
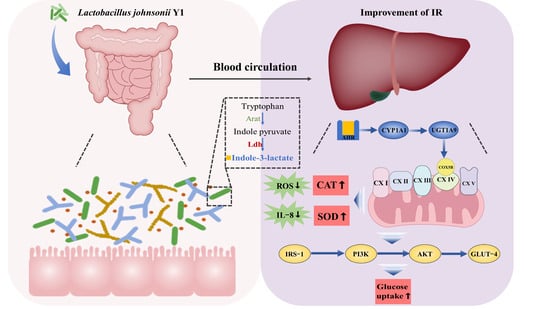
Improvement of Insulin Resistance by Lactobacillus johnsonii-Derived Indole-3-Lactic Acid

 , ,
, ,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Strain and Conditions of Culture
2.3. Animal Experiments
2.4. Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT)
2.5. Assessment of the Insulin-Resistant Index
2.6. 16S rRNA and Serum Metabolomic Sequencing and Data Analysis
2.7. Whole Genome Analysis of L. johnsonii Y1
2.8. Quantitation of ILA and IPA
2.9. Cell Cultures and Treatments
2.9.1. Cell Cytotoxicity Tests
2.9.2. Establishment and Verification of Insulin-Resistant HepG2 Cell (IR-HepG2) Models
2.9.3. Glucose Uptake Assay
2.9.4. Intracellular ROS Levels
2.9.5. Intracellular SOD and CAT Activities Assay
2.10. Extraction of RNA and RT-qPCR Analysis
2.11. RNA Sequencing and Data Analysis
2.12. Statistical Analysis
3. Results
3.1. Effect of L. johnsonii Y1 on Alleviation of IR and Remodeling Gut Microbiota
3.2. Tryptophan Metabolism as the Key Pathway Upon L. johnsonii Y1 Intervention
3.3. Genomic and Functional Basis of ILA Biosynthesis by L. johnsonii Y1
3.4. Effects of ILA on Insulin Sensitivity and Oxidative Stress in HepG2 Cells
3.5. ILA-Mediated Activation of the AHR-COX5B Axis to Mitigate Oxidative Stress
3.6. Restoration of Insulin Signaling via the IRS-1/PI3K/AKT/GLUT-4 Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, M.; Chi, X.; Wang, Y.; Setrerrahmane, S.; Xie, W.; Xu, H. Trends in insulin resistance: Insights into mechanisms and therapeutic strategy. Signal Transduct. Target. Ther. 2022, 7, 216. [Google Scholar] [CrossRef]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef]
- Diaz-Vegas, A.; Sanchez-Aguilera, P.; Krycer, J.R.; Morales, P.E.; Monsalves-Alvarez, M.; Cifuentes, M.; Rothermel, B.A.; Lavandero, S. Is mitochondrial dysfunction a common root of noncommunicable chronic diseases? Endocr. Rev. 2020, 41, bnaa005. [Google Scholar] [CrossRef]
- Zong, Y.; Li, H.; Liao, P.; Chen, L.; Pan, Y.; Zheng, Y.; Zhang, C.; Liu, D.; Zheng, M.; Gao, J.; et al. Mitochondrial dysfunction: Mechanisms and advances in therapy. Signal Transduct. Target. Ther. 2024, 9, 124. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Tsolis, R.M.; Bäumler, A.J. The microbiome and gut homeostasis. Science 2022, 377, eabp9960. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ruan, K.; An, Z.; Li, L.; Ding, C.; Xu, D.; Yang, J.; Zhang, X. Updated review of research on the role of the gut microbiota and microbiota-derived metabolites in acute pancreatitis progression and inflammation-targeted therapy. Int. J. Biol. Sci. 2025, 21, 1242. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.-K.; Kwon, B. Immune regulation through tryptophan metabolism. Exp. Mol. Med. 2023, 55, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Wang, X.; Liu, X. Microbiota-derived metabolites in the epigenetic regulation of the host. Sci. Bull. 2025, 70, 3667–3678. [Google Scholar] [CrossRef]
