Isolation, Identification, and Characterization of Pectinolytic Yeasts for Starter Culture in Coffee Fermentation



Abstract
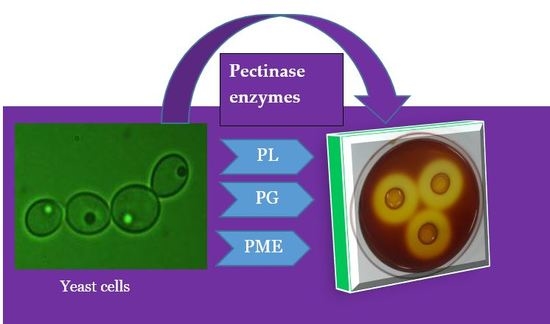
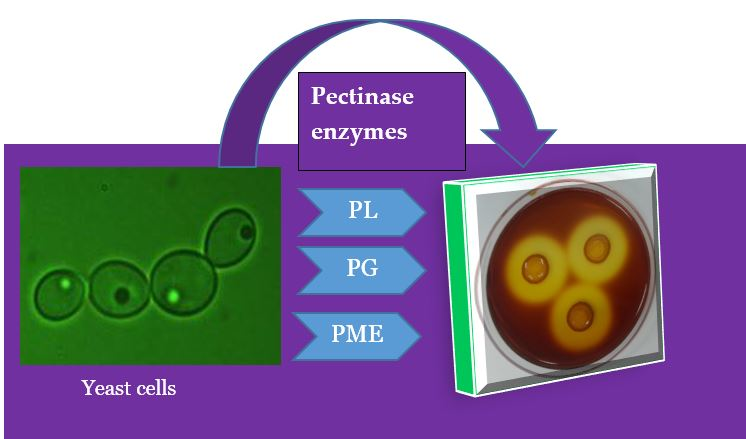
1. Introduction
2. Materials and Methods
2.1. Spontaneous Fermentation and Yeast Isolation
2.2. Pectinolytic Yeast Screening
2.3. Pectin Degradation Index (PDI) %
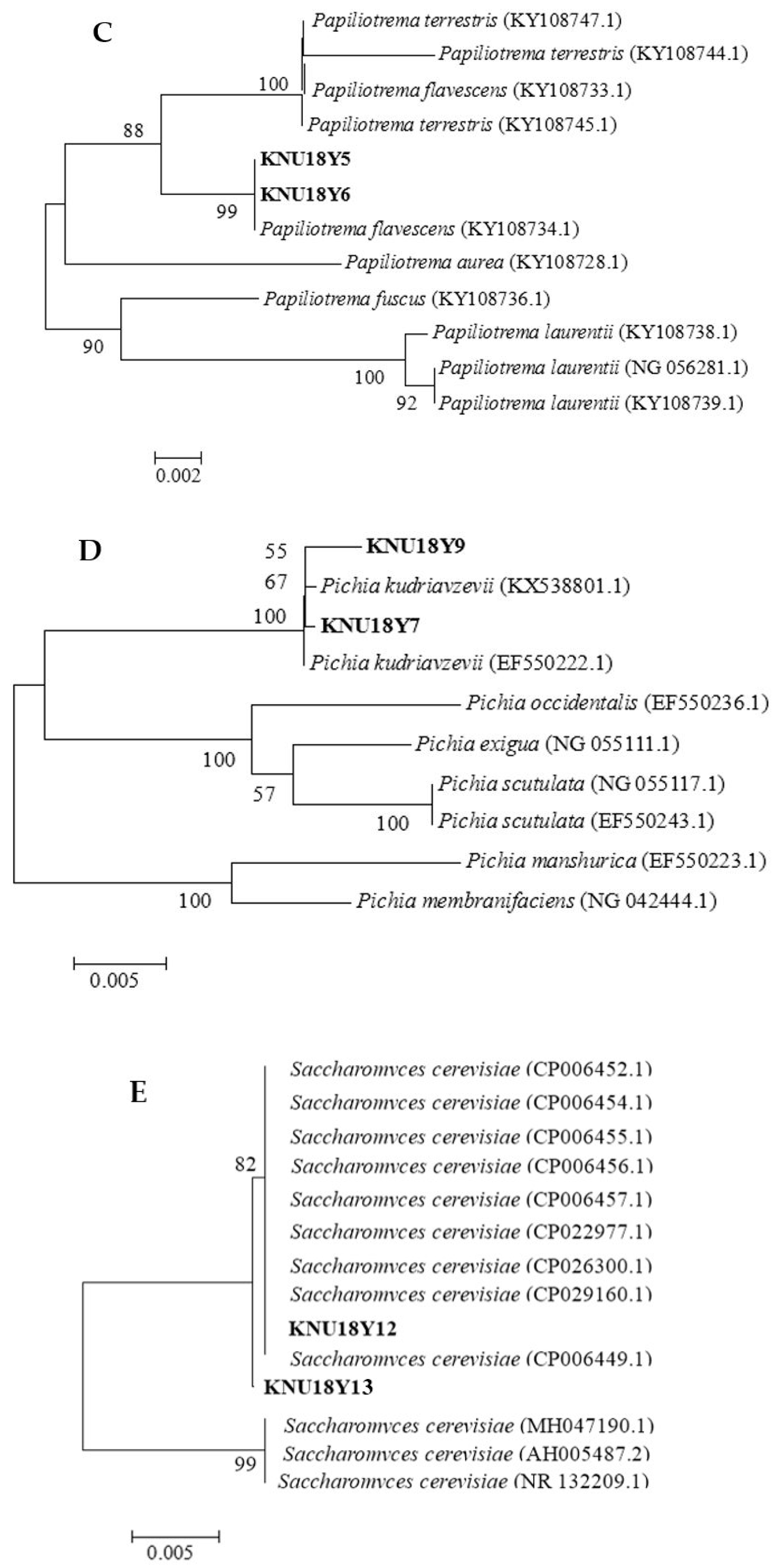
2.4. Molecular Identification and Phylogenetic Tree Analysis of Yeasts
2.5. Synthetic Pectin Media and Coffee Pulp Media Preparation for Enzymatic Assays
2.6. Enzyme Assays
2.6.1. Pectin Lyase Activity
2.6.2. Polygalacturonase Activity
2.6.3. Pectin Methylesterase
2.7. Statistical Analysis
3. Results
3.1. Isolation, Selection, and Morphological Characterization of Yeasts
3.2. Pectin Degradation Index (PDI) %
Molecular Identification and Phylogenetic Trees
3.3. Fermentation Characteristics
3.4. Enzyme Assays
3.4.1. Pectin Lyase
3.4.2. Polygalacturonase Activity
3.4.3. Pectin Methylesterase
4. Discussion
5. Conclusions
Author Contributions
Conflicts of Interest
References
- FAO. Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 4 November 2016).
