

Co-Inoculation Between Bacteria and Algae from Biological Soil Crusts and Their Effects on the Growth of Poa annua and Sandy Soils Quality
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Materials
2.2. Isolation of Functional Bacteria
2.3. Isolation of Algae
2.4. Preparation of the Inoculation Solutions
2.5. Design of Experiment
2.6. Soil Physicochemical Measurements
2.7. Microbial Biomass Measurements
2.8. Poa annua Growth and Physicochemical Measurements
2.9. Calculation of Soil Composite Index
2.10. Statistical Analysis
3. Results
3.1. Impact of Bacteria–Algae Co-Inoculation on Physicochemical Properties of Sandy Soils
3.2. Impact of Bacteria–Algal Co-Inoculation on Composition and Stability of Sandy Soils Aggregates
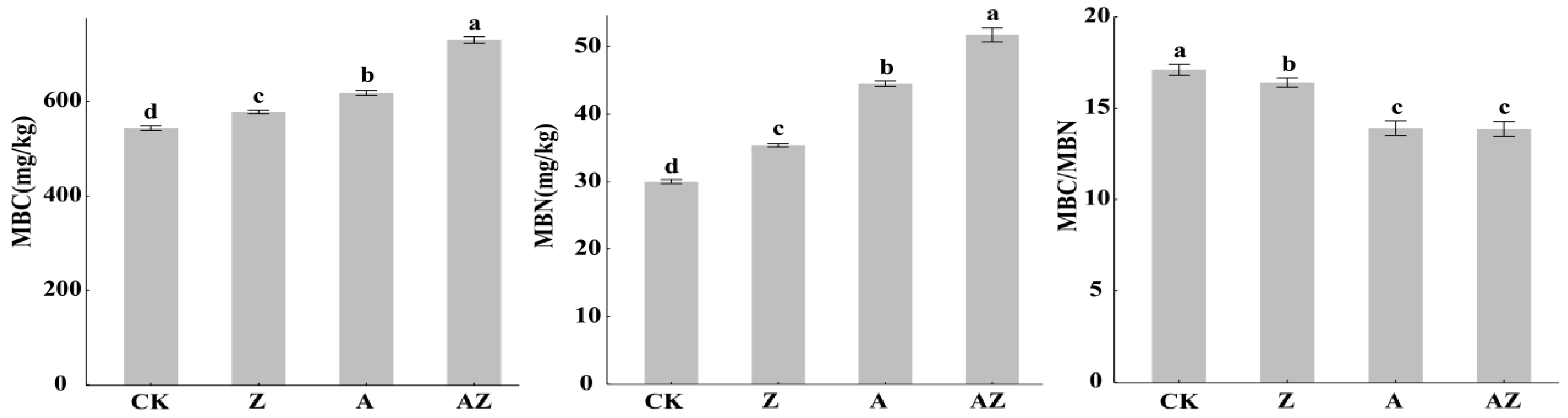
3.3. Impact of Bacteria–Algal Co-Inoculation on Microbial Biomass