- Krautkramer, K.A.; Fan, J.; Bäckhed, F. Gut microbial metabolites as multi-kingdom intermediates. Nat. Rev. Microbiol. 2021, 19, 77–94. [Google Scholar] [CrossRef]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef]
- Jia, D.; Wang, Q.; Qi, Y.; Jiang, Y.; He, J.; Lin, Y.; Sun, Y.; Xu, J.; Chen, W.; Fan, L.; et al. Microbial metabolite enhances immunotherapy efficacy by modulating T cell stemness in pan-cancer. Cell 2024, 187, 1651–1665.e21. [Google Scholar] [CrossRef]
- Qian, X.; Li, Q.; Zhu, H.; Chen, Y.; Lin, G.; Zhang, H.; Chen, W.; Wang, G.; Tian, P. Bifidobacteria with indole-3-lactic acid-producing capacity exhibit psychobiotic potential via reducing neuroinflammation. Cell Rep. Med. 2024, 5, 101798. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Y.; Xu, L.; Gu, G.; Dong, Z. Parabacteroides johnsonii inhibits the onset and progression of colorectal cancer by modulating the gut microbiota. J. Transl. Med. 2025, 23, 734. [Google Scholar] [CrossRef]
- Zhang, S.; Sun, Y.; Gao, L.; Zhang, Z.; Yu, Y.; Xu, D.; Nie, S.J.F. Function, Synergistic protective effects of β-glucan and Lactobacillus johnsonii NSP009 in ulcerative colitis. Food Funct. 2025, 16, 8345–8358. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Q.; Ciudad, M.T.; McGaha, T.L. New insights into tryptophan metabolism in cancer. Trends Cancer 2025, 11, 629–641. [Google Scholar] [CrossRef]
- Kim, J.-a.; Wei, Y.; Sowers, J.R. Role of mitochondrial dysfunction in insulin resistance. Circ. Res. 2008, 102, 401–414. [Google Scholar] [CrossRef]
- Geldon, S.; Fernández-Vizarra, E.; Tokatlidis, K. Redox-mediated regulation of mitochondrial biogenesis, dynamics, and respiratory chain assembly in yeast and human cells. Front. Cell Dev. Biol. 2021, 9, 720656. [Google Scholar] [CrossRef] [PubMed]
- Vercellino, I.; Sazanov, L.A. The assembly, regulation and function of the mitochondrial respiratory chain. Nat. Rev. Mol. Cell Biol. 2022, 23, 141–161. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, X.; Cao, K.; Zeng, M.; Fu, X.; Zheng, A.; Zhang, F.; Gao, F.; Zou, X.; Li, H.; et al. Cardiac disruption of SDHAF4-mediated mitochondrial complex II assembly promotes dilated cardiomyopathy. Nat. Commun. 2022, 13, 3947, Erratum in Nat. Commun. 2022, 13, 4690. [Google Scholar] [CrossRef]
- Tramutola, A.; Di Domenico, F.; Perluigi, M.; Barone, E. Biliverdin reductase-A is a key modulator in insulin signaling and metabolism. Trends Endocrinol. Metab. 2025, 37, 466–479. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Ye, L.; Hu, C.; Zhang, Y.; Liu, J.; Shao, L. Periodontitis as a promoting factor of T2D: Current evidence and mechanisms. Int. J. Oral Sci. 2023, 15, 25. [Google Scholar] [CrossRef]
- Gilliland, S.E.; Nelson, C.R.; Maxwell, C. Assimilation of cholesterol by Lactobacillus acidophilus. Appl. Environ. Microbiol. 1985, 49, 377–381. [Google Scholar] [CrossRef]
- GB 14922-2022; Laboratory Animal—Microbiological and Parasitical Standards and Monitoring. State Administration for Market Regulation, Standardization Administration of China: Beijing, China, 2022.
