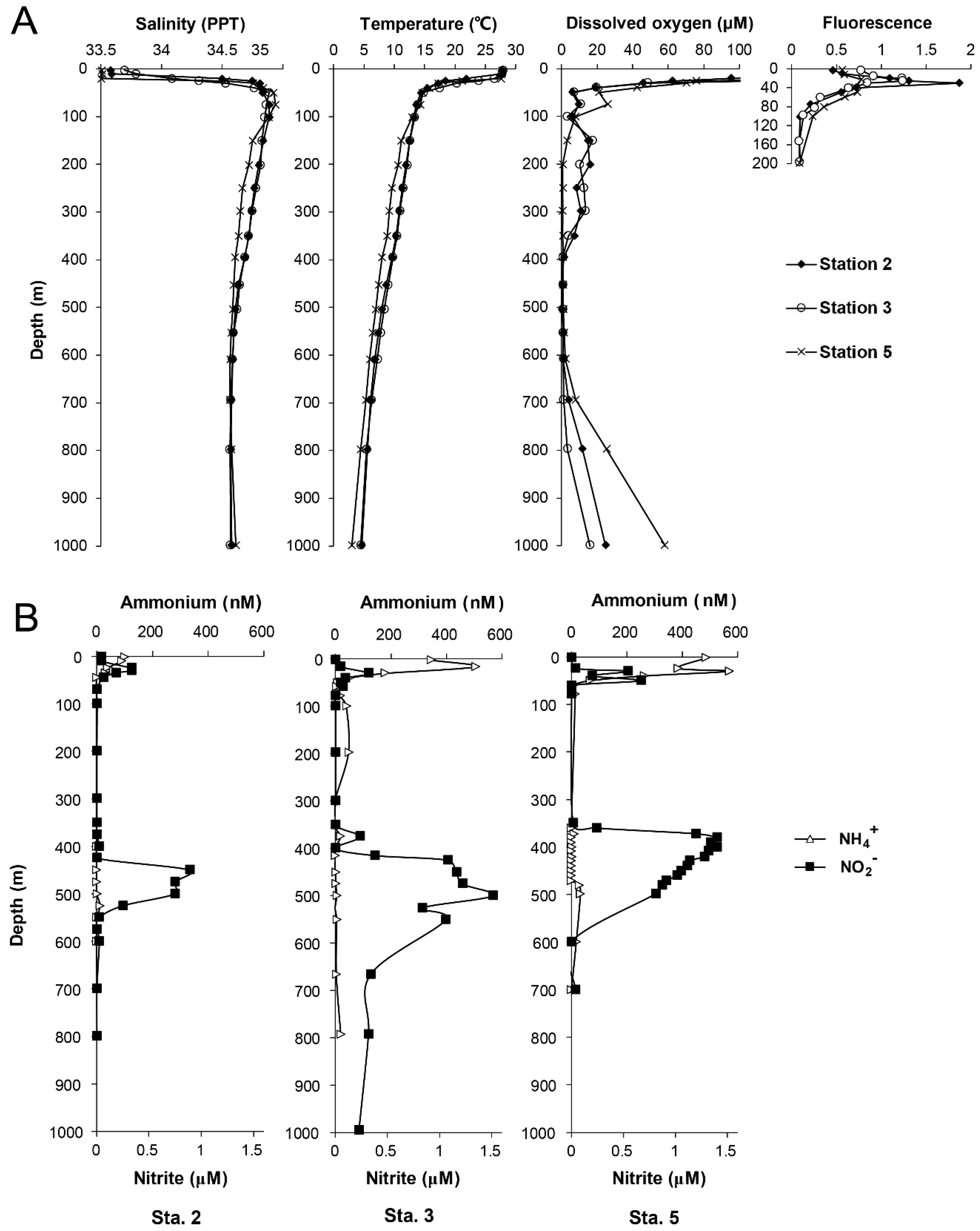
3.1. Hydrographic Conditions and Chemical Profiles in Costa Rica Dome
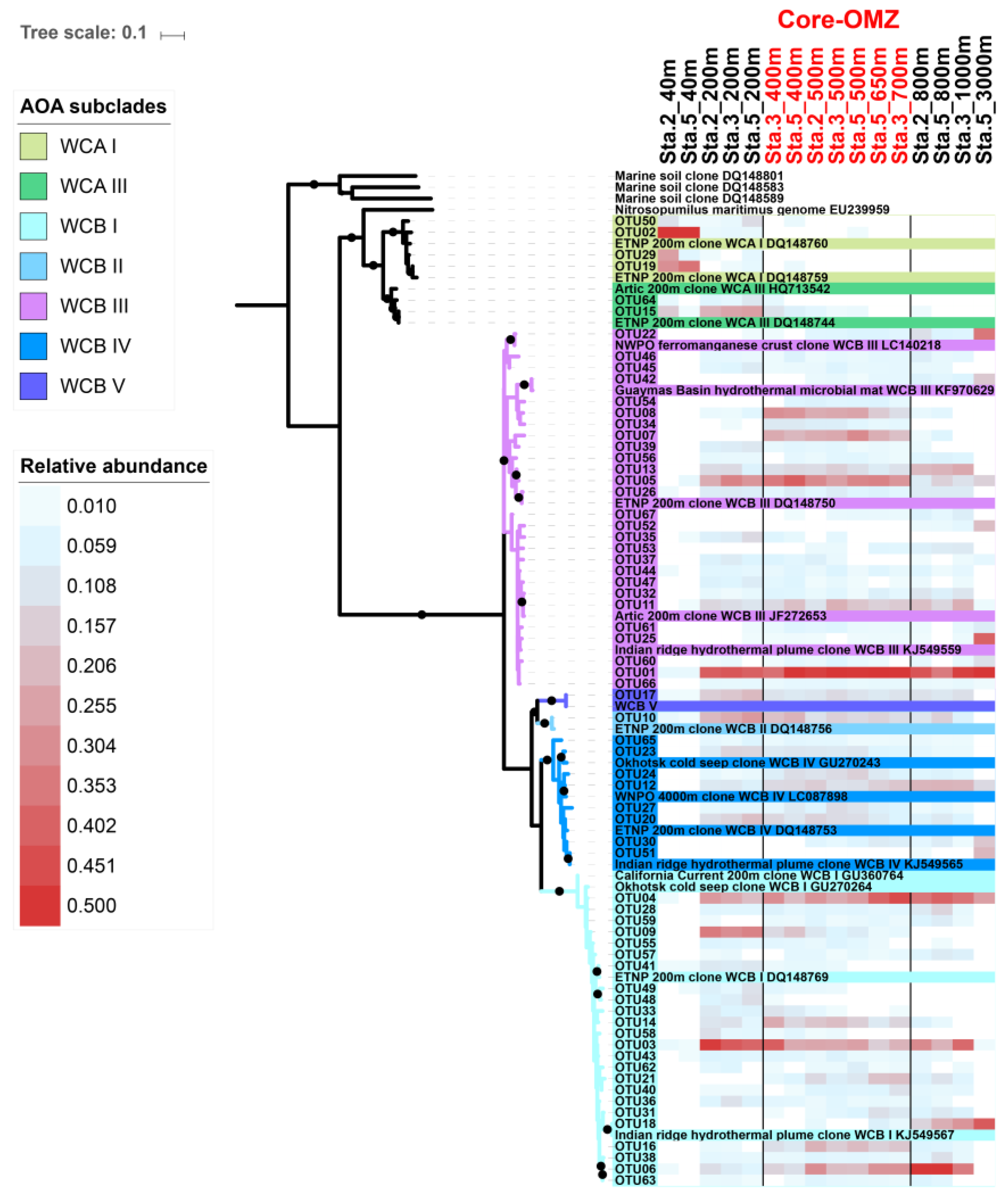
3.2. Phylogenetic Diversity of AOA in the CRD
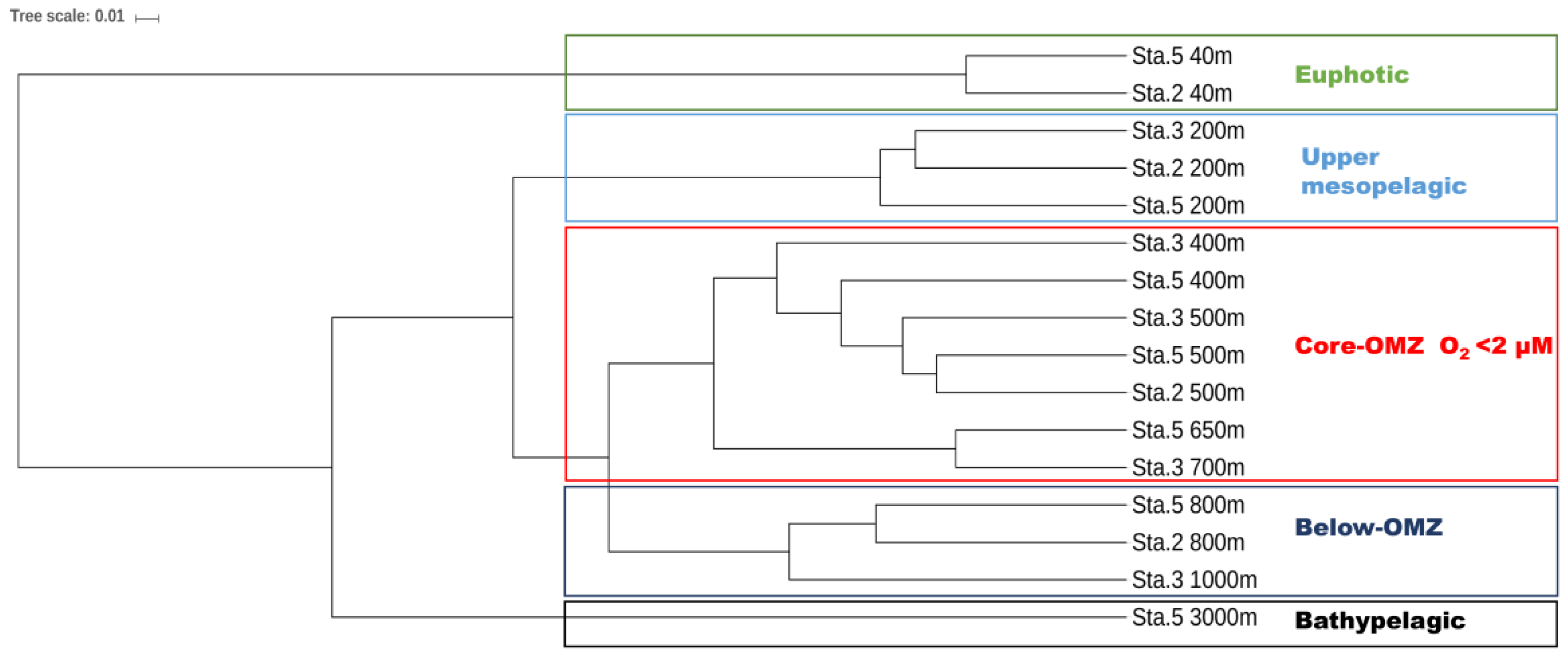
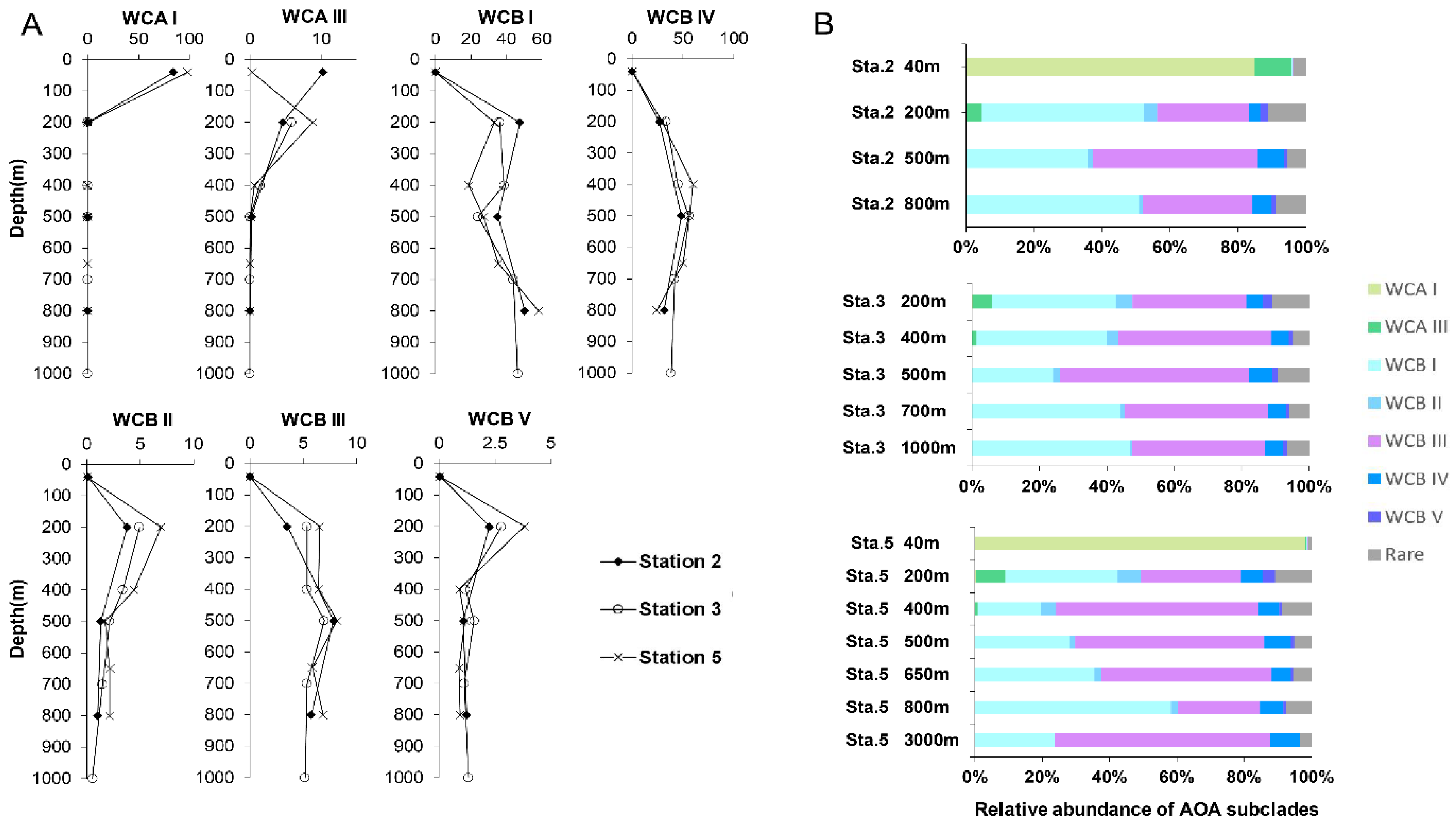
3.3. Differential Vertical Distribution of AOA Subclades
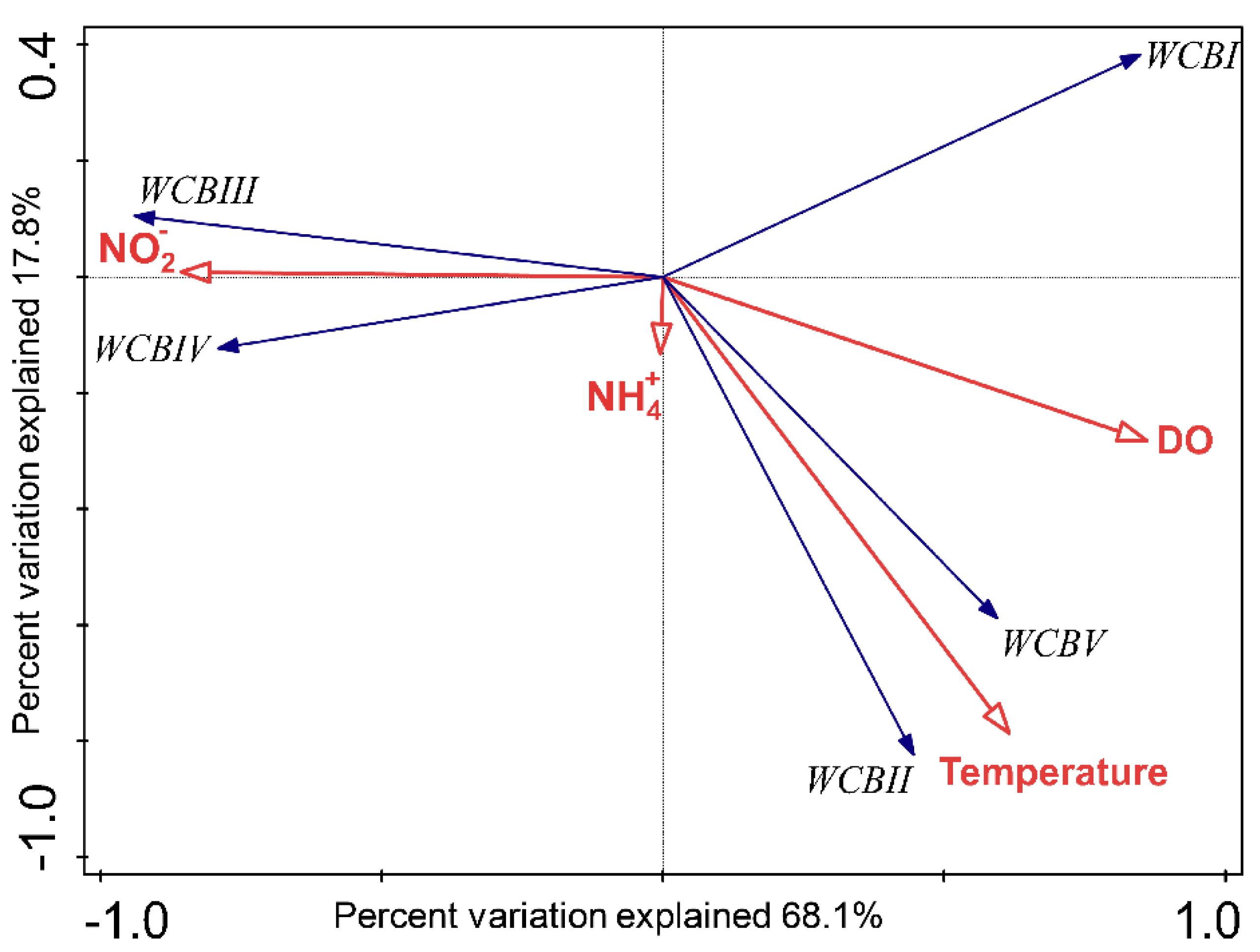
3.4. Specific WCB Phylotypes Selected by Environmental Conditions in OMZ
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wankel, S.D.; Kendall, C.; Pennington, J.T.; Chavez, F.P.; Paytan, A. Nitrification in the euphotic zone as evidenced by nitrate dual isotopic composition: Observations from Monterey Bay, California. Glob. Biogeochem. Cy. 2007, 21, GB2009. [Google Scholar] [CrossRef]
- Yool, A.; Martin, A.P.; Fernandez, C.; Clark, D.R. The significance of nitrification for oceanic new production. Nature 2007, 447, 999–1002. [Google Scholar] [CrossRef] [PubMed]
- Dore, J.E.; Popp, B.N.; Karl, D.M.; Sansone, F.J. A large source of atmospheric nitrous oxide from subtropical North Pacific surface waters. Nature 1998, 396, 63–66. [Google Scholar] [CrossRef]
- Ward, B.B. Nitrification and ammonification in aquatic systems. Life Support Biosph. Sci. 1996, 3, 25–29. [Google Scholar] [PubMed]
- Beman, J.M.; Popp, B.N.; Francis, C.A. Molecular and biogeochemical evidence for ammonia oxidation by marine Crenarchaeota in the Gulf of California. Isme. J. 2008, 2, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Francis, C.A.; Roberts, K.J.; Beman, J.M.; Santoro, A.E.; Oakley, B.B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. USA 2005, 102, 14683–14688. [Google Scholar] [CrossRef]