- Sakiyama, N.S.; Ferrao, M.A.G. Botany and production of coffee. In Cocoa and Coffee Fermentations; Schwan, R.F., Fleet, G.H., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 341–365. [Google Scholar]
- International Coffee Organization (ICO). Monthly Coffee Market. Report (2017/18); International Coffee Organization (ICO): London, UK, 2017. [Google Scholar]
- De Bruyn, F.; Zhang, S.J.; Pothakos, V.; Torres, J.; Lambot, C.; Moroni, A.V.; De Vuyst, L. Exploring the impacts of postharvest processing on the microbiota and metabolite profiles during green coffee bean production. Appl. Environ. Microbiol. 2017, 83, e02398. [Google Scholar]
- Murthy, P.S.; Naidu, M.M. Sustainable management of coffee industry by-products and value addition—A review. Resour. Conserv. Recy. 2012, 66, 45–58. [Google Scholar] [CrossRef]
- Rattan, S.; Parande, A.K.; Nagaraju, V.D.; Ghiwari, G.K. A comprehensive review on utilization of wastewater from coffee processing. Environ. Sci. Pollut. R 2015, 22, 6461–6472. [Google Scholar] [CrossRef] [PubMed]
- Silva, C. Microbial activity during coffee fermentation. Cocoa Coffee Ferment. 2014, 368–423. [Google Scholar]
- Huch, M.; Franz, C.M.A.P. Coffee. In Advances in Fermented Foods and Beverages; Woodhead Publishing: Sawston, UK, 2015; pp. 501–513. [Google Scholar]
- Vilela, D.M.; Pereira, G.V.M.; Ferreira Silva, C.; Batista, L.R.; Schwan, R.F. Molecular ecology and polyphasic characterization of the microbiota associated with semi-dry processed coffee (Coffea arabica L.). Food Microbiol. 2010, 27, 1128–1135. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, H.A.; Rodríguez-Jasso, R.M.; Rodríguez, R.; Contreras-Esquivel, J.C.; Aguilar, C.N. Pectinase production from lemon peel pomace as support and carbon source in solid-state fermentation column-tray bioreactor. Biochem. Eng. J. 2012, 65, 90–95. [Google Scholar] [CrossRef]
- Amin, F.; Bhatti, H.N.; Bilal, M. Recent advances in the production strategies of microbial pectinases—A review. Int. J. Biol. Macromol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Haile, M.; Kang, W.H. The role of microbes in coffee fermentation and their impact on coffee quality. J. Food Qual. 2019. [Google Scholar] [CrossRef]
- Combo, A.M.M.; Aguedo, M.; Goffin, D.; Wathelet, B.; Paquot, M. Enzymatic production of pectic oligosaccharides from polygalacturonic acid with commercial pectinase preparations. Food Bioprod. Process. 2012, 90, 588–596. [Google Scholar] [CrossRef]
- Patidar, M.K.; Nighojkar, S.; Kumar, A.; Nighojkar, A. Pectinolytic enzymes-solid state fermentation, assay methods and applications in fruit juice industries: A review. Biotech 2018, 8, 199. [Google Scholar] [CrossRef]
