Membrane and Cytoplasmic Proteins of Mycobacterium avium subspecies paratuberculosis that Bind to Novel Monoclonal Antibodies


Abstract
:1. Introduction
2. Materials and Methods
2.1. Map Culture and Whole Cell Extract Preparation
2.2. Map-Lambda ZAP Expression Library Construction and Screening
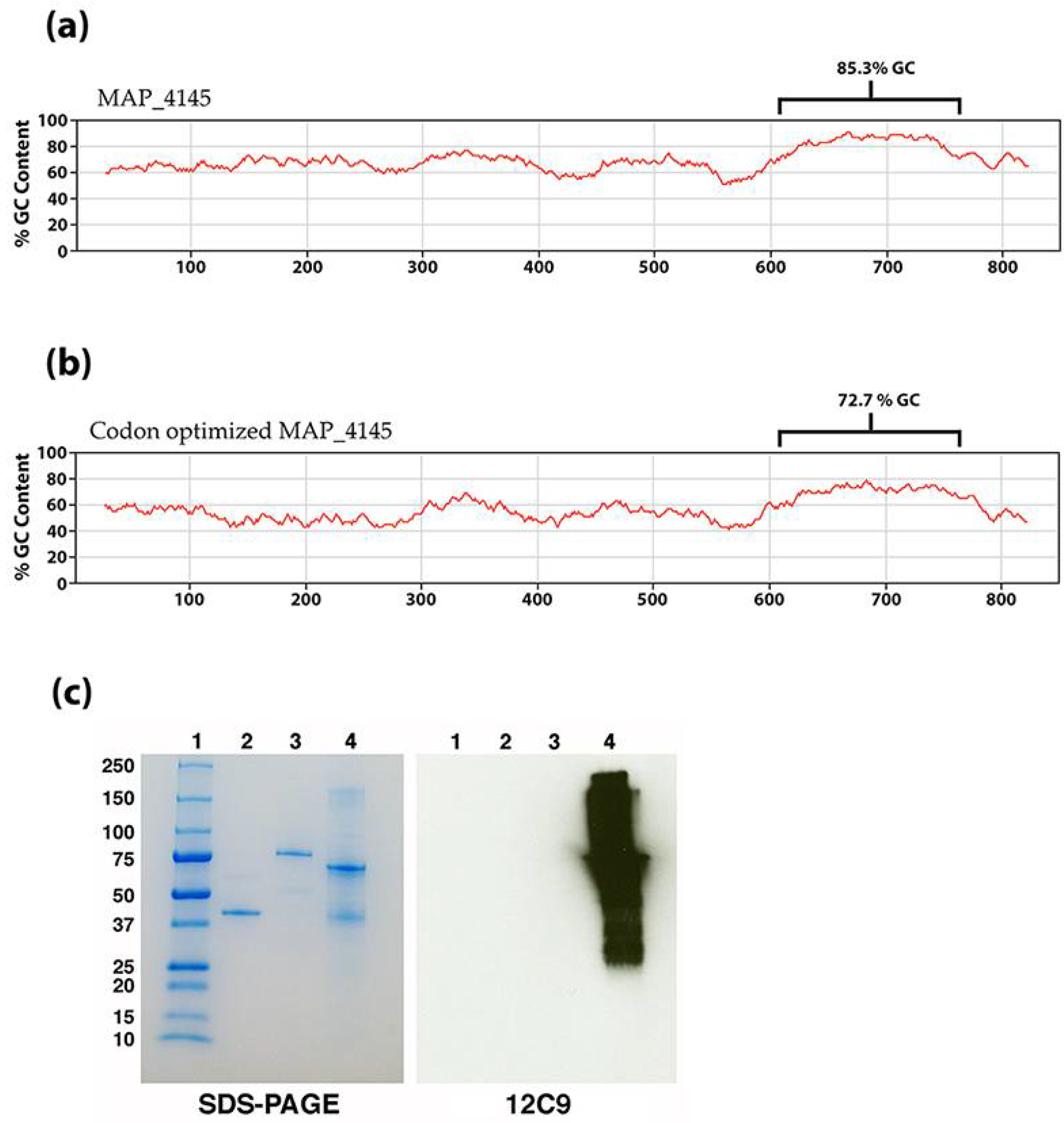
2.3. Cloning and Expression of Selected Map Open Reading Frames
2.4. Immunoblot and Dot Blot Analysis
2.5. ELISA Assay
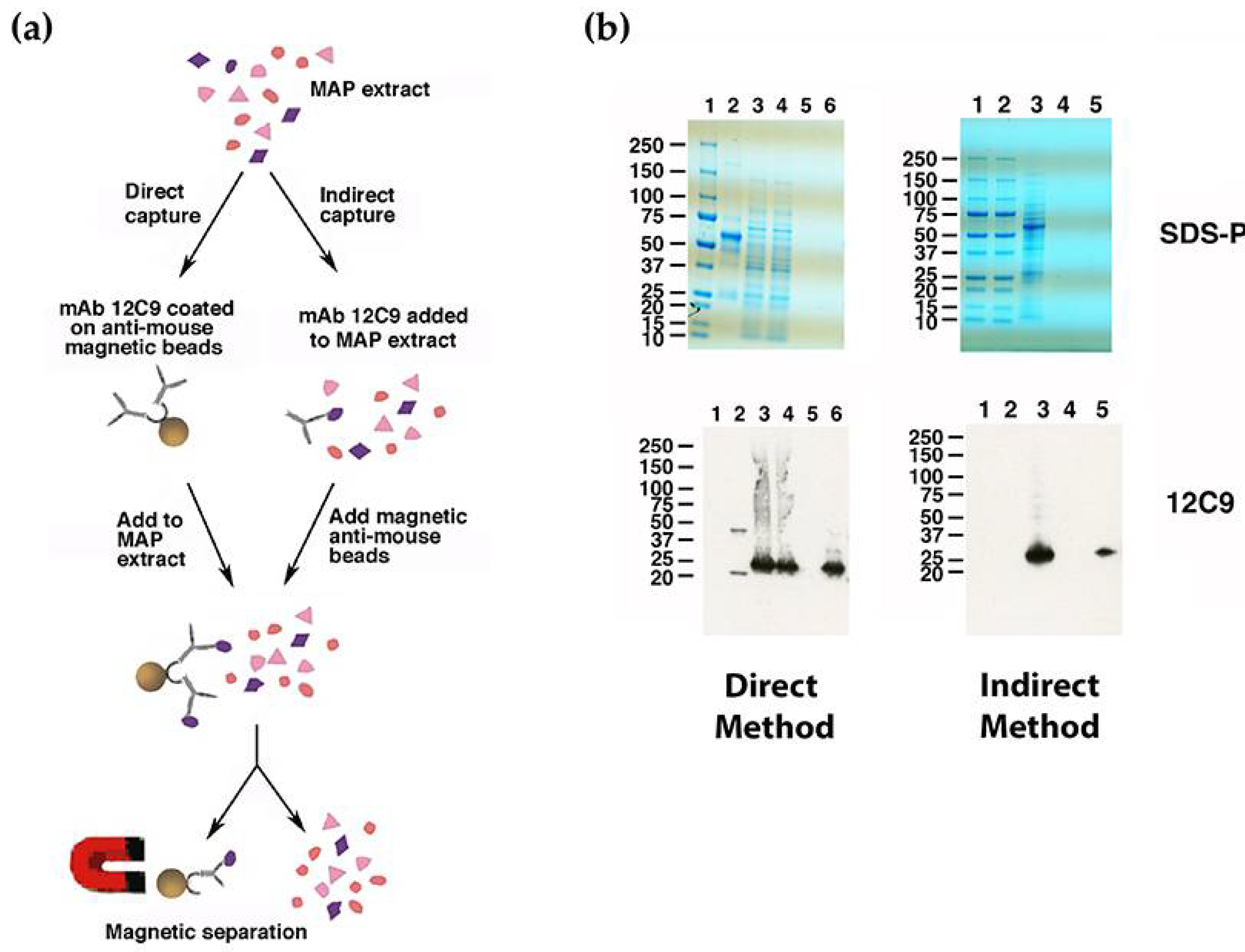
2.6. Antigen Capture and Mass Spectroscopy
2.7. Epitope Mapping Studies
3. Results
3.1. Monoclonal Antibodies to Map Proteins
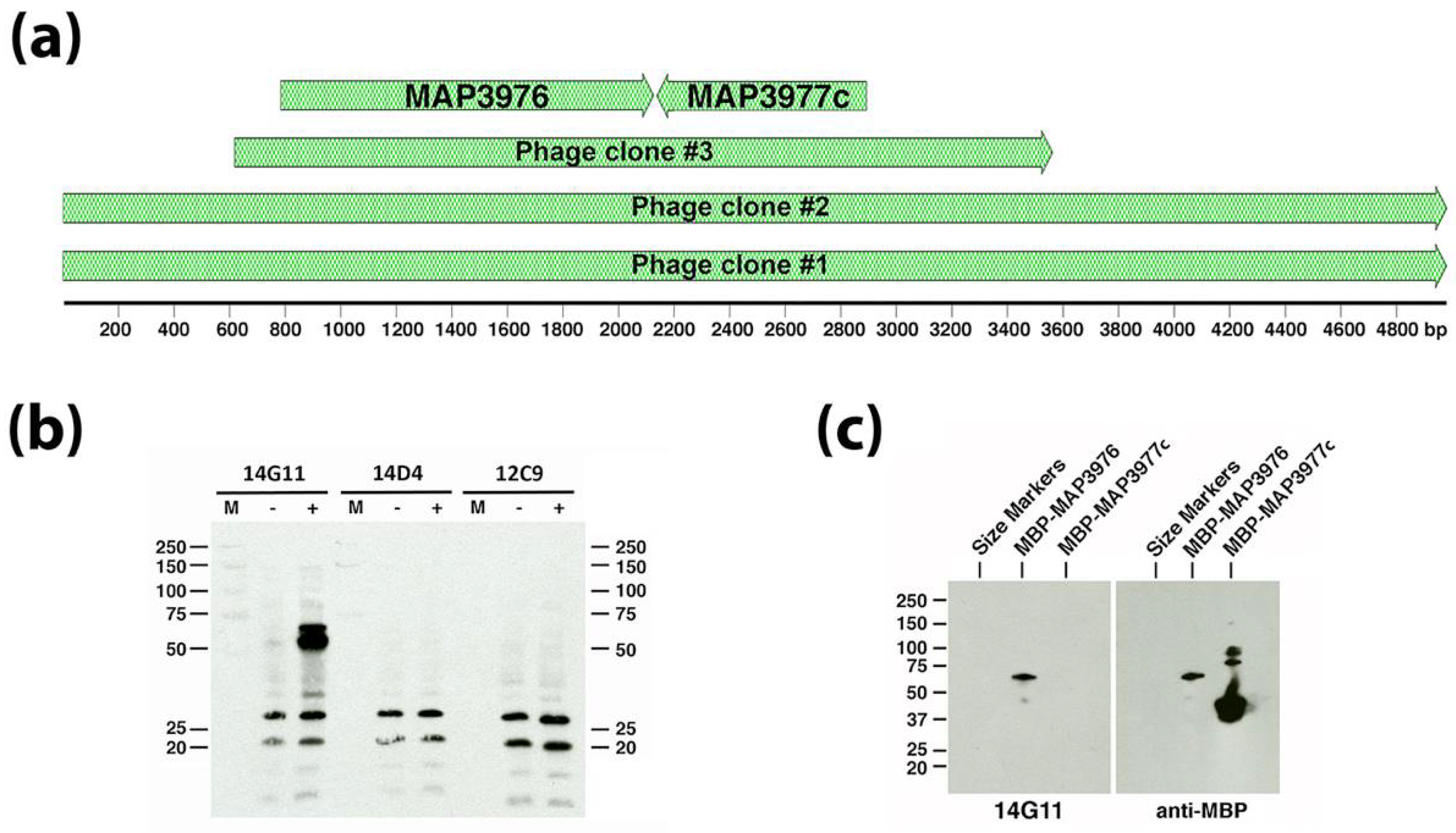
3.2. Monoclonal Antibody Screening Using the Sheep S397 Expression Library
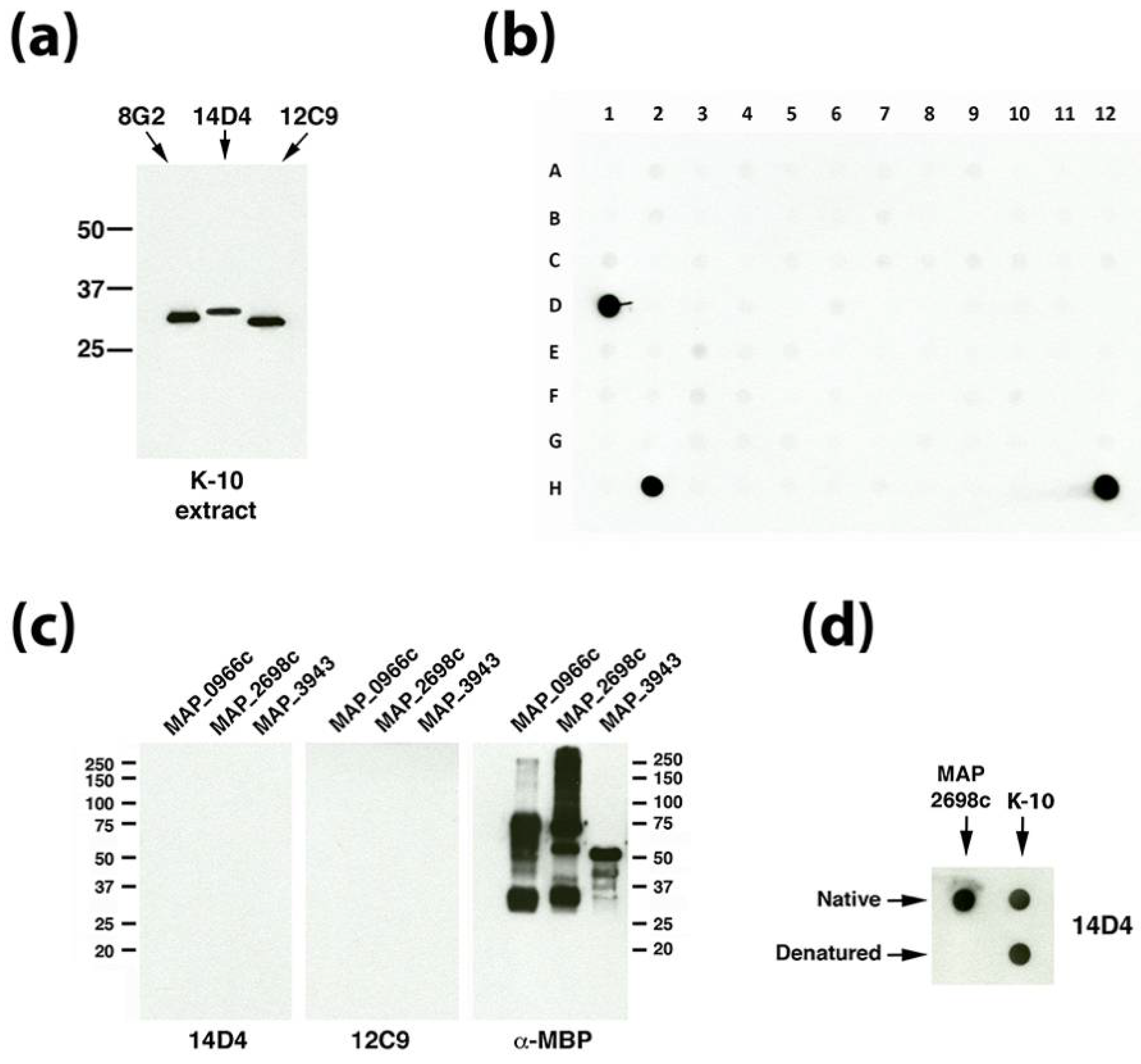
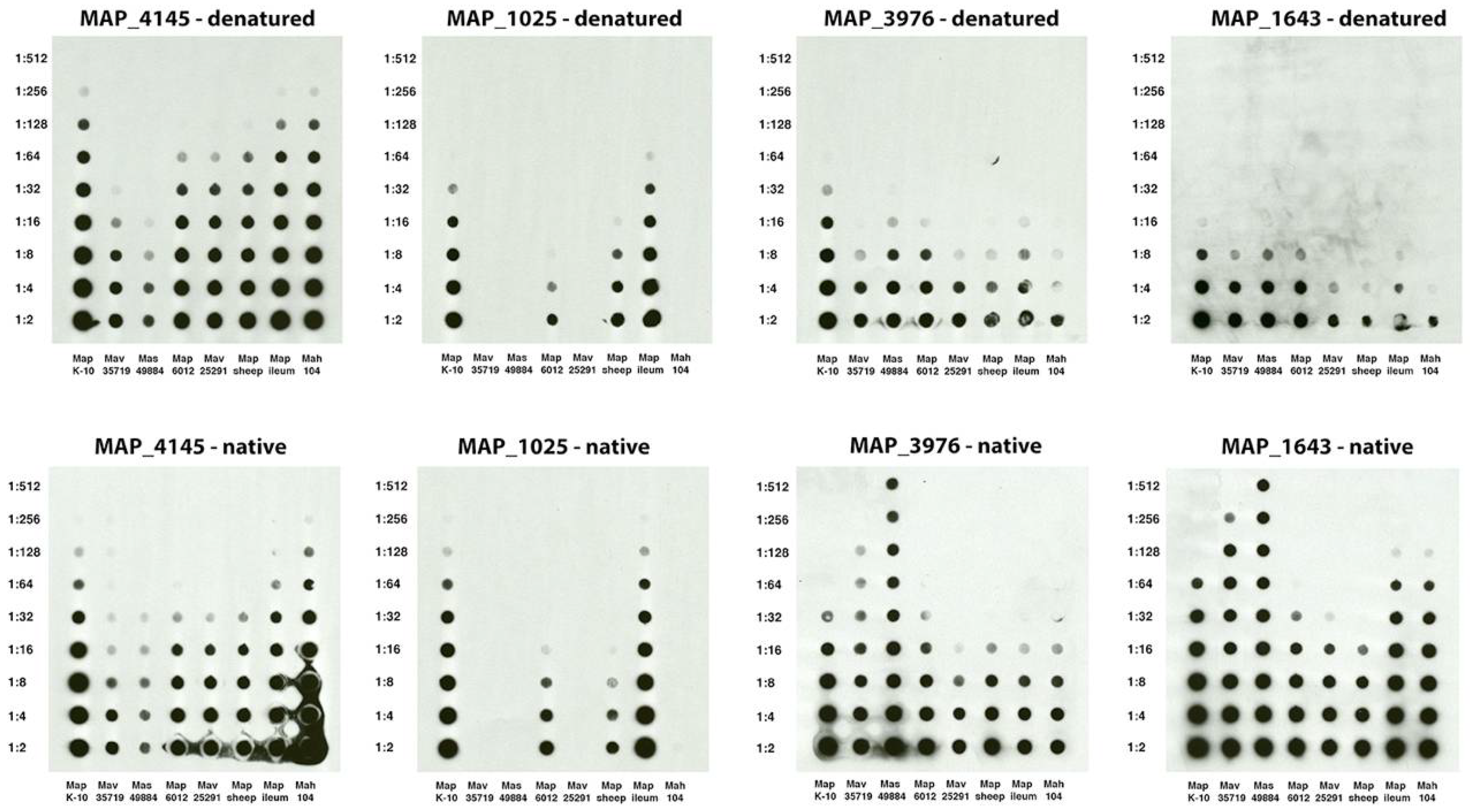
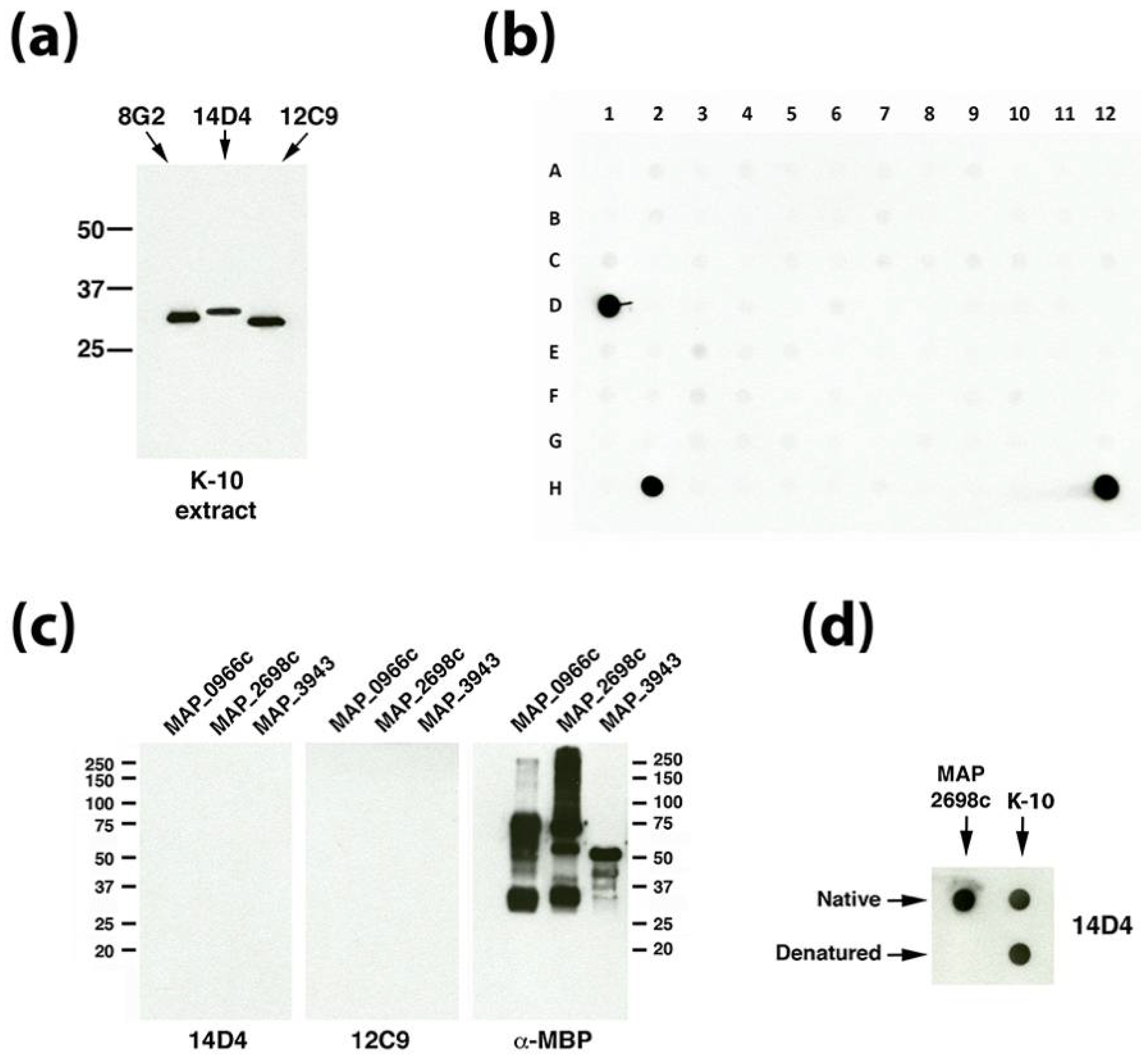
3.3. Dot Blot Array Identifies the Cognate Antigen of mAb 14D4
3.4. Identification of the 12C9 Binding Antigen by Mass Spectrometry
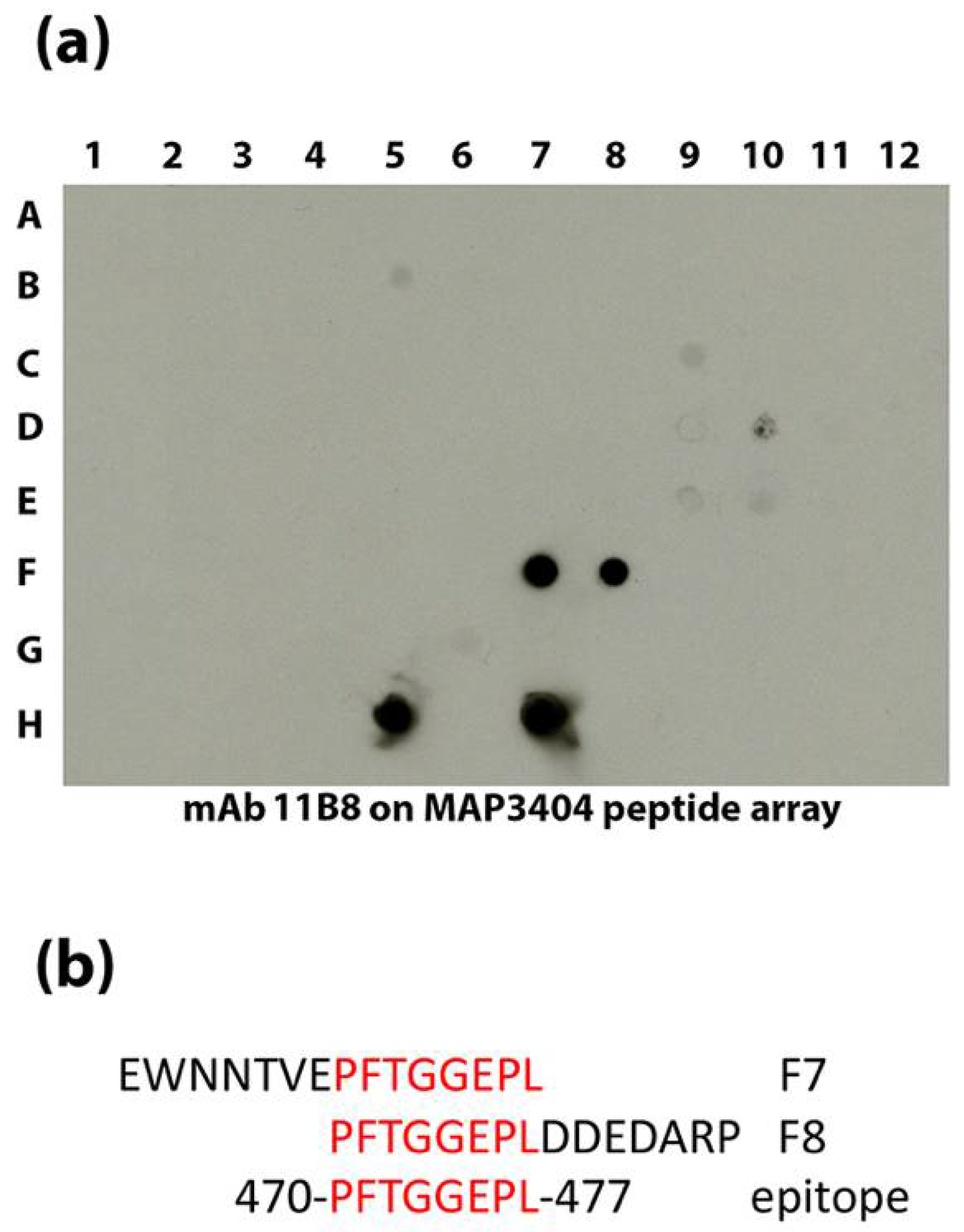
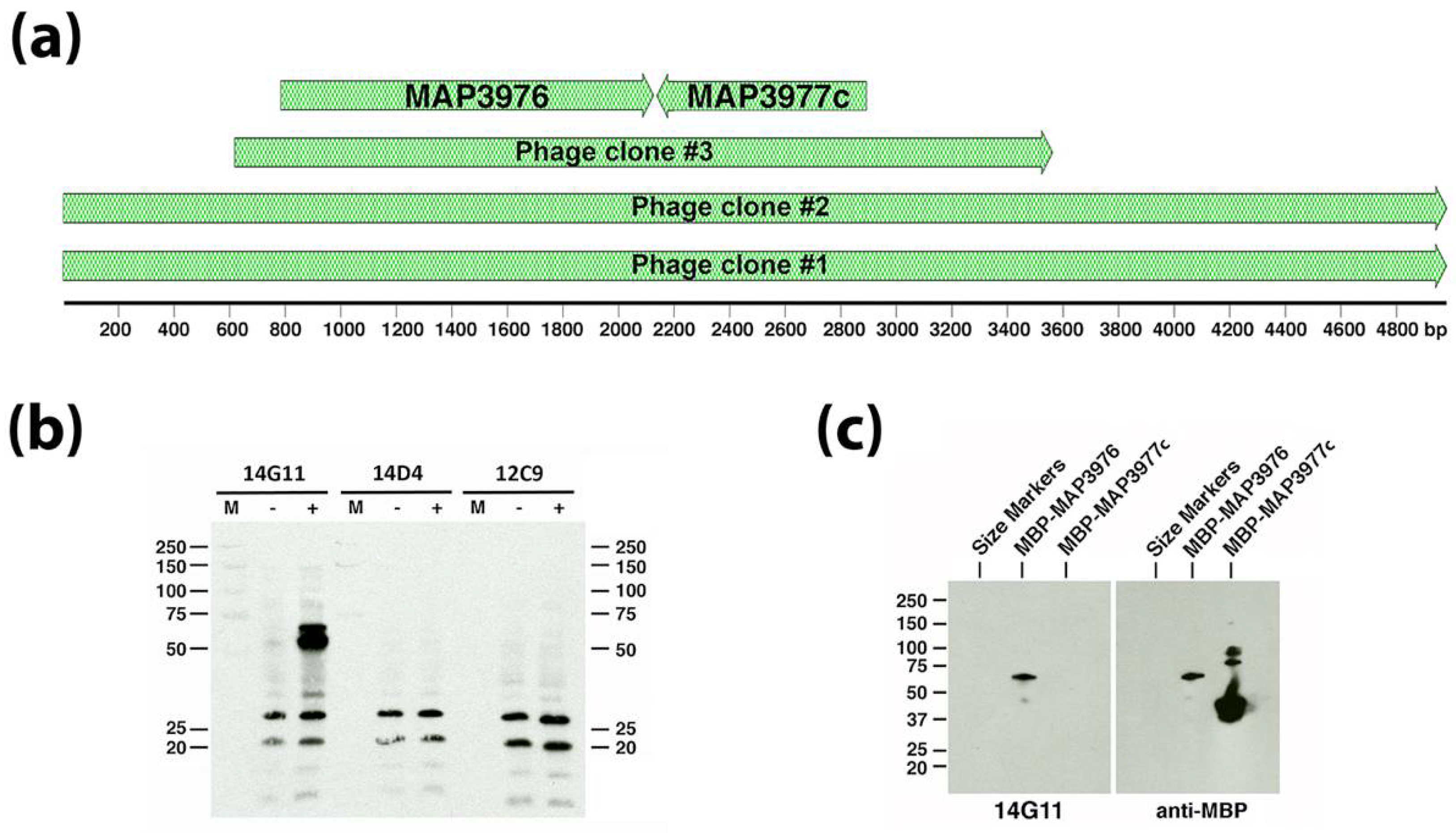
3.5. Epitope Mapping of Selected mAbs to Map Proteins
3.6. Monoclonal Antibody Use in Mycobacterial Proteomics
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Diago-Navarro, E.; Calatayud-Baselga, I.; Sun, D.; Khairallah, C.; Mann, I.; Ulacia-Hernando, A.; Sheridan, B.; Shi, M.; Fries, B.C. Antibody-based immunotherapy to treat and prevent infection with hypervirulent Klebsiella pneumoniae. Clin. Vaccine Immunol. 2017, 24, e00456-16. [Google Scholar] [CrossRef] [PubMed]
- O′Brien, L.M.; Stewart, L.D.; Strain, S.A.; Grant, I.R. Novel monoclonal antibody and peptide binders for Mycobacterium avium subsp. paratuberculosis and their application for magnetic separation. PLoS ONE 2016, 11, e0147870. [Google Scholar]