3.4. Impact of Soil Physicochemical Properties on Aggregate Stability and Microbial Biomass
3.5. Impact of Bacteria–Alga Co-Inoculation on the Growth of Poa annua
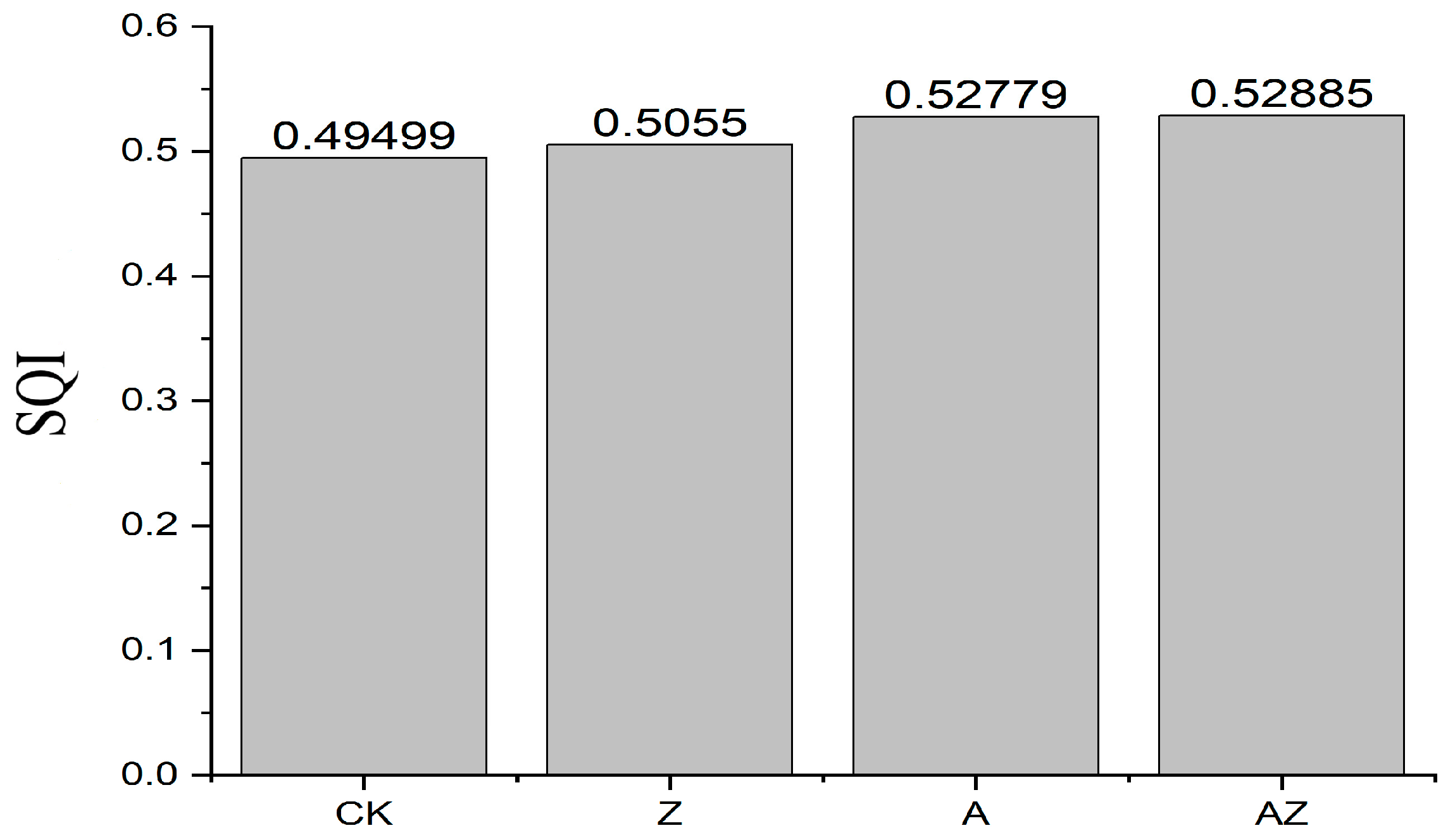
3.6. Comprehensive Index of Soil Quality Under the Combined Inoculation of Bacteria and Algae
4. Discussion
4.1. Influencing Mechanism of Bacteria–Algal Co-Inoculation on Soil Physicochemical Properties
4.2. Influencing Mechanism of Bacteria–Algal Co-Inoculation on the Formation and Stability of Aggregates
4.3. Influencing Mechanism of Bacteria–Algal Co-Inoculation on Microbial Biomass
4.4. Promoting Effect of Bacteria–Algal Co-Inoculation on Poa annua, Growth
4.5. Comprehensive Evaluation of Soil Quality Under the Bacteria–Algal Co-Inoculation
5. Conclusions
6. Patents
- Peng Xia-Wei, Chen Man, SUN Xiao-Yu et al. A strain of Streptomyces fusiformis LS159 and its application: CN202211465909.5[P]. 2023-02-03.