- Haile, M.; Kang, W.H. Antioxidant Activity, Total Polyphenol, Flavonoid and Tannin Contents of Fermented Green Coffee Beans with Selected Yeasts. Fermentation 2019, 5, 29. [Google Scholar] [CrossRef]
- Schwan, R.F.; Cooper, R.; Wheals, A.E. Endo polyglacturonase secretion by Kluyveromyces marxianus. Enzym. Microb. Tech. 1997, 4, 247–256. [Google Scholar]
- Silva, C.F.; Vilela, D.M.; de Souza Cordeiro, C.; Duarte, W.F.; Dias, D.R.; Schwan, R.F. Evaluation of a potential starter culture for enhance quality of coffee fermentation. World J. Microbiol. Biotechnol. 2013, 29, 235–247. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for quantitation of microgram quantities of protein utilize principle of protein byebinding. Anal. Biochem. 1976, 72, 248–251. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analyses of the Association of Official Analytical Chemists, 7th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000; pp. 915–922. [Google Scholar]
- Bitter, V.; Muir, H.M. Modifical uronic acid carbazole reaction. Anal. Biochem. 1962, 4, 330–334. [Google Scholar] [CrossRef]
- Pitt, O. Pectic lyase from Rhoma medicabinis var. pinodella. Methods Enzymol. 1988, 161, 350–365. [Google Scholar]
- Kashyap, D.R.; Chandra, S.; Kaul, A.; Tewari, R. Production, purification and characterization of pectinase from a Bacillus sp. DT7. World J. Microbiol. Biotechnol. 2000, 16, 277–282. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicilic acid reagent for determination of reducing sugar. Anal. Biochem. 1959, 31, 426–428. [Google Scholar]
- Baracat, M.C.; Valentin, C.; Muchovej, J.J.; Silva, D.O. Selection of pectinolytic fungi for degumming of natural fibers. Biotechnol. Lett. 1989, 11, 899–902. [Google Scholar] [CrossRef]
- Garg, G.; Singh, A.; Kaur, A.; Singh, R.; Kaur, J.; Mahajan, R. Microbial pectinases: An ecofriendly tool of nature for industries. Biotech. 2016, 6, 47. [Google Scholar] [CrossRef]
- Schwan, R.F.; Pereira, G.V.M.; Fleet, G.H. Microbial activities during cocoa fermentation. In Cocoa and Coffee Fermentations; Schwan, R.F., Fleet, G.H., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 129–192. [Google Scholar]
- Vinícius de Melo Pereira, G.; Soccol, V.T.; Brar, S.K.; Neto, E.; Soccol, C.R. Microbial ecology and starter culture technology in coffee processing. Crit. Rev. Food Sci. Nutr. 2017, 57, 2775–2788. [Google Scholar] [CrossRef]
- Kamassah, A.K.; Saalia, F.K.; Osei-Fosu, P.; Mensah-Brown, H.; Sinayobye, E.; Tano-Debrah, K. Fermentation capacity of yeasts using mango (Mangifera indica Linn.) as substrate. Food Sci. Qual. Manag. 2013, 22, 69–78. [Google Scholar]
