Abstract
Pseudomonas aeruginosa poses significant health threats due to its multidrug-resistant profile, particularly affecting immunocompromised individuals. The pathogen’s ability to produce virulence factors and antibiotic-resistant biofilms, orchestrated through quorum-sensing (QS) mechanisms, complicates conventional therapeutic interventions. This review aims to critically assess the potential of anti-QS strategies as alternatives to antibiotics against P. aeruginosa infections. Comprehensive literature searches were conducted using databases such as PubMed, Scopus, and Web of Science, focusing on studies addressing QS inhibition strategies published recently. Anti-QS strategies significantly attenuate bacterial virulence by disrupting QS-regulated genes involved in biofilm formation, motility, toxin secretion, and immune evasion. These interventions reduce the selective pressure for resistance and enhance antibiotic efficacy when used in combination therapies. Despite promising outcomes, practical application faces challenges, including specificity of inhibitors, pharmacokinetic limitations, potential cytotoxicity, and bacterial adaptability leading to resistance. Future perspectives should focus on multi-target QS inhibitors, advanced delivery systems, rigorous preclinical validations, and clinical translation frameworks. Addressing current limitations through multidisciplinary research can lead to clinically viable QS-targeted therapies, offering sustainable alternatives to traditional antibiotics and effectively managing antibiotic resistance.
1. Introduction
Pseudomonas aeruginosa is a formidable opportunistic pathogen responsible for severe infections, particularly in immunocompromised individuals, such as those with cystic fibrosis (CF), burns, or immunosuppressive conditions [1]. Its clinical significance stems from its ability to produce diverse virulence factors and form antibiotic-resistant biofilms, rendering conventional therapies ineffective. The rise of multidrug-resistant (MDR) strains due to antibiotic overuse underscores the urgent need for alternative therapeutic strategies [2].
The pathogenicity of P. aeruginosa is mediated by an arsenal of virulence factors, including pyocyanin, pyoverdin, elastase, rhamnolipids, and proteases, which collectively damage host tissues, evade immune responses, and facilitate nutrient acquisition. These factors are tightly regulated by quorum sensing (QS), a cell-density-dependent signaling system that coordinates bacterial behavior [3]. The las and rhl QS systems, governed by the lasI/lasR and rhlI/rhlR genes, regulate biofilm formation, motility, and toxin secretion. QS ensures synchronized expression of virulence determinants, enabling the bacteria to adapt dynamically to host environments [4]. For instance, biofilm formation, a hallmark of chronic infections, is intrinsically linked to QS activation, allowing bacterial communities to adhere to surfaces and resist clearance [5].
P. aeruginosa exhibits intrinsic and acquired resistance to multiple antibiotics, including β-lactams, fluoroquinolones, and aminoglycosides, due to efflux pumps, enzymatic inactivation, and membrane permeability alterations [2]. However, biofilm formation represents a critical resistance mechanism, as extracellular polymeric substances (EPS) in biofilms physically shield bacteria from antibiotics and host immune cells. Biofilms also create metabolic gradients, inducing dormant persister cells that tolerate high antibiotic concentrations [6,7]. Genetic and biofilm—mediated dual resistance strategy renders P. aeruginosa infections recurrent and challenging to eradicate, particularly in hospital-acquired pneumonia, urinary tract infections, and chronic wounds [8].
To circumvent antibiotic resistance, novel strategies targeting virulence and biofilm formation without exerting bactericidal pressure are under investigation [9]. Anti-QS therapies disrupt bacterial communication, attenuating virulence factor production and biofilm development. Natural and synthetic QS inhibitors (QSIs), such as plant-derived compounds and repurposed drugs, have shown promise in downregulating las and rhl systems [10,11]. For example, plant extracts rich in polyphenols and alkaloids inhibit QS-regulated genes, reducing pyocyanin synthesis and biofilm biomass [12]. Similarly, silver nanoparticles synthesized via green chemistry exhibit dual antibacterial and anti-QS activity, destabilizing biofilms through oxidative stress and interference with QS signal molecules [13].
Complementary approaches include bacteriophage therapy, which selectively lyses biofilms while preserving commensal microbiota, and hybrid nanoparticle systems that enhance antibiotic delivery [14]. Hybrid solid lipid nanoparticles (SLNs) co-loaded with antibiotics and natural antimicrobials improve drug solubility and biofilm penetration, synergistically reducing bacterial viability [15]. Marine-derived bioactive compounds, such as actinobacterial metabolites and mollusk-derived peptides, disrupt biofilm architecture and QS signaling pathways [16]. High-throughput screening of FDA-approved drugs has further identified candidates that suppress QS-regulated virulence genes, offering rapid translational potential [17].
The main objective of this review article is to provide a structured overview that begins with a concise examination of the virulence factors of P. aeruginosa and their clinical implications. Subsequently, this review discusses various QSIs, categorized according to their sources, including natural products, synthetic compounds, and bioengineered molecules. Furthermore, the review identifies existing limitations and challenges related to the practical therapeutic application of QSIs. It concludes by outlining future perspectives and suggesting potential research avenues that may facilitate advancements in the development and clinical implementation of QS inhibition strategies. It is important to note that although the selection of QSIs presented herein is extensive, it is not exhaustive; nonetheless, considerable effort has been made to encompass a broad representation of the relevant literature.
2. Virulence Mechanisms of P. aeruginosa
P. aeruginosa is a highly adaptable opportunistic pathogen that utilizes a variety of virulence mechanisms to establish and persist in host environments, particularly in immunocompromised individuals and patients with chronic respiratory diseases such as CF. This bacterium employs diverse strategies, including biofilm formation, QS, motility mechanisms, secretion systems, and the production of a wide range of toxins, collectively contributing to its pathogenicity and resistance to host immune defenses and antimicrobial agents.
2.1. Biofilm Formation and Antibiotic Resistance in P. aeruginosa
A defining virulence feature of P. aeruginosa is its capacity to form biofilms—structured microbial communities encased in a self-produced extracellular matrix. This matrix consists of polysaccharides (e.g., Psl, Pel, and alginate), extracellular DNA (eDNA), proteins, and lipids, which collectively shield bacteria from environmental stressors, host immune defenses, and antimicrobial agents [5,18]. The biofilm’s physical integrity is further reinforced by adhesins such as type IV pili and flagella, which mediate surface attachment, and components like rhamnolipids and amyloid-like fimbriae (ALF) encoded by the fap operon, which enhance stability and resistance to environmental perturbations [19,20,21].
Biofilm-mediated chronic infections are particularly detrimental in CF, where P. aeruginosa adopts a mucoid phenotype characterized by alginate overproduction [22,23]. This phenotype exacerbates respiratory failure by promoting bacterial persistence, impairing mucociliary clearance, and evading antibiotic penetration and host immune responses [24]. Beyond CF, biofilms contribute to the recalcitrance of wound infections, urinary tract infections, and device-associated infections (e.g., ventilators, catheters), where surface-adherent communities resist clearance and prolong host tissue damage [5,25].
Biofilm formation is tightly regulated by QS, a cell-density-dependent signaling system that coordinates the expression of virulence factors, including matrix components and adhesins [18,26]. This regulatory network enhances biofilm maturation and adaptability, further complicating treatment. The combined effects of the EPS matrix, phenotypic diversification, and QS-driven gene expression render biofilm-associated infections extraordinarily resistant to conventional antibiotics [27]. For instance, eDNA within the matrix stabilizes biofilm architecture and binds antimicrobial agents, reducing their efficacy [24]. Consequently, biofilm-related infections often necessitate prolonged, high-dose antibiotic regimens or device removal, underscoring the urgent need for novel therapeutic strategies targeting biofilm dispersal or matrix disruption [28].
2.2. QS and Virulence Regulation in P. aeruginosa
P. aeruginosa employs QS to coordinate virulence factor production and population-wide behaviors. Four interconnected QS systems—Las, Rhl, Pseudomonas quinolone signal (PQS), and integrated QS (Iqs)—govern this regulatory network [24,29,30]. These systems rely on autoinducers, small signaling molecules that synchronize gene expression across bacterial populations, enabling adaptive responses to environmental changes [31]. QS controls the expression of diverse virulence determinants critical for infection (Figure 1): Toxins and Proteases: Exotoxins (ExoS, ExoT, ExoU, ExoY), elastases (LasA, LasB), and proteases contribute to tissue damage, host cell disruption, and immune evasion [31,32].
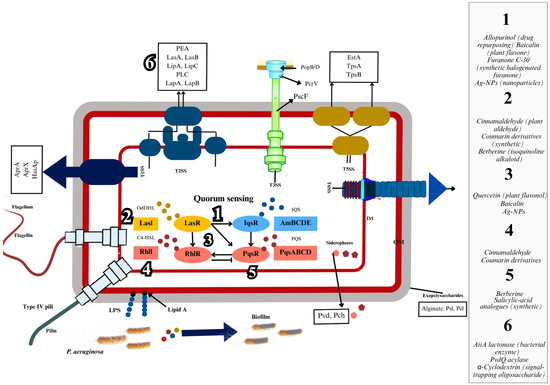
Figure 1.
QS and virulence mechanisms in P. aeruginosa. Legend. This image highlights the various QS systems (LasI/LasR, RhlI/RhlR, IqsR) that regulate the production of virulence factors such as exopolysaccharides (alginate, Psl, Pel), type IV pili, and siderophores. The image also shows the secretion systems (T2SS, T3SS) involved in the export of toxins and other virulence factors, such as PopB/D and PcrV. The interaction between QS signals, such as OdDHL and C4-HSL, leads to the activation of these systems, facilitating the pathogen’s ability to form biofilms and cause infections. This mechanism is central to the pathogenicity of P. aeruginosa, contributing to its persistence and resistance in host environments.
Biofilm Components: QS regulates exopolysaccharides and rhamnolipids, which stabilize biofilm architecture, enhance bacterial persistence, and confer resistance to host defenses [5,33].
Iron Acquisition and Secondary Metabolites: Pyocyanin and pyoverdine, regulated by QS, facilitate iron scavenging and oxidative stress induction, further compromising host immunity [34].
Secretion Systems: The Type VI secretion system (T6SS), linked to the Pseudomonas virulence factor (pvf) gene cluster, mediates host invasion and interbacterial competition [32,35].
The QS network integrates approximately 70 transcription factors (TFs), including Las, Rhl, Pqs, and Iqs, to dynamically adjust virulence gene expression [33]. This hierarchical regulation ensures precise control over biofilm maturation, motility, and toxin secretion. For instance, the pvf cluster synthesizes autoinducers that activate T6SS components and protease production, amplifying pathogenicity [32].
QS-driven adaptations significantly enhance the ability of P. aeruginosa to thrive across diverse ecological niches. Through immune evasion and tissue damage, QS-controlled factors like rhamnolipids and elastases actively degrade host cell membranes, while exotoxins disrupt vital cellular processes [29,31]. Furthermore, antibiotic resistance in P. aeruginosa is bolstered by QS-regulated biofilm formation and efflux pump activity, collectively reducing antimicrobial susceptibility and complicating treatment efforts [24,28]. Lastly, QS signaling pathways facilitate environmental adaptability, allowing rapid transitions between acute virulent states characterized by toxin release and chronic persistence phases through biofilm formation, ensuring survival in fluctuating conditions [19,36].
2.3. Motility Mechanisms and Host Colonization in P. aeruginosa
P. aeruginosa employs diverse motility mechanisms, including flagellar-mediated swimming and type IV pili (T4P)-dependent twitching motility, to colonize host tissues and establish infections [37]. These systems facilitate surface attachment, tissue penetration, and biofilm formation, which are critical for acute and chronic infections [18,38]. Flagella, composed of structural proteins such as FliC (flagellin) and FliD (filament cap), enable swimming motility through liquid environments, allowing the bacterium to navigate toward favorable niches [39]. Conversely, T4P—governed by components including PilA (pilin monomer), PilB (assembly ATPase), PilT, and PilU (retraction ATPases)—mediate twitching motility, a surface-associated movement essential for microcolony formation, biofilm initiation, and host tissue colonization [30,40].
The transition between planktonic and biofilm-associated lifestyles is dynamically regulated by intracellular signaling molecules such as cyclic di-GMP (c-di-GMP) and cAMP. Elevated c-di-GMP levels promote biofilm formation by suppressing motility, while reduced concentrations favor dispersal and motility [41]. Beyond physical movement, these motility structures act as pathogen-associated molecular patterns (PAMPs) recognized by host immune receptors like Toll-like receptor 5 (TLR5), triggering innate immune responses [41,42]. This interaction modulates immune evasion and influences bacterial persistence [43]. Adhesion is further reinforced by lectins LecA and LecB, which bind host glycoconjugates, enhancing biofilm stability and tissue colonization [44].
In clinical settings, particularly in CF airways, P. aeruginosa exploits its motility apparatus to invade tissues and form biofilms, creating protected niches resistant to host defenses and antibiotics [45]. The coordinated action of flagella and T4P enables bacterial aggregation into complex biofilm architectures, hallmarks of chronic infections. This adaptability, governed by environmental cues and signaling pathways, underscores the bacterium’s capacity to thrive in hostile host environments, contributing to its persistence in immunocompromised individuals [18,38].
2.4. Toxins and Enzymes Involved in Tissue Damage
P. aeruginosa secretes an array of potent toxins and enzymes, significantly contributing to its virulence and the pathogenesis of infections through tissue destruction and immune evasion [3]. A prominent virulence mechanism involves the secretion of exotoxins via the Type III secretion system (T3SS), which injects effector proteins directly into host cells. These exotoxins—ExoS, ExoT, ExoU, and ExoY—disrupt host cellular functions and promote immune evasion [46,47]. Specifically, ExoS and ExoT interfere with host cell signaling by modifying actin cytoskeleton dynamics by manipulating small GTPases such as Rho, Rac, and Cdc42. This interaction results in the collapse of actin stress fibers, breakdown of intercellular junctions, loss of tissue integrity, and increased bacterial invasiveness [47,48]. ExoU exhibits phospholipase activity, inducing rapid membrane damage, host cell lysis, and significant lung injury. Strains expressing ExoU are associated with hypervirulence, leading to severe infections and treatment complications [49,50]. ExoY causes the accumulation of cyclic nucleotides, destabilizing the cytoskeletal structure and thus further impairing cellular integrity and function [51].
P. aeruginosa also produces other significant toxins and enzymes through various secretion pathways. Elastases (LasA and LasB), secreted primarily through the Type II secretion system (T2SS), degrade extracellular matrix components and host cell junctions, facilitating bacterial spread within tissues [52,53]. Alkaline protease (AprA), protease IV (PrpL), and phospholipase C actively degrade host proteins, including immunoglobulins and surfactants, leading to extensive tissue damage and promoting immune evasion [54]. Exotoxin A (ExoA), released via T2SS, also significantly contributes to host cell apoptosis and immune disruption [55].
The production of pyocyanin, a phenazine pigment, induces oxidative stress by generating reactive oxygen species (ROS) [34]. These ROS damage host tissues and suppress immune cell function, exacerbating infection severity [46]. Furthermore, hydrogen cyanide (HCN), another toxic metabolite, contributes to oxidative stress and tissue injury [39,56]. Rhamnolipids, surfactant-like virulence factors essential for biofilm formation, play a dual role in biofilm stability and destruction of host tissues. Their presence enhances bacterial resistance to immune responses and antimicrobial agents, compounding infection control challenges [57,58].
Exolysin (ExlA), secreted via the Two-Partner Secretion system (TPS), forms pores in host cell membranes, triggering inflammasome activation, pyroptosis, and tissue disruption via ADAM10-mediated cleavage of cadherins, further compromising cellular integrity [59,60]. Additionally, the Type VI secretion system (T6SS) mediates bacterial competition and modulates host-pathogen interactions, thus influencing infection dynamics and persistence [19,35].
2.5. Immune Evasion Strategies
P. aeruginosa employs diverse strategies to evade host immune responses, enabling persistent infection and resistance to antimicrobial treatment. A central component of its immune evasion involves manipulating lipopolysaccharides (LPS), critical elements of its outer membrane. LPS triggers host inflammatory responses via interactions with receptors such as Toll-like receptor 4 (TLR4); however, modifications in LPS structure help the bacterium evade TLR recognition, reducing immune activation and facilitating persistent colonization [61,62].
Additionally, P. aeruginosa leverages secretion systems, particularly the T3SS and T6SS secretion systems, to deliver virulence factors directly into host cells. These effectors manipulate immune responses, promote bacterial survival, and assist in immune evasion [63]. Toxins such as ExoA disrupt host cell protein synthesis, further impairing immune responses. Elastases (LasA, LasB) and various proteases degrade host immune mediators, weakening the host’s defensive mechanisms and enhancing bacterial dissemination [33,52,60].
Biofilm formation constitutes a crucial immune evasion tactic, protecting bacterial communities within a self-produced matrix. This biofilm matrix shields bacteria from immune cell detection and antibiotic penetration [27]. Alginate production, especially significant in pulmonary infections, additionally prevents recognition by immune cells and enhances persistence within the host environment [64]. Surface-bound factors, such as pili, flagella, and LPS, promote adherence to host tissues, assist initial infection stages, and mask bacteria from immune surveillance, further aiding chronic colonization [65].
To counter host-generated oxidative stress, P. aeruginosa produces reactive ROS-scavenging enzymes, including catalases and peroxidases, regulated by transcription factors SoxR and OxyR. These enzymes neutralize oxidative attacks from immune cells, thus ensuring bacterial survival in hostile environments [66,67].
Efflux pumps, notably MexAB-OprM, significantly contribute to immune evasion by actively expelling antibiotics, enhancing intrinsic resistance, particularly among MDR and extensively drug-resistant (XDR) strains [50,68].
Furthermore, extracellular vesicles (EVs), such as outer membrane vesicles (OMVs), outer-inner membrane vesicles (OIMVs), and explosive outer membrane vesicles (EOMVs), transport virulence factors and antibiotic resistance enzymes (e.g., β-lactamases, aminoglycoside-modifying enzymes), facilitating further immune evasion and antibiotic resistance [69,70,71]. eDNA, released through bacterial autolysis, fortifies biofilm structure, enhancing resistance to immune attacks and antibiotic treatment [72,73].
2.6. Antibiotic Resistance
Antibiotic resistance significantly contributes to the virulence of P. aeruginosa, complicating therapeutic interventions and exacerbating clinical outcomes [74]. This resistance arises from intrinsic and acquired mechanisms, presenting notable challenges in managing infections.
Intrinsic resistance in P. aeruginosa involves several mechanisms, notably the reduced permeability of its outer membrane, modifications of outer membrane proteins (e.g., OprI and OprL), and the active expulsion of antibiotics via efflux pumps such as MexAB-OprM [2,75]. These efflux pumps effectively remove antibiotics and other toxic substances from bacterial cells, elevating the organism’s resistance profile [76].
Biofilm formation significantly enhances antibiotic resistance, complicating therapeutic interventions. Within biofilms, bacterial cells exhibit increased resistance to antibiotic penetration and evade immune detection, making infections more persistent and difficult to treat. This biofilm-associated resistance underscores the clinical challenge of eradicating P. aeruginosa infections, particularly in chronic cases [8,28].
Moreover, P. aeruginosa expresses a variety of antibiotic-degrading enzymes, notably β-lactamases, which neutralize key classes of antibiotics, including penicillins and cephalosporins [77]. The acquisition and horizontal transfer of resistance genes further enhances this resistance, frequently mediated by extracellular vesicles, including OMVs, OIMVs, and EOMVs. These extracellular vesicles facilitate the horizontal transfer of resistance determinants between bacterial cells, significantly accelerating the spread of resistance genes within bacterial populations [78,79].
3. Inhibitors of QS in P. aeruginosa
Strategies to quench QS in P. aeruginosa focus on disrupting key stages of its communication system to reduce pathogenicity. One approach targets the synthesis of AIs through enzymes like lactonases and acylases, which degrade or modify AIs, preventing them from activating QS receptors. Another strategy uses small molecules or structural analogs that bind to QS receptors, blocking AI binding and gene activation. Additionally, antibodies and scavenger proteins can sequester QS molecules. These QSIs can complement antibiotics by preventing biofilm formation and virulence factor production, enhancing bacterial susceptibility to the host immune system, and reducing resistance [80,81]. QSIs, as shown in Figure 1, highlight the diverse strategies targeting specific components of the QS pathway in P. aeruginosa.
3.1. Plant-Derived Natural Inhibitors of QS
Plant-derived compounds employ multifaceted strategies to disrupt P. aeruginosa QS, primarily interfering with acyl-homoserine lactone (AHLs) synthesis, receptor binding, and virulence gene expression (Table 1) [82]. Extracts from Psidium guajava leaves suppress LasR, RhlR, and CviR receptors via beta-caryophyllene and nerolidol, reducing pyocyanin, pyoverdin, and biofilm formation [83]. Similarly, Hypericum perforatum extracts inhibit LasI/R (71.33%) and RhlI/R (57.41%) systems, as demonstrated by diminished Green Fluorescent Protein (GFP) expression in lasB-GFP and rhlA-GFP reporter strains [84]. Molecular docking studies reveal that chlorogenic acid in Prunus avium stalks binds LasR, RhlR, and PqsR, reducing swarming motility (86%) and biofilm formation (75%) [85]. At the same time, Nepeta curviflora methanolic extract attenuates AHL secretion and downregulates biofilm-associated pslA and pelA genes [86]. Polyphenols like caffeoylmalic acid from Salix tetrasperma form hydrogen bonds with LasR and PqsR, impairing protease activity and motility [87].

Table 1.
Plant-based inhibitors of QS in P. aeruginosa.
Direct suppression of QS regulators is a recurring mechanism. Musa acuminata peel extract and its metabolite 5-hydroxymethylfurfural downregulate lasI, lasR, rhlI, and rhlR, reducing biofilm proteins and exopolysaccharides [88]. Green coffee extract at 2.5 mg/mL decreases lasI and lasR expression via chlorogenic acid [89]. Diphysa americana and Hibiscus sabdariffa inhibit swarming and ExoU toxin secretion, correlating with reduced murine pathogenicity [90]. European herbs like Fragaria vesca and Matricaria chamomilla disrupt AI-1/AI-2 signaling, suppressing biofilm and virulence [91], while ginseng (Panax spp.) impairs motility genes, enhancing bacterial clearance [92].
Biofilm and motility disruption are critical for reducing pathogenicity. Syzygium cumini ethyl acetate fraction inhibits biofilm by 86% via RhlG/NADP and LasR interactions [93]. Teucrium polium reduces swarming (23.66%) and swimming (35.25%) without affecting AHL production [94], whereas Persicaria maculosa and Bistorta officinalis abolish swarming at 50 μg/mL by inhibiting LasR [95]. Syzygium aromaticum and Eucalyptus camaldulensis competitively inhibit QS receptors, reducing biofilm [96]. Ginger and wild blueberry extracts downregulate ndvB and pelC by 10.4-fold, dispersing biofilms [97].
Synergy with antibiotics enhances therapeutic efficacy. Fuzheng Touxie Jiedu Huayu decoction reduces ceftazidime minimum inhibitory concentration (MIC) from 128 to 64 μg/mL by inhibiting MexAB-OprM efflux pumps and QS genes (lasI, rhlR) via quercetin and baicalein [98]. Panchvalkal formulation induces nitrosative stress, downregulating norB and nirS to disrupt the biofilm and QS [99] while altering oxidative stress and iron homeostasis genes [100]. LasR, a central QS regulator, is inhibited by boeravinone O (Boerhavia diffusa) and daidzein dimethyl ether (Albizzia lebbeck). Boeravinone O binds LasR’s ligand-binding domain with higher affinity than native autoinducers (3-oxo-C12-HSL/C4-HSL), destabilizing its structure and blocking virulence activation [101]. Similarly, daidzein dimethyl ether suppresses lasR and downstream genes (lasB), reducing biofilm, elastase, and pyocyanin by >50% [102]. Flavonoids like Mosloflavone bind LasR/RhlR, downregulating lasI, lasR, rhlI, and rhlR [103], while wogonin (Agrimonia pilosa) suppresses pqsA, pqsR, and lasB, impairing exopolysaccharides (Psl) and motility [104].
Luteolin modulates host immune responses by reducing TNF-α, IL-1β, and enhancing antioxidant genes (sod, cat) [105]. Quercetin and allicin inhibit biofilm-associated EPS, reducing proteins, carbohydrates, and eDNA [106]. Pinocembrin and vestitol target SagS in MDR strains, reducing biofilm and resistance [107]. Essential oils (EOs) from Cuminum cyminum and Carum carvi inhibit QS via cuminaldehyde and carvone, blocking LasR/CviR [108]. Cinnamomum camphora EO reduces violacein (63%) and biofilm (77.64%) by downregulating cviI, cviR, and vioA-E via linalool and eucalyptol [109]. Sunflower EOs (Agsun cultivars) bind LasR with high affinity (−66.42 kcal/mol), displacing 3OC12-HSL [110]. Zingiber cassumunar EOs synergize with tetracycline (FICI = 0.5), reducing pyocyanin and protease activity [111].
Berberine inhibits PqsA, disrupting alkylquinolone signaling and pyocyanin production [112]. Caffeine mimics 3-oxo-C12-HSL, blocking LasR activation and suppressing swarming [113]. Trigonelline hydrochloride targets Las/Rhl/Pqs systems, altering biofilm architecture [114]. Compound C25 (Plumula nelumbinis) binds LasR/PqsR, silencing QS cascades [115]. Ricinine derivative 7 destabilizes AHL signaling, reducing biofilm [116]. Bakuchiol, a monoterpenoid from Cullen corylifolium, selectively inhibits the pqs system by binding PqsR, destabilizing its structure through interactions with the ALA-168 residue. This reduces pqsA expression by 70%, suppressing pyocyanin (65%), hydrogen cyanide (40%), elastase (55%), lectin (50%), and bacterial motility (>60%) while enhancing survival in Caenorhabditis elegans and Brassica pekinensis models by 80% [117]. Carvacrol exhibits broader anti-QS activity by targeting LasI (−5.932 kcal/mol), LasR (−7.469 kcal/mol), and BswR (−4.42 kcal/mol) via hydrogen bonds at critical residues (Ile107, Tyr47, Leu57), reducing C12-AHL levels by 60% and downregulating lasR by 50% [118,119].
Isoeugenol (400 μM) inhibits QS-regulated pyocyanin, rhamnolipids, exopolysaccharides (Psl), and biofilms (>70%) by binding LasI, LasR, PqsE, and SdiA (−4.8 to −6.4 kcal/mol). At the same time, isovanillin suppresses las and pqs systems at sub-MICs, enhancing antibiotic susceptibility via destabilized QS-mediated resistance [120,121]. Paeonol downregulates lasI/R, rhlI/R, and pqsA/R genes, reduces AHL signaling, and disrupts biofilm architecture [122]. Vanillin specifically inhibits PqsR (−7.3 kcal/mol), suppressing QS signaling and synergizing with colistin to improve G. elegans survival [123], whereas ortho-vanillin blocks RhlR activation by C4-HSL, preserving LasR functionality [124].
Phillyrin (0.25 mg/mL) suppresses pyocyanin, rhamnolipids, and elastase without bactericidal effects, reducing biofilm formation by 84.48% [125]. Sesamin and sesamolin (75 μg/mL) inhibit Las and Rhl systems, downregulating lasI, rhlI, and virulence genes (phzM, rhlA), enhancing C. elegans survival [126]. 6-Gingerol broadly targets LasR, RhlR, QscR, and PqsR, reducing biofilm formation and virulence factors by 40–70% [127]. Curcumin (12.5–50 μM) and baicalein (500 μM) inhibit pyocyanin and motility but lack biofilm efficacy, highlighting variable phenolic QSI potency [128].
Farnesol and tyrosol transcriptionally inhibit lasI and rhlI, disrupting hierarchical QS [129]. Thymoquinone (0.5–2 mg/mL) suppresses Las, Rhl, and QS systems, reducing pyocyanin (~73%), proteases (~70%), and biofilms (~63%) via gene downregulation (lasI/R, pqsA-E/H) and receptor binding (LasR, RhlR) [130,131]. Psoralen binds LasR, RhlR, and PqsR, reducing proteases and motility and enhancing C. elegans survival by 70% [132]. Umbelliferone downregulates rhlR, lasA, and algL, reducing pyocyanin (55%) and biofilm biomass (50%) while quadrupling ciprofloxacin susceptibility [133]. The synthetic coumarin 4t inhibits biofilms (IC50: 3.6 μM) and chelates iron, destabilizing pyoverdine and enhancing tobramycin efficacy [134].
Receptor antagonists like chlorogenic acid, methyl gallate, and falcarindiol competitively bind LasR/RhlR/PqsR, suppressing virulence [102,135,136]. Mangiferin derivatives (Zinc D/E) outperform azithromycin in biofilm inhibition [137]. Transcriptional modulators such as malonate and pyranoanthocyanins shift QS gene expression, while lutein and sennoside A suppress las/rhl/pqs genes [138,139,140,141]. Astragaloside IV and eugenol inhibit efflux pumps, limiting autoinducer release [142].
QS signal degradation strategies include malonate-induced lactonolysis, CS-g-CA-mediated AHL/c-di-GMP reduction, and coumarin-chalcone C9 suppression of 3-oxo-C12-HSL, C4-HSL, and PQS [138,143]. Biofilm disruption via EPS reduction (1,8-cineole), oxidative stress (CS-g-CA), and swarming inhibition (cycloartane triterpenes) further attenuates pathogenicity [144,145].
