
Exploring the Use of Viral Vectors Pseudotyped with Viral Glycoproteins as Tools to Study Antibody-Mediated Neutralizing Activity
 ,
,
Abstract
1. Introduction
2. Overview and Definition of Pseudotyped Viruses
3. Vectors Used for Pseudotyping
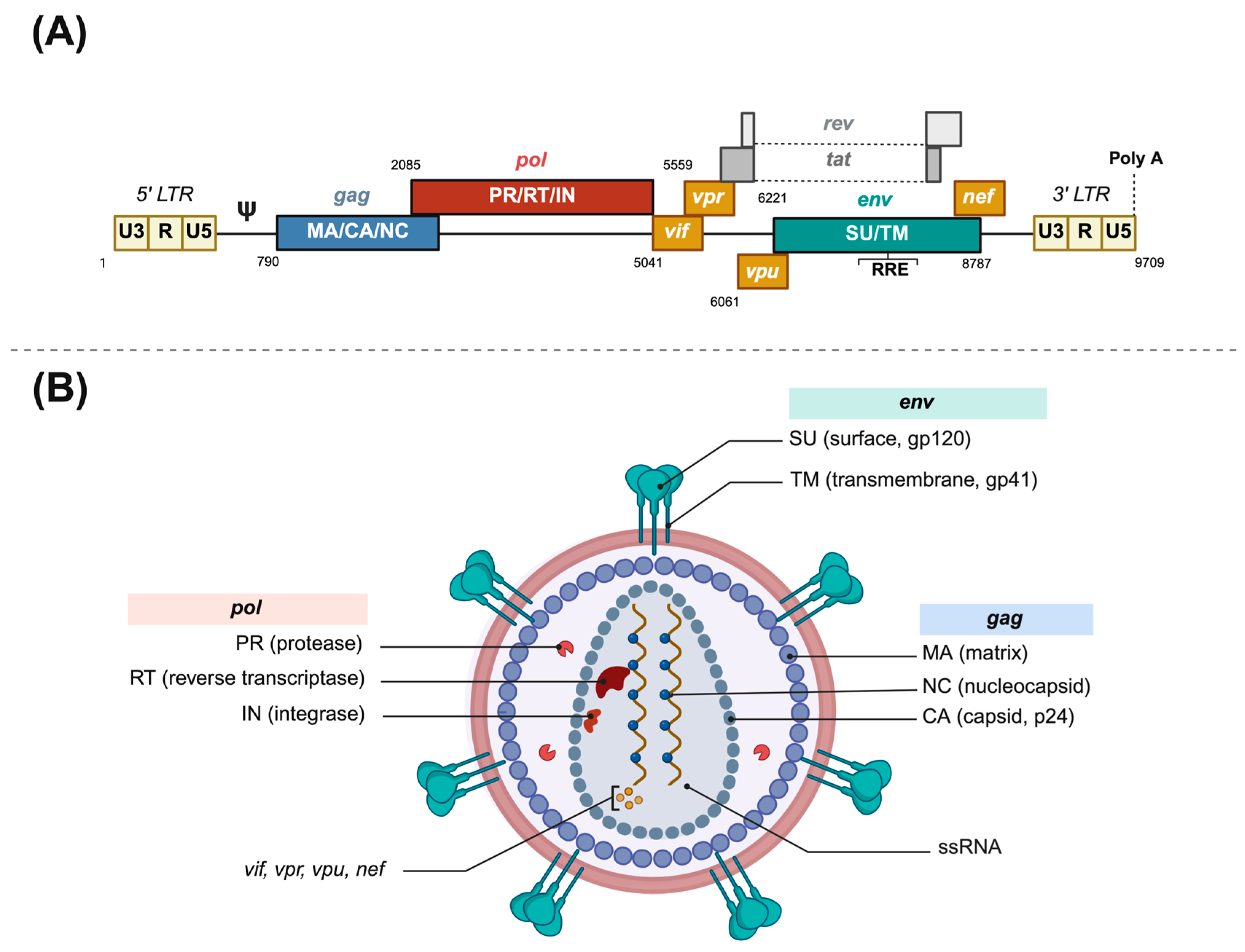
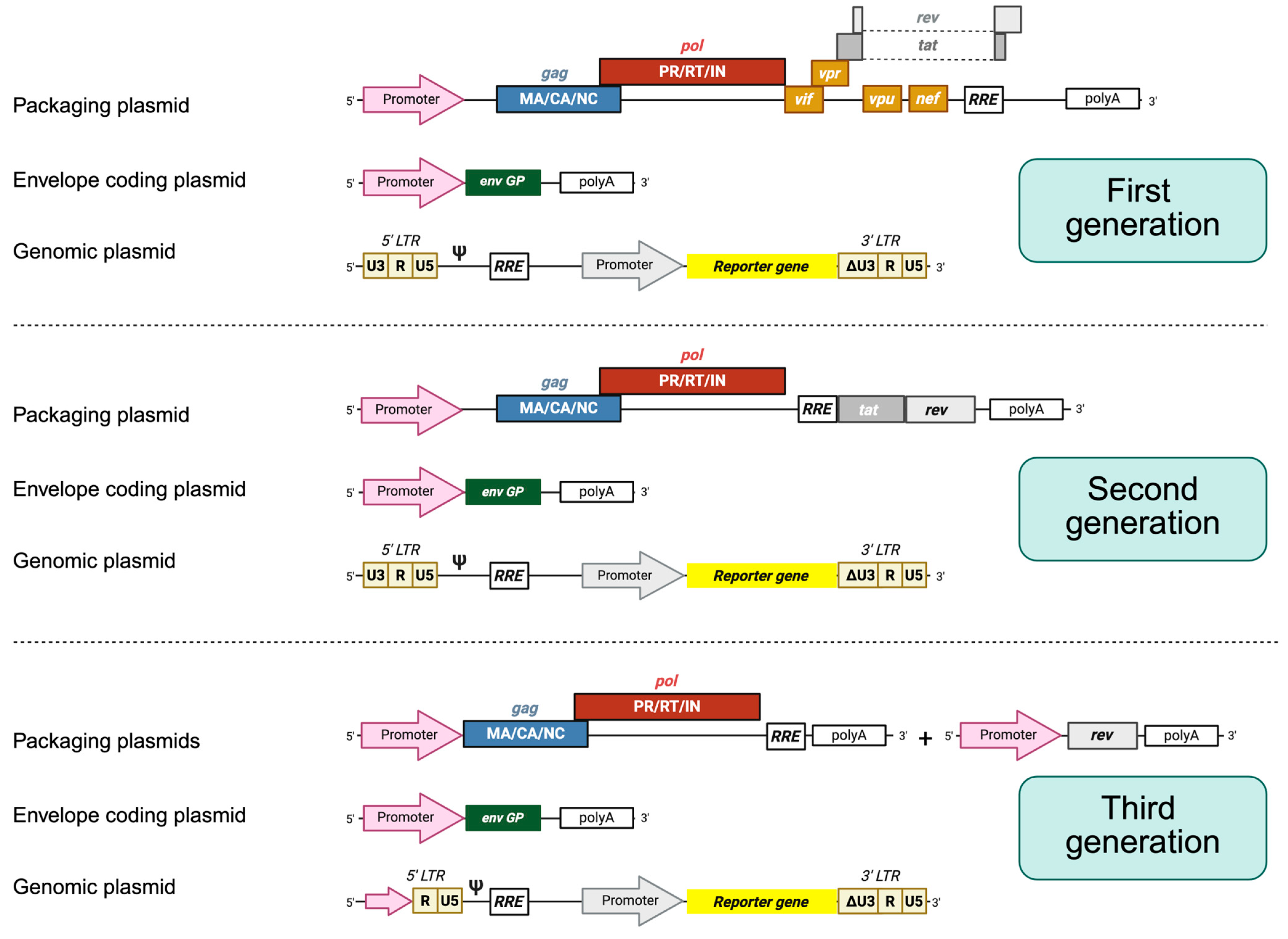
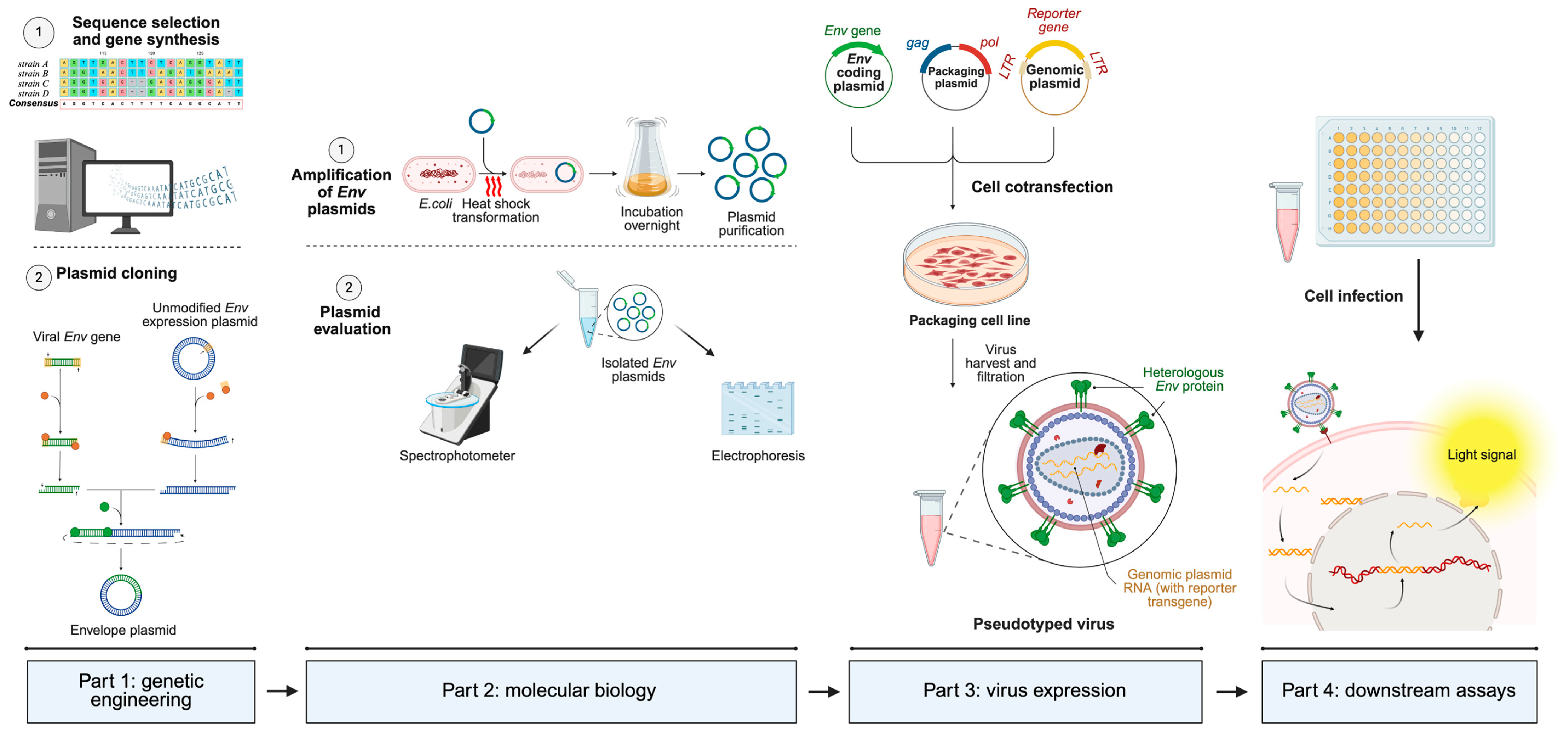
3.1. Lentiviral Vectors
3.2. Gammaretrovirus Vectors
3.3. The VSV Vector
3.4. Other Packaging Systems
3.5. Non-Enveloped Pseudotyped Viruses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Packaging System | Viral Protein | Research Line | Study |
|---|---|---|---|
| HIV-1 | EBOV GP | Gene therapy, cellular tropism, antiviral screening, cross-neutralization activity | [38,39,40] |
| HIV Env | Cellular tropism, neutralization antibody assay, Env mutation screening, antiretroviral therapy resistance, receptor recognition | [41,42,43,44] | |
| CHIKV E3 E2 E1 | Neutralization antibody assay, cellular tropism, in vivo imaging model | [45,46] | |
| RSV SH/G/F | Neutralization antibody assay, antiviral screening, identification of neutralization-escape genotypes. | [47] | |
| SARS-CoV-2 Spike | Receptor recognition and virus tropism, neutralization antibody epitopes, immune evasion, viral evolution | [48,49,50,51,52,53,54] | |
| MERS-CoV Spike | Receptor recognition, neutralization antibody assay, screening inhibitor of viral entry, viral evolution | [55,56] | |
| Influenza virus HA, NA, M2 | Mechanism of virus entry, antiviral screening, neutralization antibody assay | [17] | |
| Lassa virus | Neutralization antibody assay | [57] | |
| DENV PrM/E | Mechanism of virus entry, antiviral screening | [58] | |
| Zika virus PrM/E | Antiviral screening, oncologic virotherapy | [59,60] | |
| RVFV GP | Neutralization antibody assay | [61] | |
| Rabies virus G | Neutralization antibody assay, serosurveillance, cross-neutralizing activity, drug screening, gene delivery | [62,63,64,65,66,67] | |
| HBV GP | Entry inhibitor development | [68] | |
| MLV | Influenza HA | Neutralization antibody assay | [17,69] |
| HIV Env | Antiviral and antibody screening | [70] | |
| EBOV GP | Mutation study, infection mechanism, human spillover | [71,72,73,74] | |
| MERS-CoV Spike | Virus entry | [22,75] | |
| SIV Env | In vivo gene therapy for AIDS | [76] | |
| HCV GP | Infection mechanism, neutralization antibody assay, epitope mapping | [77,78,79] | |
| VSV | EBOV GP | Vaccine development, viral tropism, neutralization antibody assay, GP function | [80,81,82,83] |
| NiV Env | Antiviral screening, mutation study, neutralization antibody assay | [84,85,86] | |
| CCHFV GP | Neutralization antibody assay | [87] | |
| CHIKV E1 E2 | Neutralization antibody assay | [88] | |
| Measles virus H, F | Vaccine development | [89] | |
| Avian Influenza HA, NA | Antiviral screening, neutralization antibody assay | [90] | |
| HIV | HIV vaccine development | [91] | |
| SFTS virus GP | Neutralization antibody assay, vaccine development, infection mechanism | [92,93,94] | |
| HBV GP | Entry inhibitor development | [68] | |
| PFV | HIV specific epitopes | HIV vaccine development, AIDS gene therapy | [29] |
| HPV | HPV L1/L2 | Neutralization antibody assay, standardized vaccine efficacy | [95,96] |
| Self-assembly/ reverse genetics | PV capsid | Neutralization antibody assay, antigenic analysis | [97] |
| NiV M/Env | Structural analysis | [30] | |
| DENV C, prM, E | Drug discovery | [32] | |
| SARS-CoV-2 S, M, N | Viral evolution, infection mechanism | [31] | |
| WNV | Neutralization antibody assay | [33] | |
| Coxsackievirus | Neutralization antibody assay | [34,35,36] |
4. Applications of Pseudotyped Viruses
4.1. Study of Viral Tropism and Cell Infection
4.2. Assessment of Antibody-Mediated Neutralizing Activity
4.3. Pseudotypes as Immunogens/Vaccine Vectors
4.4. Antiviral Drug Screening
4.5. Analysis of Phenotypic Properties from Virus Mutants
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Judson, S.D.; Rabinowitz, P.M. Zoonoses and global epidemics. Curr. Opin. Infect. Dis. 2021, 34, 385–392. [Google Scholar] [CrossRef]
- Martínez, M.J.; Cotten, M.; Phan, M.V.T.; Becker, K.; Espasa, M.; Leegaard, T.M.; Lisby, G.; Schneider, U.V.; Casals-Pascual, C. Viral epidemic preparedness: A perspective from five clinical microbiology laboratories in Europe. Clin. Microbiol. Infect. 2023, 30, 582–585. [Google Scholar] [CrossRef]
- Mather, S.; Scott, S.; Temperton, N.; Wright, E.; King, B.; Daly, J. Current progress with serological assays for exotic emerging/re-emerging viruses. Future Virol. 2013, 8, 745–755. [Google Scholar] [CrossRef]
- Plotkin, S.A. Vaccines: Correlates of vaccine-induced immunity. Clin. Infect. Dis. 2008, 47, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Tao, L.; Zhang, X. Comprehensive and systemic optimization for improving the yield of SARS-CoV-2 spike pseudotyped virus. Mol. Ther. Methods Clin. Dev. 2021, 20, 350–356. [Google Scholar] [CrossRef]