- Murthy, P.S.; Naidu, M.M. Improvement of robusta coffee fermentation with microbial enzymes. Eur. J. Appl. Sci. 2011, 3, 130–139. [Google Scholar]
- Kohli, P.; Kalia, M.; Gupta, R. Pectin Methylesterases: A Review. J. Bioproc. Biotech. 2015, 5, 1–7. [Google Scholar]
- Carrasco, M.; Rozas, J.M.; Alcaíno, J.; Cifuentes, V.; Baeza, M. Pectinase secreted by psychrotolerant fungi: Identification, molecular characterization and heterologous expression of a cold-active polygalacturonase from Tetracladium sp. Microb. Cell Factor. 2019, 18, 45. [Google Scholar] [CrossRef] [PubMed]
- Oumer, O.J.; Abate, D. Comparative Studies of Pectinase Production by Bacillus subtilis strain Btk 27 in Submerged and Solid-State Fermentations. Biomed. Res. Int. 2018. [Google Scholar] [CrossRef] [PubMed]
- Koffi, O.; Samagaci, L.; Goualie, B.; Niamke, S. Screening of potential yeast starters with high ethanol production for a small-scale cocoa fermentation in Ivory Coast. Food Environ. Safety J. 2018, 17. [Google Scholar]
- Romero Cortes, T.; Cuervo-Parra, J.A.; Jose Robles-Olvera, V.; Rangel Cortes, E.; Lopez Perez, P.A. Experimental and kinetic production of ethanol using mucilage juice residues from cocoa processing. Int. J. Chem. React. Eng. 2018, 16. [Google Scholar] [CrossRef]
- Martos, M.A.; Zubreski, E.R.; Garro, O.A.; Hours, R.A. Production of Pectinolytic enzymes by the yeast Wickerhanomyces anomalus isolated from citrus fruits peels. Biotechnol. Res. Int. 2013. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, G.S. Risk assessment of fungi reported from humans and animals. Mycoses 1996, 39, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Dhembare, A.J.; Kakad, S.L.; Rajani, R. Effect of pH, temperature and kinetics of pectinase enzyme using Aspergillus niger by solid-state of fermentation. Der. Pharm. Sin. 2015, 6, 1–5. [Google Scholar]
- Azzaz, H.H.; Murad, H.A.; Kholif, A.M.; Morsy, T.A.; Mansour, A.M.; El-Sayed, H.M. Pectinase production optimization and its application in banana fiber degradation. Egypt. J. Nutr. Foods 2013, 16, 117–125. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence | Type |
|---|---|---|
| Forward | 5’-ACCCGCTGAACTTAAGC -3’ | Universal |
| Reverse | 5’ -TACTACCACCAAGATCT -3’ | Universal |
| Yeasts | Shape | Color | Elevation | Surface | Margin |
|---|---|---|---|---|---|
| W. anomalus KNU18Y3 | Circular | White | Raised | Smooth | Entire |
| S. fibuligera KNU18Y4 | Circular | White | Convex | Rough | Entire |
| P. flavescens KNU18Y5 | Circular | Yellow | Pulvinate | Rough | Entire |
| P. flavescens KNU18Y6 | Circular | Yellow | Pulvinate | Smooth | Entire |