Synergy with antibiotics is exemplified by DDAG-azithromycin/gentamicin combinations, cinnamoyl hydroxamates enhancing gentamicin dispersal, and compound 2i improving G. mellonella survival with ciprofloxacin/tobramycin [146,147,148]. In vivo efficacy is demonstrated by falcarinol in burned mice and compound 5f in zebrafish [98,149].
3.2. Nanoparticles as Inhibitors of QS
Nanoparticles (NPs) represent a promising class of QSIs capable of disrupting bacterial communication pathways without promoting antimicrobial resistance (Table 2) [150].
Table 2.
Nanoparticle-based inhibitors of QS in P. aeruginosa.
Silver nanoparticles (AgNPs) exhibit multifaceted anti-QS mechanisms, with propolis-synthesized AgNPs reducing AHL-dependent violacein production in Chromobacterium violaceum by 75% through AHL suppression [151]. Multimodal Ag-chitosan-acylase nanoparticles (AgCS@AC) combine acylase I-mediated AHL degradation with biofilm eradication, achieving a 55% reduction in P. aeruginosa biofilms [152]. Eruca sativa-derived AgNPs further demonstrate efficacy by suppressing pyocyanin (68.8%) and exopolysaccharide (57.6%) production via inhibition of the las and rhl systems [153]. Proteomic analyses corroborate these effects, revealing AgNP-induced downregulation of QS genes (lasI, rhlI) and virulence factors (elastase, rhamnolipids) alongside oxidative stress-mediated biofilm destabilization [154]. However, subinhibitory AgNP concentrations paradoxically enhance bacterial motility and biofilm formation in certain strains, emphasizing the critical role of dosage optimization [155].
Zinc oxide nanoparticles (ZnO-NPs) inhibit QS by repressing lasI transcription and AHL synthesis [156]. Nanospiked ZnO structures impair biofilm attachment by suppressing protease and exopolysaccharide production [157]. At the same time, curcumin-loaded ZnO nanocomposites synergize pH-dependent curcumin release and ROS generation to inhibit lasR and rhlR, reducing pyocyanin and rhamnolipid levels [158]. Selenium nanoparticles (SeNPs) exhibit dual mechanisms, docking into LasI and RhlR proteins to block AHL synthesis and biofilm gene expression [159]. SeNPs achieve 80% inhibition of violacein in C. violaceum, whereas tellurium nanoparticles (TeNPs) disrupt signal perception at lower concentrations [160].
Titanium dioxide nanoparticles (TDN) enhance antibiotic efficacy by downregulating lasI, rhlR, and efflux pump genes [161]. Cerium dioxide nanoparticles (CeO2-NPs) mimic haloperoxidases, generating halogenated compounds that reduce biofilms by 85% [162]. Functionalized NPs, such as eugenol-coated gold (Eugenol_Au) and Fe3O4@EUG, suppress virulence factors through ROS generation and QS gene modulation [163,164]. Salmonella enterica co-loaded with eugenol and ofloxacin synergistically disrupts biofilms [15], while alginate-encapsulated meta-bromo-thiolactone (mBTL) inhibits LasR/RhlR, reducing pyocyanin in a dose-dependent manner [165].
Ulusoy et al. demonstrated that Zeolite 4A adsorbs 3-oxo-C12-HSL, reducing P. aeruginosa biofilms (87%) and pyocyanin (98%) [166]. In the same way, Yttrium oxide nanospheres inhibit AHL signaling and initial biofilm adhesion [167], and copper nanoparticles (CuNPs) destabilize biofilms via membrane damage and QS gene repression [168]. In contrast, the metallopharmaceutical Sivanar Amirtham shows ambiguous QS-modulating effects, warranting further mechanistic investigation [169].
3.3. Enzymes, Peptides, and Proteins as Inhibitors of QS
QS-quenching enzymes disrupt P. aeruginosa virulence by degrading AHLs, the signaling molecules essential for QS (Table 3). For example, YtnP, an N-acyl-homoserine lactonase from Bacillus velezensis, hydrolyzes AHLs, significantly reducing EPS synthesis and biofilm formation [170]. Other lactonases, including AiiM, AidB, MomL, and AiiA KMMI17, inactivate AHLs via lactone ring cleavage, suppressing pyocyanin, elastase, and biofilm production without inhibiting bacterial growth [171,172,173]. These enzymes exhibit broad substrate specificity against various AHLs (e.g., C4-HSL, 3OC12-HSL), disrupting the las, rhl, and pqs QS systems. Their efficacy in biofilm control is demonstrated in simulated water systems and infection models, underscoring their therapeutic potential [174,175].
Table 3.
Examples of peptides, enzymes, and proteins inhibiting QS in P. aeruginosa.
QS receptors, such as LasR and RhlR, are direct targets for inhibition. The host defense peptide CRAMP synergizes with colistin to downregulate lasR, lasI, rhlR, and rhlI, reducing pyocyanin (32%) and rhamnolipid (40.5%) synthesis while destabilizing biofilms [176]. Synthetic peptides (WSF, FASK, YDVD) bind the ligand-binding domain of LasR, inhibiting biofilm-associated genes (algC, pslA, pelA) at subinhibitory concentrations [177]. Phage-derived proteins PIT2 and Aqs1 also interfere with LasR: PIT2 disrupts LasR-mediated transcription, suppressing elastase and biofilm formation [178], whereas Aqs1 obstructs LasR-DNA-binding, attenuating pyocyanin and motility [179].
Cyclic dipeptides, such as cyclo(L-Pro-L-Tyr), competitively bind LasR with an affinity comparable to 3OC12-HSL, downregulating lasI and rhlI [180]. Antimicrobial peptides like G3/C8G2 induce conformational changes in LasR, reducing virulence factor expression and biofilm biomass by >95% [180]. Nesfactin, a lipopeptide, combines AHL degradation with receptor binding, while the deep learning-optimized peptide PA-Win2 suppresses las, rhl, and pqs genes, degrading pre-formed biofilms in MDR strains [181,182].
Dual-function agents merge QS inhibition with bactericidal activity. The dendritic peptide G3KL permeabilizes bacterial membranes, causing cytoplasmic leakage, while M59 inhibits the MvfR regulator, reducing pyocyanin [183]. A β-peptide polymer (20:80-Bu: DM) disrupts membranes and suppresses QS genes (las, rhl, pqs), eradicating biofilms and outperforming levofloxacin in murine infection models [184]. These dual mechanisms enhance therapeutic efficacy by simultaneously targeting virulence and viability.
Biofilm matrix disruption indirectly impairs QS. Micafungin, a lipopeptide, reduces alginate and glucan synthesis, destabilizing biofilm architecture and enhancing antibiotic penetration [185]. Recombinant lectin rHPLOE binds biofilm-associated rhamnose, dispersing mature biofilms and suppressing di-rhamnolipid production, a QS-regulated biofilm component [186].
QSI universally attenuates virulence factors, including pyocyanin, elastase, and proteases. For instance, AiiM lactonase reduces elastase activity by 60% [171], while Olivancillaria-derived peptides inhibit pyoverdine by 72% [187]. Biofilm biomass is reduced by >90% using CRAMP/colistin combinations [176] and β-peptide polymers [184]. These effects translate to improved in vivo outcomes: PIT2 enhances survival in C. elegans and HeLa cell infection models [178], while PA-Win2 mitigates pneumonia and catheter-associated infections [182].
3.4. Synthetic and Derived Molecular Inhibitors
Synthetic and derived molecular inhibitors targeting P. aeruginosa employ diverse mechanisms to disrupt QS, efflux pumps, and virulence pathways, offering promising strategies to counter antibiotic resistance (Table 4) [188].
Table 4.
Examples of synthetic and derivative molecules inhibiting QS in P. aeruginosa.
Diclofenac sodium exemplifies dual functionality by suppressing QS and inhibiting efflux pumps via downregulating mexB, mexX, and mexY genes associated with MexAB-OprM and MexXY-OprM systems. However, compensatory upregulation of mexA and oprM is observed. This inhibition enhances gentamicin’s efficacy by restoring antibiotic susceptibility [189,190]. Similarly, ketone- and cyano-selenoesters impair efflux pump activity, improving antimicrobial retention while disrupting QS through interference with autoinducer-1 (AI-1) and autoinducer-2 (AI-2) signaling. Competitive inhibitors like 5-hydroxymethylfurfural (5-HMF) bind LasR and RhlR receptors, key regulators of the Las and Rhl QS circuits, reducing pyocyanin, protease, elastase, and chitinase production by 40–85% and impairing biofilm formation and motility at subinhibitory concentrations [191]. Azithromycin suppresses AHL synthesis, diminishing QS-mediated virulence factors such as pyocyanin and biofilms. At the same time, phenylalanine-arginine β-naphthylamide (PAβN) inhibits efflux pumps involved in QS signal export, albeit with less potency than AZM [192]. High-throughput screening has identified 30 novel PqsR antagonists that competitively bind the PQS site, blocking pqsE activation and disrupting phenazine biosynthesis without affecting bacterial growth [193]. Cephalosporins such as cefepime and ceftazidime inhibit LasR, PqsR, and CviR receptors at sub-MIC levels, reducing pyocyanin, motility, and biofilm formation [194]. Compound Y-31, derived from the AOZ-1 scaffold, interacts with QS regulatory proteins, reducing pyocyanin (22.48%), elastase (22.67%), and biofilms (40.44%). At the same time, lamivudine (LAM) docks into LasR, RhlR, and PqsR, downregulating lasI and pqsR by 82.9% and 91.2%, respectively, and protecting mice from lethal infections [195,196]. Macrolides like erythromycin downregulate lasI, lasR, rhlI, and pqsA by 65.5–81.3%, synergizing with meropenem to enhance growth inhibition [197,198]. Furazolidone reduces lasR, rhlR, and pqsR expression, impairing biofilms, motility, and proteases [199,200]. At the same time, FDA-approved drugs such as erythromycin, chloroquine, and levamisole suppress lasI and rhlI, diminishing virulence in murine sepsis models [201,202].
Efflux pump inhibitors and siderophore disruptors further complement QS-targeted strategies. Nitrofurazone and erythromycin estolate inhibit PqsE, suppressing pyocyanin, rhamnolipids, and biofilms while relieving operon repression to enhance tobramycin efficacy [201]. The siderophore inhibitor HMMN disrupts iron acquisition and QS, reducing siderophores (69.37%), biofilms (28.24%), and pyocyanin (36.06%) via downregulation of lasI, rhlR, mvfR, and pchG [203]. Collectively, these inhibitors attenuate virulence traits, with pyocyanin production reduced by 22.48–36.06%, biofilm formation impaired by 28–45%, and motility disrupted via cephalosporins and furazolidone [194,199]. Protease activity declines by 40% with LAM and furazolidone. Synergies with antibiotics are notable: erythromycin potentiates meropenem, while nitrofurazone enhances tobramycin, suggesting combinatorial therapies to counter resistance [198,201]. In vivo, efficacy is demonstrated by Y-31 extending C. elegans survival, LAM protecting mice, and repurposed FDA drugs improving survival in murine sepsis [195,196,202].
The hierarchical Las, Rhl, and Pqs QS systems are prime targets for antivirulence strategies. Las inhibitors like niclosamide reduce 3OC12-HSL production, though efficacy varies across isolates, while clofoctol competitively targets PqsR to suppress PQS synthesis with broader consistency [204]. Furanone C-30 disrupts LasR folding, impairing QS-regulated gene expression and swarming motility [205], and brominated furanone GBr suppresses pyocyanin, biofilms, and the Type III secretion system (T3SS), attenuating murine infections [206]. N-alkylimidazoles, particularly octylimidazole, inhibit pyoverdine and biofilms via LasR interaction [207], while ibuprofen selectively inhibits Rhl by reducing C4-HSL synthesis [208]. Non-competitive inhibitors MHY1383 and MHY1387 suppress LasR and RhlR at picomolar concentrations, lowering c-di-GMP levels and improving survival in infection models [209]. RhlR antagonists like alkynyl ketone compound 30 block BHL binding, inhibiting rhamnolipids and biofilms [210], and benzimidazolium salts reduce elastase and pyocyanin by up to 64% [85]. Dimetridazole with ribavirin broadly targets LasR, RhlR, and PqsR to downregulate QS genes [211], while furazolidone suppresses lasR, rhlR, and pqsR expression, alleviating murine lung injury [200].
Pqs system inhibitors include thioether-linked cefoperazone Ce-2-ones, which competitively block PqsR to reduce alkylquinolone signaling [212], and quinazolinone derivatives like compound 61, which bind PqsR with nanomolar affinity, suppressing PQS and HHQ production [213]. Thiazole-containing quinazolinones (compounds 18 and 19) inhibit PqsR at sub-300 nM IC50 [214], and quinoline derivative compound 1 competitively antagonizes PqsR, reducing pyocyanin and elastase [215]. Thiazole-based inverse agonists (compounds 27 and 34) suppress PqsR basal activity [216], while multi-target inhibitors like 4-fluorophenyl-5-methylene-2(5H)-furanone 23e inhibit Las, Rhl, and Pqs, enhancing ciprofloxacin activity [217]. Hybrid N-acylcysteines disrupt LasR and biofilms by downregulating pslA and lasI [218] and thiazolo-indolin-2-ones dual-target QS and dihydrofolate reductase [219].
Biofilm-specific strategies include QSI-polymer conjugates (e.g., P2-QSI), which enhance biofilm penetration and ciprofloxacin activity by 70% [220], and L-HSL, which disrupts EPS and motility [221]. 3-Hydroxypyridin-4(1H)-one hybrid inhibits Las and Pqs systems, increasing antibiotic susceptibility [222], while 1,3-diaminopropane suppresses flagellin synthesis and lasR expression [223]. Iron chelation synergizes with QS inhibition; compound 10d binds FpvA, disrupting pyoverdine-mediated iron uptake [224]. Repurposed drugs like cefoperazone inhibit PQS synthesis, synergizing with ciprofloxacin [225], and virtual screening identifies quinazolinones and quinolones as optimized PqsR inhibitors [226]. Resistance to Las-targeting inhibitors is more prevalent than for Pqs-targeting compounds, highlighting the latter’s therapeutic potential [204]. Non-antibiotic approaches, such as efflux pump inducers Cou-1 and Cou-2, reduce T3SS exotoxins via GacS-GacA regulation [227].
Structural insights into inhibitor-receptor interactions have advanced rational design. Ketoprofen and G20 bind PqsR through hydrophobic and hydrogen bonds [228], while benzimidazole 6f engages critical PqsR residues [229]. Halogenated phenazines stabilize LasR’s allosteric site [230], and sulfoxide derivative 6b mediates LasR binding [231]. Repurposed adrenoreceptor blockers, including terazosin and doxazosin, stabilize QS receptors in inactive conformations, reducing biofilm formation and virulence in murine models [127,232]. β-Blockers like propranolol competitively bind LuxR-type receptors, downregulating QS genes [233]. Antidiabetic drugs metformin and vildagliptin synergistically reduce biofilm and virulence factors by binding LasR, RhlR, and PqsR [234], while sitagliptin suppresses motility and QS gene expression [235].
Allopurinol disrupts QS by binding LasR and RhlR, reducing biofilm formation by 61% [236], and cilostazol suppresses QS genes, synergizing with tobramycin [237]. Secnidazole blocks AHL receptor binding [238], and ostarine transcriptionally suppresses autoinducer synthesis [239]. Paracetamol inhibits biofilm formation via AbaR binding [240]. Synthetic derivatives like N-(2- and 3-pyridinyl)benzamides exhibit anti-QS activity [241], and ceftriaxone-silver complexes destabilize biofilms [242,243]. Hydrazide benzoxazole derivatives synergize with ciprofloxacin [244], and chalcone derivatives inhibit MvfR, reducing biofilms and pyoverdine [245]. Natural compounds like trans-cinnamaldehyde suppress lasI and rhlR expression [246], while benzaldehydes enhance antibiotic efficacy against biofilms [247].
Advanced delivery systems, such as nanoparticle-in-liposome platforms, co-deliver PqsR inverse agonists and antibiotics to biofilms, achieving > 90% reduction [248]. Multi-pathway targeting by Sodium New Houttuyfonate (SNH) and itaconimides disrupts QS and virulence [249,250]. Structural interference strategies include phosphate ester derivatives of chrysin and phenyloxadiazole sulfoxides [251,252]. Repurposed drugs like guanfacine block biofilm synthesis [253], and chromone-2-carboxamides exhibit high PqsR-binding affinity [254]. Dual-target inhibitors combining LasR antagonism with nitric oxide synthase inhibition impair virulence and survival in C. elegans [255].
3.5. Microorganisms Producing QSIs
Microorganisms employ diverse QSIs to disrupt P. aeruginosa communication through enzymatic degradation of signaling molecules, receptor antagonism, transcriptional repression, or modulation of secondary metabolic pathways (Table 5) [256].
Table 5.
Some examples of inhibitors produced by microorganisms targeting QS in P. aeruginosa.
A key mechanism involves enzymatic inactivation of AHLs, the primary QS signals in P. aeruginosa. Marine bacteria such as Bacillus paralicheniformis and Vibrio alginolyticus secrete lactonases and acylases that hydrolyze AHLs, suppressing biofilm formation by over 90% [257]. Similarly, Bacillus cereus and Pseudomonas putida degrade AHLs via the aiiA (lactonase) and pvdQ/quiP (acylase) genes, disrupting biofilm formation in MDR strains [258]. Streptomycetes like Streptomyces griseoincarnatus HK12 produce fatty acyl compounds that inhibit LasI, an AHL synthase, preventing signal synthesis entirely [259]. These enzymatic strategies block QS activation while preserving bacterial viability, reducing selective pressure for resistance.
Competitive antagonism of QS receptors represents another primary inhibitory strategy. Macrolactins from Bacillus amyloliquefaciens inhibit the PQS system, suppressing pyocyanin production [260]. At the same time, fungal metabolites such as chermesiterpenoids [261] and cladodionen [262] bind LasR and PqsR with higher affinity than native ligands, reducing elastase and rhamnolipid synthesis. Curvularin selectively antagonizes RhlR, blocking N-butanoyl homoserine lactone (BHL) binding and attenuating virulence in C. elegans infection models [263]. Similarly, the fungal compound tyrosol (P. chrysogenum) binds CviR homologs, suppressing pyocyanin by 63% [264]. These receptor-targeted QSIs prevent signal transduction, rendering P. aeruginosa less pathogenic without directly killing cells.
Transcriptional repression of QS-associated genes further disrupts virulence. Chitosan from shrimp shells reduces lasR and rhlR expression, lowering pyocyanin and biofilm production [265]. Actinomycin D from Streptomyces cyaneochromogenes suppresses lasI, rhlI, and pqsR, impairing AHL synthesis [266]. The NtrBC two-component system, activated by Staphylococcus aureus metabolites, interacts with QS pathways to upregulate pyoverdine and elastase; its deletion abolishes competitive virulence [267]. Gamma irradiation (1 kGy) synergizes with halophilic bacterial metabolites to repress las/rhl genes via oxidative stress, reducing biofilm biomass [268].
QSIs also target secondary signaling systems integral to virulence. Volatile organic compounds (VOCs) from Spongiibacter nanhainus downregulate iron uptake genes (pchR, fpvA), limiting intracellular Fe2+ and QS-dependent metabolism [269]. Chermesiterpenoids reduce c-di-GMP levels, destabilizing biofilm adherence [261], while exopolysaccharide EPS273 degrades eDNA, a critical biofilm matrix component [270]. Biosurfactants from Lactiplantibacillus plantarum, such as oleic acid, structurally mimic AHLs and reduce pyocyanin by 67% [271].
These strategies collectively suppress virulence factors: pyocyanin (23–95% reduction), elastase (35–57%), and rhamnolipids (34–98%) [272,273]. Biofilm formation is inhibited via the downregulation of pslA, algP, and fleQ [270,274], while pre-formed biofilms are dispersed by VOCs [269] and actinobacterial enzymes [275]. Non-bactericidal mechanisms reduce resistance risks; for example, Delftia tsuruhatensis C18-HSL disrupts QS without affecting growth [276].
QSIs enhance conventional antibiotics by sensitizing P. aeruginosa. Chitosan reduces ceftazidime MIC by 15-fold [265], while phthalate derivatives synergize with meropenem [277]. Probiotics like L. plantarum F-10 suppress motility and biofilm formation [278], and bacteriophages complement QSIs by lysing planktonic cells [279]. Marine-derived compounds, including lyngbyoic acid, exhibit low cytotoxicity, underscoring their therapeutic potential [280,281].
3.6. Other Inhibitors
Bromotyrosine derivatives from Aplysinella rhax—psammaplin A and bisaprasin—show strain-specific QS modulation. Bisaprasin broadly suppresses lasB (IC50: 3.53 μM) and rhlA (IC50: 2.41 μM) expression, while psammaplin A is more potent against rhlA (IC50: 4.99 μM vs. 14.02 μM for lasB). Both inhibit elastase production, with psammaplin A achieving more potent suppression (50–100 μM); molecular docking indicates its thiol monomer competitively binds LasR’s ligand-binding domain, disrupting QS. Notably, bisaprasin disrupts biofilm formation at ≥100 μM, unlike psammaplin A [282].
Palmitoleic acid (PMA) targets QS by reducing 3OC12-HSL and C4-HSL, dose-dependently suppressing pyocyanin production and downregulating lasR, rhlR, and rhlI [275]. Its altered membrane permeability lowers the intracellular-to-extracellular 3OC12-HSL ratio, delaying QS activation and reducing biofilm biomass.
Guanosine from Melianthus comosus extracts antagonizes QS by competitively binding LasR and CviR, with docking scores of –8.376 and –5.969 kcal/mol, respectively. It reduces biofilm formation (78.88% at the attachment phase and 34.85% during maturation), suppresses pyocyanin synthesis, and inhibits motility, with a MIC of 0.031 mg/mL [283].
Cyclodextrins, particularly α-CD, inhibit QS by sequestering hydrophobic autoinducers into inclusion complexes. At 10 mM, α-CD reduces pyocyanin by 58% (72 h) and pyoverdine by up to 70% (48 h) without affecting bacterial growth, functioning as a QS quencher and offering a strategy to circumvent antibiotic resistance [284].
3.7. Clustered Regularly Interspaced Short Palindromic Repeats-Based Inhibition of QS
The emergence of Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-based genome-editing technologies has introduced novel strategies to disrupt QS pathways, offering a targeted approach to attenuate bacterial pathogenicity. By leveraging the precision of CRISPR systems to silence or disrupt QS-associated genes, researchers have demonstrated the potential to interfere with bacterial communication, thereby reducing virulence and biofilm-related resilience. This approach is auspicious in MDR pathogens, where traditional antibiotics often fail [285].
In Escherichia coli, CRISPR-Cas9 and CRISPR interference (CRISPRi) systems have successfully targeted QS-related genes such as luxS, fimH, and bolA. For instance, CRISPRi-mediated repression of luxS, a gene essential for autoinducer-2 (AI-2) synthesis, resulted in a 75–84% reduction in biofilm formation, alongside diminished adhesion and virulence [286]. Similarly, in Salmonella enterica, CRISPR-Cas9 targeting of sdiA, a receptor for exogenous AHLs, led to impaired biofilm formation and attenuated virulence in murine infection models [287]. The application of CRISPR-based strategies extends to Vibrio species, where Mobile-CRISPRi systems have been used to silence luxR and luxO, key regulators of bioluminescence and QS. Repression of these genes resulted in significant reductions in light production. It disrupted biofilm architecture, highlighting the potential of CRISPRi to modulate QS-regulated behaviors in environmentally and clinically relevant species [288]. Such findings in diverse bacterial models provide a foundational framework for adapting CRISPR technologies to P. aeruginosa, a notorious MDR pathogen whose QS systems are central to its virulence.
In P. aeruginosa, CRISPR-Cas9 and CRISPR-Cas3 systems have been deployed to target master regulators of QS, including lasR and rhlR. For example, CRISPR-Cas9-mediated disruption of lasR, a transcriptional activator of N-acyl-homoserine lactone (AHL) production, significantly reduced biofilm formation and pyocyanin synthesis, key virulence traits [289]. Similarly, the Type I-F CRISPR-Cas3 system was shown to degrade lasR mRNA, impairing QS signaling and attenuating virulence in both in vitro and murine infection models [290]. Computationally designed guide RNAs targeting the LasR gene, a key QS regulator, and constructed a corresponding vector for gene knockout. Experimental validation demonstrated that disrupting LasR expression significantly reduced QS activity and virulence factor production, aligning with the passage’s assertion that CRISPR systems can be adapted to disrupt bacterial communication networks [291]. These outcomes mirror the success observed in E. coli and Salmonella, where CRISPR-based QS inhibition led to diminished pathogenicity and biofilm resilience.
The convergence of evidence across bacterial species reinforces the viability of CRISPR-based QS inhibition as a therapeutic strategy for P. aeruginosa. The shared mechanistic principles—such as using guide RNAs to target conserved QS regulators and the resultant phenotypic attenuation—suggest that CRISPR systems can be broadly adapted to disrupt bacterial communication networks. For instance, the CRISPRi-mediated silencing of lasR in P. aeruginosa parallels the repression of luxS in E. coli, resulting in disrupted biofilm formation and reduced virulence [286,292]. Furthermore, the adaptability of CRISPR systems, exemplified by the use of dCas9 for reversible gene repression, offers flexibility in modulating QS without permanent genomic alterations, a feature critical for translational applications [293].
3.8. Comparative Analysis of QS Inhibitor Classes
QSIs span diverse chemical and biological classes, each with distinct mechanisms, efficacy profiles, and limitations. Microbial-derived inhibitors, such as macrolactins (e.g., amylomacrolactines A/B) and diketopiperazines (DKPs), disrupt QS by degrading AHLs or mimicking autoinducers. For instance, amylomacrolactines A/B suppress pyocyanin production [260], while DKPs from B. cereus reduce biofilm formation by 68.64% [294]. However, these compounds often exhibit narrow-spectrum activity and face scalability challenges due to complex biosynthesis pathways. In contrast, plant-derived phytochemicals like 6-gingerol and thymoquinone demonstrate broader QS inhibition by targeting multiple receptors (LasR, RhlR, and PqsR). Thymoquinone, for example, reduces pyocyanin by 73% at 2 mg/mL [131], while 6-gingerol suppresses biofilm formation by 42.74% [97]. Despite their potency, poor aqueous solubility and rapid metabolic degradation limit their therapeutic utility, though formulations like chitosan nanoparticles can enhance delivery [295] (Table 6).
Table 6.
Comparative summary of QSI classes.
Synthetic compounds, such as furanone C-30 and phenyloxadiazole sulfoxides, offer high specificity and tunability. Furanone C-30 binds LasR with a ΔG of −54.22 kcal/mol, reducing biofilm biomass by 50% [205], while compound 5b (phenyloxadiazole sulfoxide derivative) inhibits LasB elastase with an IC50 of 8.7 μM [252]. Although these molecules often outperform natural analogs in potency (nanomolar to low micromolar IC50), issues like cytotoxicity (e.g., hemolysis at >100 μM) and resistance emergence hinder clinical translation [296,297]. Peptide-based inhibitors, including CRAMP and PA-Win2, act through dual mechanisms: CRAMP disrupts membrane integrity and downregulates lasR/rhlI genes, achieving 91.05% biofilm reduction at sub-MIC doses [176], while PA-Win2 eliminates biofilms by depolarizing bacterial membranes [182]. However, susceptibility to proteolytic degradation necessitates advanced formulations, such as hydrogels, to prolong stability [298].
Nanoparticles, particularly AgNPs and ZnO-NPs, combine QS inhibition with bactericidal effects. Propolis-synthesized AgNPs reduce violacein production by 75.24% [151], and ZnO-NPs inhibit lasI transcription by 70% [156]. Despite high efficacy (>85% biofilm inhibition), concerns over cytotoxicity and environmental persistence restrict their use [299]. Enzymatic inhibitors, such as AiiA lactonase and MomL, hydrolyze AHLs, reducing pyocyanin by >60% [172,173]. While enzymes provide precise targeting, their instability in vivo demands immobilization strategies (e.g., encapsulation) to maintain activity.
4. Molecular Mechanisms of Plant-Derived Compounds That Antagonize QS Receptors
4.1. Competitive Inhibition of LasR Through Ligand Mimicry
Numerous plant phenolics and terpenoids act as structural analogs of P. aeruginosa’s native autoinducers, competitively occupying the ligand-binding domain (LBD) of the LasR receptor. For example, 6-gingerol, eugenol, and naringenin insert into LasR’s AHL pocket via hydrogen bonding with residues such as Thr75, Asp73, and Tyr56, complemented by hydrophobic interactions with Ala127, Ile52, and Trp88. This steric blockade prevents 3-oxo-C12-HSL binding, as confirmed by molecular docking, GFP reporter assays, and thermal-shift analyses. The resulting inhibition suppresses lasR transcription, downstream QS regulons (lasI, rhlI/R), and virulence phenotypes like pyocyanin, elastase, and biofilm production [300,301]. Similar mechanisms are observed for zingerone, parthenolide, and catechin-7-xyloside, which mimic autoinducer geometry while forming π-stacking or hydrogen-bond networks with conserved LasR residues (e.g., Tyr64, Leu125). These interactions reduce LasR’s affinity for its cognate signal molecule by 10–100-fold, as quantified by microscale thermophoresis and dissociation constants [302,303,304].