- Sedgwick, R.L.; ElBohy, O.; Daly, J.M. Role of pseudotyped viruses in understanding epidemiology, pathogenesis and immunity of viral diseases affecting both horses and humans. Virology 2024, 597, 110164. [Google Scholar] [CrossRef]
- Thimmiraju, S.R.; Kimata, J.T.; Pollet, J. Pseudoviruses, a safer toolbox for vaccine development against enveloped viruses. Expert Rev. Vaccines 2024, 23, 174–185. [Google Scholar] [CrossRef]
- Joglekar, A.V.; Sandoval, S. Pseudotyped Lentiviral Vectors: One Vector, Many Guises. Hum. Gene Ther. Methods 2017, 28, 291–301. [Google Scholar] [CrossRef]
- Shchaslyvyi, A.Y.; Antonenko, S.V.; Tesliuk, M.G.; Telegeev, G.D. Current State of Human Gene Therapy: Approved Products and Vectors. Pharmaceuticals 2023, 16, 1416. [Google Scholar] [CrossRef]
- Whitt, M.A. Generation of VSV pseudotypes using recombinant ΔG-VSV for studies on virus entry, identification of entry inhibitors, and immune responses to vaccines. J. Virol. Methods 2010, 169, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liu, Q.; Huang, W.; Li, X.; Wang, Y. Current status on the development of pseudoviruses for enveloped viruses. Rev. Med. Virol. 2018, 28, e1963. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, Z.; Wu, X.; Li, T.; Wu, J.; Cai, M.; Nie, J.; Wang, W.; Cui, Z. Pseudotyped Viruses; Springer: Singapore, 2023; pp. 1–27. [Google Scholar]
- Mátrai, J.; Chuah, M.K.L.; VandenDriessche, T. Recent advances in lentiviral vector development and applications. Mol. Ther. 2010, 18, 477–490. [Google Scholar] [CrossRef]
- Johnson, N.M.; Alvarado, A.F.; Moffatt, T.N.; Edavettal, J.M.; Swaminathan, T.A.; Braun, S.E. HIV-based lentiviral vectors: Origin and sequence differences. Mol. Ther. Methods Clin. Dev. 2021, 21, 451–465. [Google Scholar] [CrossRef]
- Turner, B.G.; Summers, M.F. Structural biology of HIV-1. J. Mol. Biol. 1999, 285, 1–32. [Google Scholar] [CrossRef]
- Duvergé, A.; Negroni, M. Pseudotyping Lentiviral Vectors: When the Clothes Make the Virus. Viruses 2020, 12, 1311. [Google Scholar] [CrossRef]
- Carnell, G.W.; Ferrara, F.; Grehan, K.; Thompson, C.P.; Temperton, N.J. Pseudotype-based neutralization assays for influenza: A systematic analysis. Front. Immunol. 2015, 6, 161. [Google Scholar] [CrossRef]
- Toon, K.; Bentley, E.M.; Mattiuzzo, G. More Than Just Gene Therapy Vectors: Lentiviral Vector Pseudotypes for Serological Investigation. Viruses 2021, 13, 217. [Google Scholar] [CrossRef]
- Lerner, G.; Weaver, N.; Anokhin, B.; Spearman, P. Advances in HIV-1 Assembly. Viruses 2022, 14, 478. [Google Scholar] [CrossRef] [PubMed]
- Bona, R.; Michelini, Z.; Mazzei, C.; Gallinaro, A.; Canitano, A.; Borghi, M.; Vescio, M.F.; Di Virgilio, A.; Pirillo, M.F.; Klotman, M.E.; et al. Safety and efficiency modifications of SIV-based integrase-defective lentiviral vectors for immunization. Mol. Ther. Methods Clin. Dev. 2021, 23, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Sinn, P.L.; Burnight, E.R.; Hickey, M.A.; Blissard, G.W.; McCray, P.B. Persistent Gene Expression in Mouse Nasal Epithelia following Feline Immunodeficiency Virus-Based Vector Gene Transfer. J. Virol. 2005, 79, 12818–12827. [Google Scholar] [CrossRef]
- Millet, J.; Whittaker, G. Murine Leukemia Virus (MLV)-based Coronavirus Spike-pseudotyped Particle Production and Infection. Bio Protoc. 2016, 6, e2035. [Google Scholar] [CrossRef] [PubMed]
- Sandrin, V.; Muriaux, D.; Darlix, J.L.; Cosset, F.L. Intracellular Trafficking of Gag and Env Proteins and Their Interactions Modulate Pseudotyping of Retroviruses. J. Virol. 2004, 78, 7153–7164. [Google Scholar] [CrossRef]
- Maetzig, T.; Galla, M.; Baum, C.; Schambach, A. Gammaretroviral vectors: Biology, technology and application. Viruses 2011, 3, 677–713. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Cao, W.; Salawudeen, A.; Zhu, W.; Emeterio, K.; Safronetz, D.; Banadyga, L. Vesicular Stomatitis Virus: From Agricultural Pathogen to Vaccine Vector. Pathogens 2021, 10, 1092. [Google Scholar] [CrossRef]
- Whelan, S.P.; Ball, L.A.; Barr, J.N.; Wertz, G.T. Efficient recovery of infectious vesicular stomatitis virus entirely from cDNA clones. Proc. Natl. Acad. Sci. USA 1995, 92, 8388–8392. [Google Scholar] [CrossRef]
- Erlwein, O.; McClure, M.O. Progress and prospects: Foamy virus vectors enter a new age. Gene Ther. 2010, 17, 1423–1429. [Google Scholar] [CrossRef]
- Trobridge, G.D. Foamy virus vectors for gene transfer. Expert. Opin. Biol. Ther. 2009, 9, 1427–1436. [Google Scholar] [CrossRef]
- Cho, S.Y.; Kim, K.D.; Shin, C.G. Advances in foamy virus vector systems: Development and applications. Virology 2025, 601, 110270. [Google Scholar] [CrossRef]
- Vera-Velasco, N.M.; García-Murria, M.J.; Sánchez del Pino, M.M.; Mingarro, I.; Martinez-Gil, L. Proteomic composition of Nipah virus-like particles. J. Proteom. 2018, 172, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Syed, A.M.; Taha, T.Y.; Tabata, T.; Chen, I.P.; Ciling, A.; Khalid, M.M.; Sreekumar, B.; Chen, P.-Y.; Hayashi, J.M.; Soczek, K.M.; et al. Rapid assessment of SARS-CoV-2–evolved variants using virus-like particles. Science 2021, 374, 1626–1632. [Google Scholar] [CrossRef]