- Zhou Jinxing, Chen Man, GUAN Yinghui et al. A new soil improvement microbial agent, preparation method and application: CN202210974181.2[P]. 2022-12-16.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, J.; Zhang, J.; Zhu, X.; Miao, T.; Hou, H.; Chen, G. The Effects of Microbial Composite Fertilizer CAMP on Soil Improvement and Growth of Chinese Cabbage. North. Hortic. 2018, 12, 112–118. [Google Scholar]
- Zhao, Y.; Wang, Z.; Yang, L.; Peng, D.; Xiang, D.; Fu, C.; Liu, Y.; Qin, X.; Qin, H.; Li, Q. Effects of Microalgae Combined with Bacillus subtilis on Soil Chemical Properties and Bacterial Communities in Tobacco Fields. J. Jiangxi Agric. Univ. 2024, 46, 241–250. [Google Scholar]
- Cheng, C.; Gao, M.; Zhang, Y.; Long, M.; Wu, Y.; Li, X. Effects of disturbance to moss biocrusts on soil nutrients, enzyme activities, and microbial communities in degraded karst landscapes in southwest China. Soil Biol. Biochem. 2021, 152, 108065. [Google Scholar] [CrossRef]
- Weber, B.; Belnap, J.; Büdel, B.; Antoninka, A.J.; Barger, N.N.; Chaudhary, V.B.; Darrouzet-Nardi, A.; Eldridge, D.J.; Faist, A.M.; Ferrenberg, S.; et al. What is a biocrust? A refined, contemporary definition for a broadening research community. Biol. Rev. 2022, 97, 1768–1785. [Google Scholar] [CrossRef] [PubMed]
- Gundlapally, S.R.; Garcia-Pichel, F. The community and phylogenetic diversity of biological soil crusts in the Colorado Plateau studied by molecular fingerprinting and intensive cultivation. Microb. Ecol. 2006, 52, 345–357. [Google Scholar] [CrossRef]
- Lan, S.; Ouyang, H.; Wu, L.; Zhang, D.; Hu, C. Biological soil crust community types differ in photosynthetic pigment composition, fluorescence and carbon fixation in Shapotou region of China. Appl. Soil Ecol. 2017, 111, 9–16. [Google Scholar] [CrossRef]
- Zhou, Z.; Cheng, Z.; Liu, Z. Study on algae ecology in desert crust. Acta Ecol. Sin. 1995, 4, 385–391. [Google Scholar]
- Aditya, L.; Mahlia, T.M.I.; Nguyenn, L.N.; Vu, H.P.; Nghiem, L.D. Microalgae-bacteria consortium for wastewater treatment and biomass production. Sci. Total Environ. 2022, 838, 155871. [Google Scholar] [CrossRef]
- Ju, M.; Bu, C.; Wang, Q.; Bai, X.; Li, Y.; Guo, Q.; Wei, Y. Effects of algae and microbial additions on artificial restoration of biological crust on high steep slope. Bull. Soil Water Conserv. 2019, 39, 124–128+35. [Google Scholar]
- Wang, H.; Bu, C.; Wu, S.; Wang, C.; Mo, Q. Effects of photosynthetic bacteria and chlorella on the provenological expansion of Lichenia lithophyticus. Acta Bot. Sin. Northwest China 2023, 43, 1888–1896. [Google Scholar]
- Zhou, L.; Liu, W.; Duan, H.; Dong, H.; Li, J.; Zhang, S.; Zhang, J.; Ding, S.; Xu, T.; Guo, B. Improved effects of combined application of nitrogen-fixing bacteria Azotobacter beijerinckii and microalgae Chlorella pyrenoidosa on wheat growth and saline-alkali soil quality. Chemosphere 2023, 313, 137409. [Google Scholar] [CrossRef]
- Bao, S. Soil Agrochemical Analysis, 3rd ed.; China Agricultural Publishing House: Beijing, China, 2000. [Google Scholar]
- Liu, S.; Fu, B.; Chen, L.; Qiu, J.; Lv, Y. Comparison of two quantitative evaluation methods for soil quality change. Resour. Environ. Yangtze Basin 2003, 5, 422–426. [Google Scholar]