4.2. Dual Targeting of QS Receptors and Synthases
Certain compounds disrupt QS by simultaneously binding LasR and LasI synthase, inhibiting signal perception and synthesis. Carvacrol docks into LasR’s LBD (ΔG = −7.47 kcal mol−1) through hydrogen bonds with Tyr47 and hydrophobic stacking, while also occupying LasI’s catalytic site via Zn2+ coordination and hydrogen bonding to Thr104/His399, reducing AHL production by >50%. Similarly, methyl gallate (MG) competitively engages LasR (Trp60, Arg61) and LasI (Arg30, Thr144), with ICM scores of −57.18 and −50.79 kcal mol−1, respectively. This dual interference lowers extracellular 3-oxo-C12-HSL levels and suppresses lasR/I transcription, as demonstrated by β-galactosidase assays and qRT-PCR. Eugenol-conjugated silver nanoparticles (Eu-AgNPs) extend this strategy by targeting LasR, LasI, and MvfR through hydrogen bonding (Tyr56, Ser129) and metal coordination, achieving >80% reductions in violacein, pyocyanin, and biofilm biomass [118,136,305].
4.3. Multi-Receptor Antagonism Across QS Hierarchies
Sesquiterpenoids (α-copaene, β-caryophyllene) and polyphenols (caffeoylmalic acid, epicatechin) exhibit broad-spectrum QS inhibition by targeting LasR, RhlR, and PqsR. Docking simulations reveal α-copaene forms π-alkyl interactions with Val76 (LasR) and Leu57 (CviR), while caffeoylmalic acid binds RhlR (ΔG = −74.39 kcal mol−1) via hydrogen bonds to Arg19/Asp42. These interactions suppress violacein synthesis in C, violaceum, and attenuate P. aeruginosa virulence outputs (e.g., 70% biofilm reduction, 83% pyocyanin decline). Similarly, zingerone and terpinen-4-ol competitively engage LasR, RhlR, and PqsR, reducing C4-AHL and PQS levels while downregulating rhlI/R and pqsR. Such multi-receptor targeting disrupts the interconnected Las-Rhl-PQS signaling network, amplifying antivirulence effects [83,87,306,307].
4.4. Enzymatic Interference with AHL Synthesis
Trans-cinnamaldehyde (CA) and reserpine inhibit LasI synthase, a key enzyme in AHL biosynthesis. CA occupies LasI’s S-adenosylmethionine cavity via π–π interactions with Phe27/Trp33, competitively blocking autoinducer synthesis. Reserpine further obstructs LasI’s catalytic channel through hydrogen bonds to Leu22/Ser109, reducing AHL levels by >70%. These mechanisms, validated by docking and GFP reporter assays, correlate with 102–103-fold repression of lasI and rhlI transcripts and diminished protease/rhamnolipid production [246,308].
4.5. Transcriptional Repression of QS Regulatory Genes
Subinhibitory concentrations of carvacrol, parthenolide, and Acacia nilotica polyphenols downregulate lasR and rhlR transcription by 57–90%, as quantified by qRT-PCR. This repression cascades into reduced expression of QS-controlled genes (lasB, rhlA), lowering elastase, pyocyanin, and biofilm biomass. Phenotypic assays confirm that transcriptional silencing, rather than bactericidal activity, drives virulence attenuation [118,309].
5. Mechanisms of Resistance to QSI in P. aeruginosa
Targeting QS with inhibitors has emerged as a promising antivirulence strategy, aiming to disarm the bacterium without directly killing it, thereby reducing selective pressure for resistance. However, the evolutionary adaptability of P. aeruginosa has led to the rapid development of diverse resistance mechanisms, undermining the efficacy of QSIs and complicating therapeutic outcomes [310]. These mechanisms range from genetic mutations in QS regulatory genes and compensatory activation of alternative signaling pathways to structural biofilm defenses and enzymatic degradation of inhibitors. Notably, the pathogen’s ability to overproduce signaling molecules, upregulate efflux pumps, and modify autoinducer structures further exemplifies its resilience [311]. Such adaptations are particularly problematic in chronic infections, such as those in CF patients, where biofilms and hypoxic microenvironments exacerbate resistance.
5.1. Mutations in QS Regulatory Genes
Genetic mutations in QS regulatory genes are a primary resistance mechanism, enabling P. aeruginosa to evade QSI by altering receptor function or signal transduction. Mutations in key QS regulatory genes, such as lasR, rhlR, and pqsR, are central to resistance against QSIs. LasR mutations, including point mutations, frameshift mutations, insertions, and deletions, frequently localize to the DNA-binding or ligand-binding domains, rendering the protein non-functional or constitutively active [312]. These mutations impair signal molecule binding (e.g., 3OC12-HSL) and transcriptional activation of virulence genes, leading to persistent biofilm formation and virulence factor production despite QSI exposure [313]. Similarly, rhlR and pqsR mutations disrupt ligand recognition (e.g., C4-HSL or PQS), causing signal-independent activation or reduced inhibitor efficacy [314]. Clinical isolates, particularly from chronic infections like CF, often harbor these mutations, confer adaptive advantages in hypoxic biofilm environments by activating alternative regulators like Anr [315]. Phenotypic consequences include sustained pathogenicity, reduced QSI susceptibility, and enhanced biofilm resilience [316].
5.2. Overproduction of QS Signal Molecules
P. aeruginosa bypasses QSI efficacy by overproducing signaling molecules, overwhelming inhibitor competition, and maintaining QS-mediated virulence. P. aeruginosa counteracts QSIs by overproducing signaling molecules such as AHLs and PQS [317]. Upregulation of biosynthetic enzymes (e.g., LasI, RhlI) elevates extracellular signal concentrations, saturating receptor sites, and overwhelming competitive inhibition [314]. Hyper-virulent strains leverage this mechanism to maintain virulence factor expression (e.g., pyocyanin, elastase) and biofilm formation [318]. Chemical analyses and reporter assays confirm elevated AHL levels in resistant strains, enabling persistent QS activity even under inhibitor pressure [319].
5.3. Activation of Alternative QS Pathways
When primary QS systems (e.g., LasI/LasR) are inhibited, P. aeruginosa activates compensatory pathways such as RhlI/RhlR and PqsR [314]. For example, lasR mutants exhibit upregulated rhlR and pqsR expression, enabling continued regulation of virulence genes and biofilm formation [320]. Cross-talk between systems ensures redundancy, allowing bacteria to bypass targeted QS inhibition [321]. Transcriptomic studies reveal pathway-switching dynamics, where inhibition of one system triggers overexpression of another, maintaining pathogenicity [322].
5.4. Efflux Pump Overexpression
Overexpression of efflux pumps, particularly MexAB-OprM, mediates QSI resistance by expelling inhibitors from bacterial cells [323]. Mutations in regulatory genes like mexR derepress pump expression, reducing intracellular QSI concentrations. This mechanism also confers cross-resistance to antibiotics, as MexAB-OprM exports diverse substrates [324]. Biofilm-associated strains further exploit efflux pumps, with hyperactive variants forming denser biofilms due to enhanced AHL export [325].
5.5. Catabolic Degradation of QSIs
Certain strains enzymatically degrade QSIs, such as furanone C30, via lactone ring hydrolysis. This catabolic activity neutralizes inhibitors, reducing their environmental efficacy. Prolonged QSI exposure selects for strains with enhanced degradative enzymes, posing challenges for long-term therapeutic use [326].
5.6. Biofilm-Mediated Resistance
Biofilms confer intrinsic resistance through physical and physiological barriers. The extracellular matrix (polysaccharides, eDNA) limits QSI penetration, while reduced metabolic activity in biofilm cells diminishes inhibitor susceptibility [327]. Biofilms also upregulate efflux pumps and facilitate horizontal gene transfer, accelerating resistance evolution [328].
5.7. Compensatory Activation of Virulence Pathways
QS-independent virulence pathways enable resistance by bypassing QS regulation. Mutants produce virulence factors (e.g., pyocyanin) via alternative signaling cascades, maintaining pathogenicity despite QSI application [329].
5.8. Structural Modifications of Autoinducers
Some strains modify autoinducer structures (e.g., altering AHL acyl chains or lactone rings) to evade QSI binding. These changes prevent inhibitor-receptor interactions, preserving QS signaling and virulence [330].
5.9. Environmental Modulation of QS Hierarchy
Environmental stressors (e.g., nutrient limitation, hypoxia) shift QS system dominance. Phosphate or iron scarcity upregulates RhlR or integrated quorum-sensing systems (iqs), enabling virulence factor production under suboptimal conditions [331].
5.10. Mutations Enhancing Autoinducer Affinity
Rare mutations in LuxR-like receptors increase autoinducer affinity, lowering activation thresholds. These gain-of-function mutations allow QS activation despite low signal concentrations, circumventing competitive inhibition [314].
6. Challenges and Limitations of QSIs
The emergence of antibiotic-resistant P. aeruginosa has driven interest in anti-QS therapies as innovative alternatives to conventional antibiotics. These agents aim to disrupt bacterial communication, attenuating virulence and reducing selective pressure for resistance. However, their clinical translation is hindered by multifaceted challenges spanning efficacy, safety, mechanistic understanding, and practical applicability, necessitating a critical synthesis of these limitations [332].
6.1. High Required Concentrations
The necessity for elevated concentrations to achieve therapeutic efficacy represents a pervasive challenge across multiple classes of QSI, undermining their clinical viability. Synthetic antimicrobial peptides (AMPs), such as WSF, FASK, and YDVD, exemplify this limitation, requiring doses as high as 1600 μg/mL to inhibit P. aeruginosa planktonic growth. These high concentrations raise concerns about cytotoxicity and complicate dosing regimens, as achieving such levels in vivo may exceed safe pharmacokinetic thresholds [177]. Similarly, natural product-derived inhibitors, such as ginger and wild blueberry extracts, demand impractical concentrations (5–25%) to achieve modest biofilm reduction. These levels are far beyond typical pharmacological dosing ranges, posing formulation challenges and risks of systemic toxicity due to the sheer volume or frequency of administration required [97].
Cyclodextrins further illustrate this issue while effectively disrupting QS signaling molecules like AHLs. Their antivirulence activity necessitates prolonged exposure (48–72 h) at 10–12.5 mM concentrations, rendering them unsuitable for acute infections or scenarios requiring rapid therapeutic action. Even synthetic small molecules, such as chromone-2-carboxamide derivatives, require 50 μM to inhibit 90% of biofilm formation—a concentration that may not be achievable in clinical settings due to solubility limitations or toxicity thresholds [254].
6.2. Cytotoxicity and Host Cell Damage
The non-selective activity of several QSI poses significant risks to host cells and tissues, limiting their therapeutic utility. Dendritic peptides, such as G3KL, exemplify this challenge through their membranolytic mechanism. While effectively disrupting bacterial membranes, G3KL indiscriminately targets eukaryotic cell membranes, leading to host cell lysis and tissue damage. This lack of selectivity arises from the peptide’s structural similarity to host defense peptides, compromising its ability to distinguish between pathogen and host cells [183]. Similarly, synthetic peptides like LIVRHK and LIVRRK, though potent QS suppressors, require high doses that exacerbate cytotoxicity, as seen in in vitro models of human epithelial cells [333].
Nanoparticle-based strategies introduce additional risks through oxidative stress. For instance, zinc-copper nanocomposites (ZnC-NCs) and eugenol-conjugated gold nanoparticles (Eugenol_Au NPs) generate ROS as part of their antimicrobial mechanism. While ROSs disrupt bacterial biofilms, they also induce oxidative damage in host tissues, impairing cellular repair mechanisms and exacerbating inflammation in chronic wounds or pulmonary infections [164]. AgNPs, though effective QSI, similarly risk accumulating in vital organs such as the liver and kidneys, where prolonged exposure can lead to organ dysfunction [334].
Natural product-derived inhibitors are not exempt from toxicity concerns. The venom-derived peptide PA-Win2, while effective at disrupting P. aeruginosa biofilm formation, exhibits off-target interactions with human neuronal and cardiovascular cells, necessitating extensive safety evaluations to mitigate risks of neurotoxicity or arrhythmias [182]. Mycotoxin-based compounds, such as citrinin, further illustrate this issue; even at subinhibitory concentrations, they disrupt host mitochondrial function, highlighting the delicate balance between antimicrobial activity and biocompatibility [272]. These examples underscore the critical need for selectivity profiling and structural modifications to minimize host-system collateral damage while retaining antivirulence efficacy.
6.3. Resistance Risks
The non-lethal nature of QS inhibition creates a fertile ground for bacterial adaptation, posing a significant threat to the long-term efficacy of antivirulence therapies. Subinhibitory concentrations of inhibitors, while sufficient to disrupt QS-mediated virulence, often fail to eradicate bacterial populations, enabling P. aeruginosa to develop phenotypic tolerance or genetic resistance [317]. For example, subtherapeutic doses of synthetic peptides like G3 and C8G2 exert selective pressure that favors the emergence of QS-deficient mutants or hypermutable strains, which bypass QS regulation to sustain virulence through alternative pathways [180]. Similarly, plant-derived extracts such as N. curviflora exhibit dose-dependent QS suppression. Still, at low concentrations, they inadvertently promote bacterial efflux pump activation or biofilm matrix remodeling, enhancing resilience against subsequent treatments [86].
Nanoparticles administered at sub-MICs further exacerbate resistance risks. Bio-synthesized AgNPs, while effectively reducing QS signal molecules, enhance bacterial motility and upregulate efflux systems like MexAB-OprM when used at sub-lethal doses. This adaptive response compromises the efficacy of AgNPs and may confer cross-resistance to conventional antibiotics [155]. Additionally, QSI-targeting single pathways, such as PqsR antagonists, leave Las and Rhl systems intact. Residual bacteria exploiting these redundant QS networks can regenerate virulent subpopulations, as observed in P. aeruginosa biofilms treated with high-throughput-screened PqsR inhibitors [193].
The persistence of resistant mutants is further amplified in chronic infections, where heterogeneous biofilms provide sanctuaries for QS-independent subpopulations. Enzymatic inhibitors like AHL-lactonases, which degrade signal molecules without killing bacteria, may inadvertently select for “cheater” strains that exploit public goods (e.g., proteases, toxins) produced by QS-active neighbors, perpetuating infection [171]. Combinatorial approaches—pairing QSI with antibiotics or phage therapy—are critical to mitigate these risks. For instance, coupling PqsR inhibitors with bactericidal agents like ciprofloxacin reduces bacterial load while suppressing virulence, minimizing opportunities for resistance evolution [244].
6.4. Pharmacokinetic and Formulation Challenges
The successful translation of QSI from bench to bedside is significantly hindered by instability, poor bioavailability, and complex delivery requirements. Peptide-based inhibitors, such as Nesfactin, exemplify these hurdles. While effective in vitro, Nesfactin degrades rapidly in physiological environments due to protease activity, necessitating hydrogel-based formulations to stabilize its structure and prolong its half-life. However, hydrogels introduce challenges, including variable drug release kinetics and limited tissue penetration in dense biofilms, which compromise therapeutic consistency [181]. Similarly, enzymatic inhibitors like MomL lactonase face delivery obstacles; their production relies on heterologous expression in Bacillus brevis, requiring purification and encapsulation for therapeutic use. This multi-step process escalates production costs and complicates direct administration, particularly systemic infections [335].
Nanoparticle formulations, though promising, grapple with instability in biological environments. ZnC-NCs, designed for pH-dependent release in acidic infection sites, often underperform in vivo due to physiological pH variability. For instance, in chronic wound models, fluctuating pH levels disrupt the controlled release of QSI, leading to subtherapeutic drug concentrations and inconsistent biofilm suppression [158]. Similarly, SLNs co-loaded with antibiotics and QSI face manufacturing complexities. While enhancing drug solubility, the lipid matrix is prone to destabilization during scale-up, resulting in batch variability. Chitosan coatings that improve mucosal adhesion further risk triggering immunogenic responses, limiting their use in immunocompromised patients [15].
Enzymatic QSI, such as AHL-lactonases from marine actinobacteria, suffer from poor in vivo stability. These enzymes are highly susceptible to proteolytic degradation in host tissues and lose activity at physiological temperatures or pH conditions [256]. For example, recombinant AHL-lactonase KMMI17, while effective in vitro, shows reduced activity in metal-deficient host environments, as its metallo-β-lactamase domain requires zinc ions for catalytic function—a resource often sequestered by the host during infection [172]. Natural product formulations face additional hurdles: chitosan, though effective in synergizing with antibiotics, exhibits erratic solubility in physiological fluids, leading to unpredictable bioavailability [265].
6.5. Specificity and Off-Target Effects
The lack of precision in targeting P. aeruginosa QS systems often results in unintended biological consequences, undermining both efficacy and safety. Enzymatic inhibitors, such as recombinant lectin rHPLOE, exemplify this limitation by binding exclusively to rhamnose-containing components of P. aeruginosa biofilms. While effective against strains producing rhamnolipid-rich matrices, this narrow specificity renders the enzyme useless against biofilms lacking these structures, significantly restricting its therapeutic scope [186]. Similarly, broad-spectrum enzymes like AidB degrade AHLs across bacterial species, indiscriminately disrupting QS communication in pathogenic and beneficial microbiota. This collateral damage risks dysbiosis, particularly in the gut or respiratory microbiomes, where commensal bacteria contribute to host defense and metabolic homeostasis [173].
Synthetic compounds further illustrate the trade-off between potency and specificity. Ketone-/cyano-selenoesters, designed to inhibit both AI-1 and AI-2 QS pathways, disrupt interspecies signaling networks in polymicrobial environments. While this broad activity may suppress virulence in P. aeruginosa, it destabilizes symbiotic microbial communities, potentially exacerbating infections by opportunistic pathogens [189]. Natural product-derived inhibitors, such as ortho-vanillin, selectively target the RhlR receptor but fail to inhibit Las or Pqs systems, allowing P. aeruginosa to maintain virulence through redundant QS pathways. This partial targeting is particularly problematic in chronic infections, where strain heterogeneity ensures the persistence of QS-active subpopulations [124].
Mechanistic ambiguity exacerbates specificity challenges. Cyclic dipeptides like tryptophan-containing CDPs interfere with QS in P. aeruginosa through poorly characterized interactions, raising concerns about off-target binding to host or microbial receptors unrelated to virulence [336]. Similarly, bromotyrosine-derived compounds like psammaplin A exhibit variable efficacy across QS-regulated genes (e.g., lasB vs. rhlA), suggesting inconsistent receptor affinity or unintended interactions with bacterial metabolic pathways [282]. These off-target effects complicate dose optimization, as higher concentrations may amplify unintended interactions.
6.6. Ecological and Safety Concerns
The environmental persistence and unintended biological interactions of QSI pose significant ecological and safety challenges, complicating their clinical and environmental deployment [337]. Nanoparticles, such as AgNPs and ZnO-NPs, exemplify these risks. While effective at disrupting QS signals and biofilms, AgNPs accumulate in aquatic ecosystems, persist indefinitely, and disrupt microbial communities. This environmental persistence promotes horizontal gene transfer of resistance markers among bacteria, exacerbating the global antibiotic resistance crisis [334]. Similarly, ZnO-NPs, though biodegradable under certain conditions, generate ROS that harm aquatic organisms and destabilize ecological balances in water systems [158].
Natural product-derived inhibitors introduce risks to both host microbiomes and human physiology. Fatty acyl compounds from S. griseoincarnatus non-specifically degrade QS molecules across bacterial species, disrupting beneficial commensal microbiota in niches like the gut or respiratory tract. Such dysbiosis can impair host immune function or metabolic health, creating vulnerabilities to secondary infections [259]. Phthalate derivatives, despite reported non-toxicity in initial studies, carry historical associations with endocrine disruption, raising concerns about chronic exposure effects such as hormonal imbalances or reproductive toxicity [277]. Mycotoxin-based compounds like citrinin further illustrate safety trade-offs; even at subinhibitory concentrations, they interfere with mitochondrial function in host cells, potentially exacerbating tissue damage in immunocompromised patients [272].
Non-specific therapeutic strategies amplify these risks. Though effective at generating ROS to disrupt QS, Gamma irradiation damages host tissues through oxidative stress, particularly in chronic wound or pulmonary infection models [268]. Similarly, phage-based QSI face regulatory hurdles due to their strain-specific activity and potential to provoke immune reactions, which may worsen inflammatory responses in sensitive populations [279]. Plant-derived formulations, such as multi-herbal extracts like Panchvalkal, suffer from batch variability due to inconsistent phytochemical profiles, leading to unpredictable ecological or toxicological outcomes [100].
Economic and manufacturing barriers further compound these concerns. Rare-earth nanoparticles (e.g., yttrium oxide nanosheets) and metallopharmaceuticals require resource-intensive synthesis protocols, raising costs and limiting accessibility in low-resource settings [156]. Hybrid delivery systems, such as bacteriophage-ZnO-NP conjugates, demand precise dosing regimens to balance efficacy and safety—a challenge that complicates clinical scalability [167].
6.7. Mechanistic Ambiguity
A fundamental barrier to developing QSI is the incomplete understanding of their modes of action, which stifles optimization and clinical translation. This mechanistic ambiguity arises when inhibitors interact with multiple targets or their primary biochemical interactions remain undefined, leading to unpredictable efficacy and off-target effects [338].
Natural product extracts exemplify this challenge. Multi-component formulations, such as those derived from P. maculosa or B. officinalis, exhibit synergistic anti-QS effects, but the contributions of individual phytochemicals are obscured. For instance, while these extracts reduce biofilm formation, the specific compounds responsible for LasR or RhlR inhibition are unidentified, complicating rational drug design [95]. Similarly, marine-derived B. paralicheniformis extracts require high concentrations for efficacy, yet their active principles remain uncharacterized, leaving researchers unable to isolate or enhance key molecules [257].
Synthetic and semi-synthetic compounds also suffer from unclear mechanisms. Tryptophan-containing cyclic dipeptides disrupt P. aeruginosa QS, but their precise molecular targets—receptor binding, signal molecule sequestration, or interference with transcriptional regulators—are poorly defined. This ambiguity hinders structure-activity relationship (SAR) studies, critical for potency optimization [336]. Bromotyrosine-derived inhibitors like psammaplin A further illustrate this issue. While they suppress lasB and rhlA gene expression, their variable efficacy across QS-regulated pathways (e.g., higher IC50 for lasB than rhlA) suggests non-specific interactions or off-target binding to bacterial metabolic enzymes [282].
Mycotoxin-derived compounds like citrinin introduce additional complexity by conflating anti-QS activity with host cell toxicity. Citrinin disrupts bacterial virulence at subinhibitory concentrations and impairs mitochondrial function in eukaryotic cells, implying dual mechanisms that are difficult to disentangle. This dual activity complicates dose optimization, as therapeutic windows narrow when antimicrobial and cytotoxic effects overlap [272].
Oxidative stress-inducing agents, including alkyl hydroperoxide E (AHE), exacerbate mechanistic uncertainty. While AHE inhibits QS by generating ROS, the exact pathways linking ROS to QS suppression—whether through signal molecule degradation, receptor oxidation, or transcriptional interference—are unresolved. This ambiguity risks unintended consequences, such as selecting antioxidant-rich bacterial mutants resistant to ROS and QS inhibition [339].
Fungal and actinobacterial extracts face reproducibility challenges due to uncharacterized bioactive components. For example, T. polium extracts fail to inhibit AHL-mediated QS in some studies, yet their variable phytochemical profiles across batches prevent definitive conclusions about their mechanism [94]. Similarly, P. avium cultivars exhibit inconsistent QS inhibition due to fluctuating polyphenol content, underscoring the need for standardized extraction protocols [85].
6.8. Partial Efficacy and Strain Dependency
The variable and incomplete suppression of virulence across P. aeruginosa strains or infection contexts represents a critical limitation of many QSI, undermining their reliability in clinical settings. Partial efficacy often stems from the redundancy of QS networks, where inhibitors targeting a single pathway fail to block alternative virulence mechanisms [340]. For instance, chrysin, a flavonoid, reduces pyocyanin production by only 41.4%, leaving significant residual virulence that sustains pathogenicity in chronic infections [341]. Similarly, isoeugenol achieves 70% biofilm inhibition at 400 μM—a concentration approaching cytotoxic thresholds—highlighting the narrow therapeutic window of many inhibitors [120]. Enzymatic inhibitors like AiiM lactonase exacerbate this issue by degrading AHLs without addressing downstream virulence effectors such as the T3SS, allowing P. aeruginosa to maintain cytotoxicity through non-QS pathways [171].
Strain dependency further complicates therapeutic outcomes. Farnesol, a natural sesquiterpene, exhibits reduced activity against mucoid P. aeruginosa phenotypes prevalent in CF airways. These strains overproduce alginate, a biofilm matrix component that physically shields QS receptors from inhibitor binding, rendering farnesol ineffective [129]. Similarly, plant-derived extracts from D. americana and H. abdariffa show discordant activity. While they inhibit pyocyanin production in vitro, their failure to reduce mortality in C. elegans infection models underscores the disconnect between biochemical assays and in vivo efficacy [90]. Such strain- or model-specific results challenge the extrapolation of preclinical data to human infections, where genetic and phenotypic diversity among clinical isolates is vast.
Biofilm heterogeneity amplifies these limitations. P. aeruginosa biofilms vary in matrix composition (e.g., alginate, Psl, Pel polysaccharides), and inhibitors targeting specific components, such as rhamnolipids or extracellular DNA, often fail against biofilms dominated by alternative polymers [342]. For example, recombinant lectin rHPLOE, which binds rhamnose residues, is ineffective against Pel-rich biofilms common in chronic wound infections [186]. This structural variability necessitates broad-spectrum inhibitors, yet most agents lack the versatility to adapt to dynamic biofilm architectures.
Compounding these issues, some inhibitors inadvertently exacerbate virulence. Malonate, while suppressing lasR expression, upregulates pqsC and increases pyocyanin production—a paradoxical effect that may worsen infection outcomes [138]. Similarly, pyranoanthocyanins enhance pqsE expression, potentially amplifying the production of cytotoxic factors [139].
6.9. Lack of In Vivo Validation
A critical translational gap for QSI is the overreliance on simplified in vitro or invertebrate models, leaving their efficacy and safety in mammalian systems inadequately characterized [343]. Many promising inhibitors fail to progress beyond early-stage preclinical testing due to insufficient validation in physiologically relevant in vivo environments. For instance, phage-derived proteins like PIT2 and synthetic peptides like Nesfactin have demonstrated robust anti-biofilm activity in vitro and invertebrate models (e.g., C. elegans) [181]. However, their performance in mammalian infection models—such as murine pneumonia or burn wound systems—remains unstudied, raising uncertainties about their pharmacokinetics, immunogenicity, or tissue penetration in complex host environments [178].
Natural product-derived inhibitors frequently suffer from this limitation. Psoralen, a coumarin derivative, reduces P. aeruginosa virulence in C. elegans but lacks validation in mammalian systems, where host-pathogen interactions, immune responses, and metabolic clearance pathways could diminish its efficacy [132]. Similarly, Boeravinone O, a flavonoid from Boerhaavia diffusa, exhibits potent QS inhibition in vitro but has not been tested in mammalian infection models, leaving its therapeutic potential speculative [101]. Even synthetic compounds like N-Aryl Malonamides (NAMs), despite showing efficacy in murine wound models, lack human clinical trials, leaving critical questions about dosage, toxicity, and bioavailability unanswered [344].
Nanoparticle-based strategies face similar translational hurdles. AgNPs and ZnC-NCs are often evaluated in in vitro biofilm assays or short-term invertebrate studies, bypassing chronic infection models that mimic human disease progression [345]. For example, proteomic analyses of AgNPs in P. aeruginosa-infected Galleria larvae reveal disrupted biofilm matrices. Still, these findings do not account for mammalian immune cell interactions or nanoparticle accumulation in organs like the liver or spleen [154]. Rare-earth nanoparticles (e.g., Y2O3 nanosheets) and bacteriophage-ZnO-NP conjugates, while effective in controlled lab settings, have not been assessed in large-animal models or human-relevant pharmacokinetic studies, stalling their clinical translation [156,167].
The consequences of this validation gap are profound. Invertebrate models, though cost-effective, lack the complexity of mammalian immune systems, tissue microenvironments, and metabolic pathways that influence drug efficacy. For instance, C. elegans cannot replicate the hypoxic conditions of CF lungs or the inflammatory milieu of chronic wounds, where P. aeruginosa biofilms thrive. Consequently, inhibitors like guanosine from M. comosus, validated only in C. elegans, may fail in human trials due to unanticipated host-pathogen interactions or rapid clearance by renal or hepatic systems [283,346].
6.10. Translational and Economic Barriers
The path to clinical adoption of QSI is fraught with economic, regulatory, and manufacturing challenges that extend beyond scientific limitations. Many promising compounds, particularly those derived from natural products or requiring complex synthesis, face prohibitive production costs [347]. For example, rare-earth nanoparticles like yttrium oxide nanosheets (Y2O3/NSs) demand resource-intensive synthesis protocols, including high-temperature calcination and specialized stabilizing agents, limiting scalability and affordability in low-resource healthcare settings [348]. Similarly, hybrid formulations such as SLNs co-loaded with antibiotics and QSI require precise control over lipid matrix stability during scale-up, often leading to batch variability and increased manufacturing complexity [15].