- Qing, M.; Liu, W.; Yuan, Z.; Gu, F.; Shi, P.Y. A high-throughput assay using dengue-1 virus-like particles for drug discovery. Antivir. Res. 2010, 86, 163–171. [Google Scholar] [CrossRef]
- Pierson, T.C.; Sánchez, M.D.; Puffer, B.A.; Ahmed, A.A.; Geiss, B.J.; Valentine, L.E.; Altamura, L.A.; Diamond, M.S.; Doms, R.W. A rapid and quantitative assay for measuring antibody-mediated neutralization of West Nile virus infection. Virology 2006, 346, 53–65. [Google Scholar] [CrossRef]
- Su, Y.; Chen, P.; Gao, F.; Bian, L.; Sun, S.; Dong, F.; Hu, Y.; Mao, Q.; Jiang, W.; Wu, X.; et al. A surrogate assay for measuring Coxsackievirus A6 neutralizing antibodies. Hum. Vaccin. Immunother. 2018, 14, 3034–3040. [Google Scholar] [CrossRef]
- Chen, P.; Wu, X.; Mao, Q.; Gao, F.; Hao, X.; Bian, L.; Zhu, F.; Li, W.; Xu, M.; Liang, Z. A rapid and quantitative assay for measuring neutralizing antibodies of Coxsackievirus B3. J. Virol. Methods 2016, 232, 1–7. [Google Scholar] [CrossRef]
- Chen, P.; Wu, X.; Su, Y.; Hao, X.; Mao, Q.; Liang, Z. Development of a pseudovirus based assay for measuring neutralizing antibodies against coxsackievirus B5. J. Virol. Methods 2017, 246, 21–26. [Google Scholar] [CrossRef]
- Tariq, H.; Batool, S.; Asif, S.; Ali, M.; Abbasi, B.H. Virus-Like Particles: Revolutionary Platforms for Developing Vaccines Against Emerging Infectious Diseases. Front. Microbiol. 2022, 12, 790121. [Google Scholar] [CrossRef]
- Kobinger, G.P.; Weiner, D.J.; Yu, Q.C.; Wilson, J.M. Filovirus-pseudotyped lentiviral vector can efficiently and stably transduce airway epithelia in vivo. Nat. Biotechnol. 2001, 19, 225–230. [Google Scholar] [CrossRef]
- Zhao, Y.; Ren, J.; Harlos, K.; Jones, D.M.; Zeltina, A.; Bowden, T.A.; Padilla-Parra, S.; Fry, E.E.; Stuart, D.I. Toremifene interacts with and destabilizes the Ebola virus glycoprotein. Nature 2016, 535, 169–172. [Google Scholar] [CrossRef]
- Luczkowiak, J.; Lasala, F.; Mora-Rillo, M.; Arribas, J.R.; Delgado, R. Broad Neutralizing Activity Against Ebolaviruses Lacking the Mucin-Like Domain in Convalescent Plasma Specimens From Patients With Ebola Virus Disease. J. Infect. Dis. 2018, 218 (Suppl. S5), S574–S581. [Google Scholar] [CrossRef]
- Seaman, M.S.; Janes, H.; Hawkins, N.; Grandpre, L.E.; Devoy, C.; Giri, A.; Coffey, R.T.; Harris, L.; Wood, B.; Daniels, M.G.; et al. Tiered Categorization of a Diverse Panel of HIV-1 Env Pseudoviruses for Assessment of Neutralizing Antibodies. J. Virol. 2010, 84, 1439–1452. [Google Scholar] [CrossRef]
- Solomon, M.; Liang, C. Pseudotyped Viruses for Retroviruses. In Pseudotyped Viruses; Springer: Singapore, 2023; pp. 61–84. [Google Scholar]
- Zhao, J.; Nie, J.; Jiao, Y.; Li, L.; Zhang, T.; Liu, Q.; Huang, W.; Wu, H.; Wang, Y. Effect of the maturation of neutralizing antibodies on human immunodeficiency virus (HIV) envelope evolution in HIV-infected subjects. Infect. Genet. Evol. 2016, 38, 82–89. [Google Scholar] [CrossRef]
- Nikolenko, G.N.; Svarovskaia, E.S.; Delviks, K.A.; Pathak, V.K. Antiretroviral Drug Resistance Mutations in Human Immunodeficiency Virus Type 1 Reverse Transcriptase Increase Template-Switching Frequency. J. Virol. 2004, 78, 8761–8770. [Google Scholar] [CrossRef]
- Hu, D.; Zhang, J.; Wang, H.; Liu, S.; Yu, L.; Sun, L.; Qu, Y.; Wang, T. Chikungunya Virus Glycoproteins Pseudotype with Lentiviral Vectors and Reveal a Broad Spectrum of Cellular Tropism. PLoS ONE 2014, 9, e110893. [Google Scholar] [CrossRef]
- Wu, J.; Zhao, C.; Liu, Q.; Huang, W.; Wang, Y. Development and application of a bioluminescent imaging mouse model for Chikungunya virus based on pseudovirus system. Vaccine 2017, 35, 6387–6394. [Google Scholar] [CrossRef]
- Haid, S.; Grethe, C.; Bankwitz, D.; Grunwald, T.; Pietschmann, T. Identification of a Human Respiratory Syncytial Virus Cell Entry Inhibitor by Using a Novel Lentiviral Pseudotype System. J. Virol. 2016, 90, 3065–3073. [Google Scholar] [CrossRef]
- Zost, S.J.; Gilchuk, P.; Case, J.B.; Binshtein, E.; Chen, R.E.; Nkolola, J.P.; Schäfer, A.; Reidy, J.X.; Trivette, A.; Nargi, R.S.; et al. Potently neutralizing and protective human antibodies against SARS-CoV-2. Nature 2020, 584, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.C.; Lyddon, T.D.; Suarez, R.; Salcedo, B.; Lepique, M.; Graham, M.; Ricana, C.; Robinson, C.; Ritter, D.G. Optimized Pseudotyping Conditions for the SARS-CoV-2 Spike Glycoprotein. J. Virol. 2020, 94, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Meng, B.; Abdullahi, A.; Ferreira, I.A.T.M.; Goonawardane, N.; Saito, A.; Kimura, I.; Yamasoba, D.; Gerber, P.P.; Fatihi, S.; Rathore, S.; et al. Altered TMPRSS2 usage by SARS-CoV-2 Omicron impacts infectivity and fusogenicity. Nature 2022, 603, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Cantoni, D.; Mayora-Neto, M.; Temperton, N. The role of pseudotype neutralization assays in understanding SARS CoV-2. Oxf. Open Immunol. 2021, 2, iqab005. [Google Scholar] [CrossRef]