- Mutharia, L.M.; Moreno, W.; Raymond, M. Analysis of culture filtrate and cell wall-associated antigens of Mycobacterium paratuberculosis with monoclonal antibodies. Infect. Immun. 1997, 65, 387–394. [Google Scholar]
- Bannantine, J.P.; Radosevich, T.J.; Stabel, J.R.; Sreevatsan, S.; Kapur, V.; Paustian, M.L. Development and characterization of monoclonal antibodies and aptamers against major antigens of Mycobacterium avium subsp. paratuberculosis. Clin. Vaccine Immunol. 2007, 14, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Bannantine, J.P.; Stabel, J.R.; Lamont, E.A.; Briggs, R.E.; Sreevatsan, S. Monoclonal antibodies bind a SNP-sensitive epitope that is present uniquely in Mycobacterium avium subspecies paratuberculosis. Front. Microbiol. 2011, 2, 163. [Google Scholar] [CrossRef] [PubMed]
- Bannantine, J.P.; Lingle, C.K.; Stabel, J.R.; Ramyar, K.X.; Garcia, B.L.; Raeber, A.J.; Schacher, P.; Kapur, V.; Geisbrecht, B.V. MAP1272c encodes an NlpC/p60 protein, an antigen detected in cattle with Johne’s disease. Clin. Vaccine Immunol. 2012, 19, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- Bannantine, J.P.; Lingle, C.K.; Adam, P.R.; Ramyar, K.X.; McWhorter, W.J.; Stabel, J.R.; Picking, W.D.; Geisbrecht, B.V. Nlpc/p60 domain-containing proteins of Mycobacterium avium subspecies paratuberculosis that differentially bind and hydrolyze peptidoglycan. Protein Sci. 2016, 25, 840–851. [Google Scholar] [CrossRef] [PubMed]
- Bannantine, J.P.; Radosevich, T.J.; Stabel, J.R.; Berger, S.; Griffin, J.F.; Paustian, M.L. Production and characterization of monoclonal antibodies against a major membrane protein of Mycobacterium avium subsp. paratuberculosis. Clin. Vaccine Immunol. 2007, 14, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Bannantine, J.P.; Huntley, J.F.; Miltner, E.; Stabel, J.R.; Bermudez, L.E. The Mycobacterium avium subsp. paratuberculosis 35 kda protein plays a role in invasion of bovine epithelial cells. Microbiology 2003, 149, 2061–2069. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Li, J.Y.; Wu, J.; Meng, L.; Shou, C.C. Mycoplasma infections and different human carcinomas. World J. Gastroenterol. 2001, 7, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Ning, J.Y.; Sun, G.X.; Huang, S.; Ma, H.; An, P.; Meng, L.; Song, S.M.; Wu, J.; Shou, C.C. Identification of antigens by monoclonal antibody PD4 and its expression in Escherichia coli. World J. Gastroenterol. 2003, 9, 2164–2168. [Google Scholar] [CrossRef] [PubMed]
- Waters, W.R.; Miller, J.M.; Palmer, M.V.; Stabel, J.R.; Jones, D.E.; Koistinen, K.A.; Steadham, E.M.; Hamilton, M.J.; Davis, W.C.; Bannantine, J.P. Early induction of humoral and cellular immune responses during experimental Mycobacterium avium subsp. paratuberculosis infection of calves. Infect. Immun. 2003, 71, 5130–5138. [Google Scholar] [CrossRef] [PubMed]
- Bannantine, J.P.; Stabel, J.R. Identification of two Mycobacterium avium subspecies paratuberculosis gene products differentially recognised by sera from rabbits immunised with live mycobacteria but not heat-killed mycobacteria. J. Med. Microbiol. 2001, 50, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Bannantine, J.P.; Stabel, J.R.; Bayles, D.O.; Geisbrecht, B.V. Characteristics of an extensive Mycobacterium avium subspecies paratuberculosis recombinant protein set. Protein Expr. Purif. 2010, 72, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A., 3rd; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Bannantine, J.P.; Paustian, M.L. Identification of diagnostic proteins in Mycobacterium avium subspecies paratuberculosis by a whole genome analysis approach. Methods Mol. Biol. 2006, 345, 185–196. [Google Scholar] [PubMed]