Regulatory hurdles further impede progress. Novel nanoparticle formulations or repurposed drugs must undergo rigorous safety and efficacy evaluations, which are complicated by the lack of standardized testing frameworks for antivirulence agents [349]. For instance, bacteriophage-ZnO-NP conjugates, while effective in lab settings, face uncertainty in regulatory approval due to their dual biological and inorganic components, necessitating extensive toxicological and environmental impact assessments [350]. Even well-characterized inhibitors like N-Aryl Malonamides (NAMs) struggle to advance to clinical trials without robust human pharmacokinetic data, as regulatory bodies prioritize compounds with established safety profiles [344].
Repurposed drugs, though economically attractive, introduce additional challenges. Ketoprofen derivatives, developed initially as anti-inflammatory agents, conflate QS inhibition with immune modulation, complicating regulatory approval due to ambiguous mechanisms of action [228]. Similarly, gamma irradiation—a cost-effective method to generate ROS for QS disruption—faces resistance from clinicians due to its non-specific tissue damage and lack of targeted delivery systems [268].
The economic viability of QSI is further strained by competition with conventional antibiotics, which dominate clinical practice despite rising resistance. Investors and pharmaceutical companies often prioritize antibiotics with immediate bactericidal effects over antivirulence agents, which are perceived as niche or adjunct therapies [351]. This bias stifles funding for large-scale trials, as seen with cyclic dipeptides or phage-derived proteins, which remain confined to academic research despite preclinical promise [178,336].
7. Future Perspectives
7.1. Structural Optimization and Multi-Target Inhibitor Design
The rational design of anti-QS molecules represents a cornerstone in combating P. aeruginosa resistance. It requires meticulous structural optimization to balance potency, specificity, and biocompatibility. This process integrates chemical synthesis, computational modeling, and synthetic biology to overcome the inherent limitations of early-generation QSI, such as poor pharmacokinetics, susceptibility to efflux pumps, and off-target cytotoxicity [352].
7.1.1. Chemical Modifications for Enhanced Bioavailability and Target Affinity
Structural refinement of core pharmacophores is critical to improving drug-like properties. For instance, flavones and quinazolinones—natural scaffolds with intrinsic QS inhibitory activity—are chemically tailored to enhance solubility and membrane permeability [214]. Selenoester derivatives, where sulfur atoms in thiolactone rings are replaced with selenium, demonstrate superior stability against bacterial lactonases while retaining strong LasR antagonism [353]. Similarly, halogenation of brominated furanones (GBr) increases their electrophilic reactivity, enabling covalent binding to LasR’s ligand-binding domain and irreversible disruption of QS signaling [206]. Structure-activity relationship studies guide these modifications to minimize unintended interactions with host cells.
7.1.2. Computational-Driven Design of Hybrid Inhibitors
Advances in in silico tools, such as molecular docking and molecular dynamics simulations, enable the systematic design of multi-target inhibitors [354]. For example, benzaldehyde derivatives are computationally optimized to bind conserved regions of both LasR and PqsR receptors, exploiting shared hydrophobic pockets to block autoinducer binding [124]. Machine learning algorithms further predict resistance-prone residues in QS receptors, guiding the synthesis of polymer-conjugated PqsR blockers with elongated alkyl side chains. These chains enhance biofilm penetration while avoiding recognition by efflux systems like MexAB-OprM [355]. Hybrid molecules, such as cephalosporin-QS inhibitor conjugates, exemplify dual functionality: the β-lactam core disrupts cell wall synthesis, while the appended furanone moiety inhibits RhlR-mediated virulence gene expression [194].
7.1.3. Synthetic Biology and Efflux Pump Evasion Strategies
Synthetic biology approaches re-engineer existing antimicrobial scaffolds to evade resistance mechanisms. Modified N-acylcysteine derivatives are redesigned with bulky substituents (e.g., tert-butyl groups) that sterically hinder binding to efflux pump channels, prolonging intracellular retention [356]. Similarly, selenium-doped nanoparticles loaded with QSI exploit bacterial redox systems: selenite reduction generates ROSs that weaken biofilm integrity, while concurrently releasing inhibitors like halogenated furanones to silence LasR [159]. These strategies are complemented by substrate mimicry—designing inhibitors structurally analogous to natural QS autoinducers (e.g., 3-oxo-C12-HSL) to competitively saturate receptor sites without activating virulence pathways competitively [207].
7.2. Synergistic Therapies and Combinatorial Approaches
Synergistic combinations exploit multiple pathways to disrupt bacterial virulence and potentiate antimicrobial activity. For instance, QSI, such as thymoquinone, degrade biofilm EPS, exposing embedded bacteria to antibiotics. Thymoquinone reduces exopolysaccharide production by 70%, enabling colistin to penetrate dual-species biofilms (S. aureus-P. aeruginosa) and reduce bacterial load by 4-log units [131]. Concurrently, cationic polymers like chitosan bind to MexAB-OprM efflux pumps, blocking antibiotic extrusion and increasing intracellular ceftazidime concentrations by 15-fold [265]. Subinhibitory doses of azithromycin combined with the QSI furanone C-30 enhance macrophage phagocytosis by increasing P. aeruginosa’s sensitivity to host defense mechanisms, particularly nitrosative stress. This effect is partly attributed to the interference with P. aeruginosa’s QS systems, which regulate immune evasion mechanisms such as the production of virulence factors like phzM and pvdA [357]. Engineered phage-probiomic systems, such as bacteriophage-ZnO nanocomposites, degrade mature biofilms [156] while silencing lasI via CRISPR-dCas9 interference, achieving 3-log reductions in burn infection models [358].
7.3. Engineered Biological Systems and Nanotechnology
7.3.1. Nanotechnology-Driven Delivery Platforms
NPs have emerged as versatile carriers for QSI, merging antivirulence and bactericidal functionalities. AgNPs functionalized with quorum-quenching enzymes, such as acylases, enzymatically degrade AHLs in biofilms while delivering bactericidal silver ions [152]. Hybrid nanomaterials like eugenol-conjugated AgNPs exploit hydrophobic interactions to dock into QS receptor pockets, blocking autoinducer binding and suppressing lasI/rhlR expression [305]. Similarly, ZnO-curcumin nanocomposites generate ROSs that disrupt LasR-mediated signaling while destabilizing biofilm matrices [158]. Innovations such as halogenated cerium oxide NPs mimic natural lactonase activity to address biocompatibility concerns, degrading QS signals without inducing oxidative damage to host tissues [162]. Stimuli-responsive nanocarriers further refine delivery; pH-sensitive liposomes release encapsulated caraway essential oils in acidic biofilm microenvironments, ensuring localized and sustained anti-QS activity [359].
7.3.2. Inhalable and Topical Formulations
Pulmonary and chronic wound infections demand site-specific delivery systems. Inhalable dry powder formulations, such as lactonase-loaded chitosan microparticles, target P. aeruginosa biofilms in CF airways, reducing lasB elastase production by 80% in preclinical models. These systems leverage mucoadhesive polymers to prolong residence time in the bronchial mucosa [360,361]. For chronic wounds, collagen-based hydrogels embedded with cytotoxic peptides (e.g., G3KL) enable localized delivery, disrupting biofilms without harming regenerating epithelial cells [362,363]. Cyclodextrin complexes sequester hydrophobic QSI, like hamamelitannin, enhancing solubility and enabling controlled release in exudative wound environments [364].
7.4. Translational Challenges and Ecological Integration
7.4.1. Preclinical Validation: Bridging the Gap Between Bench and Bedside
Robust preclinical models must replicate the polymicrobial, chronic infection landscapes seen in clinical settings. For instance, rodent catheter-associated biofilm models incorporate co-infections with Candida albicans and S. aureus to mimic diabetic foot ulcer microbiomes, enabling the evaluation of compounds like allopurinol-metformin hybrids under realistic conditions. These models assess biofilm eradication and host-pathogen dynamics, such as neutrophil recruitment and cytokine profiling [365,366]. Pharmacokinetic/pharmacodynamic (PK/PD) modeling is critical for optimizing dosing regimens; thymoquinone nanoformulations, for example, require adjusted dosing intervals in CF models due to altered mucus clearance rates [131]. Safety assessments extend beyond acute toxicity, with 90-day murine studies revealing that selenium-doped nanoparticles induce hepatic glutathione depletion at high doses, necessitating formulation refinements [367].
7.4.2. Standardization and Reproducibility
Natural product-derived QSI, such as propolis triterpenes and B. diffusa extracts, suffer from batch-to-batch variability due to seasonal and geographical factors [101,145]. Advanced phytochemical fingerprinting via HPLC-MS and AI-driven cultivation protocols is being deployed to standardize bioactive compound ratios [368]. Synthetic biology offers an alternative; engineered Saccharomyces cerevisiae strains now produce uniform yields of hamamelitannin analogs, achieving >95% purity for clinical-grade batches [369].
7.4.3. Ecological and Evolutionary Considerations
The ecological ripple effects of anti-QS therapies remain understudied. Horizontal gene transfer (HGT) risks are amplified in wastewater ecosystems, where engineered phages or plasmid-encoded QSI could disseminate resistance genes [370]. Metagenomic surveillance of hospital effluents post-trial revealed lasR mutations in non-target Acinetobacter spp., highlighting the need for biodegradable inhibitors [10]. For example, PLGA nanoparticles loaded with brominated furanones hydrolyze into lactic and glycolic acids, reducing environmental persistence [371,372]. Evolutionary trade-offs must also be managed: longitudinal exposure assays show that P. aeruginosa develops compensatory mutations in rhlR when treated with LasR-specific inhibitors, necessitating multi-target approaches [373]. CRISPR-Cas9-based allele-specific silencing of mutant lasR variants is being explored to preempt resistance [374].
7.4.4. Regulatory and Commercialization Hurdles
Regulatory agencies lack standardized guidelines for anti-QS therapies, particularly combination products like phage-nanoparticle conjugates. The FDA’s Complex Drug Advisory Committee now requires ecotoxicology dossiers for nanotherapeutics, including biofilm dispersal impacts on aquatic ecosystems [375]. Commercial scalability is another bottleneck; the Good Manufacturing Practice (GMP) production of stimuli-responsive nanocarriers costs 3–5 times more than conventional antibiotics, limiting low-resource market access [376]. Public-private partnerships, such as the CARB-X-funded “QuorumSafe” initiative, subsidize early-stage development to offset these barriers [377].
7.4.5. Future Directions: Toward Sustainable Anti-Infective Strategies
Next-generation translational frameworks prioritize One Health integration. Phase IV post-marketing surveillance will monitor long-term ecological impacts, using blockchain-enabled supply chains to track environmental dispersal of QSI [132]. Personalized medicine approaches, guided by patient-specific lasR SNP profiles and microbiome signatures, aim to optimize inhibitor-antibiotic pairings [378]. Meanwhile, “green” formulation strategies—such as algae-based chitosan production and solar-driven nanoparticle synthesis—reduce anti-QS therapeutics’ carbon footprint [379].
7.5. Mechanistic Insights and Resistance Management
Multi-target inhibition strategies are paramount to preempting resistance. Hybrid molecules such as efflux pump inhibitors, PAβN derivatives, covalently modify both LasR and MexAB-OprM efflux pumps, reducing virulence while enhancing intracellular antibiotic retention [192]. Natural polysaccharides like ulvan from green algae employ a dual mechanism—sequestering AHL signals via lactonase-mimetic activity and downregulating pqsABCDE through zinc ion chelation [380]. Synthetic biology approaches further enable “resistance-proof” therapies; engineered phages delivering dCas9-sgRNA complexes silence lasR and mexT simultaneously, suppressing compensatory pathway activation [381]. Combinatorial regimens, such as pairing PqsR inhibitors with tobramycin-loaded nanoparticles, exploit collateral sensitivity, where pqsR mutants exhibit 100-fold increased susceptibility to aminoglycosides due to disrupted membrane potential regulation [382].
8. Conclusions
Developing anti-QS therapies against P. aeruginosa is a promising yet complex endeavor constrained by biological redundancy, pharmacokinetic inefficiencies, and evolutionary adaptability. Overcoming these challenges requires a multidisciplinary approach, integrating structural chemistry, systems biology, and translational medicine. Multi-target inhibitors can bypass redundancy, advanced delivery systems enhance bioavailability, and rigorous in vivo validation ensures safety and efficacy. By harmonizing molecular optimization, synergistic combinations, and innovative delivery, these strategies promise to redefine infection management. Collaborative efforts across microbiology, pharmacology, and nanotechnology will translate anti-QS molecules from experimental tools into clinically viable solutions, offering a sustainable paradigm in the post-antibiotic era and curbing P. aeruginosa resistance.
Author Contributions
Conceptualization, L.T. and A.T.; methodology, L.T. and A.T.; validation, L.T., A.T. and T.I.; investigation, L.T. and A.T.; resources, L.T. and A.T. and T.I.; data curation, L.T. and T.I.; writing—original draft preparation, L.T., T.I. and A.T.; writing—review and editing, A.T. and T.I.; visualization, T.I. and N.A.I.; supervision, A.T. and T.I; project administration, A.T.; funding acquisition, N.A.I. All authors have read and agreed to the published version of the manuscript.
Funding
The Deanship of Scientific Research at Imam Mohammad Bin Saud Islamic University (IMSIU) supported and funded this work. (grant number IMSIU-DDRSP2501).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kerr, K.G.; Snelling, A.M. Pseudomonas aeruginosa: A Formidable and Ever-Present Adversary. J. Hosp. Infect. 2009, 73, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic Resistance in Pseudomonas aeruginosa: Mechanisms and Alternative Therapeutic Strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Strateva, T.; Mitov, I. Contribution of an Arsenal of Virulence Factors to Pathogenesis of Pseudomonas aeruginosa Infections. Ann Microbiol 2011, 61, 717–732. [Google Scholar] [CrossRef]
- Pearson, J.P.; Pesci, E.C.; Iglewski, B.H. Roles of Pseudomonas aeruginosa Las and Rhl Quorum-Sensing Systems in Control of Elastase and Rhamnolipid Biosynthesis Genes. J. Bacteriol. 1997, 179, 5756–5767. [Google Scholar] [CrossRef] [PubMed]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef]
- Rather, M.A.; Gupta, K.; Mandal, M. Microbial Biofilm: Formation, Architecture, Antibiotic Resistance, and Control Strategies. Braz. J. Microbiol. 2021, 52, 1701–1718. [Google Scholar] [CrossRef]
- Sharma, S.; Mohler, J.; Mahajan, S.D.; Schwartz, S.A.; Bruggemann, L.; Aalinkeel, R. Microbial Biofilm: A Review on Formation, Infection, Antibiotic Resistance, Control Measures, and Innovative Treatment. Microorganisms 2023, 11, 1614. [Google Scholar] [CrossRef]
- Haidar, A.; Muazzam, A.; Nadeem, A.; Atique, R.; Saeed, H.A.; Naveed, A.; Sharif, J.; Perveen, A.; Fatima, H.R.; Samad, A. Biofilm Formation and Antibiotic Resistance in Pseudomonas aeruginosa. Microbe 2024, 3, 100078. [Google Scholar] [CrossRef]
- Li, J.; Jia, T.; Yang, L. Targeting Anti-Virulence Factor Strategies of Bacterial Pathogens. Biosaf. Health 2025, 7, 1–4. [Google Scholar] [CrossRef]
- Aruwa, C.E.; Chellan, T.; S’thebe, N.W.; Dweba, Y.; Sabiu, S. ESKAPE Pathogens and Associated Quorum Sensing Systems: New Targets for Novel Antimicrobials Development. Health Sci. Rev. 2024, 11, 100155. [Google Scholar] [CrossRef]
- Lu, L.; Li, M.; Yi, G.; Liao, L.; Cheng, Q.; Zhu, J.; Zhang, B.; Wang, Y.; Chen, Y.; Zeng, M. Screening Strategies for Quorum Sensing Inhibitors in Combating Bacterial Infections. J. Pharm. Anal. 2022, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Truchado, P.; Larrosa, M.; Castro-Ibáñez, I.; Allende, A. Plant Food Extracts and Phytochemicals: Their Role as Quorum Sensing Inhibitors. Trends Food Sci. Technol. 2015, 43, 189–204. [Google Scholar] [CrossRef]
- Sengupta, A.; Bhattacharya, D.; Lahiri, D.; Nag, M.; Dey, A.; Ray, S. Gold and Silver Nanoparticles as Potent Quorum Quenchers: A Critical Review. BioNanoScience 2024, 15, 73. [Google Scholar] [CrossRef]
- García-Cruz, J.C.; Huelgas-Méndez, D.; Jiménez-Zúñiga, J.S.; Rebollar-Juárez, X.; Hernández-Garnica, M.; Fernández-Presas, A.M.; Husain, F.M.; Alenazy, R.; Alqasmi, M.; Albalawi, T.; et al. Myriad Applications of Bacteriophages beyond Phage Therapy. PeerJ 2023, 11, e15272. [Google Scholar] [CrossRef] [PubMed]
- Rodenak-Kladniew, B.; Scioli Montoto, S.; Sbaraglini, M.L.; Di Ianni, M.; Ruiz, M.E.; Talevi, A.; Alvarez, V.A.; Durán, N.; Castro, G.R.; Islan, G.A. Hybrid Ofloxacin/Eugenol Co-Loaded Solid Lipid Nanoparticles with Enhanced and Targetable Antimicrobial Properties. Int. J. Pharm. 2019, 569, 118575. [Google Scholar] [CrossRef]
- Chen, J.; Wang, B.; Lu, Y.; Guo, Y.; Sun, J.; Wei, B.; Zhang, H.; Wang, H. Quorum Sensing Inhibitors from Marine Microorganisms and Their Synthetic Derivatives. Mar. Drugs 2019, 17, 80. [Google Scholar] [CrossRef]
- Zhang, J.-W.; Guo, C.; Xuan, C.-G.; Gu, J.-W.; Cui, Z.-N.; Zhang, J.; Zhang, L.; Jiang, W.; Zhang, L.-Q. High-Throughput, Quantitative Screening of Quorum-Sensing Inhibitors Based on a Bacterial Biosensor. ACS Chem. Biol. 2023, 18, 2544–2554. [Google Scholar] [CrossRef]
- Monroy-Pérez, E.; Herrera-Gabriel, J.P.; Olvera-Navarro, E.; Ugalde-Tecillo, L.; García-Cortés, L.R.; Moreno-Noguez, M.; Martínez-Gregorio, H.; Vaca-Paniagua, F.; Paniagua-Contreras, G.L. Molecular Properties of Virulence and Antibiotic Resistance of Pseudomonas aeruginosa Causing Clinically Critical Infections. Pathogens 2024, 13, 868. [Google Scholar] [CrossRef]
- Sendra, E.; Fernández-Muñoz, A.; Zamorano, L.; Oliver, A.; Horcajada, J.P.; Juan, C.; Gómez-Zorrilla, S. Impact of Multidrug Resistance on the Virulence and Fitness of Pseudomonas aeruginosa: A Microbiological and Clinical Perspective. Infection 2024, 52, 1235–1268. [Google Scholar] [CrossRef]
- Newman, J.W.; Floyd, R.V.; Fothergill, J.L. The Contribution of Pseudomonas aeruginosa Virulence Factors and Host Factors in the Establishment of Urinary Tract Infections. FEMS Microbiol. Lett. 2017, 364, fnx124. [Google Scholar] [CrossRef]
- Zezzi do Valle Gomes, M.; Nitschke, M. Evaluation of Rhamnolipid and Surfactin to Reduce the Adhesion and Remove Biofilms of Individual and Mixed Cultures of Food Pathogenic Bacteria. Food Control 2012, 25, 441–447. [Google Scholar] [CrossRef]
- Høiby, N. Cystic Fibrosis and the Clinical Biofilm Revolution A Survey of the Danish CF Center’s Contribution. Biofilm 2025, 9, 100246. [Google Scholar] [CrossRef]
- Høiby, N. Understanding Bacterial Biofilms in Patients with Cystic Fibrosis: Current and Innovative Approaches to Potential Therapies. J. Cyst. Fibros. 2002, 1, 249–254. [Google Scholar] [CrossRef]
- Tuon, F.F.; Dantas, L.R.; Suss, P.H.; Tasca Ribeiro, V.S. Pathogenesis of the Pseudomonas aeruginosa Biofilm: A Review. Pathogens 2022, 11, 300. [Google Scholar] [CrossRef] [PubMed]
- Behzadi, P.; Baráth, Z.; Gajdács, M. It’s Not Easy Being Green: A Narrative Review on the Microbiology, Virulence and Therapeutic Prospects of Multidrug-Resistant Pseudomonas aeruginosa. Antibiotics 2021, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- de Kievit, T.R. Quorum Sensing in Pseudomonas aeruginosa Biofilms. Environ. Microbiol. 2009, 11, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.-C.; van Hullebusch, E.D.; Neu, T.R.; Nielsen, P.H.; Seviour, T.; Stoodley, P.; Wingender, J.; Wuertz, S. The Biofilm Matrix: Multitasking in a Shared Space. Nat. Rev. Microbiol. 2023, 21, 70–86. [Google Scholar] [CrossRef]
- Ciofu, O.; Rojo-Molinero, E.; Macià, M.D.; Oliver, A. Antibiotic Treatment of Biofilm Infections. APMIS 2017, 125, 304–319. [Google Scholar] [CrossRef]
- Sultan, M.; Alghetaa, H.; Mohammed, A.; Abdulla, O.A.; Wisniewski, P.J.; Singh, N.; Nagarkatti, P.; Nagarkatti, M. The Endocannabinoid Anandamide Attenuates Acute Respiratory Distress Syndrome by Downregulating miRNA That Target Inflammatory Pathways. Front. Pharmacol. 2021, 12, 644281. [Google Scholar] [CrossRef]
- Sánchez-Jiménez, A.; Llamas, M.A.; Marcos-Torres, F.J. Transcriptional Regulators Controlling Virulence in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2023, 24, 11895. [Google Scholar] [CrossRef]
- Wood, S.J.; Kuzel, T.M.; Shafikhani, S.H. Pseudomonas aeruginosa: Infections, Animal Modeling, and Therapeutics. Cells 2023, 12, 199. [Google Scholar] [CrossRef]
- Kretsch, A.M.; Morgan, G.L.; Acken, K.A.; Barr, S.A.; Li, B. Pseudomonas Virulence Factor Pathway Synthesizes Autoinducers That Regulate the Secretome of a Pathogen. ACS Chem. Biol. 2021, 16, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Yao, C.; Ding, Y.; Hu, H.; Qian, G.; He, M.; Deng, X. The Transcriptional Regulators of Virulence for Pseudomonas aeruginosa: Therapeutic Opportunity and Preventive Potential of Its Clinical Infections. Genes. Dis. 2023, 10, 2049–2063. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, A.A.; Kamer, A.M.A.; Al-Monofy, K.B.; Al-Madboly, L.A. Pseudomonas aeruginosa’s Greenish-Blue Pigment Pyocyanin: Its Production and Biological Activities. Microb. Cell Factories 2023, 22, 110. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zou, Y.; She, P.; Wu, Y. Composition, Function, and Regulation of T6SS in Pseudomonas aeruginosa. Microbiol. Res. 2015, 172, 19–25. [Google Scholar] [CrossRef]
- Prescott, R.D.; Decho, A.W. Flexibility and Adaptability of Quorum Sensing in Nature. Trends Microbiol. 2020, 28, 436–444. [Google Scholar] [CrossRef]
- O’Toole, G.A.; Kolter, R. Flagellar and Twitching Motility Are Necessary for Pseudomonas aeruginosa Biofilm Development. Mol. Microbiol. 1998, 30, 295–304. [Google Scholar] [CrossRef]
- Blanchard, A.C.; Waters, V.J. Microbiology of Cystic Fibrosis Airway Disease. Semin. Respir. Crit. Care Med. 2019, 40, 727–736. [Google Scholar] [CrossRef]