- Yang, Y.; Du, L.; Liu, C.; Wang, L.; Ma, C.; Tang, J.; Baric, R.S.; Jiang, S.; Li, F. Receptor usage and cell entry of bat coronavirus HKU4 provide insight into bat-to-human transmission of MERS coronavirus. Proc. Natl. Acad. Sci. USA 2014, 111, 12516–12521. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Du, L.; Ma, C.; Li, Y.; Li, L.; Poon, V.K.; Wang, L.; Yu, F.; Zheng, B.-J.; Jiang, S.; et al. A safe and convenient pseudovirus-based inhibition assay to detect neutralizing antibodies and screen for viral entry inhibitors against the novel human coronavirus MERS-CoV. Virol. J. 2013, 10, 266. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, R.; Forconi, V.; Molesti, E.; Semplici, C.; Piu, P.; Altamura, M.; Dapporto, F.; Temperton, N.; Montomoli, E.; Manenti, A. A validate and standardized pseudotyped microneutralization assay as a safe and powerful tool to measure LASSA neutralising antibodies for vaccine development and comparison. F1000Res 2024, 13, 534. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.P.; Hsieh, S.C.; King, C.C.; Wang, W.K. Characterization of retrovirus-based reporter viruses pseudotyped with the precursor membrane and envelope glycoproteins of four serotypes of dengue viruses. Virology 2007, 368, 376–387. [Google Scholar] [CrossRef]
- Pan, T.; Peng, Z.; Tan, L.; Zou, F.; Zhou, N.; Liu, B.; Liang, L.; Chen, C.; Liu, J.; Wu, L.; et al. Nonsteroidal Anti-inflammatory Drugs Potently Inhibit the Replication of Zika Viruses by Inducing the Degradation of AXL. J. Virol. 2018, 92, 10–1128. [Google Scholar] [CrossRef]
- Kretschmer, M.; Kadlubowska, P.; Hoffmann, D.; Schwalbe, B.; Auerswald, H.; Schreiber, M. Zikavirus prME Envelope Pseudotyped Human Immunodeficiency Virus Type-1 as a Novel Tool for Glioblastoma-Directed Virotherapy. Cancers 2020, 12, 1000. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, Y.; Wang, C.; Zheng, X.; Wang, H.; Gai, W.; Jin, H.; Yan, F.; Qiu, B.; Gao, Y.; et al. Packaging of Rift Valley fever virus pseudoviruses and establishment of a neutralization assay method. J. Vet. Sci. 2018, 19, 200. [Google Scholar] [CrossRef]
- Federici, T.; Kutner, R.; Zhang, X.-Y.; Kuroda, H.; Tordo, N.; Boulis, N.M.; Reiser, J. Comparative analysis of HIV-1-based lentiviral vectors bearing lyssavirus glycoproteins for neuronal gene transfer. Genet. Vaccines Ther. 2009, 7, 1. [Google Scholar] [CrossRef]
- Wu, J.; Cao, S.; Lei, S.; Liu, Q.; Li, Y.; Yu, Y.; Xie, H.; Li, Q.; Zhao, X.; Chen, R.; et al. Clofazimine: A Promising Inhibitor of Rabies Virus. Front. Pharmacol. 2021, 12, 598241. [Google Scholar] [CrossRef]
- Wright, E.; McNabb, S.; Goddard, T.; Horton, D.L.; Lembo, T.; Nel, L.H.; Weiss, R.A.; Cleaveland, S.; Fooks, A.R. A robust lentiviral pseudotype neutralisation assay for in-field serosurveillance of rabies and lyssaviruses in Africa. Vaccine 2009, 27, 7178–7186. [Google Scholar] [CrossRef]
- Nie, J.; Wu, X.; Ma, J.; Cao, S.; Huang, W.; Liu, Q.; Li, X.; Li, Y.; Wang, Y. Development of in vitro and in vivo rabies virus neutralization assays based on a high-titer pseudovirus system. Sci. Rep. 2017, 7, 42769. [Google Scholar] [CrossRef]
- Wright, E.; Temperton, N.J.; Marston, D.A.; McElhinney, L.M.; Fooks, A.R.; Weiss, R.A. Investigating antibody neutralization of lyssaviruses using lentiviral pseudotypes: A cross-species comparison. J. Gen. Virol. 2008, 89 Pt 9, 2204–2213. [Google Scholar] [CrossRef]
- Both, L.; Dolleweerd, C.; Wright, E.; Banyard, A.C.; Bulmer-Thomas, B.; Selden, D.; Altmann, F.; Fooks, A.R.; Ma, J.K. Production, characterization, and antigen specificity of recombinant 62-71-3, a candidate monoclonal antibody for rabies prophylaxis in humans. FASEB J. 2013, 27, 2055–2065. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Zhou, M.; He, Y.; Wan, Y.; Bai, W.; Tao, S.; Ren, Y.; Zhang, X.; Xu, J.; Liu, J.; et al. Efficient Inhibition of Hepatitis B Virus Infection by a preS1-binding Peptide. Sci. Rep. 2016, 6, 29391. [Google Scholar] [CrossRef] [PubMed]
- Del Rosario, J.M.M.; da Costa, K.A.S.; Temperton, N.J. Pseudotyped Viruses for Influenza. In Pseudotyped Viruses; Springer: Singapore, 2023; pp. 153–173. [Google Scholar]
- Chan, E.; Heilek-Snyder, G.; Cammack, N.; Sankuratri, S.; Ji, C. Development of a Moloney Murine Leukemia Virus–Based Pseudotype Anti-HIV Assay Suitable for Accurate and Rapid Evaluation of HIV Entry Inhibitors. SLAS Discov. 2006, 11, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Urbanowicz, R.A.; McClure, C.P.; Sakuntabhai, A.; Sall, A.A.; Kobinger, G.; Müller, M.A.; Holmes, E.C.; Rey, F.A.; Simon-Loriere, E.; Ball, J.K. Human Adaptation of Ebola Virus during the West African Outbreak. Cell 2016, 167, 1079–1087.e5. [Google Scholar] [CrossRef]
- Kuhn, J.H.; Radoshitzky, S.R.; Guth, A.C.; Warfield, K.L.; Li, W.; Vincent, M.J.; Towner, J.S.; Nichol, S.T.; Bavari, S.; Choe, H.; et al. Conserved Receptor-binding Domains of Lake Victoria Marburgvirus and Zaire Ebolavirus Bind a Common Receptor. J. Biol. Chem. 2006, 281, 15951–15958. [Google Scholar] [CrossRef]
- Wang, M.K.; Lim, S.Y.; Lee, S.M.; Cunningham, J.M. Biochemical Basis for Increased Activity of Ebola Glycoprotein in the 2013–16 Epidemic. Cell Host Microbe 2017, 21, 367–375. [Google Scholar] [CrossRef]