- Santoro, A.E.; Casciotti, K.L.; Francis, C.A. Activity, abundance and diversity of nitrifying archaea and bacteria in the central California Current. Env. Microbiol. 2010, 12, 1989–2006. [Google Scholar] [CrossRef]
- Lam, P.; Jensen, M.M.; Lavik, G.; McGinnis, D.F.; Muller, B.; Schubert, C.J.; Amann, R.; Thamdrup, B.; Kuypers, M.M. Linking crenarchaeal and bacterial nitrification to anammox in the Black Sea. Proc. Natl. Acad. Sci. USA 2007, 104, 7104–7109. [Google Scholar] [CrossRef]
- Molina, V.; Belmar, L.; Ulloa, O. High diversity of ammonia-oxidizing archaea in permanent and seasonal oxygen-deficient waters of the eastern South Pacific. Env. Microbiol. 2010, 12, 2450–2465. [Google Scholar] [CrossRef]
- Peng, X.; Jayakumar, A.; Ward, B.B. Community composition of ammonia-oxidizing archaea from surface and anoxic depths of oceanic oxygen minimum zones. Front Microbiol. 2013, 4, 177. [Google Scholar] [CrossRef]
- Mincer, T.J.; Church, M.J.; Taylor, L.T.; Preston, C.; Karl, D.M.; DeLong, E.F. Quantitative distribution of presumptive archaeal and bacterial nitrifiers in Monterey Bay and the North Pacific Subtropical Gyre. Env. Microbiol. 2007, 9, 1162–1175. [Google Scholar] [CrossRef]
- Wuchter, C.; Abbas, B.; Coolen, M.J.; Herfort, L.; van Bleijswijk, J.; Timmers, P.; Strous, M.; Teira, E.; Herndl, G.J.; Middelburg, J.J.; et al. Archaeal nitrification in the ocean. Proc. Natl. Acad. Sci. USA 2006, 103, 12317–12322. [Google Scholar] [CrossRef] [PubMed]
- Martens-Habbena, W.; Berube, P.M.; Urakawa, H.; de la Torre, J.R.; Stahl, D.A. Ammonia oxidation kinetics determine niche separation of nitrifying Archaea and Bacteria. Nature 2009, 461, 976–979. [Google Scholar] [CrossRef] [PubMed]