- Paz-Zarza, V.M.; Mangwani-Mordani, S.; Martínez-Maldonado, A.; Álvarez-Hernández, D.; Solano-Gálvez, S.G.; Vázquez-López, R. Pseudomonas aeruginosa: Pathogenicity and antimicrobial resistance in urinary tract infection. Rev. Chil. Infectologia 2019, 36, 180–189. [Google Scholar] [CrossRef]
- Ellison, C.K.; Whitfield, G.B.; Brun, Y.V. Type IV Pili: Dynamic Bacterial Nanomachines. FEMS Microbiol. Rev. 2022, 46, fuab053. [Google Scholar] [CrossRef] [PubMed]
- Kazmierczak, B.I.; Schniederberend, M.; Jain, R. Cross-Regulation of Pseudomonas Motility Systems: The Intimate Relationship between Flagella, Pili and Virulence. Curr. Opin. Microbiol. 2015, 28, 78–82. [Google Scholar] [CrossRef]
- Mogensen, T.H. Pathogen Recognition and Inflammatory Signaling in Innate Immune Defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef]
- Vance, R.E.; Isberg, R.R.; Portnoy, D.A. Patterns of Pathogenesis: Discrimination of Pathogenic and Non-Pathogenic Microbes by the Innate Immune System. Cell Host Microbe 2009, 6, 10–21. [Google Scholar] [CrossRef]
- Metelkina, O.; Konstantinović, J.; Klein, A.; Shafiei, R.; Fares, M.; Alhayek, A.; Yahiaoui, S.; Elgaher, W.A.M.; Haupenthal, J.; Titz, A.; et al. Dual Inhibitors of Pseudomonas aeruginosa Virulence Factors LecA and LasB. Chem. Sci. 2024, 15, 13333–13342. [Google Scholar] [CrossRef] [PubMed]
- Folkesson, A.; Jelsbak, L.; Yang, L.; Johansen, H.K.; Ciofu, O.; Høiby, N.; Molin, S. Adaptation of Pseudomonas aeruginosa to the Cystic Fibrosis Airway: An Evolutionary Perspective. Nat. Rev. Microbiol. 2012, 10, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.; Kollef, M. The Epidemiology and Pathogenesis and Treatment of Pseudomonas aeruginosa Infections: An Update. Drugs 2021, 81, 2117–2131. [Google Scholar] [CrossRef]
- Wagener, B.M.; Hu, R.; Wu, S.; Pittet, J.-F.; Ding, Q.; Che, P. The Role of Pseudomonas aeruginosa Virulence Factors in Cytoskeletal Dysregulation and Lung Barrier Dysfunction. Toxins 2021, 13, 776. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, J.T.; Sun, J. Pseudomonas aeruginosa ExoS and ExoT. Rev. Physiol. Biochem. Pharmacol. 2004, 152, 79–92. [Google Scholar] [CrossRef]
- Foulkes, D.M.; McLean, K.; Haneef, A.S.; Fernig, D.G.; Winstanley, C.; Berry, N.; Kaye, S.B. Pseudomonas aeruginosa Toxin ExoU as a Therapeutic Target in the Treatment of Bacterial Infections. Microorganisms 2019, 7, 707. [Google Scholar] [CrossRef]
- Zhu, Y.; Jia, P.; Yu, W.; Chu, X.; Liu, X.; Yang, Q. The Epidemiology and Virulence of Carbapenem-Resistant Pseudomonas aeruginosa in China. Lancet Microbe 2023, 4, e665. [Google Scholar] [CrossRef]
- Belyy, A.; Mechold, U.; Renault, L.; Ladant, D. ExoY, an Actin-Activated Nucleotidyl Cyclase Toxin from P. aeruginosa: A Minireview. Toxicon 2018, 149, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Wretlind, B.; Pavlovskis, O.R. Pseudomonas aeruginosa Elastase and Its Role in Pseudomonas Infections. Rev. Infect. Dis. 1983, 5, S998–S1004. [Google Scholar] [CrossRef] [PubMed]
- Everett, M.J.; Davies, D.T. Pseudomonas aeruginosa Elastase (LasB) as a Therapeutic Target. Drug Discov. Today 2021, 26, 2108–2123. [Google Scholar] [CrossRef] [PubMed]
- Galdino, A.C.M.; Branquinha, M.H.; Santos, A.L.S.; Viganor, L. Pseudomonas aeruginosa and Its Arsenal of Proteases: Weapons to Battle the Host. In Pathophysiological Aspects of Proteases; Chakraborti, S., Dhalla, N.S., Eds.; Springer: Singapore, 2017; pp. 381–397. ISBN 978-981-10-6141-7. [Google Scholar]
- West, S.E.H. Pseudomonas aeruginosa Exotoxin A: Structure/Function, Production, and Intoxication of Eukaryotic Cells. In Bacterial Protein Toxins; Aktories, K., Just, I., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 67–89. ISBN 978-3-662-05971-5. [Google Scholar]
- Chai, Y.-H.; Xu, J.-F. How Does Pseudomonas aeruginosa Affect the Progression of Bronchiectasis? Clin. Microbiol. Infect. 2020, 26, 313–318. [Google Scholar] [CrossRef]
- Soberón-Chávez, G.; González-Valdez, A.; Soto-Aceves, M.P.; Cocotl-Yañez, M. Rhamnolipids Produced by Pseudomonas: From Molecular Genetics to the Market. Microb. Biotechnol. 2020, 14, 136–146. [Google Scholar] [CrossRef]
- Maier, R.M.; Soberón-Chávez, G. Pseudomonas aeruginosa Rhamnolipids: Biosynthesis and Potential Applications. Appl. Microbiol. Biotechnol. 2000, 54, 625–633. [Google Scholar] [CrossRef]
- Bertrand, Q.; Job, V.; Maillard, A.P.; Imbert, L.; Teulon, J.-M.; Favier, A.; Pellequer, J.-L.; Huber, P.; Attrée, I.; Dessen, A. Exolysin (ExlA) from Pseudomonas aeruginosa Punctures Holes into Target Membranes Using a Molten Globule Domain. J. Mol. Biol. 2020, 432, 4466–4480. [Google Scholar] [CrossRef]
- Reboud, E.; Basso, P.; Maillard, A.P.; Huber, P.; Attrée, I. Exolysin Shapes the Virulence of Pseudomonas aeruginosa Clonal Outliers. Toxins 2017, 9, 364. [Google Scholar] [CrossRef]
- Cohen, T.S.; Parker, D.; Prince, A. Pseudomonas aeruginosa Host Immune Evasion. In Pseudomonas: Volume 7: New Aspects of Pseudomonas Biology; Ramos, J.-L., Goldberg, J.B., Filloux, A., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 3–23. ISBN 978-94-017-9555-5. [Google Scholar]
- Huszczynski, S.M.; Lam, J.S.; Khursigara, C.M. The Role of Pseudomonas aeruginosa Lipopolysaccharide in Bacterial Pathogenesis and Physiology. Pathogens 2019, 9, 6. [Google Scholar] [CrossRef]
- Sawa, T.; Shimizu, M.; Moriyama, K.; Wiener-Kronish, J.P. Association between Pseudomonas aeruginosa Type III Secretion, Antibiotic Resistance, and Clinical Outcome: A Review. Crit. Care 2014, 18, 668. [Google Scholar] [CrossRef]
- Boyd, A.; Chakrabarty, A.M. Pseudomonas aeruginosa Biofilms: Role of the Alginate Exopolysaccharide. J. Ind. Microbiol. 1995, 15, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Establishment of Pseudomonas aeruginosa Infection: Lessons from a Versatile Opportunist1. Microbes Infect. 2000, 2, 1051–1060. [Google Scholar] [CrossRef]
- Kolar, S.L.; Liu, G.Y. RAS and ROS—A Story of Pseudomonas Survival. Cell Host Microbe 2017, 21, 551–552. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Wang, Y.; Xiao, J.; Wang, Y.; Zhu, X.; Xu, B.; Wang, M. Scavenging of Reactive Oxygen Species Effectively Reduces Pseudomonas aeruginosa Biofilms through Disrupting Policing. Environ. Res. 2023, 220, 115182. [Google Scholar] [CrossRef]
- Minagawa, S.; Inami, H.; Kato, T.; Sawada, S.; Yasuki, T.; Miyairi, S.; Horikawa, M.; Okuda, J.; Gotoh, N. RND Type Efflux Pump System MexAB-OprM of Pseudomonas aeruginosa Selects Bacterial Languages, 3-Oxo-Acyl-Homoserine Lactones, for Cell-to-Cell Communication. BMC Microbiol. 2012, 12, 70. [Google Scholar] [CrossRef] [PubMed]
- Henriquez, T.; Falciani, C. Extracellular Vesicles of Pseudomonas: Friends and Foes. Antibiotics 2023, 12, 703. [Google Scholar] [CrossRef]
- Huber, P.; Basso, P.; Reboud, E.; Attrée, I. Pseudomonas aeruginosa Renews Its Virulence Factors. Environ. Microbiol. Rep. 2016, 8, 564–571. [Google Scholar] [CrossRef]
- Augustyniak, D.; Olszak, T.; Drulis-Kawa, Z. Outer Membrane Vesicles (OMVs) of Pseudomonas aeruginosa Provide Passive Resistance but Not Sensitization to LPS-Specific Phages. Viruses 2022, 14, 121. [Google Scholar] [CrossRef]
- Malhotra, S.; Hayes, D.; Wozniak, D.J. Cystic Fibrosis and Pseudomonas aeruginosa: The Host-Microbe Interface. Clin. Microbiol. Rev. 2019, 32, e00138-18. [Google Scholar] [CrossRef]
- Mulcahy, H.; Charron-Mazenod, L.; Lewenza, S. Extracellular DNA Chelates Cations and Induces Antibiotic Resistance in Pseudomonas aeruginosa Biofilms. PLoS Pathog. 2008, 4, e1000213. [Google Scholar] [CrossRef]
- Geisinger, E.; Isberg, R.R. Interplay Between Antibiotic Resistance and Virulence During Disease Promoted by Multidrug-Resistant Bacteria. J. Infect. Dis. 2017, 215, S9–S17. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Chauhan, K.; Singh, G.; Chaudhary, S.; Rathore, J.S. Decoding Antibiotic Resistance in Pseudomonas aeruginosa: Embracing Innovative Therapies beyond Conventional Antibiotics. Microbe 2025, 6, 100233. [Google Scholar] [CrossRef]
- Lorusso, A.B.; Carrara, J.A.; Barroso, C.D.N.; Tuon, F.F.; Faoro, H. Role of Efflux Pumps on Antimicrobial Resistance in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2022, 23, 15779. [Google Scholar] [CrossRef] [PubMed]
- Pechère, J.-C.; Köhler, T. Patterns and Modes of β-Lactam Resistance in Pseudomonas aeruginosa. Clin. Microbiol. Infect. 1999, 5, S15–S18. [Google Scholar] [CrossRef]
- Botelho, J.; Grosso, F.; Peixe, L. Antibiotic Resistance in Pseudomonas aeruginosa—Mechanisms, Epidemiology and Evolution. Drug Resist. Updat. 2019, 44, 100640. [Google Scholar] [CrossRef]
- Wen, A.X.; Herman, C. Horizontal Gene Transfer and beyond: The Delivery of Biological Matter by Bacterial Membrane Vesicles to Host and Bacterial Cells. Curr. Opin. Microbiol. 2024, 81, 102525. [Google Scholar] [CrossRef]
- Rodríguez-Urretavizcaya, B.; Vilaplana, L.; Marco, M.-P. Strategies for Quorum Sensing Inhibition as a Tool for Controlling Pseudomonas aeruginosa Infections. Int. J. Antimicrob. Agents 2024, 64, 107323. [Google Scholar] [CrossRef]
- Duplantier, M.; Lohou, E.; Sonnet, P. Quorum Sensing Inhibitors to Quench P. aeruginosa Pathogenicity. Pharmaceuticals 2021, 14, 1262. [Google Scholar] [CrossRef]
- Koh, C.-L.; Sam, C.-K.; Yin, W.-F.; Tan, L.Y.; Krishnan, T.; Chong, Y.M.; Chan, K.-G. Plant-Derived Natural Products as Sources of Anti-Quorum Sensing Compounds. Sensors 2013, 13, 6217–6228. [Google Scholar] [CrossRef]
- Khan, M.A.; Celik, I.; Khan, H.M.; Shahid, M.; Shahzad, A.; Kumar, S.; Ahmed, B. Antibiofilm and Anti-Quorum Sensing Activity of Psidium guajava L. Leaf Extract: In Vitro and In Silico Approach. PLoS ONE 2023, 18, e0295524. [Google Scholar] [CrossRef]
- Doğan, Ş.; Gökalsın, B.; Şenkardeş, İ.; Doğan, A.; Sesal, N.C. Anti-Quorum Sensing and Anti-Biofilm Activities of Hypericum perforatum Extracts against Pseudomonas aeruginosa. J. Ethnopharmacol. 2019, 235, 293–300. [Google Scholar] [CrossRef]
- Önem, E.; Sarısu, H.C.; Özaydın, A.G.; Muhammed, M.T.; Ak, A. Phytochemical Profile, Antimicrobial, and Anti-Quorum Sensing Properties of Fruit Stalks of Prunus avium L. Lett. Appl. Microbiol. 2021, 73, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Qaralleh, H. Chemical Composition and Quorum Sensing Inhibitory Effect of Nepeta curviflora Methanolic Extract against ESBL Pseudomonas aeruginosa. J. Pharmacopunct. 2023, 26, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, I.; Abbas, H.A.; Ashour, M.L.; Yasri, A.; El-Shazly, A.M.; Wink, M.; Sobeh, M. Polyphenols from Salix tetrasperma Impair Virulence and Inhibit Quorum Sensing of Pseudomonas aeruginosa. Molecules 2020, 25, 1341. [Google Scholar] [CrossRef]
- Vijayakumar, K.; Ramanathan, T. Musa acuminata and Its Bioactive Metabolite 5-Hydroxymethylfurfural Mitigates Quorum Sensing (Las and Rhl) Mediated Biofilm and Virulence Production of Nosocomial Pathogen Pseudomonas aeruginosa In Vitro. J. Ethnopharmacol. 2020, 246, 112242. [Google Scholar] [CrossRef]
- Jamalifar, H.; Samadi, N.; Nowroozi, J.; Dezfulian, M.; Fazeli, M.R. Down-Regulatory Effects of Green Coffee Extract on Las I and Las R Virulence-Associated Genes in Pseudomonas aeruginosa. DARU J. Pharm. Sci. 2019, 27, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Núñez, J.L.; Pérez-López, M.; Espinosa, N.; Campos-Hernández, N.; García-Contreras, R.; Díaz-Guerrero, M.; Cortes-López, H.; Vázquez-Sánchez, M.; Quezada, H.; Martínez-Vázquez, M.; et al. Anti-Virulence Properties of Plant Species: Correlation between In Vitro Activity and Efficacy in a Murine Model of Bacterial Infection. Microorganisms 2021, 9, 2424. [Google Scholar] [CrossRef]
- Křížkovská, B.; Hoang, L.; Brdová, D.; Klementová, K.; Szemerédi, N.; Loučková, A.; Kronusová, O.; Spengler, G.; Kaštánek, P.; Hajšlová, J.; et al. Modulation of the Bacterial Virulence and Resistance by Well-Known European Medicinal Herbs. J. Ethnopharmacol. 2023, 312, 116484. [Google Scholar] [CrossRef]
- Wang, L.; Huang, Y.; Yin, G.; Wang, J.; Wang, P.; Chen, Z.-Y.; Wang, T.; Ren, G. Antimicrobial Activities of Asian Ginseng, American Ginseng, and Notoginseng. Phytother Res. 2020, 34, 1226–1236. [Google Scholar] [CrossRef]
- Gupta, K.; Singh, S.P.; Manhar, A.K.; Saikia, D.; Namsa, N.D.; Konwar, B.K.; Mandal, M. Inhibition of Staphylococcus aureus and Pseudomonas aeruginosa Biofilm and Virulence by Active Fraction of Syzygium cumini (L.) Skeels Leaf Extract: In-Vitro and In Silico Studies. Indian J. Microbiol. 2019, 59, 13–21. [Google Scholar] [CrossRef]
- Alreshidi, M.; Noumi, E.; Bouslama, L.; Ceylan, O.; Veettil, V.N.; Adnan, M.; Danciu, C.; Elkahoui, S.; Badraoui, R.; Al-Motair, K.A.; et al. Phytochemical Screening, Antibacterial, Antifungal, Antiviral, Cytotoxic, and Anti-Quorum-Sensing Properties of Teucrium polium L. Aerial Parts Methanolic Extract. Plants 2020, 9, 1418. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, M.; Morić, I.; Nikolić, B.; Pavić, A.; Svirčev, E.; Šenerović, L.; Mitić-Ćulafić, D. Anti-Virulence Potential and In Vivo Toxicity of Persicaria maculosa and Bistorta officinalis Extracts. Molecules 2020, 25, 1811. [Google Scholar] [CrossRef]
- Moradi, F.; Hadi, N.; Bazargani, A. Evaluation of Quorum-Sensing Inhibitory Effects of Extracts of Three Traditional Medicine Plants with Known Antibacterial Properties. New Microbes New Infect 2020, 38, 100769. [Google Scholar] [CrossRef] [PubMed]
- Miari, M.; Rasheed, S.S.; Haidar Ahmad, N.; Itani, D.; Abou Fayad, A.; Matar, G.M. Natural Products and Polysorbates: Potential Inhibitors of Biofilm Formation in Pseudomonas aeruginosa. J. Infect. Dev. Ctries. 2020, 14, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Xu, H.; Huang, T.; Liu, G.; Cao, H.; Lin, Y.; Li, Y.; Li, Y.; Yao, X. Fuzheng Touxie Jiedu Huayu Decoction Inhibits the MexAB-OprM Efflux Pump and Quorum Sensing-Mediated Biofilm Formation in Difficult-to-Treat Multidrug Resistance Pseudomonas aeruginosa. J. Ethnopharmacol. 2024, 332, 118365. [Google Scholar] [CrossRef]
- Ruparel, F.J.; Shah, S.K.; Patel, J.H.; Thakkar, N.R.; Gajera, G.N.; Kothari, V.O. Network Analysis for Identifying Potential Anti-Virulence Targets from Whole Transcriptome of Pseudomonas aeruginosa and Staphylococcus aureus Exposed to Certain Anti-Pathogenic Polyherbal Formulations. Drug Target Insights 2023, 17, 58–69. [Google Scholar] [CrossRef]
- Joshi, C.; Patel, P.; Palep, H.; Kothari, V. Validation of the Anti-Infective Potential of a Polyherbal “Panchvalkal” Preparation, and Elucidation of the Molecular Basis Underlining Its Efficacy against Pseudomonas aeruginosa. BMC Complement. Altern. Med. 2019, 19, 19. [Google Scholar] [CrossRef]
- Shravani, V.; Girija, A.S.; Krishnan, M.; Babu, S. Anti-Quorum Sensing Activity of Boerhavia diffusa against Pseudomonas aeruginosa PAO1. Bioinformation 2023, 19, 310–318. [Google Scholar] [CrossRef]
- Zhao, Y.; Seenivasan, B.; Li, R.; Li, C.; Zhang, Y.; Ravichandran, V.; Zhong, L.; Li, A. Exploring Daidzein Dimethyl Ether from Albizzia lebbeck as a Novel Quorum Sensing Inhibitor against Pseudomonas aeruginosa: Insights from In Vitro and In Vivo Studies. Bioorg. Chem. 2025, 156, 108168. [Google Scholar] [CrossRef]
- Hnamte, S.; Parasuraman, P.; Ranganathan, S.; Ampasala, D.R.; Reddy, D.; Kumavath, R.N.; Suchiang, K.; Mohanty, S.K.; Busi, S. Mosloflavone Attenuates the Quorum Sensing Controlled Virulence Phenotypes and Biofilm Formation in Pseudomonas aeruginosa PAO1: In Vitro, In Vivo and In Silico Approach. Microb. Pathog. 2019, 131, 128–134. [Google Scholar] [CrossRef]
- Wang, S.; Feng, Y.; Han, X.; Cai, X.; Yang, L.; Liu, C.; Shen, L. Inhibition of Virulence Factors and Biofilm Formation by Wogonin Attenuates Pathogenicity of Pseudomonas aeruginosa PAO1 via Targeting Pqs Quorum-Sensing System. Int. J. Mol. Sci. 2021, 22, 12699. [Google Scholar] [CrossRef]
- Nayak, S.P.R.R.; Boopathi, S.; Priya, P.S.; Pasupuleti, M.; Pachaiappan, R.; Almutairi, B.O.; Arokiyaraj, S.; Arockiaraj, J. Luteolin, a Promising Quorum Quencher Mitigates Virulence Factors Production in Pseudomonas aeruginosa—In Vitro and In Vivoapproach. Microb. Pathog. 2023, 180, 106123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, S.; Cheng, Y. Antibiofilm Activity of Allicin and Quercetin in Treating Biofilm-Associated Orthopaedics Infection. Appl. Biochem. Biotechnol. 2022, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Behera, S.K.; Panda, A.K.; Mishra, R.; Mahanty, A.; Bisht, S.S. Structure Based Virtual Screening and Molecular Dynamics of Natural Anti-Biofilm Compounds against SagS Response Regulator/Sensor Kinase in Pseudomonas aeruginosa. J. Biomol. Struct. Dyn. 2023, 41, 6011–6026. [Google Scholar] [CrossRef] [PubMed]
- Ghannay, S.; Aouadi, K.; Kadri, A.; Snoussi, M. GC-MS Profiling, Vibriocidal, Antioxidant, Antibiofilm, and Anti-Quorum Sensing Properties of Carum carvi L. Essential Oil: In Vitro and In Silico Approaches. Plants 2022, 11, 1072. [Google Scholar] [CrossRef]
- Wang, W.; Li, D.; Huang, X.; Yang, H.; Qiu, Z.; Zou, L.; Liang, Q.; Shi, Y.; Wu, Y.; Wu, S.; et al. Study on Antibacterial and Quorum-Sensing Inhibition Activities of Cinnamomum camphora Leaf Essential Oil. Molecules 2019, 24, 3792. [Google Scholar] [CrossRef]
- S’thebe, N.W.; Aribisala, J.O.; Sabiu, S. Cheminformatics Bioprospection of Sunflower Seeds’ Oils against Quorum Sensing System of Pseudomonas aeruginosa. Antibiotics 2023, 12, 504. [Google Scholar] [CrossRef]
- Wira Septama, A.; Arvia Chiara, M.; Turnip, G.; Nur Tasfiyati, A.; Triana Dewi, R.; Anggrainy Sianipar, E.; Jaisi, A. Essential Oil of Zingiber cassumunar Roxb. and Zingiber officinale Rosc.: A Comparative Study on Chemical Constituents, Antibacterial Activity, Biofilm Formation, and Inhibition of Pseudomonas aeruginosa Quorum Sensing System. Chem. Biodivers 2023, 20, e202201205. [Google Scholar] [CrossRef]
- Yi, Y.; Zhou, Y.; Lin, S.; Shi, K.; Mei, J.; Ying, G.; Wu, S. Screening and Isolation of Quorum Sensing Inhibitors of Pseudomonas aeruginosa from Phellodendron amurense Extracts Using Bio-Affinity Chromatography. J. Sep. Sci. 2024, 47, e2400222. [Google Scholar] [CrossRef]
- Chakraborty, P.; Dastidar, D.G.; Paul, P.; Dutta, S.; Basu, D.; Sharma, S.R.; Basu, S.; Sarker, R.K.; Sen, A.; Sarkar, A.; et al. Inhibition of Biofilm Formation of Pseudomonas aeruginosa by Caffeine: A Potential Approach for Sustainable Management of Biofilm. Arch. Microbiol. 2020, 202, 623–635. [Google Scholar] [CrossRef]
- Kar, A.; Mukherjee, S.K.; Barik, S.; Hossain, S.T. Antimicrobial Activity of Trigonelline Hydrochloride Against Pseudomonas aeruginosa and Its Quorum-Sensing Regulated Molecular Mechanisms on Biofilm Formation and Virulence. ACS Infect. Dis. 2024, 10, 746–762. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wu, L.; Su, Y.; Huang, Z.; Wang, L.; Xia, Z.; Huang, H.; Wang, W.; Fang, J.; Gu, Z.; et al. Inhibitory Effects of Compounds from Plumula nelumbinis on Biofilm and Quorum Sensing Against P. aeruginosa. Curr. Microbiol. 2022, 79, 236. [Google Scholar] [CrossRef]
- El-Naggar, N.E.-A.; Bashir, S.I.; Rabei, N.H.; Saber, W.I.A. Innovative Biosynthesis, Artificial Intelligence-Based Optimization, and Characterization of Chitosan Nanoparticles by Streptomyces Microflavus and Their Inhibitory Potential against Pectobacterium Carotovorum. Sci. Rep. 2022, 12, 21851. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Ma, X.; Zheng, Y.; Liu, D.; Ning, W.; Xiao, W.; Mao, Q.; Bai, Z.; Mao, R.; Cheng, J.; et al. Traditional Chinese Medicine Monomer Bakuchiol Attenuates the Pathogenicity of Pseudomonas aeruginosa via Targeting PqsR. Int. J. Mol. Sci. 2024, 26, 243. [Google Scholar] [CrossRef]
- Tapia-Rodriguez, M.R.; Bernal-Mercado, A.T.; Gutierrez-Pacheco, M.M.; Vazquez-Armenta, F.J.; Hernandez-Mendoza, A.; Gonzalez-Aguilar, G.A.; Martinez-Tellez, M.A.; Nazzaro, F.; Ayala-Zavala, J.F. Virulence of Pseudomonas aeruginosa Exposed to Carvacrol: Alterations of the Quorum Sensing at Enzymatic and Gene Levels. J. Cell Commun. Signal. 2019, 13, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Singh, V.; Nag, S.; Roy, D.N. Carvacrol, a Monoterpenoid, Binds Quorum Sensing Proteins (LasI and LasR) and Swarming Motility Protein BswR of Pseudomonas aeruginosa, Resulting in Loss of Pathogenicity: An In Silico Approach. Can. J. Microbiol. 2025, 71, 1–15. [Google Scholar] [CrossRef]
- Shastry, R.P.; Kanekar, S.; Pandial, A.S.; Rekha, P.D. Isoeugenol Suppresses Multiple Quorum Sensing Regulated Phenotypes and Biofilm Formation of Pseudomonas aeruginosa PAO1. Nat. Prod. Res. 2022, 36, 1663–1667. [Google Scholar] [CrossRef]
- Deng, J.; Yuan, Y.; Wu, Y.; Wen, F.; Yang, X.; Gou, S.; Chu, Y.; Zhao, K. Isovanillin Decreases the Virulence Regulated by the Quorum Sensing System of Pseudomonas aeruginosa. Microb. Pathog. 2024, 196, 107010. [Google Scholar] [CrossRef]
- Yang, D.; Hao, S.; Zhao, L.; Shi, F.; Ye, G.; Zou, Y.; Song, X.; Li, L.; Yin, Z.; He, X.; et al. Paeonol Attenuates Quorum-Sensing Regulated Virulence and Biofilm Formation in Pseudomonas aeruginosa. Front. Microbiol. 2021, 12, 692474. [Google Scholar] [CrossRef]
- Mok, N.; Chan, S.Y.; Liu, S.Y.; Chua, S.L. Vanillin Inhibits PqsR-Mediated Virulence in Pseudomonas aeruginosa. Food Funct. 2020, 11, 6496–6508. [Google Scholar] [CrossRef]
- Woods, K.E.; Akhter, S.; Rodriguez, B.; Townsend, K.A.; Smith, N.; Smith, B.; Wambua, A.; Craddock, V.; Abisado-Duque, R.G.; Santa, E.E.; et al. Characterization of Natural Product Inhibitors of Quorum Sensing Reveals Competitive Inhibition of Pseudomonas aeruginosa RhlR by Ortho-Vanillin. Microbiol. Spectr. 2024, 12, e0068124. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhang, A.; Chu, W. Phillyrin Is an Effective Inhibitor of Quorum Sensing with Potential as an Anti-Pseudomonas aeruginosa Infection Therapy. J. Vet. Med. Sci. 2019, 81, 473–479. [Google Scholar] [CrossRef]
- Anju, V.T.; Busi, S.; Ranganathan, S.; Ampasala, D.R.; Kumar, S.; Suchiang, K.; Kumavath, R.; Dyavaiah, M. Sesamin and Sesamolin Rescues Caenorhabditis elegans from Pseudomonas aeruginosa Infection through the Attenuation of Quorum Sensing Regulated Virulence Factors. Microb. Pathog. 2021, 155, 104912. [Google Scholar] [CrossRef]
- Elfaky, M.A.; Elbaramawi, S.S.; Eissa, A.G.; Ibrahim, T.S.; Khafagy, E.-S.; Ali, M.A.M.; Hegazy, W.A.H. Drug Repositioning: Doxazosin Attenuates the Virulence Factors and Biofilm Formation in Gram-Negative Bacteria. Appl. Microbiol. Biotechnol. 2023, 107, 3763–3778. [Google Scholar] [CrossRef] [PubMed]
- Lima, E.M.F.; de Almeida, F.A.; Pinto, U.M. Exploring the Antivirulence Potential of Phenolic Compounds to Inhibit Quorum Sensing in Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2025, 41, 32. [Google Scholar] [CrossRef]
- Kalgudi, R.; Tamimi, R.; Kyazze, G.; Keshavarz, T. Effect of Quorum Quenchers on Virulence Factors Production and Quorum Sensing Signalling Pathway of Non-Mucoid, Mucoid, and Heavily Mucoid Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2022, 38, 163. [Google Scholar] [CrossRef]