- Yang, Z.Y.; Delgado, R.; Xu, L.; Todd, R.F.; Nabel, E.G.; Sanchez, A.; Nabel, G.J. Distinct Cellular Interactions of Secreted and Transmembrane Ebola Virus Glycoproteins. Science 1998, 279, 1034–1037. [Google Scholar] [CrossRef]
- Millet, J.K.; Tang, T.; Nathan, L.; Jaimes, J.A.; Hsu, H.-L.; Daniel, S.; Whittaker, G.R. Production of Pseudotyped Particles to Study Highly Pathogenic Coronaviruses in a Biosafety Level 2 Setting. J. Vis. Exp. 2019, 145, 10-3791. [Google Scholar] [CrossRef]
- Steidl, S.; Stitz, J.; Schmitt, I.; König, R.; Flory, E.; Schweizer, M.; Cichutek, K. Coreceptor Switch of [MLV(SIVagm)] Pseudotype Vectors by V3-Loop Exchange. Virology 2002, 300, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Denolly, S.; Cosset, F.L.; Freitas, N. Membrane Fusion Assays for Studying Entry Hepatitis C Virus into Cells. In Hepatitis C Virus Protocols; Humana Press: New York, NY, USA, 2019; pp. 219–234. [Google Scholar]
- Riva, L.; Dubuisson, J. Similarities and Differences Between HCV Pseudoparticle (HCVpp) and Cell Culture HCV (HCVcc) in the Study of HCV. In Hepatitis C Virus Protocols; Humana Press: New York, NY, USA, 2019; pp. 33–45. [Google Scholar]
- Mosa, A.I.; Urbanowicz, R.A.; AbouHaidar, M.G.; Tavis, J.E.; Ball, J.K.; Feld, J.J. A bivalent HCV peptide vaccine elicits pan-genotypic neutralizing antibodies in mice. Vaccine 2020, 38, 6864–6867. [Google Scholar] [CrossRef]
- Anderson, E.M.; Coller, B.A.G. Translational success of fundamental virology: A VSV-vectored Ebola vaccine. J. Virol. 2024, 98. [Google Scholar] [CrossRef] [PubMed]
- Steeds, K.; Hall, Y.; Slack, G.S.; Longet, S.; Strecker, T.; Fehling, S.K.; Wright, E.; Bore, J.A.; Koundouno, F.R.; Konde, M.K.; et al. Pseudotyping of VSV with Ebola virus glycoprotein is superior to HIV-1 for the assessment of neutralising antibodies. Sci. Rep. 2020, 10, 14289. [Google Scholar] [CrossRef]
- Konduru, K.; Shurtleff, A.C.; Bavari, S.; Kaplan, G. High degree of correlation between Ebola virus BSL-4 neutralization assays and pseudotyped VSV BSL-2 fluorescence reduction neutralization test. J. Virol. Methods 2018, 254, 1–7. [Google Scholar] [CrossRef]
- Takada, A.; Robison, C.; Goto, H.; Sanchez, A.; Murti, K.G.; Whitt, M.A.; Kawaoka, Y. A system for functional analysis of Ebola virus glycoprotein. Proc. Natl. Acad. Sci. USA 1997, 94, 14764–14769. [Google Scholar] [CrossRef]
- Aljofan, M.; Sganga, M.L.; Lo, M.K.; Rootes, C.L.; Porotto, M.; Meyer, A.G.; Saubern, S.; Moscona, A.; Mungall, B.A. Antiviral activity of gliotoxin, gentian violet and brilliant green against Nipah and Hendra virus in vitro. Virol. J. 2009, 6, 187. [Google Scholar] [CrossRef]
- Negrete, O.A.; Chu, D.; Aguilar, H.C.; Lee, B. Single Amino Acid Changes in the Nipah and Hendra Virus Attachment Glycoproteins Distinguish EphrinB2 from EphrinB3 Usage. J. Virol. 2007, 81, 10804–10814. [Google Scholar] [CrossRef]
- Kaku, Y.; Noguchi, A.; Marsh, G.A.; McEachern, J.A.; Okutani, A.; Hotta, K.; Bazartseren, B.; Fukushi, S.; Broder, C.C.; Yamada, A.; et al. A neutralization test for specific detection of Nipah virus antibodies using pseudotyped vesicular stomatitis virus expressing green fluorescent protein. J. Virol. Methods 2009, 160, 7–13. [Google Scholar] [CrossRef]
- Vasmehjani, A.A.; Salehi-Vaziri, M.; Azadmanesh, K.; Nejati, A.; Pouriayevali, M.H.; Gouya, M.M.; Parsaeian, M.; Shahmahmoodi, S. Efficient production of a lentiviral system for displaying Crimean-Congo hemorrhagic fever virus glycoproteins reveals a broad range of cellular susceptibility and neutralization ability. Arch. Virol. 2020, 165, 1109–1120. [Google Scholar] [CrossRef]
- Tong, W.; Yin, X.X.; Lee, B.J.; Li, Y.G. Preparation of vesicular stomatitis virus pseudotype with Chikungunya virus envelope protein. Acta Virol. 2015, 59, 189–193. [Google Scholar] [CrossRef]
- Schlereth, B.; Buonocore, L.; Tietz, A.; Meulen Vter Rose, J.K.; Niewiesk, S. Successful mucosal immunization of cotton rats in the presence of measles virus-specific antibodies depends on degree of attenuation of vaccine vector and virus dose. J. Gen. Virol. 2003, 84, 2145–2151. [Google Scholar] [CrossRef]
- Zimmer, G.; Locher, S.; Berger Rentsch, M.; Halbherr, S.J. Pseudotyping of vesicular stomatitis virus with the envelope glycoproteins of highly pathogenic avian influenza viruses. J. Gen. Virol. 2014, 95, 1634–1639. [Google Scholar] [CrossRef] [PubMed]
- Rose, N.F.; Marx, P.A.; Luckay, A.; Nixon, D.F.; Moretto, W.J.; Donahoe, S.M.; Montefiori, D.; Roberts, A.; Buonocore, L.; Rose, J.K. An Effective AIDS Vaccine Based on Live Attenuated Vesicular Stomatitis Virus Recombinants. Cell 2001, 106, 539–549. [Google Scholar] [CrossRef]
- Hofmann, H.; Li, X.; Zhang, X.; Liu, W.; Kühl, A.; Kaup, F.; Soldan, S.S.; González-Scarano, F.; Weber, F.; He, Y.; et al. Severe Fever with Thrombocytopenia Virus Glycoproteins Are Targeted by Neutralizing Antibodies and Can Use DC-SIGN as a Receptor for pH-Dependent Entry into Human and Animal Cell Lines. J. Virol. 2013, 87, 4384–4394. [Google Scholar] [CrossRef] [PubMed]