- Konneke, M.; Schubert, D.M.; Brown, P.C.; Hugler, M.; Standfest, S.; Schwander, T.; Schada von Borzyskowski, L.; Erb, T.J.; Stahl, D.A.; Berg, I.A. Ammonia-oxidizing archaea use the most energy-efficient aerobic pathway for CO2 fixation. Proc. Natl. Acad. Sci. USA 2014, 111, 8239–8244. [Google Scholar] [CrossRef] [PubMed]
- Cheung, S.; Mak, W.; Xia, X.; Lu, Y.; Cheung, Y.; Liu, H. Overlooked Genetic Diversity of Ammonia Oxidizing Archaea Lineages in the Global Oceans. J. Geophysi. Res.: Biogeosciences 2019. [Google Scholar] [CrossRef]
- Biller, S.J.; Mosier, A.C.; Wells, G.F.; Francis, C.A. Global biodiversity of aquatic ammonia-oxidizing archaea is partitioned by habitat. Front Microbiol. 2012, 3. [Google Scholar] [CrossRef]
- Erguder, T.H.; Boon, N.; Wittebolle, L.; Marzorati, M.; Verstraete, W. Environmental factors shaping the ecological niches of ammonia-oxidizing archaea. Fems. Microbiol. Rev. 2009, 33, 855–869. [Google Scholar] [CrossRef]
- Luo, H.; Tolar, B.B.; Swan, B.K.; Zhang, C.L.; Stepanauskas, R.; Ann Moran, M.; Hollibaugh, J.T. Single-cell genomics shedding light on marine Thaumarchaeota diversification. Isme. J. 2014, 8, 732–736. [Google Scholar] [CrossRef]
- Qin, W.; Meinhardt, K.A.; Moffett, J.W.; Devol, A.H.; Virginia Armbrust, E.; Ingalls, A.E.; Stahl, D.A. Influence of oxygen availability on the activities of ammonia-oxidizing archaea. Env. Microbiol. Rep. 2017, 9, 250–256. [Google Scholar] [CrossRef]
- Santoro, A.E.; Richter, R.A.; Dupont, C.L. Planktonic Marine Archaea. Annu. Rev. Mar. Sci. 2019, 11, 131–158. [Google Scholar] [CrossRef]
- Alves, R.J.E.; Minh, B.Q.; Urich, T.; von Haeseler, A.; Schleper, C. Unifying the global phylogeny and environmental distribution of ammonia-oxidising archaea based on amoA genes. Nat. Commun. 2018, 9, 1517. [Google Scholar] [CrossRef] [PubMed]