- Zhang, Q.; Song, Y.; You, W. The Relationship between Secondary Succession of Plant Communities in Zhejiang Tian Tong Area and Soil Fertility. Acta Ecol. Sin. 1999, 19, 32–36. [Google Scholar]
- Chen, J.; Yu, S.; Fu, J.; Xu, G.; Yu, B.; Lai, X.; Hu, S.; Zhang, K.; Liu, J. Soil quality characteristics and influencing factors of different utilization methods in red bed geomorphic regions of South China: A case study of Nanxiong Basin. Chin. J. Ecol. Environ. 2022, 31, 918–930. [Google Scholar]
- Zheng, F.; Zhang, F.; Wang, B. Quantitative evaluation of soil quality degradation under vegetation destruction and erosion in recent 100 years. Acta Ecol. Sin. 2010, 30, 6044–6051. [Google Scholar]
- Magdoff, F. Defining Soil Quality for a Sustainable Environment. Edited by J.W. Doran, D.C. Coleman, D.F. Bezdicek, and B.A. Stewart. 1994. Soil Sci. Soc. Amer. Spec. Pub. 35. American Society of Agronomy, 677 South Segoe Road, Madison, WI 53711-1086. $30.00, paper, xxiii + 244 pp. Am. J. Altern. Agric. 1995, 10, 46. [Google Scholar]
- Zhu, J.; Qin, F.; Li, L.; Yang, Z.; Fang, F.; Zhao, Q. Comprehensive evaluation on soil fertility of typical stand of plantation in loess hilly area. China Soil Fertil. 2022, 2, 9–16. [Google Scholar]
- Tang, D.; Qing, R.; Fu, H.; Lan, L. Study on the improvement of poor soil by soil microalgae. J. Sichuan Univ. 2003, 40, 352–355. [Google Scholar]
- Peng, X.; Wang, T.; Ma, J.; Tong, Y.; Wang, X.; Zhang, M.; Ye, Y.; Chen, S. Study on the improvement of soil acid-alkalinity by microbial agent. Tianjin Sci. Technol. 2021, 48, 42–58. [Google Scholar]
- Maltas, A.S.; Tavali, I.E.; Uz, I.; Kaplan, M. Monitoring the effects of pH and EC regulated drip fertigation on microbial dynamics of calcareous soil in tomato (Solanum lycopersicum L.) cultivation under greenhouse conditions in a Mediterranean climate. Sci. Hortic. 2022, 306, 111448. [Google Scholar] [CrossRef]
- Yinga, W.; En, W.; Lan, W.; Yan, Z. Effects of composite microbial fertilizer on biological properties and soil fertility of alkaline soil. China Soil Fertil. 2021, 1, 197–203. [Google Scholar]
- Deng, L.; Wang, T.; Luo, W.; He, L.; Liang, Z. Effects of a compound microbial agent and plants on soil properties, enzyme activities, and bacterial composition of Pisha sandstone. Environ. Sci. Pollut. Res. 2021, 28, 53353–53364. [Google Scholar] [CrossRef] [PubMed]
- Belnap, J.; Phillips, S.L.; Smith, S.D. Dynamics of cover, UV-protective pigments, and quantum yield in biological soil crust communities of an undisturbed Mojave Desert shrubland. Flora Morphol. Distrib. Funct. Ecol. Plants 2007, 202, 674–686. [Google Scholar] [CrossRef]
- Qiu, Z.; Gou, Y.; Zhang, Z.; Wang, Z.; Meng, P.; Zhou, J.; Jiang, Y. Research progress on application of algae resources in agricultural planting. J. Agric. Resour. Environ. 2023, 40, 840–851. [Google Scholar]
- Deng, H.; Gao, M.; Long, Y.; Lai, J.X.; Wang, Y.Y.; Wang, Z.F. Effects of biochar and straw returning on soil aggregate and organic carbon in purple soil on dry slope. J. Environ. Sci. 2018, 42, 5481–5490. [Google Scholar]