- Tani, H.; Shimojima, M.; Fukushi, S.; Yoshikawa, T.; Fukuma, A.; Taniguchi, S.; Morikawa, S.; Saijo, M. Characterization of Glycoprotein-Mediated Entry of Severe Fever with Thrombocytopenia Syndrome Virus. J. Virol. 2016, 90, 5292–5301. [Google Scholar] [CrossRef]
- Dong, F.; Li, D.; Wen, D.; Li, S.; Zhao, C.; Qi, Y.; Jangra, R.K.; Wu, C.; Xia, D.; Zhang, X.; et al. Single dose of a rVSV-based vaccine elicits complete protection against severe fever with thrombocytopenia syndrome virus. NPJ Vaccines 2019, 4, 5. [Google Scholar] [CrossRef] [PubMed]
- Krajden, M.; Cook, D.; Yu, A.; Chow, R.; Su, Q.; Mei, W.; McNeil, S.; Money, D.; Dionne, M.; Palefsky, J.; et al. Assessment of HPV 16 and HPV 18 antibody responses by pseudovirus neutralization, Merck cLIA and Merck total IgG LIA immunoassays in a reduced dosage quadrivalent HPV vaccine trial. Vaccine 2014, 32, 624–630. [Google Scholar] [CrossRef]
- WHO. WHO Expert Committee on Biological Standardization: Recommendations to Assure the Quality, Safety and Efficacy of Recombinant Human Papillomavirus Virus-Like Particle Vaccines; World Health Organization Technical Report Series; WHO: Geneva, Switzerland, 2016; pp. 1–267. [Google Scholar]
- Arita, M.; Iwai-Itamochi, M. Evaluation of antigenic differences between wild and Sabin vaccine strains of poliovirus using the pseudovirus neutralization test. Sci. Rep. 2019, 9, 11970. [Google Scholar] [CrossRef] [PubMed]
- Cantoni, D.; Wilkie, C.; Bentley, E.M.; Mayora-Neto, M.; Wright, E.; Scott, S.; Ray, S.; Castillo-Olivares, J.; Heeney, J.L.; Mattiuzzo, G.; et al. Correlation between pseudotyped virus and authentic virus neutralisation assays, a systematic review and meta-analysis of the literature. Front. Immunol. 2023, 14, 1184362. [Google Scholar] [CrossRef]
- Del Rosario, J.M.M.; da Costa, K.A.S.; Asbach, B.; Ferrara, F.; Ferrari, M.; Wells, D.A.; Mann, G.S.; Ameh, V.O.; Sabeta, C.T.; Banyard, A.C.; et al. Exploiting Pan Influenza A and Pan Influenza B Pseudotype Libraries for Efficient Vaccine Antigen Selection. Vaccines 2021, 9, 741. [Google Scholar] [CrossRef]
- Sampson, A.T.; Heeney, J.; Cantoni, D.; Ferrari, M.; Sans, M.S.; George, C.; Di Genova, C.; Neto, M.M.; Einhauser, S.; Asbach, B.; et al. Coronavirus Pseudotypes for All Circulating Human Coronaviruses for Quantification of Cross-Neutralizing Antibody Responses. Viruses 2021, 13, 1579. [Google Scholar] [CrossRef] [PubMed]
- Schoofs, T.; Klein, F.; Braunschweig, M.; Kreider, E.F.; Feldmann, A.; Nogueira, L.; Oliveira, T.; Lorenzi, J.C.C.; Parrish, E.H.; Learn, G.H.; et al. HIV-1 therapy with monoclonal antibody 3BNC117 elicits host immune responses against HIV-1. Science 2016, 352, 997–1001. [Google Scholar] [CrossRef]
- Steffen, I.; Lu, K.; Yamamoto, L.K.; Hoff, N.A.; Mulembakani, P.; Wemakoy, E.O.; Muyembe-Tamfum, J.-J.; Ndembi, N.; Brennan, C.A.; Hackett, J.; et al. Serologic Prevalence of Ebola Virus in Equatorial Africa. Emerg. Infect. Dis. 2019, 25, 911–918. [Google Scholar] [CrossRef]
- Steffen, I.; Lu, K.; Hoff, N.A.; Mulembakani, P.; Wemakoy, E.O.; Muyembe-Tamfum, J.-J.; Ndembi, N.; Brennan, C.A.; Hackett, J.; Switzer, W.M.; et al. Seroreactivity against Marburg or related filoviruses in West and Central Africa. Emerg. Microbes Infect. 2020, 9, 124–128. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Accardi, G.; Aiello, A.; Calabrò, A.; Caruso, C.; Ligotti, M.E.; Marchi, S.; Montomoli, E.; Neto, M.M.; Temperton, N.; et al. Centenarians, semi and supercentenarians, COVID-19 and Spanish flu: A serological assessment to gain insight into the resilience of older centenarians to COVID-19. Immun. Ageing 2024, 21, 44. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.; Park, Y.J.; Beltramello, M.; Walls, A.C.; Tortorici, M.A.; Bianchi, S.; Jaconi, S.; Culap, K.; Zatta, F.; De Marco, A.; et al. Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody. Nature 2020, 583, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wu, J.; Nie, J.; Zhang, L.; Hao, H.; Liu, S.; Zhao, C.; Zhang, Q.; Liu, H.; Nie, L.; et al. The Impact of Mutations in SARS-CoV-2 Spike on Viral Infectivity and Antigenicity. Cell 2020, 182, 1284–1294.e9. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Das, R.; Dijokaite-Guraliuc, A.; Zhou, D.; Mentzer, A.J.; Supasa, P.; Selvaraj, M.; Duyvesteyn, H.M.E.; Ritter, T.G.; Temperton, N.; et al. Emerging variants develop total escape from potent monoclonal antibodies induced by BA.4/5 infection. Nat. Commun. 2024, 15, 3284. [Google Scholar] [CrossRef]
- Collier, D.A.; De Marco, A.; Ferreira, I.A.T.M.; Meng, B.; Datir, R.P.; Walls, A.C.; Kemp, S.A.; Bassi, J.; Pinto, D.; Silacci-Fregni, C.; et al. Sensitivity of SARS-CoV-2 B.1.1.7 to mRNA vaccine-elicited antibodies. Nature 2021, 593, 136–141. [Google Scholar] [CrossRef]
- Palmer, C.D.; Scallan, C.D.; Tardif, L.D.K.; Kachura, M.A.; Rappaport, A.R.; Koralek, D.O.; Uriel, A.; Gitlin, L.; Klein, J.; Davis, M.J.; et al. GRT-R910: A self-amplifying mRNA SARS-CoV-2 vaccine boosts immunity for ≥6 months in previously-vaccinated older adults. Nat. Commun. 2023, 14, 3274. [Google Scholar] [CrossRef]