| P. kudriavzevii KNU18Y7 | Circular | White | Umbonate | Smooth | Entire |
| P. kudriavzevii KNU18Y9 | Circular | White | Pulvinate | Smooth | Entire |
| S. cerevisiae KNU18Y12 | Circular | White | Convex | Smooth | Entire |
| S. cerevisiae KNU18Y13 | Circular | White | Pulvinate | Smooth | Entire |
| Yeasts | PDI % |
|---|---|
| W. anomalus KNU18Y3 | 160b ± 1.73 |
| S. fibuligera KNU18Y4 | 178a ± 4.04 |
| P. flavescens KNU18Y5 | 129c ± 4.62 |
| P. flavescens KNU18Y6 | 152b ± 4.04 |
| P. kudriavzevii KNU18Y7 | 110d ± 2.89 |
| P. kudriavzevii KNU18Y9 | 125c ± 4.04 |
| S. cerevisiae KNU18Y12 | 121cd ± 2.31 |
| S. cerevisiae KNU18Y13 | 118cd ± 4.62 |
| Accession Number | Yeast Species Name | Strain Code |
|---|---|---|
| MH483547 | Wickerhamomyces anomalus | KNU18Y3 |
| MH483929 | Saccharomycopsis fibuligera | KNU18Y4 |
| MH484046 | Papiliotrema flavescens | KNU18Y5 |
| MH485393 | Papiliotrema flavescens | KNU18Y6 |
| MH488970 | Pichia kudriavzevii | KNU18Y7 |
| MH487468 | Pichia kudriavzevii | KNU18Y9 |
| MH491423 | Saccharomyces cerevisiae | KNU18Y12 |
| MH488975 | Saccharomyces cerevisiae | KNU18Y13 |
| Yeasts | Log CFU mL−1 | |||
|---|---|---|---|---|
| 24 h | 48 h | 72 h | 96 h | |
| W. anomalus KNU18Y3 | 7.08f ± 0.23 | 8.21b ± 0.32 | 7.22a ± 0.52 | 7.23a ± 0.10 |
| S. fibuligera KNU18Y4 | 6.80g ± 0.11 | 8.04e ± 0.52 | 7.11d ± 0.41 | 7.12c ± 0.08 |
| P. flavescens KNU18Y5 | 7.10e ± 0.21 | 8.17d ± 0.46 | 7.16b ± 0.36 | 7.15b ± 0.15 |
| P. flavescens KNU18Y6 | 7.13d ±0.14 | 8.20b ± 0.26 | 7.23a ± 0.21 | 7.22a ± 0.17 |
| P. kudriavzevii KNU18Y7 | 7.17c ±0.12 | 8.19c ± 0.13 | 7.15c ± 0.25 | 7.15bc ± 0.20 |
| P. kudriavzevii KNU18Y9 | 7.13d ± 0.15 | 8.18cd ± 0.26 | 7.16b ±0.14 | 7.17b ± 0.16 |
| S. cerevisiae KNU18Y12 | 7.19b ± 0.09 | 8.18cd ± 0.18 | 7.17b ± 0.11 | 7.16b ± 0.19 |
| S. cerevisiae KNU18Y13 | 7.22a ± 0.18 | 8.23a ± 0.12 | 7.22a ±0.32 | 7.21a ± 0.15 |
| Strains Code | pH | Brix° | ||||||
|---|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 96 h | 24 h | 48 h | 72 h | 96 h | |
| KNU18Y3 | 4.93b ± 0.21 | 4.62g ± 0.14 | 4.60e ± 0.23 | 4.75h ± 0.41 | 2.13b ± 0.11 | 0.9d ± 0.04 | 0.46d ± 0.02 | 0.40b ± 0.01 |
| KNU18Y4 | 5.23a ± 0.42 | 5.01a ± 0.11 | 4.86c ± 0.27 | 4.83f ± 0.35 | 2.93a ± 0.23 | 1.77a ± 0.12 | 1.75a ± 0.04 | 0.96a ± 0.02 |
| KNU18Y5 | 5.04ab ± 0.12 | 4.97b ± 0.13 | 5.28b ± 0.17 | 5.86c ± 0.23 | 2.10b ± 0.14 | 1.66ab ± 0.08 | 1.15b ± 0.03 | 1.03a ± 0.05 |
| KNU18Y6 | 4.94b ± 0.13 | 4.77e ± 0.21 | 4.75cd ± 0.31 | 4.80g ± 0.24 | 2.06b ± 0.19 | 1.16cd ± 0.01 | 0.55c ± 0.06 | 0.46b ± 0.04 |
| KNU18Y7 | 4.95b ± 0.14 | 4.73f ± 0.23 | 5.68a ± 0.30 | 5.91b ± 0.41 | 1.80c ± 0.09 | 1.70ab ± 0.10 | 1.15b ± 0.07 | 1.01a ± 0.03 |