- Jiang, K.; Jia, Y.; Yang, Y.; Chen, J.; Yu, P. Effects of land use change on aggregate stability and organic carbon in karst area of Southwest China. Environ. Sci. 2024, 45, 2840–2847. [Google Scholar] [CrossRef]
- Shao, H.; Li, Z.; Liu, D.; Li, Y.; Lu, L.; Wang, X.; Zhang, A.; Wang, Y. Effects of organic fertilizer application on soil organic carbon components and aggregate stability. Environ. Sci. 2019, 40, 4691–4699. [Google Scholar]
- Luo, C.; Yang, J.; Chen, W.; Han, F. Effect of biochar on soil properties on the Loess Plateau: Results from field experiments. Geoderma 2020, 369, 114323. [Google Scholar] [CrossRef]
- An, Y.; Ji, Q.; Zhao, S.-x.; Wang, X.-d. Effects of biochar on soil aggregate distribution and water retention in orchards. Environ. Sci. 2016, 37, 293–300. [Google Scholar]
- Brodowski, S.; John, B.; Flessa, H.; Amelung, W. Aggregate-occluded black carbon in soil. Eur. J. Soil Sci. 2006, 57, 539–546. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, P.; Bao, J.-T.; Zhao, J.-C.; Song, G.; Yang, H.-T.; Huang, L.; He, M.-Z.; Li, X.-R. Comparison of cyanobacterial communities in temperate deserts: A cue for artificial inoculation of biological soil crusts. Sci. Total Environ. 2020, 745, 140970. [Google Scholar] [CrossRef]
- Adessi, A.; Cruz de Carvalho, R.; De Philippis, R.; Branquinho, C.; Marques da Silva, J. Microbial extracellular polymeric substances improve water retention in dryland biological soil crusts. Soil Biol. Biochem. 2018, 116, 67–69. [Google Scholar] [CrossRef]
- Zhuang, J.; Xian, J.; Wang, X.; Li, Z.; Cheng, C. Changes of soil microbial biomass carbon and nitrogen and its influencing factors in planted forests in low hilly areas of North China. Fujian J. Agric. Sci. 2023, 38, 1459–1465. [Google Scholar] [CrossRef]
- Zhou, Z.; Feng, H.; Ding, J.; Wang, Y.; Li, Z.; Qi, Y. Effects of compound microbial fertilizer on the stability and enzyme activity of tobacco rhizosphere soil aggregate. J. South. Agric. Sci. 2022, 53, 3088–3097. [Google Scholar]
- Yang, Y.J.; Liu, S.R.; Wang, H.; Chen, L.; Lu, L.H.; Cai, D.X. Reduction in throughfall reduces soil aggregate stability in two subtropical plantations. Eur. J. Soil Sci. 2019, 2, 70. [Google Scholar] [CrossRef]
- Wang, Y.; Niu, W.; Li, X.; Wang, J.; Guan, Y.; Dong, J. Effects of biochar on soil aggregate structure in solar greenhouse. Bull. Soil Water Conserv. 2019, 39, 190–195. [Google Scholar]
- Shirazi, M.A.; Boersma, L. A Unifying Quantitative Analysis of Soil Texture. Soil Sci. Soc. Am. J. 1984, 48, 142–147. [Google Scholar] [CrossRef]
- Qiao, D.; Wu, M.; Zhang, Q.; Han, Y.; Zhang, Y.; Li, P.; Li, H. Effects of straw returning to field and biochar application on aggregate stability and organic carbon accumulation of yellow brown soil. China Soil Fertil. 2018, 3, 92–99. [Google Scholar]
- van Bavel, C.H.M. Mean Weight-Diameter of Soil Aggregates as a Statistical Index of Aggregation. Soil Sci. Soc. Am. J. 1950, 14, 20–23. [Google Scholar] [CrossRef]
- Dong, T.; Deng, Z.; Yang, J.; Dai, Q.; Nie, Y. Characteristics of soil aggregate stability at different vegetation restoration stages in karst degraded farmland. Res. Soil Water Conserv. 2024, 31, 33–42. [Google Scholar]