- Chen, H.; Ji, P.-C.; Qi, Y.-H.; Chen, S.-J.; Wang, C.-Y.; Yang, Y.-J.; Zhao, X.-Y.; Zhou, J.-W. Inactivation of Pseudomonas aeruginosa Biofilms by Thymoquinone in Combination with Nisin. Front. Microbiol. 2022, 13, 1029412. [Google Scholar] [CrossRef] [PubMed]
- Al-Rabia, M.W.; Asfour, H.Z.; Alhakamy, N.A.; Abdulaal, W.H.; Ibrahim, T.S.; Abbas, H.A.; Salem, I.M.; Hegazy, W.A.H.; Nazeih, S.I. Thymoquinone Is a Natural Antibiofilm and Pathogenicity Attenuating Agent in Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2024, 14, 1382289. [Google Scholar] [CrossRef]
- Wen, F.; Wu, Y.; Yuan, Y.; Yang, X.; Ran, Q.; Gan, X.; Guo, Y.; Wang, X.; Chu, Y.; Zhao, K. Discovery of Psoralen as a Quorum Sensing Inhibitor Suppresses Pseudomonas aeruginosa Virulence. Appl. Microbiol. Biotechnol. 2024, 108, 222. [Google Scholar] [CrossRef]
- Kasthuri, T.; Barath, S.; Nandhakumar, M.; Karutha Pandian, S. Proteomic Profiling Spotlights the Molecular Targets and the Impact of the Natural Antivirulent Umbelliferone on Stress Response, Virulence Factors, and the Quorum Sensing Network of Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2022, 12, 998540. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, S.-Y.; Hu, J.-Y.; Chen, Q.-X.; Jiao, S.-M.; Xiao, H.-C.; Zhang, Q.; Xu, J.; Zhao, J.-F.; Zhou, H.-B.; et al. Novel Coumarin Derivatives Inhibit the Quorum Sensing System and Iron Homeostasis as Antibacterial Synergists against Pseudomonas aeruginosa. J. Med. Chem. 2023, 66, 14735–14754. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chu, W.; Ye, C.; Gaeta, B.; Tao, H.; Wang, M.; Qiu, Z. Chlorogenic Acid Attenuates Virulence Factors and Pathogenicity of Pseudomonas aeruginosa by Regulating Quorum Sensing. Appl. Microbiol. Biotechnol. 2019, 103, 903–915. [Google Scholar] [CrossRef]
- Naga, N.G.; Zaki, A.A.; El-Badan, D.E.; Rateb, H.S.; Ghanem, K.M.; Shaaban, M.I. Inhibition of Pseudomonas aeruginosa Quorum Sensing by Methyl Gallate from Mangifera Indica. Sci. Rep. 2023, 13, 17942. [Google Scholar] [CrossRef] [PubMed]
- Nortje, N.Q.; Aribisala, J.O.; Pillay, C.; Sabiu, S. Molecular Modelling and Experimental Validation of Mangiferin and Its Related Compounds as Quorum Sensing Modulators of Pseudomonas aeruginosa. Arch. Microbiol. 2025, 207, 53. [Google Scholar] [CrossRef] [PubMed]
- Elmassry, M.M.; Bisht, K.; Colmer-Hamood, J.A.; Wakeman, C.A.; San Francisco, M.J.; Hamood, A.N. Malonate Utilization by Pseudomonas aeruginosa Affects Quorum-Sensing and Virulence and Leads to Formation of Mineralized Biofilm-like Structures. Mol. Microbiol. 2021, 116, 516–537. [Google Scholar] [CrossRef]
- Coelho, P.; Oliveira, J.; Fernandes, I.; Araújo, P.; Pereira, A.R.; Gameiro, P.; Bessa, L.J. Pyranoanthocyanins Interfering with the Quorum Sensing of Pseudomonas aeruginosa and Staphylococcus aureus. Int. J. Mol. Sci. 2021, 22, 8559. [Google Scholar] [CrossRef]
- Sampathkumar, S.J.; Srivastava, P.; Ramachandran, S.; Sivashanmugam, K.; Gothandam, K.M. Lutein: A Potential Antibiofilm and Antiquorum Sensing Molecule from Green Microalga Chlorella Pyrenoidosa. Microb. Pathog. 2019, 135, 103658. [Google Scholar] [CrossRef]
- Han, X.; Nan, M.; Cai, X.; Qiao, B.; Chen, L.; Shen, L. Sennoside A Inhibits Quorum Sensing System to Attenuate Its Regulated Virulence and Pathogenicity via Targeting LasR in Pseudomonas aeruginosa. Front. Microbiol. 2022, 13, 1042214. [Google Scholar] [CrossRef]
- Wang, W.-Q.; Feng, X.-C.; Shi, H.-T.; Wang, Y.-M.; Jiang, C.-Y.; Xiao, Z.-J.; Xu, Y.-J.; Zhang, X.; Yuan, Y.; Ren, N.-Q. Biofilm Inhibition Based on Controlling the Transmembrane Transport and Extracellular Accumulation of Quorum Sensing Signals. Environ. Res. 2023, 221, 115218. [Google Scholar] [CrossRef]
- Zhang, Y.; Bhasme, P.; Reddy, D.S.; Liu, D.; Yu, Z.; Zhao, T.; Zheng, Y.; Kumar, A.; Yu, H.; Ma, L.Z. Dual Functions: A Coumarin–Chalcone Conjugate Inhibits Cyclic-Di-GMP and Quorum-Sensing Signaling to Reduce Biofilm Formation and Virulence of Pathogens. mLife 2023, 2, 283–294. [Google Scholar] [CrossRef]
- Karuppiah, V.; Thirunanasambandham, R.; Thangaraj, G. Anti-Quorum Sensing and Antibiofilm Potential of 1,8-Cineole Derived from Musa Paradisiaca against Pseudomonas aeruginosa Strain PAO1. World J. Microbiol. Biotechnol. 2021, 37, 66. [Google Scholar] [CrossRef] [PubMed]
- Tamfu, A.N.; Ceylan, O.; Cârâc, G.; Talla, E.; Dinica, R.M. Antibiofilm and Anti-Quorum Sensing Potential of Cycloartane-Type Triterpene Acids from Cameroonian Grassland Propolis: Phenolic Profile and Antioxidant Activity of Crude Extract. Molecules 2022, 27, 4872. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, M.; Misra, T.K.; Roy, D.N. In Vitro Anti-Biofilm Activity of 14-Deoxy-11,12-Didehydroandrographolide from Andrographis Paniculata against Pseudomonas aeruginosa. Braz. J. Microbiol. 2020, 51, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Wu, H.; Li, J.-J.; Wang, B.; Jia, A.-Q. Two Cinnamoyl Hydroxamates as Potential Quorum Sensing Inhibitors against Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2024, 14, 1424038. [Google Scholar] [CrossRef]
- Zhang, Z.-S.; He, Z.; Shi, Y.; Guan, M.; Zhao, D.-S.; Zhu, D.; Xiong, L.-T.; Li, Y.; Deng, X.; Cui, Z.-N. Structure-Based Discovery of Symmetric Disulfides from Garlic Extract as Pseudomonas aeruginosa Quorum Sensing Inhibitors. J. Agric. Food. Chem. 2024, 72, 20299–20307. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, Q.; Chen, L.; Duan, K. Virulence-Inhibiting Herbal Compound Falcarindiol Significantly Reduced Mortality in Mice Infected with Pseudomonas aeruginosa. Antibiotics 2020, 9, 136. [Google Scholar] [CrossRef]
- Qais, F.A.; Khan, M.S.; Ahmad, I. Nanoparticles as Quorum Sensing Inhibitor: Prospects and Limitations. In Biotechnological Applications of Quorum Sensing Inhibitors; Kalia, V.C., Ed.; Springer: Singapore, 2018; pp. 227–244. ISBN 978-981-10-9026-4. [Google Scholar]
- Ilk, S.; Tan, G.; Emül, E.; Sağlam, N. Investigation the Potential Use of Silver Nanoparticles Synthesized by Propolis Extract as N-Acyl-Homoserine Lactone-Mediated Quorum Sensing Systems Inhibitor. Turk. J. Med. Sci. 2020, 50, 1147–1156. [Google Scholar] [CrossRef]
- Ferreres, G.; Ivanova, K.; Torrent-Burgués, J.; Tzanov, T. Multimodal Silver-Chitosan-Acylase Nanoparticles Inhibit Bacterial Growth and Biofilm Formation by Gram-Negative Pseudomonas aeruginosa Bacterium. J. Colloid Interface Sci. 2023, 646, 576–586. [Google Scholar] [CrossRef]
- Awadelkareem, A.M.; Al-Shammari, E.; Elkhalifa, A.O.; Adnan, M.; Siddiqui, A.J.; Patel, M.; Khan, M.I.; Mehmood, K.; Ashfaq, F.; Badraoui, R.; et al. Biosynthesized Silver Nanoparticles from Eruca sativa Miller Leaf Extract Exhibits Antibacterial, Antioxidant, Anti-Quorum-Sensing, Antibiofilm, and Anti-Metastatic Activities. Antibiotics 2022, 11, 853. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, X.; Liao, S.; Jiang, C.; Wang, L.; Tang, Y.; Wu, G.; Dai, G.; Chen, L. Quantitative Proteomics Reveals the Mechanism of Silver Nanoparticles against Multidrug-Resistant Pseudomonas aeruginosa Biofilms. J. Proteome Res. 2020, 19, 3109–3122. [Google Scholar] [CrossRef]
- Saeki, E.K.; Yamada, A.Y.; de Araujo, L.A.; Anversa, L.; Garcia, D.d.O.; de Souza, R.L.B.; Martins, H.M.; Kobayashi, R.K.T.; Nakazato, G. Subinhibitory Concentrations of Biogenic Silver Nanoparticles Affect Motility and Biofilm Formation in Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2021, 11, 656984. [Google Scholar] [CrossRef]
- Alipour-Khezri, E.; Moqadami, A.; Barzegar, A.; Mahdavi, M.; Skurnik, M.; Zarrini, G. Bacteriophages and Green Synthesized Zinc Oxide Nanoparticles in Combination Are Efficient against Biofilm Formation of Pseudomonas aeruginosa. Viruses 2024, 16, 897. [Google Scholar] [CrossRef]
- Khan, M.F.; Husain, F.M.; Zia, Q.; Ahmad, E.; Jamal, A.; Alaidarous, M.; Banawas, S.; Alam, M.M.; Alshehri, B.A.; Jameel, M.; et al. Anti-Quorum Sensing and Anti-Biofilm Activity of Zinc Oxide Nanospikes. ACS Omega 2020, 5, 32203–32215. [Google Scholar] [CrossRef] [PubMed]
- Prateeksha; Rao, C.V.; Das, A.K.; Barik, S.K.; Singh, B.N. ZnO/Curcumin Nanocomposites for Enhanced Inhibition of Pseudomonas aeruginosa Virulence via LasR-RhlR Quorum Sensing Systems. Mol. Pharm. 2019, 16, 3399–3413. [Google Scholar] [CrossRef]
- Kanak, K.R.; Dass, R.S.; Pan, A. Anti-Quorum Sensing Potential of Selenium Nanoparticles against LasI/R, RhlI/R, and PQS/MvfR in Pseudomonas aeruginosa: A Molecular Docking Approach. Front. Mol. Biosci. 2023, 10, 1203672. [Google Scholar] [CrossRef]
- Gómez-Gómez, B.; Arregui, L.; Serrano, S.; Santos, A.; Pérez-Corona, T.; Madrid, Y. Selenium and Tellurium-Based Nanoparticles as Interfering Factors in Quorum Sensing-Regulated Processes: Violacein Production and Bacterial Biofilm Formation. Metallomics 2019, 11, 1104–1114. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.Y.; Aly, U.F.; Abd El-Baky, R.M.; Waly, N.G.F.M. Effect of Titanium Dioxide Nanoparticles on the Expression of Efflux Pump and Quorum-Sensing Genes in MDR Pseudomonas aeruginosa Isolates. Antibiotics 2021, 10, 625. [Google Scholar] [CrossRef] [PubMed]
- Sarif, M.; Jegel, O.; Gazanis, A.; Hartmann, J.; Plana-Ruiz, S.; Hilgert, J.; Frerichs, H.; Viel, M.; Panthöfer, M.; Kolb, U.; et al. High-Throughput Synthesis of CeO2 Nanoparticles for Transparent Nanocomposites Repelling Pseudomonas aeruginosa Biofilms. Sci. Rep. 2022, 12, 3935. [Google Scholar] [CrossRef]
- Mohammed, H.B.; Rayyif, S.M.I.; Curutiu, C.; Birca, A.C.; Oprea, O.-C.; Grumezescu, A.M.; Ditu, L.-M.; Gheorghe, I.; Chifiriuc, M.C.; Mihaescu, G.; et al. Eugenol-Functionalized Magnetite Nanoparticles Modulate Virulence and Persistence in Pseudomonas aeruginosa Clinical Strains. Molecules 2021, 26, 2189. [Google Scholar] [CrossRef]
- Chen, H.; Hu, P.; Wang, Y.; Liu, H.; Zheng, J.; Huang, Z.; Zhang, X.; Liu, Y.; Zhou, T. From Quorum Sensing Inhibition to Antimicrobial Defense: The Dual Role of Eugenol-Gold Nanoparticles against Carbapenem-Resistant Pseudomonas aeruginosa. Colloids Surf. B Biointerfaces 2025, 247, 114415. [Google Scholar] [CrossRef]
- Eltayb, E.K.; Alqahtani, F.Y.; Alkahtani, H.M.; Alsarra, I.A.; Alfaraj, R.; Aleanizy, F.S. Attenuation of Pseudomonas aeruginosa Quorum Sensing Virulence of Biofilm and Pyocyanin by mBTL-Loaded Calcium Alginate Nanoparticles. Polymers 2022, 14, 3655. [Google Scholar] [CrossRef]
- Ulusoy, S.; Akalin, R.B.; Çevikbaş, H.; Berisha, A.; Oral, A.; Boşgelmez-Tinaz, G. Zeolite 4A as a Jammer of Bacterial Communication in Chromobacterium violaceum and Pseudomonas aeruginosa. Future Microbiol. 2022, 17, 861–871. [Google Scholar] [CrossRef] [PubMed]
- Husain, F.M.; Ansari, A.A.; Khan, A.; Ahmad, N.; Albadri, A.; Albalawi, T.H. Mitigation of Acyl-Homoserine Lactone (AHL) Based Bacterial Quorum Sensing, Virulence Functions, and Biofilm Formation by Yttrium Oxide Core/Shell Nanospheres: Novel Approach to Combat Drug Resistance. Sci. Rep. 2019, 9, 18476. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Rajwade, J.; Paknikar, K.M. Transcriptome Analysis of Silver Nanoparticles Treated Staphylococcus aureus Reveals Potential Targets for Biofilm Inhibition. Colloids Surf. B Biointerfaces 2019, 175, 487–497. [Google Scholar] [CrossRef]
- Malarvizhi, K.; Ramyadevi, D.; Vedha Hari, B.N.; Sarveswari, H.B.; Solomon, A.P.; Fang, H.; Luo, R.H.; Zheng, Y.T. Mercuric-Sulphide Based Metallopharmaceutical Formulation as an Alternative Therapeutic to Combat Viral and Multidrug-Resistant (MDR) Bacterial Infections. Sci. Rep. 2023, 13, 16706. [Google Scholar] [CrossRef]
- Sun, X.; Hill, P.; Liu, J.; Qian, J.; Ma, Y.; Zhou, S. Marine-Source Quorum Quenching Enzyme YtnP to Improve Hygiene Quality in Dental Units. Mar. Drugs 2021, 19, 225. [Google Scholar] [CrossRef] [PubMed]
- López-Jácome, L.E.; Garza-Ramos, G.; Hernández-Durán, M.; Franco-Cendejas, R.; Loarca, D.; Romero-Martínez, D.; Nguyen, P.T.D.; Maeda, T.; González-Pedrajo, B.; Díaz-Guerrero, M.; et al. AiiM Lactonase Strongly Reduces Quorum Sensing Controlled Virulence Factors in Clinical Strains of Pseudomonas aeruginosa Isolated From Burned Patients. Front. Microbiol. 2019, 10, 2657. [Google Scholar] [CrossRef]
- Anandan, K.; Vittal, R.R. Quorum Quenching Activity of AiiA Lactonase KMMI17 from Endophytic Bacillus Thuringiensis KMCL07 on AHL- Mediated Pathogenic Phenotype in Pseudomonas aeruginosa. Microb. Pathog. 2019, 132, 230–242. [Google Scholar] [CrossRef]
- Zhang, J.-W.; Xuan, C.-G.; Lu, C.-H.; Guo, S.; Yu, J.-F.; Asif, M.; Jiang, W.-J.; Zhou, Z.-G.; Luo, Z.-Q.; Zhang, L.-Q. AidB, a Novel Thermostable N-Acylhomoserine Lactonase from the Bacterium bosea sp. Appl. Environ. Microbiol. 2019, 85, e02065-19. [Google Scholar] [CrossRef]
- Rémy, B.; Plener, L.; Decloquement, P.; Armstrong, N.; Elias, M.; Daudé, D.; Chabrière, É. Lactonase Specificity Is Key to Quorum Quenching in Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 76. [Google Scholar] [CrossRef]
- Guendouze, A.; Plener, L.; Bzdrenga, J.; Jacquet, P.; Rémy, B.; Elias, M.; Lavigne, J.-P.; Daudé, D.; Chabrière, E. Effect of Quorum Quenching Lactonase in Clinical Isolates of Pseudomonas aeruginosa and Comparison with Quorum Sensing Inhibitors. Front. Microbiol 2017, 8, 227. [Google Scholar] [CrossRef]
- Zhang, Y.; He, X.; Cheng, P.; Li, X.; Wang, S.; Xiong, J.; Li, H.; Wang, Z.; Yi, H.; Du, H.; et al. Effects of a Novel Anti-Biofilm Peptide CRAMP Combined with Antibiotics on the Formation of Pseudomonas aeruginosa Biofilms. Microb. Pathog. 2021, 152, 104660. [Google Scholar] [CrossRef] [PubMed]
- Aflakian, F.; Rad, M.; Hashemitabar, G.; Lagzian, M.; Ramezani, M. Design and Assessment of Novel Synthetic Peptides to Inhibit Quorum Sensing-Dependent Biofilm Formation in Pseudomonas aeruginosa. Biofouling 2022, 38, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Schroven, K.; Putzeys, L.; Swinnen, A.-L.; Hendrix, H.; Paeshuyse, J.; Lavigne, R. The Phage-Encoded Protein PIT2 Impacts Pseudomonas aeruginosa Quorum Sensing by Direct Interaction with LasR. iScience 2023, 26, 107745. [Google Scholar] [CrossRef]
- Shah, M.; Taylor, V.L.; Bona, D.; Tsao, Y.; Stanley, S.Y.; Pimentel-Elardo, S.M.; McCallum, M.; Bondy-Denomy, J.; Howell, P.L.; Nodwell, J.R.; et al. A Phage-Encoded Anti-Activator Inhibits Quorum Sensing in Pseudomonas aeruginosa. Mol. Cell 2021, 81, 571–583.e6. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xu, Z.; Cao, R.; Li, J.; Wu, C.-J.; Wang, Y.; Zhu, H. Effects of Hydroxyl Group in Cyclo(Pro-Tyr)-like Cyclic Dipeptides on Their Anti-QS Activity and Self-Assembly. iScience 2023, 26, 107048. [Google Scholar] [CrossRef]
- Kiran, G.S.; Sajayan, A.; Priyadharshini, G.; Balakrishnan, A.; Prathiviraj, R.; Sabu, A.; Selvin, J. A Novel Anti-Infective Molecule Nesfactin Identified from Sponge Associated Bacteria Nesterenkonia sp. MSA31 against Multidrug Resistant Pseudomonas aeruginosa. Microb. Pathog. 2021, 157, 104923. [Google Scholar] [CrossRef]
- Oh, J.W.; Shin, M.K.; Park, H.-R.; Kim, S.; Lee, B.; Yoo, J.S.; Chi, W.-J.; Sung, J.-S. PA-Win2: In Silico-Based Discovery of a Novel Peptide with Dual Antibacterial and Anti-Biofilm Activity. Antibiotics 2024, 13, 1113. [Google Scholar] [CrossRef]
- Jafari, P.; Luscher, A.; Siriwardena, T.; Michetti, M.; Que, Y.-A.; Rahme, L.G.; Reymond, J.-L.; Raffoul, W.; Van Delden, C.; Applegate, L.A.; et al. Antimicrobial Peptide Dendrimers and Quorum-Sensing Inhibitors in Formulating Next-Generation Anti-Infection Cell Therapy Dressings for Burns. Molecules 2021, 26, 3839. [Google Scholar] [CrossRef]
- Li, W.; Xiao, X.; Qi, Y.; Lin, X.; Hu, H.; Shi, M.; Zhou, M.; Jiang, W.; Liu, L.; Chen, K.; et al. Host-Defense-Peptide-Mimicking β-Peptide Polymer Acting as a Dual-Modal Antibacterial Agent by Interfering Quorum Sensing and Killing Individual Bacteria Simultaneously. Research 2023, 6, 0051. [Google Scholar] [CrossRef]
- Hijazi, D.M.; Dahabiyeh, L.A.; Abdelrazig, S.; Alqudah, D.A.; Al-Bakri, A.G. Micafungin Effect on Pseudomonas aeruginosa Metabolome, Virulence and Biofilm: Potential Quorum Sensing Inhibitor. AMB Express 2023, 13, 20. [Google Scholar] [CrossRef]
- Fu, T.-K.; Ng, S.-K.; Chen, Y.-E.; Lee, Y.-C.; Demeter, F.; Herczeg, M.; Borbás, A.; Chiu, C.-H.; Lan, C.-Y.; Chen, C.-L.; et al. Rhamnose Binding Protein as an Anti-Bacterial Agent-Targeting Biofilm of Pseudomonas aeruginosa. Mar. Drugs 2019, 17, 355. [Google Scholar] [CrossRef]
- Gasu, E.N.; Ahor, H.S.; Borquaye, L.S. Peptide Mix from Olivancillaria Hiatula Interferes with Cell-to-Cell Communication in Pseudomonas aeruginosa. Biomed Res. Int. 2019, 2019, 5313918. [Google Scholar] [CrossRef]
- Abbas, S.; Mujawar, S.; Yasmin, A.; Perveen, S.; Malik, P.A.; Gill, M.S.A.; Chohan, T.A. Targeting Quorum Sensing in Pseudomonas aeruginosa with High-Affinity Inhibitors: A High-Throughput Screening and in-Silico Analysis. Comput. Biol. Chem. 2025, 117, 108419. [Google Scholar] [CrossRef]
- Szemerédi, N.; Kincses, A.; Rehorova, K.; Hoang, L.; Salardón-Jiménez, N.; Sevilla-Hernández, C.; Viktorová, J.; Domínguez-Álvarez, E.; Spengler, G. Ketone- and Cyano-Selenoesters to Overcome Efflux Pump, Quorum-Sensing, and Biofilm-Mediated Resistance. Antibiotics 2020, 9, 896. [Google Scholar] [CrossRef]
- Dadkhah, F.; Moniri, F.; Zamani, H.; Hadavi, M. Anti-Virulence Effects of Diclofenac Sodium in Combination with Gentamicin on Pseudomonas aeruginosa: Attenuation of Quorum Sensing Related Traits and Efflux Pump Systems. Braz. J. Microbiol. 2025. [Google Scholar] [CrossRef] [PubMed]
- Rajkumari, J.; Borkotoky, S.; Reddy, D.; Mohanty, S.K.; Kumavath, R.; Murali, A.; Suchiang, K.; Busi, S. Anti-Quorum Sensing and Anti-Biofilm Activity of 5-Hydroxymethylfurfural against Pseudomonas aeruginosa PAO1: Insights from In Vitro, In Vivo and In Silico Studies. Microbiol. Res. 2019, 226, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Elsheredy, A.; El-Soudany, I.; Elsherbini, E.; Metwally, D.; Ghazal, A. Effect of Azithromycin and Phenylalanine-Arginine Beta-Naphthylamide on Quorum Sensing and Virulence Factors in Clinical Isolates of Pseudomonas aeruginosa. Iran. J. Microbiol. 2021, 13, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Valastyan, J.S.; Shine, E.E.; Mook, R.A.; Bassler, B.L. Inhibitors of the PqsR Quorum-Sensing Receptor Reveal Differential Roles for PqsE and RhlI in Control of Phenazine Production. bioRxiv 2025. bioRxiv:2025.02.10.637488. [Google Scholar] [CrossRef]
- Kumar, L.; Brenner, N.; Brice, J.; Klein-Seetharaman, J.; Sarkar, S.K. Cephalosporins Interfere With Quorum Sensing and Improve the Ability of Caenorhabditis elegans to Survive Pseudomonas aeruginosa Infection. Front. Microbiol. 2021, 12, 598498. [Google Scholar] [CrossRef]
- Yan, X.; Hou, S.; Xing, C.; Zhang, Y.; Chang, J.; Xiao, J.; Lin, F. Design, Synthesis, and Biological Evaluation of the Quorum-Sensing Inhibitors of Pseudomonas aeruginosa PAO1. Molecules 2024, 29, 2211. [Google Scholar] [CrossRef]
- Alyahyawy, O.Y.; Munshi, R.M.; Badr-Eldin, S.M.; Aldawsari, H.M.; Abualsunun, W.; Abbas, H.A.; Salem, I.M.; Hegazy, W.A.H.; Nazeih, S.I. Reprofiling Lamivudine as an Antibiofilm and Anti-Pathogenic Agent against Pseudomonas aeruginosa. AMB Express 2025, 15, 33. [Google Scholar] [CrossRef]
- Kai, T.; Tateda, K.; Kimura, S.; Ishii, Y.; Ito, H.; Yoshida, H.; Kimura, T.; Yamaguchi, K. A Low Concentration of Azithromycin Inhibits the mRNA Expression of N-Acyl Homoserine Lactone Synthesis Enzymes, Upstream of lasI or rhlI, in Pseudomonas aeruginosa. Pulm. Pharmacol. Ther. 2009, 22, 483–486. [Google Scholar] [CrossRef] [PubMed]
- Apostolaki, K.; Gagaletsios, L.A.; Papagiannitsis, C.C.; Petinaki, E. Macrolides Impact the Growth Ability of Clinical Pseudomonas aeruginosa through Quorum-Sensing Systems. J. Chemother. 2024, 36, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhao, K.; Li, H.; Liu, K.; Li, J.; Chu, Y.; Prithiviraj, B.; Yue, B.; Zhang, X. Antibacterial and Anti-Virulence Effects of Furazolidone on Trueperella Pyogenes and Pseudomonas aeruginosa. BMC Vet. Res. 2022, 18, 114. [Google Scholar] [CrossRef]
- Yang, N.; Li, H.; Yang, X.; Wu, Y.; Lv, Z.; Zhang, Z.; Ma, X.; Zhou, X.; Zhang, X.; Zhao, K.; et al. Furazolidone Reduces the Pathogenesis of Trueperella Pyogenes and Pseudomonas aeruginosa Co-Infection in a Mouse Model. Heliyon 2024, 10, e39629. [Google Scholar] [CrossRef] [PubMed]
- Baldelli, V.; D’Angelo, F.; Pavoncello, V.; Fiscarelli, E.V.; Visca, P.; Rampioni, G.; Leoni, L. Identification of FDA-Approved Antivirulence Drugs Targeting the Pseudomonas aeruginosa Quorum Sensing Effector Protein PqsE. Virulence 2020, 11, 652–668. [Google Scholar] [CrossRef] [PubMed]
- Seleem, N.M.; Abd El Latif, H.K.; Shaldam, M.A.; El-Ganiny, A. Drugs with New Lease of Life as Quorum Sensing Inhibitors: For Combating MDR Acinetobacter Baumannii Infections. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1687–1702. [Google Scholar] [CrossRef]
- Yin, L.; Shen, W.; Liu, J.-S.; Jia, A.-Q. 2-Hydroxymethyl-1-Methyl-5-Nitroimidazole, One Siderophore Inhibitor, Occludes Quorum Sensing in Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2022, 12, 955952. [Google Scholar] [CrossRef]
- Collalto, D.; Giallonardi, G.; Fortuna, A.; Meneghini, C.; Fiscarelli, E.; Visca, P.; Imperi, F.; Rampioni, G.; Leoni, L. In Vitro Activity of Antivirulence Drugs Targeting the Las or Pqs Quorum Sensing Against Cystic Fibrosis Pseudomonas aeruginosa Isolates. Front. Microbiol. 2022, 13, 845231. [Google Scholar] [CrossRef]
- Markus, V.; Golberg, K.; Teralı, K.; Ozer, N.; Kramarsky-Winter, E.; Marks, R.S.; Kushmaro, A. Assessing the Molecular Targets and Mode of Action of Furanone C-30 on Pseudomonas aeruginosa Quorum Sensing. Molecules 2021, 26, 1620. [Google Scholar] [CrossRef]
- Muñoz-Cázares, N.; Castillo-Juárez, I.; García-Contreras, R.; Castro-Torres, V.A.; Díaz-Guerrero, M.; Rodríguez-Zavala, J.S.; Quezada, H.; González-Pedrajo, B.; Martínez-Vázquez, M. A Brominated Furanone Inhibits Pseudomonas aeruginosa Quorum Sensing and Type III Secretion, Attenuating Its Virulence in a Murine Cutaneous Abscess Model. Biomedicines 2022, 10, 1847. [Google Scholar] [CrossRef] [PubMed]
- Mensah, C.N.; Ampomah, G.B.; Mensah, J.O.; Gasu, E.N.; Aboagye, C.I.; Ekuadzi, E.; Boadi, N.O.; Borquaye, L.S. N-Alkylimidazole Derivatives as Potential Inhibitors of Quorum Sensing in Pseudomonas aeruginosa. Heliyon 2022, 8, e12581. [Google Scholar] [CrossRef]
- Dai, L.; Wu, T.-Q.; Xiong, Y.-S.; Ni, H.-B.; Ding, Y.; Zhang, W.-C.; Chu, S.-P.; Ju, S.-Q.; Yu, J. Ibuprofen-Mediated Potential Inhibition of Biofilm Development and Quorum Sensing in Pseudomonas aeruginosa. Life Sci. 2019, 237, 116947. [Google Scholar] [CrossRef]
- Hwang, H.-J.; Choi, H.; Hong, S.; Moon, H.R.; Lee, J.-H. Antipathogenic Compounds That Are Effective at Very Low Concentrations and Have Both Antibiofilm and Antivirulence Effects against Pseudomonas aeruginosa. Microbiol. Spectr. 2021, 9, e0024921. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.; Ham, S.-Y.; Kwon, H.; Kim, H.-S.; Moon, S.; Lee, J.-H.; Lim, T.; Son, S.-H.; Park, H.-D.; Byun, Y. Discovery and Characterization of Pure RhlR Antagonists against Pseudomonas aeruginosa Infections. J. Med. Chem. 2020, 63, 8388–8407. [Google Scholar] [CrossRef]