- Konneke, M.; Bernhard, A.E.; de la Torre, J.R.; Walker, C.B.; Waterbury, J.B.; Stahl, D.A. Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 2005, 437, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A.E.; Dupont, C.L.; Richter, R.A.; Craig, M.T.; Carini, P.; McIlvin, M.R.; Yang, Y.; Orsi, W.D.; Moran, D.M.; Saito, M.A. Genomic and proteomic characterization of “Candidatus Nitrosopelagicus brevis”: An ammonia-oxidizing archaeon from the open ocean. Proc. Natl. Acad. Sci. USA 2015, 112, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Newell, S.E.; Babbin, A.R.; Jayakumar, A.; Ward, B.B. Ammonia oxidation rates and nitrification in the Arabian Sea. Glob. Biogeochem. Cy. 2011, 25. [Google Scholar] [CrossRef]
- Peng, X.F.; Fuchsman, C.A.; Jayakumar, A.; Warner, M.J.; Devol, A.H.; Ward, B.B. Revisiting nitrification in the Eastern Tropical South Pacific: A focus on controls. J. Geophys. Res.-Ocean. 2016, 121, 1667–1684. [Google Scholar] [CrossRef]
- Stewart, F.J.; Ulloa, O.; DeLong, E.F. Microbial metatranscriptomics in a permanent marine oxygen minimum zone. Env. Microbiol. 2012, 14, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Bristow, L.A.; Dalsgaard, T.; Tiano, L.; Mills, D.B.; Bertagnolli, A.D.; Wright, J.J.; Hallam, S.J.; Ulloa, O.; Canfield, D.E.; Revsbech, N.P. Ammonium and nitrite oxidation at nanomolar oxygen concentrations in oxygen minimum zone waters. Proc. Natl. Acad. Sci. USA 2016, 113, 10601–10606. [Google Scholar] [CrossRef]