- Wang, P.; Tian, T.; Zhang, Y.; Xu, J.; Li, Z.; Shi, J. Effects of compound amendments on acidified soil and aggregates in facilities. Environ. Sci. Technol. 2021, 44, 167–176. [Google Scholar]
- Zhu, Z.; Bai, Y.; Lv, M.; Tian, G.; Zhang, X.; Li, L.; Jiang, Y.; Ge, S. Soil Fertility, Microbial Biomass, and Microbial Functional Diversity Responses to Four Years Fertilization in an Apple Orchard in North China. Hortic. Plant J. 2020, 6, 223–230. [Google Scholar] [CrossRef]
- Tian, L.; Liu, J.; Zhao, B.; Mi, J.; Li, Y.; Wang, Y.; Fei, N. Effects of combined application of water-retaining agents and microbial fertilizer on soil microbial biomass carbon, nitrogen content and enzyme activity in dryland oat. J. Soil Water Conserv. 2020, 34, 361–368. [Google Scholar] [CrossRef]
- Chen, M.; Sheng, R.; Zhang, W.; Hou, H.; Wei, W.; Ge, T.; Wang, S. Effects of multiple cropping methods on soil microbial biomass carbon and nitrogen in red soil paddy fields. Res. Agric. Mod. 2023, 44, 692–700. [Google Scholar]
- Paradelo, R.; Barral, M.T. Effect of moisture and disaggregation on the microbial activity of soil. Soil Tillage Res. 2009, 104, 317–319. [Google Scholar] [CrossRef]
- Xu, G.; Lv, Y.; Sun, J.; Shao, H.; Wei, L. Recent Advances in Biochar Applications in Agricultural Soils: Benefits and Environmental Implications. CLEAN Soil Air Water 2012, 40, 1093–1098. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, C.; Franklin, O.; Dieckmann, U.; Richter, A. Microbial community dynamics alleviate stoichiometric constraints during litter decay. Ecol. Lett. 2014, 17, 680–690. [Google Scholar] [CrossRef] [PubMed]
- Fisk, M.; Santangelo, S.; Minick, K. Carbon mineralization is promoted by phosphorus and reduced by nitrogen addition in the organic horizon of northern hardwood forests. Soil Biol. Biochem. 2015, 81, 212–218. [Google Scholar] [CrossRef]
- Chen, S. Microalgal crust and its role in fertilizing karst rocky desertification area. China Sci. Technol. Inf. 2005, 18, 64–65. [Google Scholar]
- Sun, X.; Li, X.; Tang, S.; Lin, K.; Zhao, T.; Chen, X. A review on algal-bacterial symbiosis system for aquaculture tail water treatment. Sci. Total Environ. 2022, 847, 157620. [Google Scholar] [CrossRef]
- Kohler, J.; Caravaca, F.; Carrasco, L.; Roldán, A. Interactions between a plant growth-promoting rhizobacterium, an AM fungus and a phosphate-solubilising fungus in the rhizosphere of Lactuca sativa. Appl. Soil Ecol. 2007, 35, 480–487. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.-X.; Guo, X.; Qin, Y.; Garrido-Oter, R.; Schulze-Lefert, P.; Bai, Y. High-throughput cultivation and identification of bacteria from the plant root microbiota. Nat. Protoc. 2021, 16, 988–1012. [Google Scholar] [CrossRef]
- Lu, N. Application of Algal Cell Suspension Culture in High-Throughput Biological Screening for Antibacterial and Insecticidal Agents; Gansu Agricultural University: Lanzhou, China, 2004. [Google Scholar]
- Rana, A.; Joshi, M.; Prasanna, R.; Shivay, Y.S.; Nain, L. Biofortification of wheat through inoculation of plant growth promoting rhizobacteria and cyanobacteria. Eur. J. Soil Biol. 2012, 50, 118–126. [Google Scholar] [CrossRef]