- Chen, G.-L.; Li, X.-F.; Dai, X.-H.; Li, N.; Cheng, M.-L.; Huang, Z.; Shen, J.; Ge, Y.-H.; Shen, Z.-W.; Deng, Y.-Q.; et al. Safety and immunogenicity of the SARS-CoV-2 ARCoV mRNA vaccine in Chinese adults: A randomised, double-blind, placebo-controlled, phase 1 trial. Lancet Microbe 2022, 3, e193–e202. [Google Scholar] [CrossRef]
- Zhang, Y.; Zeng, G.; Pan, H.; Li, C.; Hu, Y.; Chu, K.; Han, W.; Chen, Z.; Tang, R.; Yin, W.; et al. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine in healthy adults aged 18-59 years: A randomised, double-blind, placebo-controlled, phase 1/2 clinical trial. Lancet Infect. Dis. 2021, 21, 181–192. [Google Scholar] [CrossRef]
- Paciello, I.; Pierleoni, G.; Pantano, E.; Antonelli, G.; Pileri, P.; Maccari, G.; Cardamone, D.; Realini, G.; Perrone, F.; Neto, M.M.; et al. Antigenic sin and multiple breakthrough infections drive converging evolution of COVID-19 neutralizing responses. Cell Rep. 2024, 43, 114645. [Google Scholar] [CrossRef] [PubMed]
- Roozendaal, R.; Hendriks, J.; van Effelterre, T.; Spiessens, B.; Dekking, L.; Solforosi, L.; Czapska-Casey, D.; Bockstal, V.; Stoop, J.; Splinter, D.; et al. Nonhuman primate to human immunobridging to infer the protective effect of an Ebola virus vaccine candidate. NPJ Vaccines 2020, 5, 112. [Google Scholar] [CrossRef] [PubMed]
- Gilchuk, P.; Murin, C.D.; Cross, R.W.; Ilinykh, P.A.; Huang, K.; Kuzmina, N.; Borisevich, V.; Agans, K.N.; Geisbert, J.B.; Zost, S.J.; et al. Pan-ebolavirus protective therapy by two multifunctional human antibodies. Cell 2021, 184, 5593–5607.e18. [Google Scholar] [CrossRef]
- Corti, D.; Misasi, J.; Mulangu, S.; Stanley, D.A.; Kanekiyo, M.; Wollen, S.; Ploquin, A.; Doria-Rose, N.A.; Staupe, R.P.; Bailey, M.; et al. Protective monotherapy against lethal Ebola virus infection by a potently neutralizing antibody. Science 2016, 351, 1339–1342. [Google Scholar] [CrossRef] [PubMed]
- Montefiori, D.C. Measuring HIV neutralization in a luciferase reporter gene assay. Methods Mol. Biol. 2009, 485, 395–405. [Google Scholar]
- Laher, F.; Moodie, Z.; Cohen, K.W.; Grunenberg, N.; Bekker, L.G.; Allen, M.; Yates, N.L.; Morris, L.; Malahleha, M.; Mngadi, K.; et al. Safety and immune responses after a 12-month booster in healthy HIV-uninfected adults in HVTN 100 in South Africa: A randomized double-blind placebo-controlled trial of ALVAC-HIV (vCP2438) and bivalent subtype C gp120/MF59 vaccines. PLoS Med. 2020, 17, e1003038. [Google Scholar] [CrossRef]
- Lee, H.J.; Choi, H.; Park, K.H.; Jang, Y.; Hong, Y.J.; Kim, Y.B. Retention of neutralizing antibodies to Japanese encephalitis vaccine in age groups above fifteen years in Korea. Int. J. Infect. Dis. 2020, 100, 53–58. [Google Scholar] [CrossRef]
- Bellier, B.; Huret, C.; Miyalou, M.; Desjardins, D.; Frenkiel, M.-P.; Despres, P.; Tangy, F.; Dalba, C.; Klatzmann, D. DNA vaccines expressing retrovirus-like particles are efficient immunogens to induce neutralizing antibodies. Vaccine 2009, 27, 5772–5780. [Google Scholar] [CrossRef]
- Rao, S.; Erku, D.; Mahalingam, S.; Taylor, A. Immunogenicity, safety and duration of protection afforded by chikungunya virus vaccines undergoing human clinical trials. J. Gen. Virol. 2024, 105, 001965. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Perini, D.; Mather, S.; Temperton, N.; Montomoli, E. Overview of Serological Techniques for Influenza Vaccine Evaluation: Past, Present and Future. Vaccines 2014, 2, 707–734. [Google Scholar] [CrossRef]
- Moss, B.; Smith, G.L.; Gerin, J.L.; Purcell, R.H. Live recombinant vaccinia virus protects chimpanzees against hepatitis B. Nature 1984, 311, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Vrba, S.M.; Kirk, N.M.; Brisse, M.E.; Liang, Y.; Ly, H. Development and Applications of Viral Vectored Vaccines to Combat Zoonotic and Emerging Public Health Threats. Vaccines 2020, 8, 680. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.K.; Hendry, R.M.; Singh, V.; Rose, J.K.; Seligman, S.J.; Klug, B.; Kochhar, S.; Mac, L.M.; Carbery, B.; Chen, R.T. Live virus vaccines based on a vesicular stomatitis virus (VSV) backbone: Standardized template with key considerations for a risk/benefit assessment. Vaccine 2016, 34, 6597–6609. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, J.D.; Frank, I.; Elizaga, M.L.; Allen, M.; Frahm, N.; Kochar, N.; Li, S.; Edupuganti, S.; Kalams, S.A.; Tomaras, G.D.; et al. First-in-Human Evaluation of the Safety and Immunogenicity of a Recombinant Vesicular Stomatitis Virus Human Immunodeficiency Virus-1 gag Vaccine (HVTN 090). Open Forum Infect. Dis. 2015, 2, ofv082. [Google Scholar] [CrossRef]
- Yahalom-Ronen, Y.; Erez, N.; Fisher, M.; Tamir, H.; Politi, B.; Achdout, H.; Melamed, S.; Glinert, I.; Weiss, S.; Cohen-Gihon, I.; et al. Neutralization of SARS-CoV-2 Variants by rVSV-ΔG-Spike-Elicited Human Sera. Vaccines 2022, 10, 291. [Google Scholar] [CrossRef]