- Bannantine, J.P.; Paustian, M.L.; Waters, W.R.; Stabel, J.R.; Palmer, M.V.; Li, L.; Kapur, V. Profiling bovine antibody responses to Mycobacterium avium subsp. paratuberculosis infection by using protein arrays. Infect. Immun. 2008, 76, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Capsel, R.T.; Thoen, C.O.; Reinhardt, T.A.; Lippolis, J.D.; Olsen, R.; Stabel, J.R.; Bannantine, J.P. Composition and potency characterization of Mycobacterium avium subsp. paratuberculosis purified protein derivatives. PLoS ONE 2016, 11, e0154685. [Google Scholar] [CrossRef] [PubMed]
- Bannantine, J.P.; Wu, C.W.; Hsu, C.; Zhou, S.; Schwartz, D.C.; Bayles, D.O.; Paustian, M.L.; Alt, D.P.; Sreevatsan, S.; Kapur, V.; et al. Genome sequencing of ovine isolates of Mycobacterium avium subspecies paratuberculosis offers insights into host association. BMC Genom. 2012, 13, 89. [Google Scholar] [CrossRef]
- Li, L.; Bannantine, J.P.; Zhang, Q.; Amonsin, A.; May, B.J.; Alt, D.; Banerji, N.; Kanjilal, S.; Kapur, V. The complete genome sequence of Mycobacterium avium subspecies paratuberculosis. Proc. Natl. Acad. Sci. USA 2005, 102, 12344–12349. [Google Scholar] [CrossRef] [PubMed]
- Karuppusamy, S.; Mutharia, L.; Kelton, D.; Karrow, N.; Kirby, G. Identification of antigenic proteins from Mycobacterium avium subspecies paratuberculosis cell envelope by comparative proteomic analysis. Microbiology 2018, 164, 322–337. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. Psortb 3.0: Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Sigal, G.B.; Pinter, A.; Lowary, T.L.; Kawasaki, M.; Li, A.; Mathew, A.; Tsionsky, M.; Zheng, R.B.; Plisova, T.; Shen, K.; et al. A novel ultra-sensitive immunoassay targeting the MTX-lipoarabinomannan epitope meets the WHO’s performance target for tuberculosis diagnosis. J. Clin. Microbiol. 2018, 56, e01338-18. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.; Baars, L.; Ytterberg, A.J.; Klussmeier, A.; Wagner, C.S.; Nord, O.; Nygren, P.A.; van Wijk, K.J.; de Gier, J.W. Consequences of membrane protein overexpression in Escherichia coli. Mol. Cell. Proteom. 2007, 6, 1527–1550. [Google Scholar] [CrossRef]