- Landry, M.R.; De Verneil, A.; Goes, J.I.; Moffett, J.W. Plankton dynamics and biogeochemical fluxes in the Costa Rica Dome: Introduction to the CRD Flux and Zinc Experiments. J. Plankton Res. 2016, 38, 167–182. [Google Scholar] [CrossRef]
- Jing, H.M.; Cheung, S.Y.; Xia, X.M.; Suzuki, K.; Nishioka, J.; Liu, H.B. Geographic Distribution of Ammonia-Oxidizing Archaea along the Kuril Islands in the Western Subarctic Pacific. Front Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Holmes, R.M.; Aminot, A.; Kerouel, R.; Hooker, B.A.; Peterson, B.J. A simple and precise method for measuring ammonium in marine and freshwater ecosystems. Can J. Fish Aquat. Sci. 1999, 56, 1801–1808. [Google Scholar] [CrossRef]
- Buchwald, C.; Santoro, A.E.; Stanley, R.H.R.; Casciotti, K.L. Nitrogen cycling in the secondary nitrite maximum of the eastern tropical North Pacific off Costa Rica. Glob. Biogeochem. Cy. 2015, 29, 2061–2081. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Env. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Acland, A.; Agarwala, R.; Barrett, T.; Beck, J.; Benson, D.A.; Bollin, C.; Bolton, E.; Bryant, S.H.; Canese, K.; Church, D.M.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2014, 42, D7–D17. [Google Scholar] [CrossRef]
- Logares, R.; Audic, S.; Bass, D.; Bittner, L.; Boutte, C.; Christen, R.; Claverie, J.M.; Decelle, J.; Dolan, J.R.; Dunthorn, M. Patterns of rare and abundant marine microbial eukaryotes. Curr. Biol. 2014, 24, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Šmilauer, P.; Lepš, J. Multivariate analysis of ecological data using Canoco 5, 2nd ed.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. xii–362. [Google Scholar]