- Yuan, Y.; Yang, X.; Zeng, Q.; Li, H.; Fu, R.; Du, L.; Liu, W.; Zhang, Y.; Zhou, X.; Chu, Y.; et al. Repurposing Dimetridazole and Ribavirin to Disarm Pseudomonas aeruginosa Virulence by Targeting the Quorum Sensing System. Front. Microbiol. 2022, 13, 978502. [Google Scholar] [CrossRef] [PubMed]
- Sabir, S.; Suresh, D.; Subramoni, S.; Das, T.; Bhadbhade, M.; Black, D.S.; Rice, S.A.; Kumar, N. Thioether-Linked Dihydropyrrol-2-One Analogues as PqsR Antagonists against Antibiotic Resistant Pseudomonas aeruginosa. Bioorg Med. Chem. 2021, 31, 115967. [Google Scholar] [CrossRef]
- Soukarieh, F.; Mashabi, A.; Richardson, W.; Oton, E.V.; Romero, M.; Roberston, S.N.; Grossman, S.; Sou, T.; Liu, R.; Halliday, N.; et al. Design and Evaluation of New Quinazolin-4(3H)-One Derived PqsR Antagonists as Quorum Sensing Quenchers in Pseudomonas aeruginosa. ACS Infect. Dis. 2021, 7, 2666–2685. [Google Scholar] [CrossRef]
- Grossman, S.; Soukarieh, F.; Richardson, W.; Liu, R.; Mashabi, A.; Emsley, J.; Williams, P.; Cámara, M.; Stocks, M.J. Novel Quinazolinone Inhibitors of the Pseudomonas aeruginosa Quorum Sensing Transcriptional Regulator PqsR. Eur. J. Med. Chem. 2020, 208, 112778. [Google Scholar] [CrossRef]
- Huang, X.-H.; She, M.-T.; Zhang, Y.-H.; Liu, Y.-F.; Zhong, D.-X.; Zhang, Y.-H.; Zheng, J.-X.; Sun, N.; Wong, W.-L.; Lu, Y.-J. Novel Quinoline-Based Derivatives as the PqsR Inhibitor against Pseudomonas aeruginosa PAO1. J. Appl. Microbiol. 2022, 133, 2167–2181. [Google Scholar] [CrossRef] [PubMed]
- Abdelsamie, A.S.; Hamed, M.M.; Schütz, C.; Röhrig, T.; Kany, A.M.; Schmelz, S.; Blankenfeldt, W.; Hirsch, A.K.H.; Hartmann, R.W.; Empting, M. Discovery and Optimization of Thiazole-Based Quorum Sensing Inhibitors as Potent Blockers of Pseudomonas aeruginosa Pathogenicity. Eur. J. Med. Chem. 2024, 276, 116685. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Chen, W.; Ma, Y.-C.; Bai, B.; Sun, G.; Zhang, S.; Chang, X.; Wang, Y.; Jiang, N.; Zhang, X.; et al. Design and Synthesis of 4-Fluorophenyl-5-Methylene-2(5H)-Furanone Derivatives as Potent Quorum Sensing Inhibitors. J. Med. Chem. 2023, 66, 8441–8463. [Google Scholar] [CrossRef] [PubMed]
- Abo-Dya, N.E.; Agha, K.A.; Abbas, H.A.; Abu-Kull, M.E.; Alahmdi, M.I.; Osman, N.A. Hybrid N-Acylcysteines as Dual-Acting Matrix Disruptive and Anti-Quorum Sensing Agents Fighting Pseudomonas aeruginosa Biofilms: Design, Synthesis, Molecular Docking Studies, and In Vitro Assays. ACS Omega 2022, 7, 19879–19891. [Google Scholar] [CrossRef]
- Alzahrani, A.Y.; Ammar, Y.A.; Abu-Elghait, M.; Salem, M.A.; Assiri, M.A.; Ali, T.E.; Ragab, A. Development of Novel Indolin-2-One Derivative Incorporating Thiazole Moiety as DHFR and Quorum Sensing Inhibitors: Synthesis, Antimicrobial, and Antibiofilm Activities with Molecular Modelling Study. Bioorg Chem. 2022, 119, 105571. [Google Scholar] [CrossRef]
- Soukarieh, F.; Gurnani, P.; Romero, M.; Halliday, N.; Stocks, M.; Alexander, C.; Cámara, M. Design of Quorum Sensing Inhibitor-Polymer Conjugates to Penetrate Pseudomonas aeruginosa Biofilms. ACS Macro Lett. 2023, 12, 314–319. [Google Scholar] [CrossRef]
- Tang, D.; Lin, Y.; Yao, H.; Liu, Y.; Xi, Y.; Li, M.; Mao, A. Effect of L-HSL on Biofilm and Motility of Pseudomonas aeruginosa and Its Mechanism. Appl. Microbiol. Biotechnol. 2024, 108, 418. [Google Scholar] [CrossRef]
- Liu, J.; Meng, Y.; Yang, M.-H.; Zhang, X.-Y.; Zhao, J.-F.; Sun, P.-H.; Chen, W.-M. Design, Synthesis and Biological Evaluation of Novel 3-Hydroxypyridin-4(1H)-Ones Based Hybrids as Pseudomonas aeruginosa Biofilm Inhibitors. Eur. J. Med. Chem. 2023, 259, 115665. [Google Scholar] [CrossRef]
- She, P.; Zhou, L.; Li, S.; Zeng, X.; Wu, Y. Inhibitory Effects of 1,3-Diaminopropane on the Biofilm Formation of Pseudomonas aeruginosa via Interaction with Quorum Sensing System. J. Cent. South Univ. Med. Sci. 2021, 46, 942–948. [Google Scholar] [CrossRef]
- Liu, J.; Hou, J.-S.; Li, Y.-B.; Miao, Z.-Y.; Sun, P.-H.; Lin, J.; Chen, W.-M. Novel 2-Substituted 3-Hydroxy-1,6-Dimethylpyridin-4(1H)-Ones as Dual-Acting Biofilm Inhibitors of Pseudomonas aeruginosa. J. Med. Chem. 2020, 63, 10921–10945. [Google Scholar] [CrossRef]
- Vieira, T.F.; Leitão, M.M.; Cerqueira, N.M.F.S.A.; Sousa, S.F.; Borges, A.; Simões, M. Montelukast and Cefoperazone Act as Antiquorum Sensing and Antibiofilm Agents against Pseudomonas aeruginosa. J. Appl. Microbiol. 2024, 135, lxae088. [Google Scholar] [CrossRef]
- Xiao, H.; Li, J.; Yang, D.; Du, J.; Li, J.; Lin, S.; Zhou, H.; Sun, P.; Xu, J. Multidimensional Criteria for Virtual Screening of PqsR Inhibitors Based on Pharmacophore, Docking, and Molecular Dynamics. Int. J. Mol. Sci. 2024, 25, 1869. [Google Scholar] [CrossRef]
- Laborda, P.; Alcalde-Rico, M.; Chini, A.; Martínez, J.L.; Hernando-Amado, S. Discovery of Inhibitors of Pseudomonas aeruginosa Virulence through the Search for Natural-like Compounds with a Dual Role as Inducers and Substrates of Efflux Pumps. Environ. Microbiol. 2021, 23, 7396–7411. [Google Scholar] [CrossRef] [PubMed]
- Tajani, A.S.; Jangi, E.; Davodi, M.; Golmakaniyoon, S.; Ghodsi, R.; Soheili, V.; Fazly Bazzaz, B.S. Anti-Quorum Sensing Potential of Ketoprofen and Its Derivatives against Pseudomonas aeruginosa: Insights to In Silico and In Vitro Studies. Arch. Microbiol. 2021, 203, 5123–5132. [Google Scholar] [CrossRef] [PubMed]
- Soukarieh, F.; Mashabi, A.; Richardson, W.; Oton, E.V.; Romero, M.; Dubern, J.-F.; Robertson, S.N.; Lucanto, S.; Markham-Lee, Z.; Sou, T.; et al. Design, Synthesis, and Evaluation of New 1H-Benzo[d]Imidazole Based PqsR Inhibitors as Adjuvant Therapy for Pseudomonas aeruginosa Infections. J. Med. Chem. 2024, 67, 1008–1023. [Google Scholar] [CrossRef]
- Nketia, P.B.; Manu, P.; Osei-Poku, P.; Kwarteng, A. Phenazine Scaffolds as a Potential Allosteric Inhibitor of LasR Protein in Pseudomonas aeruginosa. Bioinform. Biol. Insights 2025, 19, 11779322251319594. [Google Scholar] [CrossRef]
- Mao, S.; Li, Q.; Yang, Z.; Li, Y.; Ye, X.; Wang, H. Design, Synthesis, and Biological Evaluation of Benzoheterocyclic Sulfoxide Derivatives as Quorum Sensing Inhibitors in Pseudomonas aeruginosa. J. Enzyme. Inhib. Med. Chem. 2023, 38, 2175820. [Google Scholar] [CrossRef]
- Almalki, A.J.; Ibrahim, T.S.; Taher, E.S.; Mohamed, M.F.A.; Youns, M.; Hegazy, W.A.H.; Al-Mahmoudy, A.M.M. Synthesis, Antimicrobial, Anti-Virulence and Anticancer Evaluation of New 5(4H)-Oxazolone-Based Sulfonamides. Molecules 2022, 27, 671. [Google Scholar] [CrossRef] [PubMed]
- Alotaibi, H.F.; Alotaibi, H.; Darwish, K.M.; Khafagy, E.-S.; Abu Lila, A.S.; Ali, M.A.M.; Hegazy, W.A.H.; Alshawwa, S.Z. The Anti-Virulence Activities of the Antihypertensive Drug Propranolol in Light of Its Anti-Quorum Sensing Effects against Pseudomonas aeruginosa and Serratia Marcescens. Biomedicines 2023, 11, 3161. [Google Scholar] [CrossRef]
- Khayat, M.T.; Abbas, H.A.; Ibrahim, T.S.; Khayyat, A.N.; Alharbi, M.; Darwish, K.M.; Elhady, S.S.; Khafagy, E.-S.; Safo, M.K.; Hegazy, W.A.H. Anti-Quorum Sensing Activities of Gliptins against Pseudomonas aeruginosa and Staphylococcus aureus. Biomedicines 2022, 10, 1169. [Google Scholar] [CrossRef]
- Abbas, H.A.; Shaldam, M.A.; Eldamasi, D. Curtailing Quorum Sensing in Pseudomonas aeruginosa by Sitagliptin. Curr. Microbiol. 2020, 77, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Saqr, A.A.; Aldawsari, M.F.; Khafagy, E.-S.; Shaldam, M.A.; Hegazy, W.A.H.; Abbas, H.A. A Novel Use of Allopurinol as A Quorum-Sensing Inhibitor in Pseudomonas aeruginosa. Antibiotics 2021, 10, 1385. [Google Scholar] [CrossRef] [PubMed]
- Al-Rabia, M.W.; Asfour, H.Z.; Alhakamy, N.A.; Bazuhair, M.A.; Ibrahim, T.S.; Abbas, H.A.; Mansour, B.; Hegazy, W.A.H.; Seleem, N.M. Cilostazol Is a Promising Anti-Pseudomonal Virulence Drug by Disruption of Quorum Sensing. AMB Express 2024, 14, 87. [Google Scholar] [CrossRef]
- Saleh, M.M.; Abbas, H.A.; Askoura, M.M. Repositioning Secnidazole as a Novel Virulence Factors Attenuating Agent in Pseudomonas aeruginosa. Microb. Pathog. 2019, 127, 31–38. [Google Scholar] [CrossRef]
- Dong, L.; Sun, L.; Hu, X.; Nie, T.; Pang, J.; Wang, X.; Yang, X.; Li, C.; Yao, K.; Zhang, Y.; et al. Ostarine Attenuates Pyocyanin in Pseudomonas aeruginosa by Interfering with Quorum Sensing Systems. J. Antibiot. 2021, 74, 863–873. [Google Scholar] [CrossRef]
- Seleem, N.M.; Atallah, H.; Abd El Latif, H.K.; Shaldam, M.A.; El-Ganiny, A.M. Could the Analgesic Drugs, Paracetamol and Indomethacin, Function as Quorum Sensing Inhibitors? Microb. Pathog. 2021, 158, 105097. [Google Scholar] [CrossRef]
- Sharma, N.; Srivastava, N.; Devi, B.; Kumar, L.; Kumar, R.; Kumar Yadav, A. Synthesis, Biological Evaluation and In Silico Study of N-(2- and 3-Pyridinyl)Benzamide Derivatives as Quorum Sensing Inhibitors against Pseudomonas aeruginosa. Chem. Biodivers. 2023, 20, e202201191. [Google Scholar] [CrossRef]
- Siqueira, F.D.S.; Alves, C.F.D.S.; Machado, A.K.; Siqueira, J.D.; Santos, T.D.; Mizdal, C.R.; Moreira, K.S.; Teixeira Carvalho, D.; Bonez, P.C.; Urquhart, C.G.; et al. Molecular Docking, Quorum Quenching Effect, Antibiofilm Activity and Safety Profile of Silver-Complexed Sulfonamide on Pseudomonas aeruginosa. Biofouling 2021, 37, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Naga, N.G.; El-Badan, D.E.; Mabrouk, M.E.M.; Rateb, H.S.; Ghanem, K.M.; Shaaban, M.I. Innovative Application of Ceftriaxone as a Quorum Sensing Inhibitor in Pseudomonas aeruginosa. Sci. Rep. 2025, 15, 5022. [Google Scholar] [CrossRef]
- Zhang, P.; Ma, Y.; Wang, Y.; Dong, E.; Ma, S. Design, Synthesis, and Biological Evaluation of 2-Phenoxyalkylhydrazide Benzoxazole Derivatives as Quorum Sensing Inhibitors with Strong Antibiofilm Effect. J. Med. Chem. 2024, 67, 5721–5743. [Google Scholar] [CrossRef]
- Arami, N.; Tajani, A.S.; Hashemi, M.; Rezaei, T.; Ghodsi, R.; Soheili, V.; Bazzaz, B.S.F. Targeted Inhibition of PqsR in Pseudomonas aeruginosa PAO1 Quorum-Sensing Network by Chalcones as Promising Antibacterial Compounds. Mol. Biol. Rep. 2025, 52, 175. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.A.K.S.; Rudden, M.; Smyth, T.J.; Dooley, J.S.G.; Marchant, R.; Banat, I.M. Natural Quorum Sensing Inhibitors Effectively Downregulate Gene Expression of Pseudomonas aeruginosa Virulence Factors. Appl. Microbiol. Biotechnol. 2019, 103, 3521–3535. [Google Scholar] [CrossRef] [PubMed]
- Leitão, M.M.; Vieira, T.F.; Sousa, S.F.; Borges, F.; Simões, M.; Borges, A. Dual Action of Benzaldehydes: Inhibiting Quorum Sensing and Enhancing Antibiotic Efficacy for Controlling Pseudomonas aeruginosa Biofilms. Microb. Pathog. 2024, 191, 106663. [Google Scholar] [CrossRef]
- Fakhoury, N.E.; Mansour, S.; Abdel-Halim, M.; Hamed, M.M.; Empting, M.; Boese, A.; Loretz, B.; Lehr, C.-M.; Tammam, S.N. Nanoparticles in Liposomes: A Platform for Increased Antibiotic Selectivity in Multidrug Resistant Bacteria in Respiratory Tract Infections. Drug Deliv. Transl. Res. 2025, 15, 1193–1209. [Google Scholar] [CrossRef]
- Zhao, Y.; Mei, L.; Si, Y.; Wu, J.; Shao, J.; Wang, T.; Yan, G.; Wang, C.; Wu, D. Sodium New Houttuyfonate Affects Transcriptome and Virulence Factors of Pseudomonas aeruginosa Controlled by Quorum Sensing. Front. Pharmacol. 2020, 11, 572375. [Google Scholar] [CrossRef]
- Fong, J.; Mortensen, K.T.; Nørskov, A.; Qvortrup, K.; Yang, L.; Tan, C.H.; Nielsen, T.E.; Givskov, M. Itaconimides as Novel Quorum Sensing Inhibitors of Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 2018, 8, 443. [Google Scholar] [CrossRef]
- Xie, Y.; Chen, J.; Wang, B.; Peng, A.-Y.; Mao, Z.-W.; Xia, W. Inhibition of Quorum-Sensing Regulator from Pseudomonas aeruginosa Using a Flavone Derivative. Molecules 2022, 27, 2439. [Google Scholar] [CrossRef]
- Ye, X.; Mao, S.; Li, Y.; Yang, Z.; Du, A.; Wang, H. Design, Synthesis, and Biological Evaluation of Phenyloxadiazole Sulfoxide Derivatives as Potent Pseudomonas aeruginosa Biofilm Inhibitors. Molecules 2023, 28, 3879. [Google Scholar] [CrossRef] [PubMed]
- Okada, B.K.; Li, A.; Seyedsayamdost, M.R. Identification of the Hypertension Drug Guanfacine as an Antivirulence Agent in Pseudomonas aeruginosa. Chembiochem 2019, 20, 2005–2011. [Google Scholar] [CrossRef]
- Trognon, J.; Vera, G.; Rima, M.; Stigliani, J.-L.; Amielet, L.; El Hage, S.; Lajoie, B.; Roques, C.; El Garah, F. Investigation of Direct and Retro Chromone-2-Carboxamides Based Analogs of Pseudomonas aeruginosa Quorum Sensing Signal as New Anti-Biofilm Agents. Pharmaceuticals 2022, 15, 417. [Google Scholar] [CrossRef]
- Gajera, G.; Henriksen, N.; Cox, B.; Kothari, V. Identification of Anti-Pathogenic Activity Among In Silico Predicted Small-Molecule Inhibitors of Pseudomonas aeruginosa LasR or Nitric Oxide Reductase (NOR). Drug Target Insights 2023, 17, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, X.; Hou, X.; Quan, C.; Chen, M. Widespread Existence of Quorum Sensing Inhibitors in Marine Bacteria: Potential Drugs to Combat Pathogens with Novel Strategies. Mar. Drugs 2019, 17, 275. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Almada, K.; González-Acosta, B.; Borges-Souza, J.M.; Aguila-Ramírez, R.N. Marine Bacteria Associated with Shallow Hydrothermal Systems in the Gulf of California with the Capacity to Produce Biofilm Inhibiting Compounds. Arch. Microbiol. 2020, 202, 1477–1488. [Google Scholar] [CrossRef] [PubMed]
- Khalid, S.J.; Ain, Q.; Khan, S.J.; Jalil, A.; Siddiqui, M.F.; Ahmad, T.; Badshah, M.; Adnan, F. Targeting Acyl Homoserine Lactones (AHLs) by the Quorum Quenching Bacterial Strains to Control Biofilm Formation in Pseudomonas aeruginosa. Saudi J. Biol. Sci. 2022, 29, 1673–1682. [Google Scholar] [CrossRef]
- Kamarudheen, N.; Rao, K.V.B. Fatty Acyl Compounds from Marine Streptomyces griseoincarnatus Strain HK12 against Two Major Bio-Film Forming Nosocomial Pathogens; an In Vitro and In Silico Approach. Microb. Pathog. 2019, 127, 121–130. [Google Scholar] [CrossRef]
- Song, Y.; Zhou, Y.; Cong, M.; Deng, S.; Chen, Y.; Pang, X.; Liu, Y.; Liao, L.; Yang, L.; Wang, J. New 24-Membered Macrolactines from an Arctic Bacterium Bacillus amyloliquefaciens SCSIO 41392 and Their Anti-Pathogenicity Evaluation. Mar. Drugs 2024, 22, 484. [Google Scholar] [CrossRef]
- Li, D.-D.; Wang, Y.; Li, H.; Niu, W.-X.; Hong, J.; Jung, J.H.; Lee, J.-H. Multifaceted Antipathogenic Activity of Two Novel Natural Products, Chermesiterpenoid B and Chermesiterpenoid B Seco Acid Methyl Ester, Against Pseudomonas aeruginosa. Microb. Biotechnol. 2025, 18, e70101. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, L.; Wu, H.; Zhao, C.; Gong, Q.; Yu, W. Cladodionen Is a Potential Quorum Sensing Inhibitor Against Pseudomonas aeruginosa. Mar. Drugs 2020, 18, 205. [Google Scholar] [CrossRef]
- Choi, H.-Y.; Le, D.D.; Kim, W.-G. Curvularin Isolated From Phoma Macrostoma Is an Antagonist of RhlR Quorum Sensing in Pseudomonas aeruginosa. Front. Microbiol. 2022, 13, 913882. [Google Scholar] [CrossRef]
- Chang, A.; Sun, S.; Li, L.; Dai, X.; Li, H.; He, Q.; Zhu, H. Tyrosol from Marine Fungi, a Novel Quorum Sensing Inhibitor against Chromobacterium violaceum and Pseudomonas aeruginosa. Bioorg Chem. 2019, 91, 103140. [Google Scholar] [CrossRef]
- Riahi, A.; Mabudi, H.; Tajbakhsh, E.; Roomiani, L.; Momtaz, H. Optimizing Chitosan Derived from Metapenaeus Affinis: A Novel Anti-Biofilm Agent against Pseudomonas aeruginosa. AMB Express 2024, 14, 77. [Google Scholar] [CrossRef]
- Zeng, Y.-X.; Liu, J.-S.; Wang, Y.-J.; Tang, S.; Wang, D.-Y.; Deng, S.-M.; Jia, A.-Q. Actinomycin D: A Novel Pseudomonas aeruginosa Quorum Sensing Inhibitor from the Endophyte Streptomyces cyaneochromogenes RC1. World J. Microbiol. Biotechnol. 2022, 38, 170. [Google Scholar] [CrossRef] [PubMed]
- Alford, M.A.; Mann, S.; Akhoundsadegh, N.; Hancock, R.E.W. Competition between Pseudomonas aeruginosa and Staphylococcus aureus Is Dependent on Intercellular Signaling and Regulated by the NtrBC Two-Component System. Sci. Rep. 2022, 12, 9027. [Google Scholar] [CrossRef] [PubMed]
- Talaat, R.; Abu El-Naga, M.N.; El-Bialy, H.A.A.; El-Fouly, M.Z.; Abouzeid, M.A. Quenching of Quorum Sensing in Multi-Drug Resistant Pseudomonas aeruginosa: Insights on Halo-Bacterial Metabolites and Gamma Irradiation as Channels Inhibitors. Ann. Clin. Microbiol. Antimicrob. 2024, 23, 31. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Liu, G.; Sun, C.; Wu, S. Volatile Organic Compounds Produced by a Deep-Sea Bacterium Efficiently Inhibit the Growth of Pseudomonas aeruginosa PAO1. Mar. Drugs. 2024, 22, 233. [Google Scholar] [CrossRef]
- Wu, Z.; Zheng, R.; Zhang, J.; Wu, S. Transcriptional Profiling of Pseudomonas aeruginosa PAO1 in Response to Anti-Biofilm and Anti-Infection Agent Exopolysaccharide EPS273. J. Appl. Microbiol. 2021, 130, 265–277. [Google Scholar] [CrossRef]
- Patel, M.; Siddiqui, A.J.; Ashraf, S.A.; Surti, M.; Awadelkareem, A.M.; Snoussi, M.; Hamadou, W.S.; Bardakci, F.; Jamal, A.; Jahan, S.; et al. Lactiplantibacillus plantarum-Derived Biosurfactant Attenuates Quorum Sensing-Mediated Virulence and Biofilm Formation in Pseudomonas aeruginosa and Chromobacterium violaceum. Microorganisms 2022, 10, 1026. [Google Scholar] [CrossRef]
- Ji, H.; Zhao, L.; Lv, K.; Zhang, Y.; Gao, H.; Gong, Q.; Yu, W. Citrinin Is a Potential Quorum Sensing Inhibitor against Pseudomonas aeruginosa. Mar. Drugs 2023, 21, 296. [Google Scholar] [CrossRef]
- Chen, X.; Chen, J.; Yan, Y.; Chen, S.; Xu, X.; Zhang, H.; Wang, H. Quorum Sensing Inhibitors from Marine Bacteria Oceanobacillus sp. XC22919. Nat. Prod. Res. 2019, 33, 1819–1823. [Google Scholar] [CrossRef]
- Goel, N.; Ghosh, M.; Jain, D.; Sinha, R.; Khare, S.K. Inhibition and Eradication of Pseudomonas aeruginosa Biofilms by Secondary Metabolites of Nocardiopsis Lucentensis EMB25. RSC Med. Chem. 2023, 14, 745–756. [Google Scholar] [CrossRef]
- Han, L.; Ren, J.; Xue, Y.; Xie, G.; Gao, J.; Fu, Q.; Shao, P.; Zhu, H.; Zhang, M.; Ding, F. Palmitoleic Acid Inhibits Pseudomonas aeruginosa Quorum Sensing Activation and Protects Lungs from Infectious Injury. Respir. Res. 2024, 25, 423. [Google Scholar] [CrossRef]
- Malešević, M.; Di Lorenzo, F.; Filipić, B.; Stanisavljević, N.; Novović, K.; Senerovic, L.; Polović, N.; Molinaro, A.; Kojić, M.; Jovčić, B. Pseudomonas aeruginosa Quorum Sensing Inhibition by Clinical Isolate Delftia Tsuruhatensis 11304: Involvement of N-Octadecanoylhomoserine Lactones. Sci. Rep. 2019, 9, 16465. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.D.; Patel, H.; Saiyad, S.M.; Bajpai, B. Effect of a Phthalate Derivative Purified from Bacillus Zhangzhouensis SK4 on Quorum Sensing Regulated Virulence Factors of Pseudomonas aeruginosa. Microb. Pathog. 2024, 191, 106664. [Google Scholar] [CrossRef]
- Onbas, T.; Osmanagaoglu, O.; Kiran, F. Potential Properties of Lactobacillus Plantarum F-10 as a Bio-Control Strategy for Wound Infections. Probiotics Antimicrob. Proteins 2019, 11, 1110–1123. [Google Scholar] [CrossRef]
- Issa, R.; Chanishvili, N.; Caplin, J.; Kakabadze, E.; Bakuradze, N.; Makalatia, K.; Cooper, I. Antibiofilm Potential of Purified Environmental Bacteriophage Preparations against Early Stage Pseudomonas aeruginosa Biofilms. J. Appl. Microbiol. 2019, 126, 1657–1667. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, M.H.; Elgaml, A.; Abdel Bar, F.M.; Badria, F.A. Antimicrobial and Antiquorum-Sensing Activity of Ricinus Communis Extracts and Ricinine Derivatives. Nat. Prod. Res. 2019, 33, 1556–1562. [Google Scholar] [CrossRef]
- Salleh, N.F.; Wang, J.; Kundukad, B.; Oluwabusola, E.T.; Goh, D.X.Y.; Phyo, M.Y.; Tong, J.J.L.; Kjelleberg, S.; Tan, L.T. Cyclopropane-Containing Specialized Metabolites from the Marine Cyanobacterium cf. Lyngbya sp. Molecules 2023, 28, 3965. [Google Scholar] [CrossRef] [PubMed]
- Oluwabusola, E.T.; Katermeran, N.P.; Poh, W.H.; Goh, T.M.B.; Tan, L.T.; Diyaolu, O.; Tabudravu, J.; Ebel, R.; Rice, S.A.; Jaspars, M. Inhibition of the Quorum Sensing System, Elastase Production and Biofilm Formation in Pseudomonas aeruginosa by Psammaplin A and Bisaprasin. Molecules 2022, 27, 1721. [Google Scholar] [CrossRef]
- Baloyi, I.T.; Adeosun, I.J.; Yusuf, A.A.; Cosa, S. In Silico and In Vitro Screening of Antipathogenic Properties of Melianthus comosus (Vahl) against Pseudomonas aeruginosa. Antibiotics 2021, 10, 679. [Google Scholar] [CrossRef]
- Fekete-Kertész, I.; Berkl, Z.; Buda, K.; Fenyvesi, É.; Szente, L.; Molnár, M. Quorum Quenching Effect of Cyclodextrins on the Pyocyanin and Pyoverdine Production of Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2024, 108, 271. [Google Scholar] [CrossRef]
- Bano, S.; Hassan, N.; Rafiq, M.; Hassan, F.; Rehman, M.; Iqbal, N.; Ali, H.; Hasan, F.; Kang, Y.-Q. Biofilms as Battlefield Armor for Bacteria against Antibiotics: Challenges and Combating Strategies. Microorganisms 2023, 11, 2595. [Google Scholar] [CrossRef] [PubMed]
- Alshammari, M.; Ahmad, A.; AlKhulaifi, M.; Al Farraj, D.; Alsudir, S.; Alarawi, M.; Takashi, G.; Alyamani, E. Reduction of Biofilm Formation of Escherichia coli by Targeting Quorum Sensing and Adhesion Genes Using the CRISPR/Cas9-HDR Approach, and Its Clinical Application on Urinary Catheter. J. Infect. Public Health 2023, 16, 1174–1183. [Google Scholar] [CrossRef] [PubMed]
- Askoura, M.; Almalki, A.J.; Lila, A.S.A.; Almansour, K.; Alshammari, F.; Khafagy, E.-S.; Ibrahim, T.S.; Hegazy, W.A.H. Alteration of Salmonella enterica Virulence and Host Pathogenesis through Targeting sdiA by Using the CRISPR-Cas9 System. Microorganisms 2021, 9, 2564. [Google Scholar] [CrossRef]
- Geyman, L.J.; Tanner, M.P.; Rosario-Meléndez, N.; Peters, J.M.; Mandel, M.J.; van Kessel, J.C. Mobile-CRISPRi as a Powerful Tool for Modulating Vibrio Gene Expression. Appl. Environ. Microbiol. 2024, 90, e0006524. [Google Scholar] [CrossRef]
- Elshobary, M.E.; Badawy, N.K.; Ashraf, Y.; Zatioun, A.A.; Masriya, H.H.; Ammar, M.M.; Mohamed, N.A.; Mourad, S.; Assy, A.M. Combating Antibiotic Resistance: Mechanisms, Multidrug-Resistant Pathogens, and Novel Therapeutic Approaches: An Updated Review. Pharmaceuticals 2025, 18, 402. [Google Scholar] [CrossRef]
- Li, R.; Fang, L.; Tan, S.; Yu, M.; Li, X.; He, S.; Wei, Y.; Li, G.; Jiang, J.; Wu, M. Type I CRISPR-Cas Targets Endogenous Genes and Regulates Virulence to Evade Mammalian Host Immunity. Cell Res. 2016, 26, 1273–1287. [Google Scholar] [CrossRef]
- Radha KesavanNair, L. Computational Design of Guide RNAs and Vector to Knockout LasR Gene of Pseudomonas aeruginosa. Gene Genome Ed. 2023, 6, 100028. [Google Scholar] [CrossRef]
- Ghosh, S.; Lahiri, D.; Nag, M.; Sarkar, T.; Pati, S.; Edinur, H.A.; Kumar, M.; Mohd Zain, M.R.A.; Ray, R.R. Precision Targeting of Food Biofilm-Forming Genes by Microbial Scissors: CRISPR-Cas as an Effective Modulator. Front. Microbiol. 2022, 13, 964848. [Google Scholar] [CrossRef] [PubMed]
- Ghavami, S.; Pandi, A. Chapter Five—CRISPR Interference and Its Applications. In Progress in Molecular Biology and Translational Science; Singh, V., Ed.; Reprogramming the Genome: Applications of CRISPR-Cas in Non-mammalian Systems Part B; Academic Press: Cambridge, MA, USA, 2021; pp. 123–140. [Google Scholar]