- He, Z.; De Buck, J. Localization of proteins in the cell wall of Mycobacterium avium subsp. paratuberculosis K10 by proteomic analysis. Proteome Sci. 2010, 8, 21. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Identified Proteins | Accession Number | Weight | Locus Tag | Total Read Counts | No. of Unique Peptides | ||
|---|---|---|---|---|---|---|---|
| MBP | 12C9 | MBP | 12C9 | ||||
| Uncharacterized protein M. paratuberculosis (strain ATCC BAA-968/K-10) GN=MAP_4145 PE = 4 SV = 1 | Q73SC9_MYCPA | 30 kDa | MAP_4145 | 0 | 24 | 3 | |
| 60 kDa chaperonin 2 M. paratuberculosis (strain ATCC BAA-968/K-10) GN=groL2 PE = 3 SV = 4 | CH602_MYCPA | 57 kDa | MAP_3936 | 10 | 0 | 3 | |
| Uncharacterized oxidoreductase MAP_3007 M. paratuberculosis (strain ATCC BAA-968/K-10) GN=MAP_3007 PE = 3 SV = 1 | Y3007_MYCPA | 30 kDa | MAP_3007 | 12 | 0 | 4 | |
| 35kd_ag M. paratuberculosis (strain ATCC BAA-968/K-10) GN=MAP_2855c PE = 4 SV = 1 | Q73W06_MYCPA | 29 kDa | MAP_2855c | 0 | 4 | 2 | |
| Uncharacterized protein M. paratuberculosis (strain ATCC BAA-968/K-10) GN=MAP_2720c PE = 4 SV = 1 | Q73WE0_MYCPA | 33 kDa | MAP_2720c | 0 | 4 | 2 | |
| Haloalkane dehalogenase M. paratuberculosis (strain ATCC BAA-968/K-10) GN=dhmA PE = 3 SV = 1 | DHMA_MYCPA | 34 kDa | MAP_2057c | 0 | 4 | 2 | |
| Wag31 M. paratuberculosis (strain ATCC BAA-968/K-10) GN=wag31 PE = 4 SV = 1 | Q73YR6_MYCPA | 28 kDa | MAP_1889c | 6 | 0 | 2 | |
| Malate dehydrogenase M. paratuberculosis (strain ATCC BAA-968/K-10) GN=mdh PE = 3 SV = 1 | MDH_MYCPA | 35 kDa | MAP_2541c | 0 | 4 | 2 | |
| Uncharacterized protein M. paratuberculosis (strain ATCC BAA-968/K-10) GN=MAP_0398c PE = 4 SV = 1 | Q744E3_MYCPA | 25 kDa | MAP_0398c | 0 | 4 | 2 | |