- Santoro, A.E.; Saito, M.A.; Goepfert, T.J.; Lamborg, C.H.; Dupont, C.L.; DiTullio, G.R. Thaumarchaeal ecotype distributions across the equatorial Pacific Ocean and their potential roles in nitrification and sinking flux attenuation. Limnol Oceanogr. 2017, 62, 1984–2003. [Google Scholar] [CrossRef]
- Xia, X.; Guo, W.; Liu, H. Basin Scale Variation on the Composition and Diversity of Archaea in the Pacific Ocean. Front Microbiol. 2017, 8, 2057. [Google Scholar] [CrossRef]
- Reji, L.; Tolar, B.B.; Smith, J.M.; Chavez, F.P.; Francis, C.A. Differential co-occurrence relationships shaping ecotype diversification within Thaumarchaeota populations in the coastal ocean water column. Multidiscip. J. Microb. Ecol. 2019, 13, 1144–1158. [Google Scholar] [CrossRef]
- Hallam, S.J.; Mincer, T.J.; Schleper, C.; Preston, C.M.; Roberts, K.; Richardson, P.M.; DeLong, E.F. Pathways of carbon assimilation and ammonia oxidation suggested by environmental genomic analyses of marine Crenarchaeota. Plos. Biol. 2006, 4, 520–536. [Google Scholar] [CrossRef] [PubMed]
- Prosser, J.I.; Nicol, G.W. Archaeal and bacterial ammonia-oxidisers in soil: The quest for niche specialisation and differentiation. Trends Microbiol. 2012, 20, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Amin, S.A.; Martens-Habbena, W.; Walker, C.B.; Urakawa, H.; Devol, A.H.; Ingalls, A.E.; Moffett, J.W.; Armbrust, E.V.; Stahl, D.A. Marine ammonia-oxidizing archaeal isolates display obligate mixotrophy and wide ecotypic variation. Proc. Natl. Acad. Sci. USA 2014, 111, 12504–12509. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.L.; Feng, X.Y.; Huang, Y.J.; Wang, H.; Hu, Z.; Clingenpeel, S.; Swan, B.K.; Fonseca, M.M.; Posada, D.; Stepanauskas, R.; et al. Phylogenomics suggests oxygen availability as a driving force in Thaumarchaeota evolution. Multidiscip. J. Microb. Ecol. 2019, 13, 2150–2161. [Google Scholar] [CrossRef]