- Dewar, A.E.; Belcher, L.J.; West, S.A. A phylogenetic approach to comparative genomics. Nat. Rev. Genet. 2025, 26, 395–405. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Fei, F.; Ding, L.-L.; Wen, S.-Y.; Ren, C.-F.; Gong, A.-H. Integrated gut microbiome and metabolome analysis reveals the inhibition effect of Lactobacillus plantarum CBT against colorectal cancer. Food Funct. 2024, 15, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Xiao, H.; Lin, C.; Wong, H.L.X.; Lam, Y.Y.; Gong, M.; Wu, G.; Ning, Z.; Huang, C.; Zhang, Y. Gut microbiota-derived tryptamine and phenethylamine impair insulin sensitivity in metabolic syndrome and irritable bowel syndrome. Nat. Commun. 2023, 14, 4986. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhong, X.; Liu, X.; Wang, X.; Gao, X. Therapeutic and improving function of lactobacilli in the prevention and treatment of cardiovascular-related diseases: A novel perspective from gut microbiota. Front. Nutr. 2021, 8, 693412. [Google Scholar] [CrossRef]
- Tian, S.; Lei, Y.; Zhao, F.; Che, J.; Wu, Y.; Lei, P.; Kang, Y.E.; Shan, Y. Improving insulin resistance by sulforaphane via activating the Bacteroides and Lactobacillus SCFAs–GPR–GLP1 signal axis. Food Funct. 2024, 15, 8644–8660. [Google Scholar] [CrossRef]
- Lyu, W.; Chen, L.; Li, D.F.; Li, S.Y.; Dai, Q.; Zhou, H.L.; Liu, Y.Y.; Zhou, J.Y.; Liang, X.J.; Wang, L. Lactobacillus johnsonii-FM1 modulates gut microbiota and secretes anticancer metabolite vanillic acid to inhibit colorectal tumorigenesis. iMetaOmics 2025, 2, e70050. [Google Scholar] [CrossRef]
- Araújo, J.R.; Marques, C.; Rodrigues, C.; Calhau, C.; Faria, A. The metabolic and endocrine impact of diet-derived gut microbiota metabolites on ageing and longevity. Ageing Res. Rev. 2024, 100, 102451. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Yang, X.; Won, K.-J.; Leone, V.A.; Chang, E.B.; Guzman, G.; Ko, Y.; Bae, O.-N.; Lee, H.; Jeong, H. Phenylpropionic acid produced by gut microbiota alleviates acetaminophen-induced hepatotoxicity. Gut Microbes 2023, 15, 2231590. [Google Scholar] [CrossRef]
- Montgomery, T.L.; Eckstrom, K.; Lile, K.H.; Caldwell, S.; Heney, E.R.; Lahue, K.G.; D’Alessandro, A.; Wargo, M.J.; Krementsov, D.N. Lactobacillus reuteri tryptophan metabolism promotes host susceptibility to CNS autoimmunity. Microbiome 2022, 10, 198. [Google Scholar] [CrossRef]
- Boyd, B.M.; James, I.; Johnson, K.P.; Weiss, R.B.; Bush, S.E.; Clayton, D.H.; Dale, C. Stochasticity, determinism, and contingency shape genome evolution of endosymbiotic bacteria. Nat. Commun. 2024, 15, 4571. [Google Scholar] [CrossRef]
- Kuzmin, E.; Taylor, J.S.; Boone, C. Retention of duplicated genes in evolution. Trends Genet. 2022, 38, 59–72, Erratum in Trends Genet. 2022, 38, 883. [Google Scholar] [CrossRef]
- Arnold, B.J.; Huang, I.T.; Hanage, W.P. Horizontal gene transfer and adaptive evolution in bacteria. Nat. Rev. Microbiol. 2022, 20, 206–218. [Google Scholar] [CrossRef]