- Sharma, S.-B.; Sayyed, R.-Z.; Trivedi, M.-H.; Gobi, T.-A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 587. [Google Scholar] [CrossRef]
- Etesami, H.; Emami, S.; Alikhani, H.-A. Potassium solubilizing bacteria (KSB): Mechanisms, promotion of plant growth, and future prospects—A review. J. Soil Sci. Plant Nutr. 2017, 17, 897–911. [Google Scholar] [CrossRef]
- de Bruijn, F. Biological Nitrogen Fixation. In Principles of Plant-Microbe Interactions; Lugtenberg, B., Ed.; Springer: Cham, Switzerland, 2015. [Google Scholar] [CrossRef]
- Ahmed, E.; Holmström, S.J.M. Siderophores in environmental research. Microb. Biotechnol. 2014, 7, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Patten, C.-L.; Glick, B.-R. Bacterial biosynthesis of indole-3-acetic acid. Can. J. Microbiol. 1996, 42, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Decho, A.-W.; Gutierrez, T. Microbial extracellular polymeric substances (EPS) in ocean systems. Front. Microbiol. 2017, 8, 922. [Google Scholar] [CrossRef] [PubMed]
- Bernard, R.G.; Penrose, D.-M.; Li, J.-P. A Model For the Lowering of Plant Ethylene Concentrations by Plant Growth-promoting Bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [CrossRef]
- Bell, C.-W.; Fricks, B.-E.; Rocca, J.-D.; Steinweg, J.-M.; McMahon, S.-K.; Wallenstein, M.-D. High-throughput fluorometric measurement of potential soil extracellular enzyme activities. J. Vis. Exp. 2013, 81, e50961. [Google Scholar] [CrossRef] [PubMed]
- Brookes, P.-C.; Landman, A.; Pruden, G.; Jenkinson, D.-S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.-D.; Brookes, P.-C.; Jenkinson, D.-S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Lu, R.-K. Soil Agricultural Chemistry Analysis Methods; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | CK | Z | A | AZ |
|---|---|---|---|---|
| pH | 8.11 ± 0.13a | 8.15 ± 0.13a | 8.18 ± 0.13a | 8.27 ± 0.17a |
| EC/(μs·cm−1) | 59.93 ± 5.64b | 55.3 ± 7.07b | 66.53 ± 3.46b | 87.9 ± 13.48a |
| SWC/% | 9.3 ± 1.1bc | 12.4 ± 0.7a | 9.1 ± 0.6c | 11.3 ± 0.9ab |
| SOC/(g·kg−1) | 7.09 ± 0.72c | 8.09 ± 0.33bc | 9.44 ± 0.55b | 15.23 ± 1.36a |
| TN/(g·kg−1) | 0.26 ± 0.01c | 0.41 ± 0.01b | 0.43 ± 0.03b | 0.47 ± 0.03a |
| TP/(g·kg−1) | 0.13 ± 0.01c | 0.34 ± 0.01a | 0.31 ± 0.01b | 0.34 ± 0.01a |
| C/N | 27.45 ± 3.68b | 19.96 ± 1.4c | 22.19 ± 1.84c | 32.09 ± 1.09a |
| C/P | 54.7 ± 9.33a | 24 ± 1.39b | 30.02 ± 1.11b | 45.28 ± 5.3a |
| N/P | 1.36 ± 0.07bc | 1.41 ± 0.12b | 1.99 ± 0.13a | 1.2 ± 0.03c |
| AN/(mg·kg−1) | 102.67 ± 10.69b | 149.33 ± 14.57a | 172.67 ± 10.69a | 184.33 ± 39.8a |
| AP/(mg·kg−1) | 15.77 ± 0.6d | 38.68 ± 0.85a | 28.28 ± 1.86c | 31.56 ± 1.75b |
| AK/(mg·kg−1) | 20.4 ± 1.9c | 21.27 ± 0.95c | 31 ± 0.36b | 38.03 ± 0.72a |
| EPS/(mg·kg−1) | 0.0485 ± 0.0004d | 0.8339 ± 0.0021b | 0.6837 ± 0.0017c | 0.9131 ± 0.0022a |
| Treatments | MWD (mm) | GMD (mm) | R0.25 (%) | PAD (%) | ELT (%) | K |
|---|---|---|---|---|---|---|
| CK | 0.61 ± 0.01c | 0.44 ± 0.02b | 0.48 ± 0.01b | 0.43 ± 0.01a | 0.52 ± 0.01a | 0.21 ± 0.01a |
| Z | 0.74 ± 0.01b | 0.49 ± 0.04ab | 0.5 ± 0.01b | 0.42 ± 0.02a | 0.5 ± 0.01a | 0.2 ± 0.01ab |
| A | 0.74 ± 0.01b | 0.5 ± 0.02ab | 0.53 ± 0.06b | 0.39 ± 0.07ab | 0.47 ± 0.06a | 0.2 ± 0.01ab |
| AZ | 0.99 ± 0.03a | 0.64 ± 0.12a | 0.65 ± 0.01a | 0.29 ± 0.01b | 0.35 ± 0.01b | 0.16 ± 0.02b |