- Kachhadia, R.; Kapadia, C.; Singh, S.; Gandhi, K.; Jajda, H.; Alfarraj, S.; Ansari, M.J.; Danish, S.; Datta, R. Quorum Sensing Inhibitory and Quenching Activity of Bacillus cereus RC1 Extracts on Soft Rot-Causing Bacteria Lelliottia Amnigena. ACS Omega 2022, 7, 25291–25308. [Google Scholar] [CrossRef]
- Ghosh, S.; Nandi, S.; Basu, T. Nano-Antibacterials Using Medicinal Plant Components: An Overview. Front. Microbiol. 2021, 12, 768739. [Google Scholar] [CrossRef]
- Antunes, J.C.; Domingues, J.M.; Miranda, C.S.; Silva, A.F.G.; Homem, N.C.; Amorim, M.T.P.; Felgueiras, H.P. Bioactivity of Chitosan-Based Particles Loaded with Plant-Derived Extracts for Biomedical Applications: Emphasis on Antimicrobial Fiber-Based Systems. Mar. Drugs 2021, 19, 359. [Google Scholar] [CrossRef] [PubMed]
- Fattah, R.A.F.A.; Fathy, F.E.Z.Y.; Mohamed, T.A.H.; Elsayed, M.S. Effect of Chitosan Nanoparticles on Quorum Sensing-Controlled Virulence Factors and Expression of LasI and RhlI Genes among Pseudomonas aeruginosa Clinical Isolates. AIMS Microbiol. 2021, 7, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Micale, N.; Citarella, A.; Molonia, M.S.; Speciale, A.; Cimino, F.; Saija, A.; Cristani, M. Hydrogels for the Delivery of Plant-Derived (Poly)Phenols. Molecules 2020, 25, 3254. [Google Scholar] [CrossRef]
- Wong, M.Y.; Lin, B.-S.; Hu, P.-S.; Huang, T.-Y.; Huang, Y.-K. Nanoparticles of Cs0.33WO3 as Antibiofilm Agents and Photothermal Treatment to Inhibit Biofilm Formation. ACS Omega 2024, 9, 28144–28154. [Google Scholar] [CrossRef]
- Kim, H.-S.; Lee, S.-H.; Byun, Y.; Park, H.-D. 6-Gingerol Reduces Pseudomonas aeruginosa Biofilm Formation and Virulence via Quorum Sensing Inhibition. Sci. Rep. 2015, 5, 8656. [Google Scholar] [CrossRef]
- Rathinam, P.; Vijay Kumar, H.S.; Viswanathan, P. Eugenol Exhibits Anti-Virulence Properties by Competitively Binding to Quorum Sensing Receptors. Biofouling 2017, 33, 624–639. [Google Scholar] [CrossRef]
- Zhong, L.; Ravichandran, V.; Zhang, N.; Wang, H.; Bian, X.; Zhang, Y.; Li, A. Attenuation of Pseudomonas aeruginosa Quorum Sensing by Natural Products: Virtual Screening, Evaluation and Biomolecular Interactions. Int. J. Mol. Sci. 2020, 21, 2190. [Google Scholar] [CrossRef]
- Kalia, M.; Yadav, V.K.; Singh, P.K.; Sharma, D.; Narvi, S.S.; Agarwal, V. Exploring the Impact of Parthenolide as Anti-Quorum Sensing and Anti-Biofilm Agent against Pseudomonas aeruginosa. Life Sci. 2018, 199, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Hernando-Amado, S.; Alcalde-Rico, M.; Gil-Gil, T.; Valverde, J.R.; Martínez, J.L. Naringenin Inhibition of the Pseudomonas aeruginosa Quorum Sensing Response Is Based on Its Time-Dependent Competition With N-(3-Oxo-Dodecanoyl)-L-Homoserine Lactone for LasR Binding. Front. Mol. Biosci. 2020, 7, 25. [Google Scholar] [CrossRef]
- Shah, S.; Gaikwad, S.; Nagar, S.; Kulshrestha, S.; Vaidya, V.; Nawani, N.; Pawar, S. Biofilm Inhibition and Anti-Quorum Sensing Activity of Phytosynthesized Silver Nanoparticles against the Nosocomial Pathogen Pseudomonas aeruginosa. Biofouling 2019, 35, 34–49. [Google Scholar] [CrossRef]
- Kumar, L.; Chhibber, S.; Kumar, R.; Kumar, M.; Harjai, K. Zingerone Silences Quorum Sensing and Attenuates Virulence of Pseudomonas aeruginosa. Fitoterapia 2015, 102, 84–95. [Google Scholar] [CrossRef]
- Bose, S.K.; Chauhan, M.; Dhingra, N.; Chhibber, S.; Harjai, K. Terpinen-4-Ol Attenuates Quorum Sensing Regulated Virulence Factors and Biofilm Formation in Pseudomonas aeruginosa. Future Microbiol. 2020, 15, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Parai, D.; Banerjee, M.; Dey, P.; Chakraborty, A.; Islam, E.; Mukherjee, S.K. Effect of Reserpine on Pseudomonas aeruginosa Quorum Sensing Mediated Virulence Factors and Biofilm Formation. Biofouling 2018, 34, 320–334. [Google Scholar] [CrossRef] [PubMed]
- Samreen; Qais, F.A.; Ahmad, I. Anti-Quorum Sensing and Biofilm Inhibitory Effect of Some Medicinal Plants against Gram-Negative Bacterial Pathogens: In Vitro and In Silico Investigations. Heliyon 2022, 8, e11113. [Google Scholar] [CrossRef] [PubMed]
- Maiga, A.; Ampomah-Wireko, M.; Li, H.; Fan, Z.; Lin, Z.; Zhen, H.; Kpekura, S.; Wu, C. Multidrug-Resistant Bacteria Quorum-Sensing Inhibitors: A Particular Focus on Pseudomonas aeruginosa. Eur. J. Med. Chem. 2025, 281, 117008. [Google Scholar] [CrossRef]
- Azimzadeh, M.; Greco, G.; Farmani, A.; Nourian, A.; Pourhajibagher, M.; Taherkhani, A.; Alikhani, M.Y.; Bahador, A. Biofilm Inhibition of Multidrug-Resistant Pseudomonas aeruginosa Using Green-Synthesized Silver Nanoparticles and Colistin. Sci. Rep. 2025, 15, 14993. [Google Scholar] [CrossRef]
- Soukarieh, F.; Williams, P.; Stocks, M.J.; Cámara, M. Pseudomonas aeruginosa Quorum Sensing Systems as Drug Discovery Targets: Current Position and Future Perspectives. J. Med. Chem. 2018, 61, 10385–10402. [Google Scholar] [CrossRef]
- Chuang, S.K.; Vrla, G.D.; Fröhlich, K.S.; Gitai, Z. Surface Association Sensitizes Pseudomonas aeruginosa to Quorum Sensing. Nat. Commun. 2019, 10, 4118. [Google Scholar] [CrossRef]
- Simanek, K.A.; Schumacher, M.L.; Mallery, C.P.; Shen, S.; Li, L.; Paczkowski, J.E. Quorum-Sensing Synthase Mutations Re-Calibrate Autoinducer Concentrations in Clinical Isolates of Pseudomonas aeruginosa to Enhance Pathogenesis. Nat. Commun. 2023, 14, 7986. [Google Scholar] [CrossRef]
- O’Connor, K.; Zhao, C.Y.; Mei, M.; Diggle, S.P. Frequency of Quorum-Sensing Mutations in Pseudomonas aeruginosa Strains Isolated from Different Environments. Microbiology 2022, 168, 001265. [Google Scholar] [CrossRef]
- Schlichter Kadosh, Y.; Muthuraman, S.; Nisaa, K.; Ben-Zvi, A.; Karsagi Byron, D.L.; Shagan, M.; Brandis, A.; Mehlman, T.; Gopas, J.; Saravana Kumar, R.; et al. Pseudomonas aeruginosa Quorum Sensing and Biofilm Attenuation by a Di-Hydroxy Derivative of Piperlongumine (PL-18). Biofilm 2024, 8, 100215. [Google Scholar] [CrossRef] [PubMed]
- Simanek, K.A.; Paczkowski, J.E. Resistance Is Not Futile: The Role of Quorum Sensing Plasticity in Pseudomonas aeruginosa Infections and Its Link to Intrinsic Mechanisms of Antibiotic Resistance. Microorganisms 2022, 10, 1247. [Google Scholar] [CrossRef] [PubMed]
- Brindhadevi, K.; LewisOscar, F.; Mylonakis, E.; Shanmugam, S.; Verma, T.N.; Pugazhendhi, A. Biofilm and Quorum Sensing Mediated Pathogenicity in Pseudomonas aeruginosa. Process Biochem. 2020, 96, 49–57. [Google Scholar] [CrossRef]
- Yang, Y.-X.; Xu, Z.-H.; Zhang, Y.-Q.; Tian, J.; Weng, L.-X.; Wang, L.-H. A New Quorum-Sensing Inhibitor Attenuates Virulence and Decreases Antibiotic Resistance in Pseudomonas aeruginosa. J. Microbiol. 2012, 50, 987–993. [Google Scholar] [CrossRef]
- Kostylev, M.; Kim, D.Y.; Smalley, N.E.; Salukhe, I.; Greenberg, E.P.; Dandekar, A.A. Evolution of the Pseudomonas aeruginosa Quorum-Sensing Hierarchy. Proc. Natl. Acad. Sci. USA 2019, 116, 7027–7032. [Google Scholar] [CrossRef] [PubMed]
- Koul, S.; Prakash, J.; Mishra, A.; Kalia, V.C. Potential Emergence of Multi-Quorum Sensing Inhibitor Resistant (MQSIR) Bacteria. Indian J. Microbiol. 2016, 56, 1–18. [Google Scholar] [CrossRef]
- Jeske, A.; Arce-Rodriguez, A.; Thöming, J.G.; Tomasch, J.; Häussler, S. Evolution of Biofilm-Adapted Gene Expression Profiles in lasR-Deficient Clinical Pseudomonas aeruginosa Isolates. npj Biofilms Microbiomes 2022, 8, 6. [Google Scholar] [CrossRef]
- Pan, Y.-P.; Xu, Y.-H.; Wang, Z.-X.; Fang, Y.-P.; Shen, J.-L. Overexpression of MexAB-OprM Efflux Pump in Carbapenem-Resistant Pseudomonas aeruginosa. Arch. Microbiol. 2016, 198, 565–571. [Google Scholar] [CrossRef]
- Abdelraheem, W.M.; Yassin, H.I.; Zaki, S.; Esmail, M.A.M. Zinc Sulfate Acts as an Efflux Pump Inhibitor on Pseudomonas aeruginosa Clinical Isolates. World J. Microbiol. Biotechnol. 2025, 41, 139. [Google Scholar] [CrossRef]
- Akinduti, P.A.; George, O.W.; Ohore, H.U.; Ariyo, O.E.; Popoola, S.T.; Adeleye, A.I.; Akinwande, K.S.; Popoola, J.O.; Rotimi, S.O.; Olufemi, F.O.; et al. Evaluation of Efflux-Mediated Resistance and Biofilm Formation in Virulent Pseudomonas aeruginosa Associated with Healthcare Infections. Antibiotics 2023, 12, 626. [Google Scholar] [CrossRef]
- Bové, M.; Bao, X.; Sass, A.; Crabbé, A.; Coenye, T. The Quorum-Sensing Inhibitor Furanone C-30 Rapidly Loses Its Tobramycin-Potentiating Activity against Pseudomonas aeruginosa Biofilms during Experimental Evolution. Antimicrob. Agents Chemother. 2021, 65, e0041321. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, M.; Yu, C.; Li, J.; Zhou, X. Biofilm Formation: Mechanistic Insights and Therapeutic Targets. Mol. Biomed. 2023, 4, 49. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Prentice, E.L.; Webber, M.A. Mechanisms of Antimicrobial Resistance in Biofilms. npj Antimicrob. Resist. 2024, 2, 27. [Google Scholar] [CrossRef]
- Tchadi, B.V.; Derringer, J.J.; Detweiler, A.K.; Taylor, I.R. PqsE Adapts the Activity of the Pseudomonas aeruginosa Quorum-Sensing Transcription Factor RhlR to Both Autoinducer Concentration and Promoter Sequence Identity. J. Bacteriol. 2025, 7, e00516-24. [Google Scholar] [CrossRef]
- Ampomah-Wireko, M.; Luo, C.; Cao, Y.; Wang, H.; Nininahazwe, L.; Wu, C. Chemical Probe of AHL Modulators on Quorum Sensing in Gram-Negative Bacteria and as Antiproliferative Agents: A Review. Eur. J. Med. Chem. 2021, 226, 113864. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Ahator, S.D.; Zhang, L.-H. Molecular Mechanisms of Phosphate Stress Activation of Pseudomonas aeruginosa Quorum Sensing Systems. mSphere 2020, 5, e00119-20. [Google Scholar] [CrossRef]
- Krzyżek, P. Challenges and Limitations of Anti-Quorum Sensing Therapies. Front. Microbiol. 2019, 10, 2473. [Google Scholar] [CrossRef]
- Taha, M.N.; Saafan, A.E.; Ahmedy, A.; El Gebaly, E.; Khairalla, A.S. Two Novel Synthetic Peptides Inhibit Quorum Sensing-Dependent Biofilm Formation and Some Virulence Factors in Pseudomonas aeruginosa PAO1. J. Microbiol. 2019, 57, 618–625. [Google Scholar] [CrossRef]
- Gondil, V.S.; Kalaiyarasan, T.; Bharti, V.K.; Chhibber, S. Antibiofilm Potential of Seabuckthorn Silver Nanoparticles (SBT@AgNPs) against Pseudomonas aeruginosa. 3 Biotech 2019, 9, 402. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.; Feng, T.; Du, R.; Tian, X.; Wang, Y.; Zhang, X.-H. Heterologous Expression of the Marine-Derived Quorum Quenching Enzyme MomL Can Expand the Antibacterial Spectrum of Bacillus Brevis. Mar. Drugs 2019, 17, 128. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, Q.; Li, L.; Pan, L.; Zhu, H. Anti-Quorum-Sensing Activity of Tryptophan-Containing Cyclic Dipeptides. Mar. Drugs 2022, 20, 85. [Google Scholar] [CrossRef] [PubMed]
- Sikdar, R.; Elias, M. Quorum Quenching Enzymes and Their Effects on Virulence, Biofilm and Microbiomes: A Review of Recent Advances. Expert Rev. Anti Infect Ther. 2020, 18, 1221–1233. [Google Scholar] [CrossRef]
- Hetta, H.F.; Ramadan, Y.N.; Rashed, Z.I.; Alharbi, A.A.; Alsharef, S.; Alkindy, T.T.; Alkhamali, A.; Albalawi, A.S.; Battah, B.; Donadu, M.G. Quorum Sensing Inhibitors: An Alternative Strategy to Win the Battle against Multidrug-Resistant (MDR) Bacteria. Molecules 2024, 29, 3466. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-W.; Jia, A.-Q.; Jiang, H.; Li, P.-L.; Chen, H.; Tan, X.-J.; Liu, E.-Q. 1-(4-Amino-2-Hydroxyphenyl)Ethanone from Phomopsis Liquidambari Showed Quorum Sensing Inhibitory Activity against Pseudomonas aeruginosa. Appl. Microbiol Biotechnol. 2021, 105, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Lesic, B.; Starkey, M.; He, J.; Hazan, R.; Rahme, L.G. Quorum Sensing Differentially Regulates Pseudomonas aeruginosa Type VI Secretion Locus I and Homologous Loci II and III, Which Are Required for Pathogenesis. Microbiology 2009, 155, 2845–2855. [Google Scholar] [CrossRef]
- Chang, A.; He, Q.; Li, L.; Yu, X.; Sun, S.; Zhu, H. Exploring the Quorum Sensing Inhibition of Isolated Chrysin from Penicillium Chrysogenum DXY-1. Bioorg. Chem. 2021, 111, 104894. [Google Scholar] [CrossRef]
- Yaeger, L.N.; Ranieri, M.R.M.; Chee, J.; Karabelas-Pittman, S.; Rudolph, M.; Giovannoni, A.M.; Harvey, H.; Burrows, L.L. A Genetic Screen Identifies a Role for oprF in Pseudomonas aeruginosa Biofilm Stimulation by Subinhibitory Antibiotics. npj Biofilms Microbiomes 2024, 10, 30. [Google Scholar] [CrossRef]
- Ma, Y.; Tang, W.S.; Liu, S.Y.; Khoo, B.L.; Chua, S.L. Juglone as a Natural Quorum Sensing Inhibitor against Pseudomonas aeruginosa Pqs-Mediated Virulence and Biofilms. ACS Pharmacol. Transl. Sci. 2024, 7, 533–543. [Google Scholar] [CrossRef]
- Singh, V.K.; Almpani, M.; Maura, D.; Kitao, T.; Ferrari, L.; Fontana, S.; Bergamini, G.; Calcaterra, E.; Pignaffo, C.; Negri, M.; et al. Tackling Recalcitrant Pseudomonas aeruginosa Infections in Critical Illness via Anti-Virulence Monotherapy. Nat. Commun. 2022, 13, 5103. [Google Scholar] [CrossRef]
- Cao, Y.-H.; Cai, W.-J.; He, X.-W.; Song, H.-L.; Gao, J.; Yang, Y.-L.; Zhou, J. A Review of Advances & Potential of Applying Nanomaterials for Biofilm Inhibition. npj Clean Water 2024, 7, 131. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L.; Yuan, X.; Wang, D. Treatment with Paeoniflorin Increases Lifespan of Pseudomonas aeruginosa Infected Caenorhabditis elegans by Inhibiting Bacterial Accumulation in Intestinal Lumen and Biofilm Formation. Front. Pharmacol. 2023, 14, 1114219. [Google Scholar] [CrossRef]
- Vadakkan, K.; Sathishkumar, K.; Mapranathukaran, V.O.; Ngangbam, A.K.; Nongmaithem, B.D.; Hemapriya, J.; Nair, J.B. Critical Review on Plant-Derived Quorum Sensing Signaling Inhibitors in Pseudomonas aeruginosa. Bioorg. Chem. 2024, 151, 107649. [Google Scholar] [CrossRef]
- Rajakumar, G.; Mao, L.; Bao, T.; Wen, W.; Wang, S.; Gomathi, T.; Gnanasundaram, N.; Rebezov, M.; Shariati, M.A.; Chung, I.-M.; et al. Yttrium Oxide Nanoparticle Synthesis: An Overview of Methods of Preparation and Biomedical Applications. Appl. Sci. 2021, 11, 2172. [Google Scholar] [CrossRef]
- Theuretzbacher, U. The Global Resistance Problem and the Clinical Antibacterial Pipeline. Nat. Rev. Microbiol. 2025. [Google Scholar] [CrossRef]
- Szymczak, M.; Pankowski, J.A.; Kwiatek, A.; Grygorcewicz, B.; Karczewska-Golec, J.; Sadowska, K.; Golec, P. An Effective Antibiofilm Strategy Based on Bacteriophages Armed with Silver Nanoparticles. Sci. Rep. 2024, 14, 9088. [Google Scholar] [CrossRef]
- Gad, A.I.; El-Ganiny, A.M.; Eissa, A.G.; Noureldin, N.A.; Nazeih, S.I. Miconazole and Phenothiazine Hinder the Quorum Sensing Regulated Virulence in Pseudomonas aeruginosa. J. Antibiot. 2024, 77, 454–465. [Google Scholar] [CrossRef] [PubMed]
- Shaker, B.; Ahmad, S.; Thai, T.D.; Eyun, S.-I.; Na, D. Rational Drug Design for Pseudomonas aeruginosa PqsA Enzyme: An In Silico Guided Study to Block Biofilm Formation. Front. Mol. Biosci. 2020, 7, 577316. [Google Scholar] [CrossRef] [PubMed]
- Elshaer, S.L.; Shaaban, M.I. Inhibition of Quorum Sensing and Virulence Factors of Pseudomonas aeruginosa by Biologically Synthesized Gold and Selenium Nanoparticles. Antibiotics 2021, 10, 1461. [Google Scholar] [CrossRef]
- Maddeboina, K.; Yada, B.; Kumari, S.; McHale, C.; Pal, D.; Durden, D.L. Recent Advances in Multitarget-Directed Ligands via In Silico Drug Discovery. Drug Discov. Today 2024, 29, 103904. [Google Scholar] [CrossRef]
- Carneiro, J.; Magalhães, R.P.; de la Oliva Roque, V.M.; Simões, M.; Pratas, D.; Sousa, S.F. TargIDe: A Machine-Learning Workflow for Target Identification of Molecules with Antibiofilm Activity against Pseudomonas aeruginosa. J. Comput. Aided Mol. Des. 2023, 37, 265–278. [Google Scholar] [CrossRef]
- Mishra, R.; Kushveer, J.S.; Khan, M.I.K.; Pagal, S.; Meena, C.K.; Murali, A.; Dhayalan, A.; Venkateswara Sarma, V. 2,4-Di-Tert-Butylphenol Isolated From an Endophytic Fungus, Daldinia Eschscholtzii, Reduces Virulence and Quorum Sensing in Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 1668. [Google Scholar] [CrossRef]
- Suzuki, S.; Morita, Y.; Ishige, S.; Kai, K.; Kawasaki, K.; Matsushita, K.; Ogura, K.; Miyoshi-Akiyama†, T.; Shimizu, T. Effects of Quorum Sensing–Interfering Agents, Including Macrolides and Furanone C-30, and an Efflux Pump Inhibitor on Nitrosative Stress Sensitivity in Pseudomonas aeruginosa. Microbiology 2024, 170, 001464. [Google Scholar] [CrossRef]
- Wang, J.; Liu, F.; Long, J.; Jin, Y.; Chen, S.; Duan, G.; Yang, H. The Application of CRISPR-Cas System in Staphylococcus aureus Infection. Heliyon 2024, 10, e34383. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wu, Y. A Review on Colloidal Delivery Vehicles Using Carvacrol as a Model Bioactive Compound. Food Hydrocoll. 2021, 120, 106922. [Google Scholar] [CrossRef]
- Rano, S.; Bhaduri, A.; Singh, M. Nanoparticle-Based Platforms for Targeted Drug Delivery to the Pulmonary System as Therapeutics to Curb Cystic Fibrosis: A Review. J. Microbiol. Methods 2024, 217, 106876. [Google Scholar] [CrossRef] [PubMed]
- Parihar, A.; Prajapati, B.G.; Paliwal, H.; Shukla, M.; Khunt, D.; Devrao Bahadure, S.; Dyawanapelly, S.; Junnuthula, V. Advanced Pulmonary Drug Delivery Formulations for the Treatment of Cystic Fibrosis. Drug Discov. Today 2023, 28, 103729. [Google Scholar] [CrossRef]
- Min, J.G.; Sanchez Rangel, U.J.; Franklin, A.; Oda, H.; Wang, Z.; Chang, J.; Fox, P.M. Topical Antibiotic Elution in a Collagen-Rich Hydrogel Successfully Inhibits Bacterial Growth and Biofilm Formation In Vitro. Antimicrob. Agents Chemother. 2020, 64, e00136-20. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.D.; Jarman, E.H.; Kuppalli, K.; Murphy, M.J.; Longaker, M.T.; Gurtner, G.; Fox, P.M. Successful Topical Treatment of Human Biofilms Using Multiple Antibiotic Elution from a Collagen-Rich Hydrogel. Sci. Rep. 2024, 14, 5621. [Google Scholar] [CrossRef]
- Brackman, G.; Garcia-Fernandez, M.J.; Lenoir, J.; De Meyer, L.; Remon, J.-P.; De Beer, T.; Concheiro, A.; Alvarez-Lorenzo, C.; Coenye, T. Dressings Loaded with Cyclodextrin-Hamamelitannin Complexes Increase Staphylococcus aureus Susceptibility Toward Antibiotics Both in Single as Well as in Mixed Biofilm Communities. Macromol. Biosci. 2016, 16, 859–869. [Google Scholar] [CrossRef]
- Sørensen, S.; Kvich, L.; Xu, Y.; Thomsen, T.R.; Bjarnsholt, T.; Thaarup, I. Development of a Tri-Species Wound Model for Studying Fungal-Bacterial Interactions and Antimicrobial Therapies. Biofilm 2025, 9, 100256. [Google Scholar] [CrossRef]
- Jing, Q.; Liu, R.; Jiang, Q.; Liu, Y.; He, J.; Zhou, X.; Yu, O.Y.; Chu, C.-H.; Cheng, L.; Ren, B.; et al. Staphylococcus aureus Wraps around Candida albicans and Synergistically Escapes from Neutrophil Extracellular Traps. Front. Immunol. 2024, 15, 1422440. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, N.; Jeong, J.; Yoon, D.; Kim, S.; Choi, J. Global Metabolomics Approach in In Vitro and In Vivo Models Reveals Hepatic Glutathione Depletion Induced by Amorphous Silica Nanoparticles. Chem. Biol. Interact. 2018, 293, 100–106. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, Y.; Xie, X.; Li, R.; You, J.; Zhao, X.; Wang, Y.; Guo, J. Development and Characterization of LasR Immobilized Monolithic Column for Screening Active Ingredients as Quorum Sensing Inhibitors Against P. Aeruginosa in Natural Products. Drug Des. Dev. Ther. 2025, 19, 2051–2064. [Google Scholar] [CrossRef] [PubMed]
- Maneira, C.; Chamas, A.; Lackner, G. Engineering Saccharomyces Cerevisiae for Medical Applications. Microb. Cell Fact. 2025, 24, 12. [Google Scholar] [CrossRef] [PubMed]
- León-Félix, J.; Villicaña, C. The Impact of Quorum Sensing on the Modulation of Phage-Host Interactions. J. Bacteriol. 2021, 203, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Subhaswaraj, P.; Barik, S.; Macha, C.; Chiranjeevi, P.V.; Siddhardha, B. Anti Quorum Sensing and Anti Biofilm Efficacy of Cinnamaldehyde Encapsulated Chitosan Nanoparticles against Pseudomonas aeruginosa PAO1. LWT 2018, 97, 752–759. [Google Scholar] [CrossRef]
- Mir, M.; Ahmed, N.; Rehman, A. ur Recent Applications of PLGA Based Nanostructures in Drug Delivery. Colloids Surf. B Biointerfaces 2017, 159, 217–231. [Google Scholar] [CrossRef]
- Lim, T.; Ham, S.-Y.; Nam, S.; Kim, M.; Lee, K.Y.; Park, H.-D.; Byun, Y. Recent Advance in Small Molecules Targeting RhlR of Pseudomonas aeruginosa. Antibiotics 2022, 11, 274. [Google Scholar] [CrossRef]
- Ali Agha, A.S.A.; Al-Samydai, A.; Aburjai, T. New Frontiers in CRISPR: Addressing Antimicrobial Resistance with Cas9, Cas12, Cas13, and Cas14. Heliyon 2025, 11, e42013. [Google Scholar] [CrossRef]
- Banala, V.T.; Mukherjee, D.; Mahajan, S.; Singh, P.K. Chapter 20—Current Status of FDA-Approved Marketed Nano Drug Products: Regulatory Considerations. In Multifunctional Nanocarriers; Mehra, N.K., Srivastava, S., Madan, J., Singh, P.K., Eds.; Micro and Nano Technologies; Elsevier: Amsterdam, The Netherlands, 2022; pp. 501–521. ISBN 978-0-323-85041-4. [Google Scholar]
- Milewska, S.; Niemirowicz-Laskowska, K.; Siemiaszko, G.; Nowicki, P.; Wilczewska, A.Z.; Car, H. Current Trends and Challenges in Pharmacoeconomic Aspects of Nanocarriers as Drug Delivery Systems for Cancer Treatment. Int. J. Nanomed. 2021, 16, 6593–6644. [Google Scholar] [CrossRef]
- CARB-X Is Funding a German Team of Scientists to Develop a New Treatment for Difficult-to-Treat Pseudomonas aeruginosa Infections in Cystic Fibrosis Patients. Carb-X. Available online: https://carb-x.org/carb-x-news/carb-x-is-funding-a-german-team-of-scientists-to-develop-a-new-treatment-for-difficult-to-treat-pseudomonas-aeruginosa-infections-in-cystic-fibrosis-patients/ (accessed on 10 January 2019).
- Zhao, K.; Yang, X.; Zeng, Q.; Zhang, Y.; Li, H.; Yan, C.; Li, J.S.; Liu, H.; Du, L.; Wu, Y.; et al. Evolution of lasR Mutants in Polymorphic Pseudomonas aeruginosa Populations Facilitates Chronic Infection of the Lung. Nat. Commun. 2023, 14, 5976. [Google Scholar] [CrossRef] [PubMed]
- Behera, M.; Behera, P.R.; Bhuyan, P.P.; Singh, L.; Pradhan, B. Algal Nanoparticles and Their Antibacterial Activity: Current Research Status and Future Prospectives. Drugs Drug Candidates 2023, 2, 554–570. [Google Scholar] [CrossRef]
- Shariati, A.; Noei, M.; Askarinia, M.; Khoshbayan, A.; Farahani, A.; Chegini, Z. Inhibitory Effect of Natural Compounds on Quorum Sensing System in Pseudomonas aeruginosa: A Helpful Promise for Managing Biofilm Community. Front. Pharmacol. 2024, 15, 1350391. [Google Scholar] [CrossRef] [PubMed]
- Nath, A.; Bhattacharjee, R.; Nandi, A.; Sinha, A.; Kar, S.; Manoharan, N.; Mitra, S.; Mojumdar, A.; Panda, P.K.; Patro, S.; et al. Phage Delivered CRISPR-Cas System to Combat Multidrug-Resistant Pathogens in Gut Microbiome. Biomed. Pharmacother. 2022, 151, 113122. [Google Scholar] [CrossRef]
- Schütz, C.; Ho, D.; Hamed, M.M.; Abdelsamie, A.S.; Röhrig, T.; Herr, C.; Kany, A.M.; Rox, K.; Schmelz, S.; Siebenbürger, L.; et al. A New PqsR Inverse Agonist Potentiates Tobramycin Efficacy to Eradicate Pseudomonas aeruginosa Biofilms. Adv. Sci. 2021, 8, 2004369. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).