- Chen, R.; Qian, J.; Wang, Q.; Li, Y.; Xu, Z.; Zhang, M.; Wang, M.; Nie, H.; Yang, W.; Tong, X.; et al. Periodontitis exacerbates metabolic dysfunction-associated steatotic liver disease via the gut microbiota-derived tryptophan metabolism-AHR axis in obesity. eBioMedicine 2025, 122, 106037. [Google Scholar] [CrossRef] [PubMed]
- Chi, Y.-Y.; Xiang, J.-Y.; Li, H.-M.; Shi, H.-Y.; Ning, K.; Shi, C.; Xiang, H.; Xie, Q. Schisandra chinensis polysaccharide prevents alcohol-associated liver disease in mice by modulating the gut microbiota-tryptophan metabolism-AHR pathway axis. Int. J. Biol. Macromol. 2024, 282, 136843. [Google Scholar] [CrossRef]
- Du, H.; Xu, T.; Yu, S.; Wu, S.; Zhang, J. Mitochondrial metabolism and cancer therapeutic innovation. Signal Transduct. Target. Ther. 2025, 10, 245. [Google Scholar] [CrossRef]
- Ježek, P.; Jabůrek, M.; Holendová, B.; Engstová, H.; Dlasková, A. Mitochondrial cristae morphology reflecting metabolism, superoxide formation, redox homeostasis, and pathology. Antioxid. Redox Signal. 2023, 39, 635–683. [Google Scholar] [CrossRef] [PubMed]
- Brzezinski, P.; Moe, A.; Adelroth, P. Structure and mechanism of respiratory III–IV supercomplexes in bioenergetic membranes. Chem. Rev. 2021, 121, 9644–9673. [Google Scholar] [CrossRef]
- Dumas, A.; Knaus, U.G. Raising the ‘Good’ Oxidants for Immune Protection. Front. Immunol. 2021, 12, 698042. [Google Scholar] [CrossRef] [PubMed]
- Vella, V.; De Francesco, E.M.; Bonavita, E.; Lappano, R.; Belfiore, A. IFN-I signaling in cancer: The connection with dysregulated Insulin/IGF axis. Trends Endocrinol. Metab. 2022, 33, 569–586. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.-F.; Liu, C.-J.; Hao, K.-X.; Fan, X.-D.; Jiang, J.-G. Polysaccharides from Flos Sophorae Immaturus ameliorates insulin resistance in IR-HepG2 cells by co-regulating signaling pathways of AMPK and IRS-1/PI3K/AKT. Int. J. Biol. Macromol. 2024, 280, 136088. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Zhan, J.-L.; Wang, Y.-T.; Yu, Y.-S.; Xu, Y.-J.; Wu, J.-J.; Wen, J.; Zou, B.; Wang, H.; Xu, Z.-L.; Wen, P.; et al. Improvement of Insulin Resistance by Lactobacillus johnsonii-Derived Indole-3-Lactic Acid. Microorganisms 2026, 14, 1231. https://doi.org/10.3390/microorganisms14061231
Zhan J-L, Wang Y-T, Yu Y-S, Xu Y-J, Wu J-J, Wen J, Zou B, Wang H, Xu Z-L, Wen P, et al. Improvement of Insulin Resistance by Lactobacillus johnsonii-Derived Indole-3-Lactic Acid. Microorganisms. 2026; 14(6):1231. https://doi.org/10.3390/microorganisms14061231
Chicago/Turabian StyleZhan, Jie-Lin, Ya-Ting Wang, Yuan-Shan Yu, Yu-Juan Xu, Ji-Jun Wu, Jing Wen, Bo Zou, Hong Wang, Zhen-Lin Xu, Peng Wen, and et al. 2026. "Improvement of Insulin Resistance by Lactobacillus johnsonii-Derived Indole-3-Lactic Acid" Microorganisms 14, no. 6: 1231. https://doi.org/10.3390/microorganisms14061231
APA StyleZhan, J.-L., Wang, Y.-T., Yu, Y.-S., Xu, Y.-J., Wu, J.-J., Wen, J., Zou, B., Wang, H., Xu, Z.-L., Wen, P., Hu, T.-G., & Bu, Z.-B. (2026). Improvement of Insulin Resistance by Lactobacillus johnsonii-Derived Indole-3-Lactic Acid. Microorganisms, 14(6), 1231. https://doi.org/10.3390/microorganisms14061231