| Ornithine carbamoyltransferase M. paratuberculosis (strain ATCC BAA-968/K-10) GN=argF PE = 3 SV = 1 | OTC_MYCPA | 34 kDa | MAP_1365 | 6 | 0 | 3 | |
| Pyrroline-5-carboxylate reductase M. paratuberculosis (strain ATCC BAA-968/K-10) GN=proC PE = 3 SV = 1 | Q73ST2_MYCPA | 30 kDa | MAP_3991 | 4 | 0 | 2 | |
| Hsp90 co-chaperone Cdc37 OS=Bos taurus GN=CDC37 PE = 2 SV = 1 | CDC37_BOVIN (+1) | 45 kDa | 4 | 0 | 2 | ||
| mAb | mAb | Cognate | Antigen | Antigen | Reference | ||
|---|---|---|---|---|---|---|---|
| Name | Isotype a | Antigen | Epitope Location | Antigen Description | Size (kDa) | Location b | |
| 17A12-3B10 c | IgG1 | MAP_1025 | 28-HPGGQQP-34 | Proline rich antigen (Pra); RDD family protein | 25.0 | Cytoplasmic membrane | Bannantine et al. [5] |
| 14C5 d | IgG2a | MAP_1272c | Between amino acids 26 and 74 | NlpC/P60 domain protein; peptidoglycan hydrolase | 29.2 | Cytoplasmic membrane | Bannantine et al. [19] |
| 8G6 d | IgG1 | MAP_1272c | Between amino acids 74 and 128 | NlpC/P60 domain protein; peptidoglycan hydrolase | 29.2 | Cytoplasmic membrane | Bannantine et al. [19] |
| 9G10 | IgG2a | MAP_1643 | C-terminal half of protein | isocitrate lyase, AceAb | 85.2 | Cytoplasmic | Bannantine et al. [4] |
| 11F6 | IgG1 | MAP_1643 | C-terminal half of protein | isocitrate lyase, AceAb | 85.2 | Cytoplasmic | Bannantine et al. [4] |
| 8C6 | ND | MAP_1643 | isocitrate lyase, AceAb | 85.2 | Cytoplasmic | This study e | |
| 10D2 | IgG2b | MAP_1643 | isocitrate lyase, AceAb | 85.2 | Cytoplasmic | This study | |
| 1C8 | ND f | MAP_1643 | isocitrate lyase, AceAb | 85.2 | Cytoplasmic | This study | |
| 13E1-4E3 g | IgG1 | MAP_2121c | Binds only to full length protein | Major membrane protein (MMP) | 33.6 | Cytoplasmic membrane | Bannantine et al. [8] |