| KNU18Y9 | 4.99b ± 0.15 | 4.95c ±0.25 | 5.65a ± 0.24 | 5.96a ± 0.21 | 1.90c ± 0.18 | 1.77a ± 0.08 | 1.10b ± 0.07 | 1.01a ± 0.04 |
| KNU18Y12 | 4.86b ± 0.15 | 4.85d ± 0.15 | 4.82c ± 0.18 | 5.13d ± 0.17 | 1.30d ± 0.17 | 1.03d ± 0.06 | 0.65c ± 0.04 | 0.60b ± 0.05 |
| KNU18Y13 | 4.90b ± 0.17 | 4.85d ± 0.14 | 4.64de ± 0.19 | 5.08e ± 0.12 | 1.20d ± 0.13 | 1.40bc ± 0.10 | 0.60c ± 0.08 | 0.36b ± 0.04 |
| Strains Code | Enzymatic Activity (U mL−1) | |||||||
|---|---|---|---|---|---|---|---|---|
| CPM | SPM | |||||||
| 24 h | 48 h | 72 h | 96 h | 24 h | 48 h | 72 h | 96 h | |
| KNU18Y3 | 16.91b ± 0.16 | 15.44b ± 0.06 | 8.56ab ± 0.23 | 8.23a ± 0.10 | 16.83ab ± 0.12 | 15.32b ± 0.40 | 8.41b ± 0.69 | 7.12a ± 0.23 |
| KNU18Y4 | 17.66 a ± 0.13 | 16.25a ± 0.03 | 8.75a ± 0.04 | 8.24a ± 0.12 | 17.55a ± 0.29 | 16.13a ± 0.21 | 8.65a ± 0.23 | 7.14a ± 0.40 |
| KNU18Y5 | 16.16 cd ± 0.12 | 14.68c ± 0.06 | 8.11d ± 0.08 | 7.75b ± 0.05 | 16.12abc ± 0.46 | 14.6cd ± 0.12 | 8.98d ± 0.35 | 7.69bc ± 0.23 |
| KNU18Y6 | 16.42bcd ± 0.39 | 14.34de± 0.07 | 8.47abc ± 0.06 | 7.32c ± 0.08 | 16.31abc ± 0.12 | 14.21de ± 0.29 | 8.34b ± 0.29 | 7.28cde ± 0.40 |
| KNU18Y7 | 16.45bc ± 0.21 | 14.57cd ± 0.12 | 8.21cd ± 0.04 | 7.81b ± 0.09 | 16.23abc ± 0.10 | 14.51cd ± 0.06 | 8.11cd ± 0.49 | 7.76b ± 0.20 |
| KNU18Y9 | 16.20cd ± 0.19 | 14.13e ± 0.07 | 8.38bcd ± 0.09 | 7.41c ± 0.14 | 16.13abc ± 0.29 | 14.09e ± 0.29 | 8.26bc ± 0.15 | 7.31d ± 0.40 |
| KNU18Y12 | 14.93e ± 0.01 | 14.78c ± 0.10 | 8.39bcd ± 0.04 | 7.31c ± 0.18 | 14.86c ± 1.59 | 14.59cd ± 0.06 | 8.21bc ± 0.71 | 7.21e ± 0.12 |
| KNU18Y13 | 15.80d ± 0.23 | 14.78c ± 0.06 | 7.67 e ± 0.03 | 7.78b ± 0.07 | 15.62bc ± 0.23 | 14.66c ± 0.17 | 7.59e ± 0.23 | 7.64c ± 0.13 |
| Strains Code | Enzymatic Activity (U mL−1) | |||||||
|---|---|---|---|---|---|---|---|---|
| CPM | SPM | |||||||
| 24 h | 48 h | 72 h | 96 h | 24 h | 48 h | 72 h | 96 h | |
| KNU18Y3 | 8.01ab ± 0.47 | 7.76b ± 0.12 | 6.08b ± 0.96 | 6.01c ± 0.05 | 7.98b ± 0.35 | 7.71b ± 0.23 | 6.02bcd ± 0.17 | 5.91d ± 0.23 |
| KNU18Y4 | 8.21a ± 0.50 | 8.28a ± 0.07 | 8.01a ± 0.11 | 7.75a ± 0.04 | 8.15a ± 0.29 | 8.21a ± 0.29 | 7.96a ± 0.29 | 7.71a ± 0.35 |
| KNU18Y5 | 7.61abc ± 0.41 | 7.39c ± 0.06 | 6.07b ± 0.08 | 5.33d ± 0.09 | 7.51d ± 0.23 | 7.32d ± 0.23 | 6.01bcd ± 0.40 | 5.23f ± 0.52 |
| KNU18Y6 | 7.98ab ± 0.47 | 7.68b ± 0.05 | 5.97b ± 0.18 | 5.88c ± 0.02 | 7.82c ± 0.46 | 7.61c ± 0.12 | 5.89d ± 0.52 | 5.7e ± 0.10 |
| KNU18Y7 | 6.54cd ± 0.25 | 6.42e ± 0.09 | 6.03b ± 0.19 | 6.16c ± 0.21 | 6.41g ± 0.35 | 6.36g ± 0.20 | 5.94cd ± 0.21 | 6.02c ± 0.23 |