- Beman, J.M.; Popp, B.N.; Alford, S.E. Quantification of ammonia oxidation rates and ammonia-oxidizing archaea and bacteria at high resolution in the Gulf of California and eastern tropical North Pacific Ocean. Limnol Oceanogr. 2012, 57, 711–726. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Lon. (W) | Lat. (N) | Depth (m) | DO (μM) | Sequence number | Shannon index | Margalef Richness | Good’s Coverage index |
|---|---|---|---|---|---|---|---|---|
| Sta. 2 | 90.51 | 8.98 | 40 200 500 800 | 19.219 15.87 30.310 11.680 | 4586 7394 6946 6554 | 0.984 3.111 3.026 2.838 | 4.11 19.18 13.39 14.75 | 0.996 0.981 0.980 0.977 |
| Sta. 3 | 92.92 | 9.95 | 200 400 500 700 1000 | 10.261 0.651 0.740 1.096 15.920 | 7438 6569 5413 6569 2435 | 3.170 2.935 3.162 2.782 2.710 | 18.51 10.67 15.67 12.08 9.90 | 0.981 0.984 0.971 0.983 0.981 |
| Sta. 5 | 87.66 | 9.35 | 40 200 400 500 650 800 3000 | 75.825 7.702 0.640 0.739 1.379 7.850 108.754 | 5764 6258 7689 7521 7171 5426 3874 | 0.456 3.239 3.081 2.954 2.839 2.841 2.295 | 1.28 14.98 15.62 11.83 12.60 11.76 4.74 | 0.997 0.981 0.982 0.986 0.985 0.984 0.996 |
| OTU No. | Subclades | OMZs | Accession No. | Sequence similarity | Spearman correlation (rs) with DO |
|---|---|---|---|---|---|
| OTU07 | WCB III | Arabian Sea | KF512370 | 100.00% | −0.782 |
| OTU08 | WCB III | Gulf of California | EU340498 | 100.00% | −0.888 |
| OTU16 | WCB I | Gulf of California | KC596407 | 100.00% | −0.780 |
| OTU06 | WCB I | Arabian Sea | KF512353 | 100.00% | −0.718 |
| OTU14 | WCB I | Arabian Se | KF512366 | 98.73% | −0.815 |
| OTU34 | WCB III | Gulf of California | EU340551 | 100.00% | −0.879 |