| 8G2-2A10 g | IgG1 | MAP_2121c | Between amino acids 78 and 153 | Major membrane protein (MMP) | 33.6 | Cytoplasmic membrane | Bannantine et al. [8] |
| 14G3 | IgG2a | Unknown | ~40 | Bannantine et al. [4] | |||
| 14D4 | IgG3 | MAP_2698c | Conformational epitope f | Fatty acid desaturase | 31.5 | Unknown h | Bannantine et al. [4] |
| 6C9 | IgG1 | MAP_3060c | Electron transfer protein, a-subunit | 32.0 | Cytoplasmic | This study | |
| 3G5 | No react i | MAP_3404 | Acetyl-CoA carboxylase, biotin carboxylase subunit | 64.3 | Cytoplasmic | This study | |
| 7C8 | IgG2a | MAP_3404 | Acetyl-CoA carboxylase, biotin carboxylase subunit | 64.3 | Cytoplasmic | This study | |
| 9H3 | IgG2b | MAP_3404 | 316-TEETAGIDLVLQQFK-330 | Acetyl-CoA carboxylase, biotin carboxylase subunit | 64.3 | Cytoplasmic | This study |
| 12E4 | IgG1 | MAP_3404 | Acetyl-CoA carboxylase, biotin carboxylase subunit | 64.3 | Cytoplasmic | This study | |
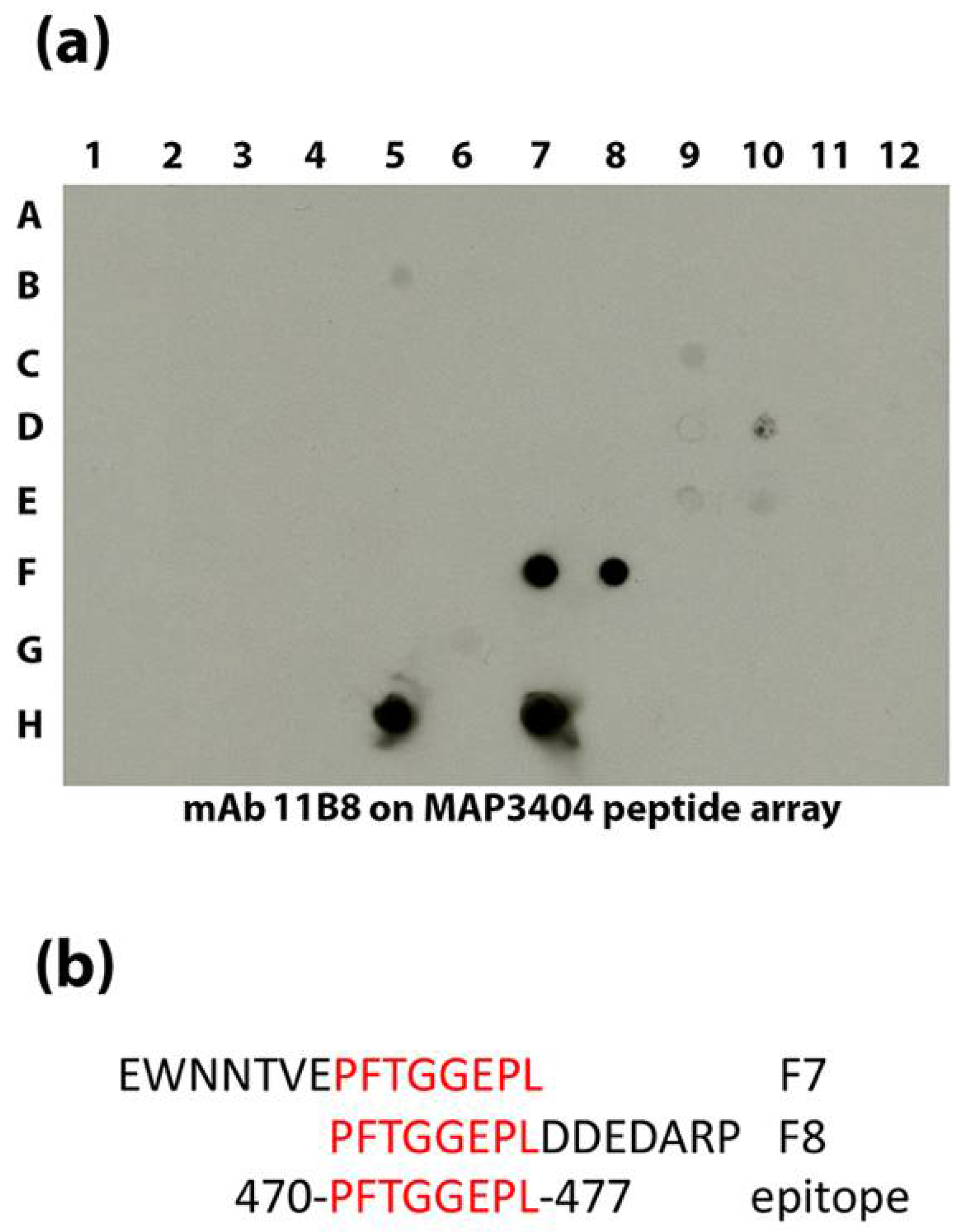
| 11B8 | IgG1 | MAP_3404 | 470-PFTGGEPL-477 | Acetyl-CoA carboxylase, biotin carboxylase subunit | 64.3 | Cytoplasmic | This study |
| 11G4 | IgG1 | MAP_3840 | N-terminal half of protein | DnaK chaperone; Heat shock protein | 66.5 | Cytoplasmic | Bannantine et al. [4] |
| 13A4 | IgG2b | MAP_3840 | Binds only to full length protein | DnaK chaperone; Heat shock protein | 66.5 | Cytoplasmic | Bannantine et al. [4] |
| 7A6 | IgG2a | MAP_3936 | Molecular chaperone GroEL2 | 56.6 | Cytoplasmic | This study | |
| 11F8 | IgG2b | MAP_3936 | Molecular chaperone GroEL2 | 56.6 | Cytoplasmic | This study | |
| 10C12 | IgG1 | MAP_3936 | Molecular chaperone GroEL2 | 56.6 | Cytoplasmic | This study | |
| 14G11 | IgG1 | MAP_3976 | Not determined | Lipoprotein anchoring transpeptidase | 47.0 | Cytoplasmic membrane | Bannantine et al. [4] |
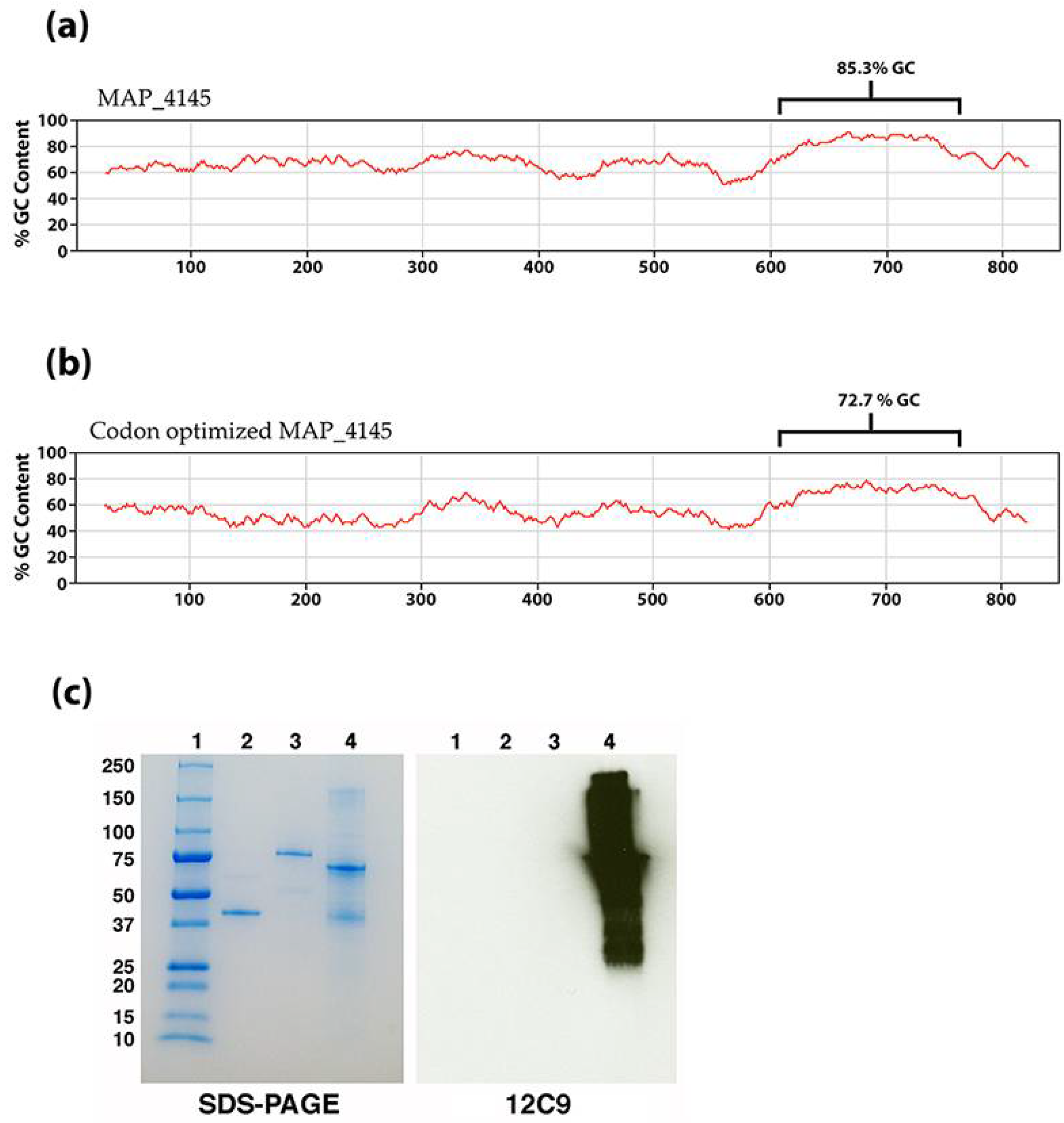
| 12C9 | IgG1 | MAP_4145 | Between amino acids 71 and 212 | Membrane protein with a short C-terminal domain | 30.0 | Cytoplasmic membrane | Bannantine et al. [4] |