- Robbins, J.A.; Tait, D.; Huang, Q.; Dubey, S.; Crumley, T.; Cote, J.; Luk, J.; Sachs, J.R.; Rutkowski, K.; Park, H.; et al. Safety and immunogenicity of intramuscular, single-dose V590 (rVSV-SARS-CoV-2 Vaccine) in healthy adults: Results from a phase 1 randomised, double-blind, placebo-controlled, dose-ranging trial. EBioMedicine 2022, 82, 104138. [Google Scholar] [CrossRef]
- Basu, A.; Mills, D.M.; Bowlin, T.L. High-throughput screening of viral entry inhibitors using pseudotyped virus. Curr. Protoc. Pharmacol. 2010, 51, 13B-3. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, K.; Guo, Y. The antifungal isavuconazole inhibits the entry of lassa virus by targeting the stable signal peptide-GP2 subunit interface of lassa virus glycoprotein. Antivir. Res. 2020, 174, 104701. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Q.; Li, Q.; Li, Y.; Liu, Z.; Deng, H.; Tang, S.; Wang, Y.; Wang, Y.; Song, D. Synthesis and biological evaluation of novel tricyclic matrinic derivatives as potential anti-filovirus agents. Acta Pharm. Sin. B 2018, 8, 629–638. [Google Scholar] [CrossRef]
- Côté, M.; Misasi, J.; Ren, T.; Bruchez, A.; Lee, K.; Filone, C.M.; Hensley, L.; Li, Q.; Ory, D.; Chandran, K.; et al. Small molecule inhibitors reveal Niemann–Pick C1 is essential for Ebola virus infection. Nature 2011, 477, 344–348. [Google Scholar] [CrossRef]
- Chong, H.; Zhu, Y.; Yu, D.; He, Y. Structural and Functional Characterization of Membrane Fusion Inhibitors with Extremely Potent Activity against Human Immunodeficiency Virus Type 1 (HIV-1), HIV-2, and Simian Immunodeficiency Virus. J. Virol. 2018, 92, 10–1128. [Google Scholar] [CrossRef]
- Saladini, F.; Giannini, A.; Giammarino, F.; Maggiolo, F.; Vichi, F.; Corbelli, G.M.; Galli, A.; Bigoloni, A.; Poli, A.; Santoro, M.M.; et al. In vitro susceptibility to fostemsavir is not affected by long-term exposure to antiviral therapy in MDR HIV-1-infected patients. J. Antimicrob. Chemother. 2020, 75, 2547–2553. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Maeda, Y.; Terasawa, H.; Monde, K.; Harada, S.; Yusa, K. A combination of polymorphic mutations in V3 loop of HIV-1 gp120 can confer noncompetitive resistance to maraviroc. Virology 2011, 413, 293–299. [Google Scholar] [CrossRef] [PubMed]
- An, T.; Ouyang, W.; Pan, W.; Guo, D.; Li, J.; Li, L.; Chen, G.; Yang, J.; Wu, S.; Tien, P. Amino acid derivatives of the (−) enantiomer of gossypol are effective fusion inhibitors of human immunodeficiency virus type 1. Antivir. Res. 2012, 94, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.W.; Yuan, S.; Chu, H.; Sridhar, S.; Yuen, K.Y. COVID-19 drug discovery and treatment options. Nat. Rev. Microbiol. 2024, 22, 391–407. [Google Scholar] [CrossRef]
- Huang, L.; Zhu, L.; Xie, H.; Goodwin, J.S.; Rana, T.; Xie, L.; Chen, C.-H. Quinolizidines as Novel SARS-CoV-2 Entry Inhibitors. Int. J. Mol. Sci. 2022, 23, 9659. [Google Scholar] [CrossRef]
- Yang, L.; Pei, R.-J.; Li, H.; Ma, X.-N.; Zhou, Y.; Zhu, F.-H.; He, P.-L.; Tang, W.; Zhang, Y.-C.; Xiong, J.; et al. Identification of SARS-CoV-2 entry inhibitors among already approved drugs. Acta Pharmacol. Sin. 2021, 42, 1347–1353. [Google Scholar] [CrossRef]
- Zhang, L.; Jackson, C.B.; Mou, H.; Ojha, A.; Peng, H.; Quinlan, B.D.; Rangarajan, E.S.; Pan, A.; Vanderheiden, A.; Suthar, M.S.; et al. SARS-CoV-2 spike-protein D614G mutation increases virion spike density and infectivity. Nat. Commun. 2020, 11, 6013. [Google Scholar] [CrossRef] [PubMed]
- Paciello, I.; Maccari, G.; Pantano, E.; Andreano, E.; Rappuoli, R. High-resolution map of the Fc functions mediated by COVID-19-neutralizing antibodies. Proc. Natl. Acad. Sci. USA 2024, 121, e2314730121. [Google Scholar] [CrossRef] [PubMed]
- Azizi, A.; Manak, M.; Bernasconi, V. The CEPI centralized laboratory network for COVID-19 will help prepare for future outbreaks. Nat. Med. 2023, 29, 2684–2685. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos-Cela, M.; Forconi, V.; Antonelli, R.; Manenti, A.; Montomoli, E. Exploring the Use of Viral Vectors Pseudotyped with Viral Glycoproteins as Tools to Study Antibody-Mediated Neutralizing Activity. Microorganisms 2025, 13, 1785. https://doi.org/10.3390/microorganisms13081785
Ramos-Cela M, Forconi V, Antonelli R, Manenti A, Montomoli E. Exploring the Use of Viral Vectors Pseudotyped with Viral Glycoproteins as Tools to Study Antibody-Mediated Neutralizing Activity. Microorganisms. 2025; 13(8):1785. https://doi.org/10.3390/microorganisms13081785
Chicago/Turabian StyleRamos-Cela, Miguel, Vittoria Forconi, Roberta Antonelli, Alessandro Manenti, and Emanuele Montomoli. 2025. "Exploring the Use of Viral Vectors Pseudotyped with Viral Glycoproteins as Tools to Study Antibody-Mediated Neutralizing Activity" Microorganisms 13, no. 8: 1785. https://doi.org/10.3390/microorganisms13081785
APA StyleRamos-Cela, M., Forconi, V., Antonelli, R., Manenti, A., & Montomoli, E. (2025). Exploring the Use of Viral Vectors Pseudotyped with Viral Glycoproteins as Tools to Study Antibody-Mediated Neutralizing Activity. Microorganisms, 13(8), 1785. https://doi.org/10.3390/microorganisms13081785