| OTU31 | WCB I | Gulf of California | KC596421 | 100.00% | |
| OTU33 | WCB I | Gulf of California | KC596422 | 99.68% | |
| OTU54 | WCB III | Gulf of California | EU340552 | 100.00% | −0.763 |
| OTU30 | WCB IV | Gulf of California | KC596405 | 100.00% | |
| OTU11 | WCB III | Gulf of California | KC596420 | 100.00% | |
| OTU57 | WCB I | Arabian Sea | KF512363 | 100.00% | −0.619 |
| OTU21 | WCB I | CRD | This study | −0.734 | |
| OTU24 | WCB IV | CRD | This study | −0.825 | |
| OTU27 | WCB IV | CRD | This study | −0.679 | |
| OTU61 | WCB III | CRD | This study | −0.837 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Xia, X.; Cheung, S.; Jing, H.; Liu, H. Differential Distribution and Determinants of Ammonia Oxidizing Archaea Sublineages in the Oxygen Minimum Zone off Costa Rica. Microorganisms 2019, 7, 453. https://doi.org/10.3390/microorganisms7100453
Lu Y, Xia X, Cheung S, Jing H, Liu H. Differential Distribution and Determinants of Ammonia Oxidizing Archaea Sublineages in the Oxygen Minimum Zone off Costa Rica. Microorganisms. 2019; 7(10):453. https://doi.org/10.3390/microorganisms7100453
Chicago/Turabian StyleLu, Yanhong, Xiaomin Xia, Shunyan Cheung, Hongmei Jing, and Hongbin Liu. 2019. "Differential Distribution and Determinants of Ammonia Oxidizing Archaea Sublineages in the Oxygen Minimum Zone off Costa Rica" Microorganisms 7, no. 10: 453. https://doi.org/10.3390/microorganisms7100453
APA StyleLu, Y., Xia, X., Cheung, S., Jing, H., & Liu, H. (2019). Differential Distribution and Determinants of Ammonia Oxidizing Archaea Sublineages in the Oxygen Minimum Zone off Costa Rica. Microorganisms, 7(10), 453. https://doi.org/10.3390/microorganisms7100453