| 4B6 | IgG1 | Unknown | Unknown | A highly conserved, unidentified mycobacterial protein | ~42 | Cytoplasmic | Bannantine et al. [4] |
| 12F9 | IgG1 | Unknown | This study | ||||
| 5E6 | IgG1 | Unknown | This study | ||||
| 10D4 | IgG1 | Unknown | This study | ||||
| 11F9 | IgG1 | Unknown | This study | ||||
| 13G7 | ND f | Unknown | This study | ||||
| 2D12 | IgG1 | Unknown | This study | ||||
| 1F2 | IgG2a | Unknown | This study | ||||
| 4A9 | IgG1 | Unknown | This study | ||||
| 4G1 | IgM | Unknown | This study | ||||
| 11F5 | ND | Unknown | Binds to a protein of similar size to MAP_3404 | ~65 | This study |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bannantine, J.P.; Stabel, J.R.; Lippolis, J.D.; Reinhardt, T.A. Membrane and Cytoplasmic Proteins of Mycobacterium avium subspecies paratuberculosis that Bind to Novel Monoclonal Antibodies. Microorganisms 2018, 6, 127. https://doi.org/10.3390/microorganisms6040127
Bannantine JP, Stabel JR, Lippolis JD, Reinhardt TA. Membrane and Cytoplasmic Proteins of Mycobacterium avium subspecies paratuberculosis that Bind to Novel Monoclonal Antibodies. Microorganisms. 2018; 6(4):127. https://doi.org/10.3390/microorganisms6040127
Chicago/Turabian StyleBannantine, John P., Judith R. Stabel, John D. Lippolis, and Timothy A. Reinhardt. 2018. "Membrane and Cytoplasmic Proteins of Mycobacterium avium subspecies paratuberculosis that Bind to Novel Monoclonal Antibodies" Microorganisms 6, no. 4: 127. https://doi.org/10.3390/microorganisms6040127
APA StyleBannantine, J. P., Stabel, J. R., Lippolis, J. D., & Reinhardt, T. A. (2018). Membrane and Cytoplasmic Proteins of Mycobacterium avium subspecies paratuberculosis that Bind to Novel Monoclonal Antibodies. Microorganisms, 6(4), 127. https://doi.org/10.3390/microorganisms6040127