| Treatments | Total Length (cm) | Plant Height (cm) | Root Length (cm) | Total Weight (g) | Above-Ground Biomass (g) | Underground Biomass (g) | LNC (g/kg) | LPC (g/kg) |
|---|---|---|---|---|---|---|---|---|
| CK | 8.5 ± 0.9j | 4.5 ± 0.2h | 4 ± 0.9g | 1.66 ± 0.25i | 0.35 ± 0.03c | 1.31 ± 0.28i | 20.3 ± 0.35i | 0.23 ± 0.01k |
| Z | 16.6 ± 1i | 8.4 ± 0.4g | 8.2 ± 0.7ef | 4.64 ± 1.33def | 0.71 ± 0.74bc | 3.93 ± 0.6efg | 25.67 ± 0.44g | 1.18 ± 0.17cdefg |
| A | 20.8 ± 0.8def | 10.4 ± 0.3cde | 10.4 ± 0.5abc | 5.66 ± 0.6cde | 0.53 ± 0.02bc | 5.13 ± 0.58de | 38.7 ± 0.66c | 1.61 ± 0.04ab |
| AZ | 24.2 ± 2.5b | 12.6 ± 1.2b | 11.6 ± 1.4a | 13.15 ± 1.05b | 1.15 ± 0.09ab | 12 ± 1.06c | 43.25 ± 0.74a | 1.74 ± 0.03a |
| Index | Principal Component 1 | Principal Component 2 | Principal Component 3 | Weight |
|---|---|---|---|---|
| Loading Coefficient | Loading Coefficient | Loading Coefficient | ||
| pH | 0.973 | 0.229 | 0.034 | 0.0534 |
| EC | 0.82 | 0.573 | 0.01 | 0.0554 |
| SOC | 0.922 | 0.369 | 0.121 | 0.0562 |
| AN | 0.952 | −0.189 | −0.241 | 0.0544 |
| AP | 0.62 | −0.744 | 0.25 | 0.0543 |
| TN | 0.954 | −0.285 | −0.092 | 0.0549 |
| TP | 0.83 | −0.557 | −0.023 | 0.0556 |
| SWC | 0.328 | −0.425 | 0.844 | 0.0404 |
| C/N | 0.355 | 0.9 | 0.253 | 0.0461 |
| C/P | −0.261 | 0.948 | 0.181 | 0.0419 |
| N/P | −0.668 | 0.743 | 0.029 | 0.0532 |
| AK | 0.905 | 0.347 | −0.245 | 0.0567 |
| EPS | 0.877 | −0.472 | 0.088 | 0.0565 |
| MBC | 0.986 | 0.164 | −0.024 | 0.0520 |
| MBN | 0.97 | 0.131 | −0.206 | 0.0531 |
| MBC/MBN | −0.908 | 0.035 | 0.417 | 0.0506 |
| MWD | 0.966 | 0.163 | 0.202 | 0.0537 |
| GMD | 0.947 | 0.26 | 0.189 | 0.0554 |
| R0.25 | 0.92 | 0.376 | 0.111 | 0.0562 |
| Characteristic Root | 13.107 | 4.563 | 1.329 | / |
| Variance Contribution Rate % | 68.99 | 24.02 | 7.00 | / |
| Cumulative Variance Contribution Rate % | 68.987 | 93.004 | 100 | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, L.; Xie, X.; Chen, M.; Qiao, F.; Liu, X.; Zhao, Y.; Peng, X.; Liu, F. Co-Inoculation Between Bacteria and Algae from Biological Soil Crusts and Their Effects on the Growth of Poa annua and Sandy Soils Quality. Microorganisms 2025, 13, 1778. https://doi.org/10.3390/microorganisms13081778
Peng L, Xie X, Chen M, Qiao F, Liu X, Zhao Y, Peng X, Liu F. Co-Inoculation Between Bacteria and Algae from Biological Soil Crusts and Their Effects on the Growth of Poa annua and Sandy Soils Quality. Microorganisms. 2025; 13(8):1778. https://doi.org/10.3390/microorganisms13081778
Chicago/Turabian StylePeng, Lin, Xuqiang Xie, Man Chen, Fengjie Qiao, Xingyu Liu, Yutong Zhao, Xiawei Peng, and Fangchun Liu. 2025. "Co-Inoculation Between Bacteria and Algae from Biological Soil Crusts and Their Effects on the Growth of Poa annua and Sandy Soils Quality" Microorganisms 13, no. 8: 1778. https://doi.org/10.3390/microorganisms13081778
APA StylePeng, L., Xie, X., Chen, M., Qiao, F., Liu, X., Zhao, Y., Peng, X., & Liu, F. (2025). Co-Inoculation Between Bacteria and Algae from Biological Soil Crusts and Their Effects on the Growth of Poa annua and Sandy Soils Quality. Microorganisms, 13(8), 1778. https://doi.org/10.3390/microorganisms13081778