| KNU18Y9 | 6.98bcd ± 0.32 | 6.77d ± 0.10 | 6.22b ± 0.05 | 6.59b ± 0.20 | 6.82e ± 0.29 | 6.67e ± 0.10 | 6.17bc ± 0.13 | 6.48b ± 0.40 |
| KNU18Y12 | 6.71cd ± 0.28 | 6.58de ± 0.09 | 6.02b ± 0.05 | 6.15c ± 0.15 | 6.68f ± 0.23 | 6.49f ± 0.40 | 6.21b ± 0.06 | 6.05c ± 0.23 |
| KNU18Y13 | 6.06d ± 0.13 | 5.94f ± 0.10 | 5.75c ± 0.12 | 5.22d ± 0.14 | 6.03h ± 0.18 | 5.88h ± 0.35 | 5.75d ± 0.27 | 5.32f ± 0.32 |
| Strains CODE | Enzymatic Activity (U mL−1) | |||||||
|---|---|---|---|---|---|---|---|---|
| CPM | SPM | |||||||
| 24 h | 48 h | 72 h | 96 h | 24 h | 48 h | 72 h | 96 h | |
| KNU18Y3 | 15.11c ± 0.47 | 15.68c ± 0.14 | 13.61bcd ± 0.81 | 12.25cd ± 0.92 | 15.05b ± 0.35 | 15.56f ± 0.09 | 13.49bc ± 0.25 | 12.15bc ± 0.13 |
| KNU18Y4 | 15.23c ± 0.21 | 15.96bc ± 0.12 | 13.86bc ± 0.92 | 12.56bc ± 0.13 | 15.07b ± 0.15 | 15.72ef ± 0.12 | 13.71abc ± 0.15 | 12.41b ± 0.10 |
| KNU18Y5 | 15.21c ± 0.29 | 16.34b ± 0.16 | 13.31d ± 0.12 | 12.01d ± 0.18 | 14.98b ± 0.18 | 16.25c ± 0.11 | 13.26c ± 0.12 | 11.98c ± 0.06 |
| KNU18Y6 | 15.32bc ± 0.24 | 16.07bc ± 0.18 | 13.57cd ± 0.10 | 12.38bcd ± 0.16 | 15.17b ± 0.19 | 16.02cde ±0.11 | 13.33c ± 0.07 | 12.19bc ± 0.22 |
| KNU18Y7 | 15.15c ± 0.17 | 16.19bc ± 0.20 | 13.56cd ± 0.11 | 12.26bcd ± 0.06 | 15.02b ± 0.12 | 15.94de ± 0.17 | 13.36c ± 0.16 | 12.12bc ± 0.12 |
| KNU18Y9 | 15.23c± 0.08 | 16.3b ± 0.19 | 13.83bc ± 0.15 | 12.43bc ± 0.08 | 14.98b ± 0.14 | 16.15cd ± 0.08 | 13.68abc ± 0.09 | 12.27bc ± 0.11 |
| KNU18Y12 | 16.53a ± 0.13 | 18.25a ± 0.12 | 14.20a ± 0.09 | 13.00a ± 0.15 | 16.32a ± 0.07 | 18.01a ± 0.08 | 14.06a ± 0.15 | 12.85a ± 0.14 |
| KNU18Y13 | 16.02ab ± 0.11 | 17.86a ± 0.23 | 13.90ab ± 0.08 | 12.64ab ± 0.06 | 15.89a ± 0.16 | 17.63b ± 0.06 | 13.81ab ± 0.13 | 12.42b ± 0.09 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haile, M.; Kang, W.H. Isolation, Identification, and Characterization of Pectinolytic Yeasts for Starter Culture in Coffee Fermentation. Microorganisms 2019, 7, 401. https://doi.org/10.3390/microorganisms7100401
Haile M, Kang WH. Isolation, Identification, and Characterization of Pectinolytic Yeasts for Starter Culture in Coffee Fermentation. Microorganisms. 2019; 7(10):401. https://doi.org/10.3390/microorganisms7100401
Chicago/Turabian StyleHaile, Mesfin, and Won Hee Kang. 2019. "Isolation, Identification, and Characterization of Pectinolytic Yeasts for Starter Culture in Coffee Fermentation" Microorganisms 7, no. 10: 401. https://doi.org/10.3390/microorganisms7100401
APA StyleHaile, M., & Kang, W. H. (2019). Isolation, Identification, and Characterization of Pectinolytic Yeasts for Starter Culture in Coffee Fermentation. Microorganisms, 7(10), 401. https://doi.org/10.3390/microorganisms7100401