
Interferon-Induced Transmembrane Protein 3 (IFITM3) Restricts PRRSV Replication via Post-Entry Mechanisms

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. IFITM3 Transfection in MARC-145 Cells
2.2. siRNA-Induced Knockdown of IFITM3 in MARC-145 Cells
2.3. Western Blot Analysis
2.4. Interferon Stimulation of IFITM3 Gene in MARC-145 Cells
2.5. Immunofluorescence Assay and Flow Cytometry
2.6. Colocalization Study Using Confocal Microscopy
2.7. Cytotoxicity Assay
2.8. Real-Time Reverse Transcription PCR (RT-PCR)
2.9. TCID50 Titer
2.10. Statistical Analysis
3. Results
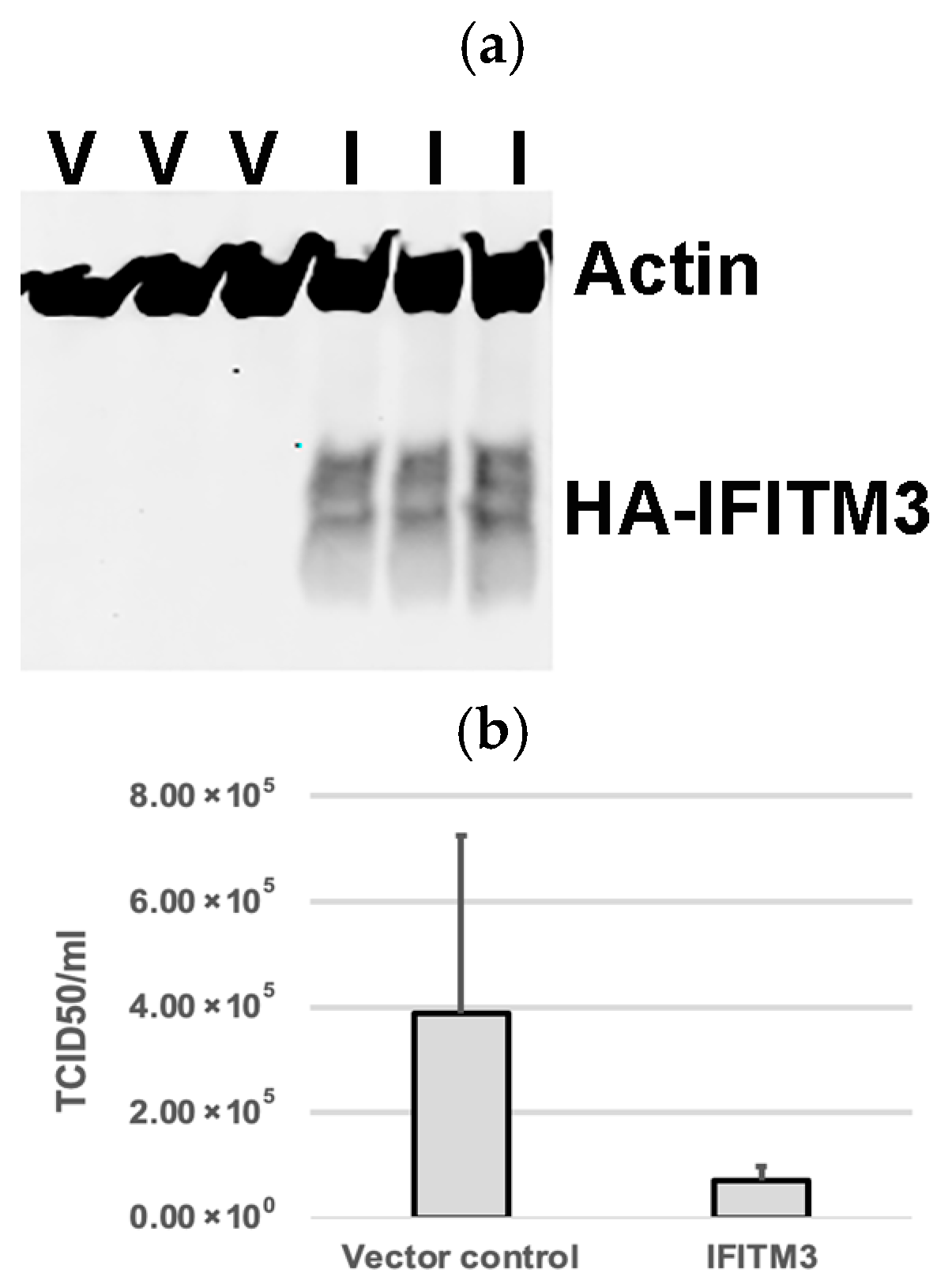
3.1. Over-Expression of Exogenous IFITM3 Reduces PRRSV Replication
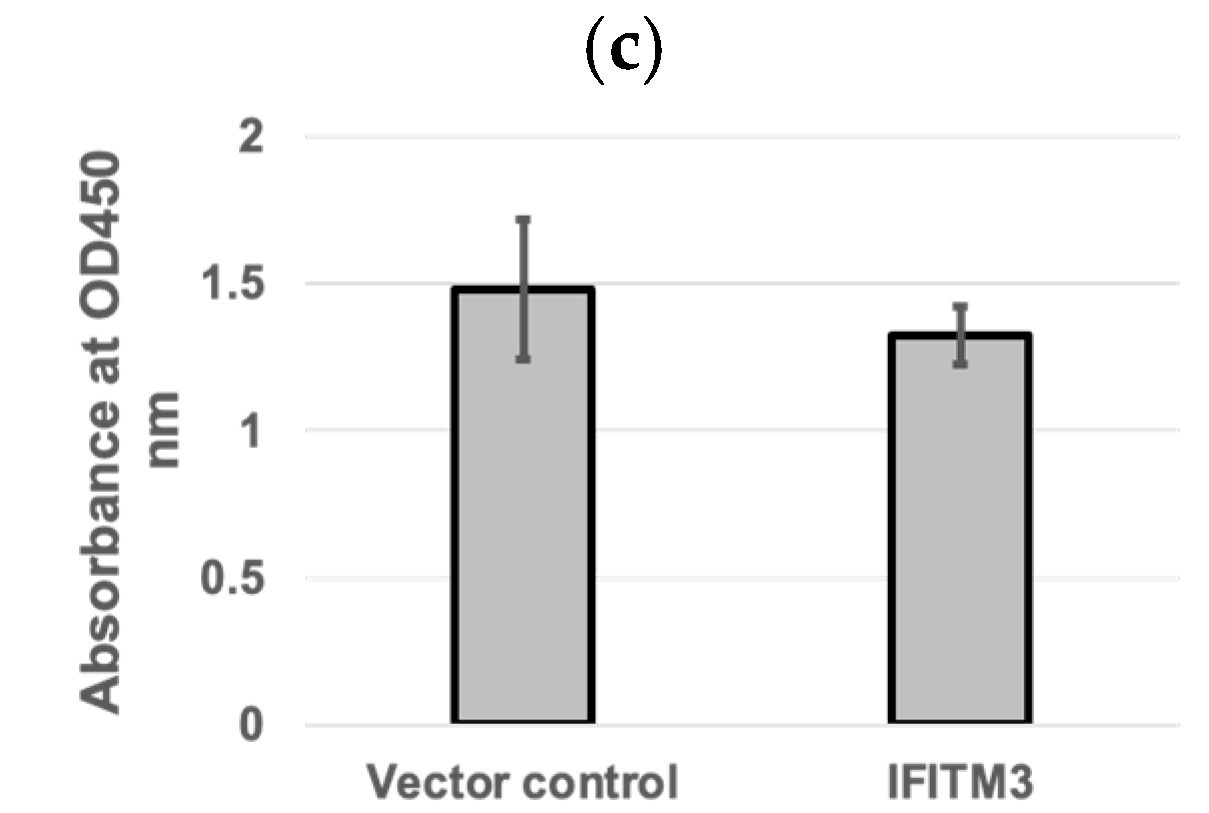
3.2. Knockdown of IFITM3 by siRNA Enhances PRRSV Replication
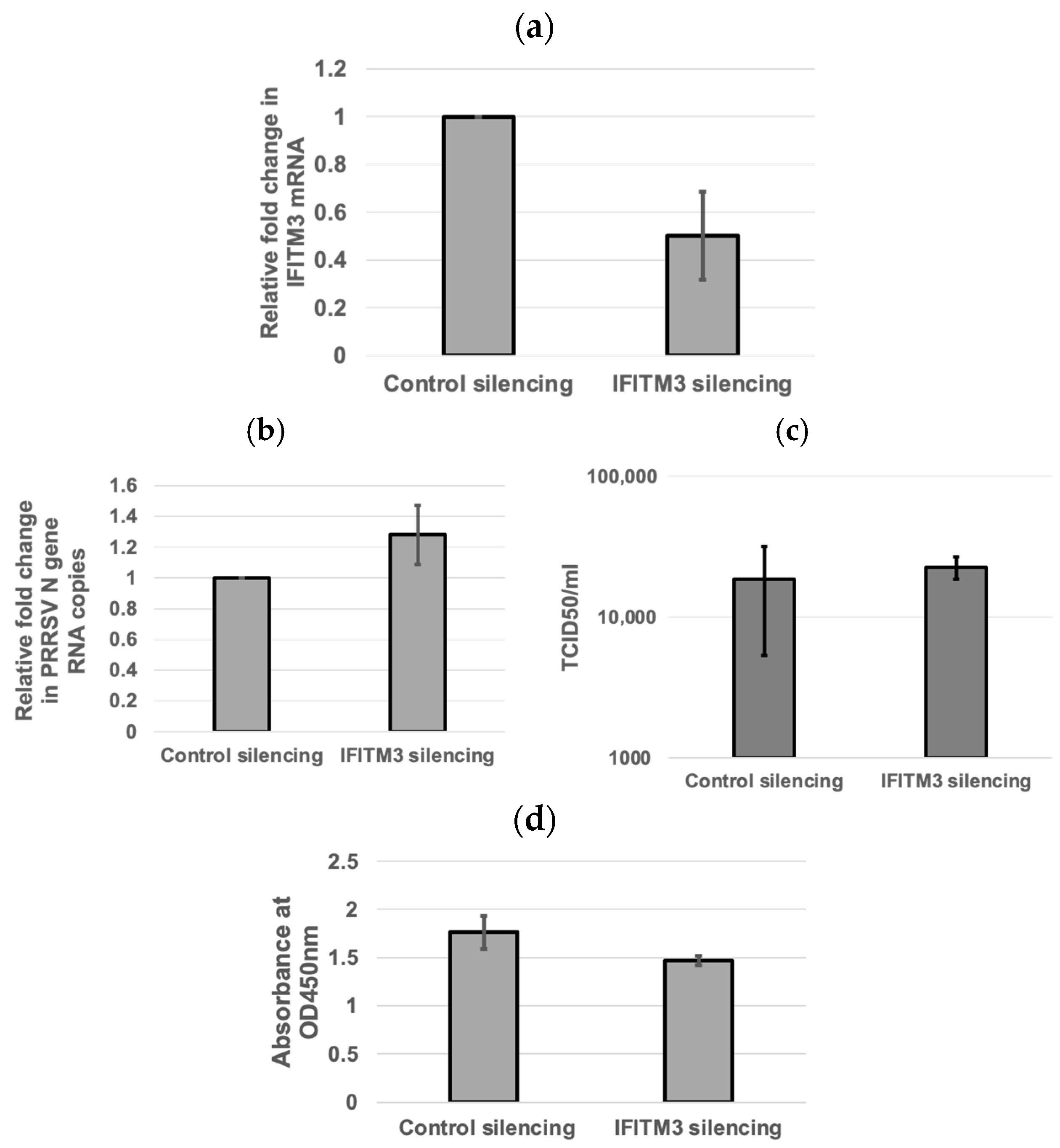
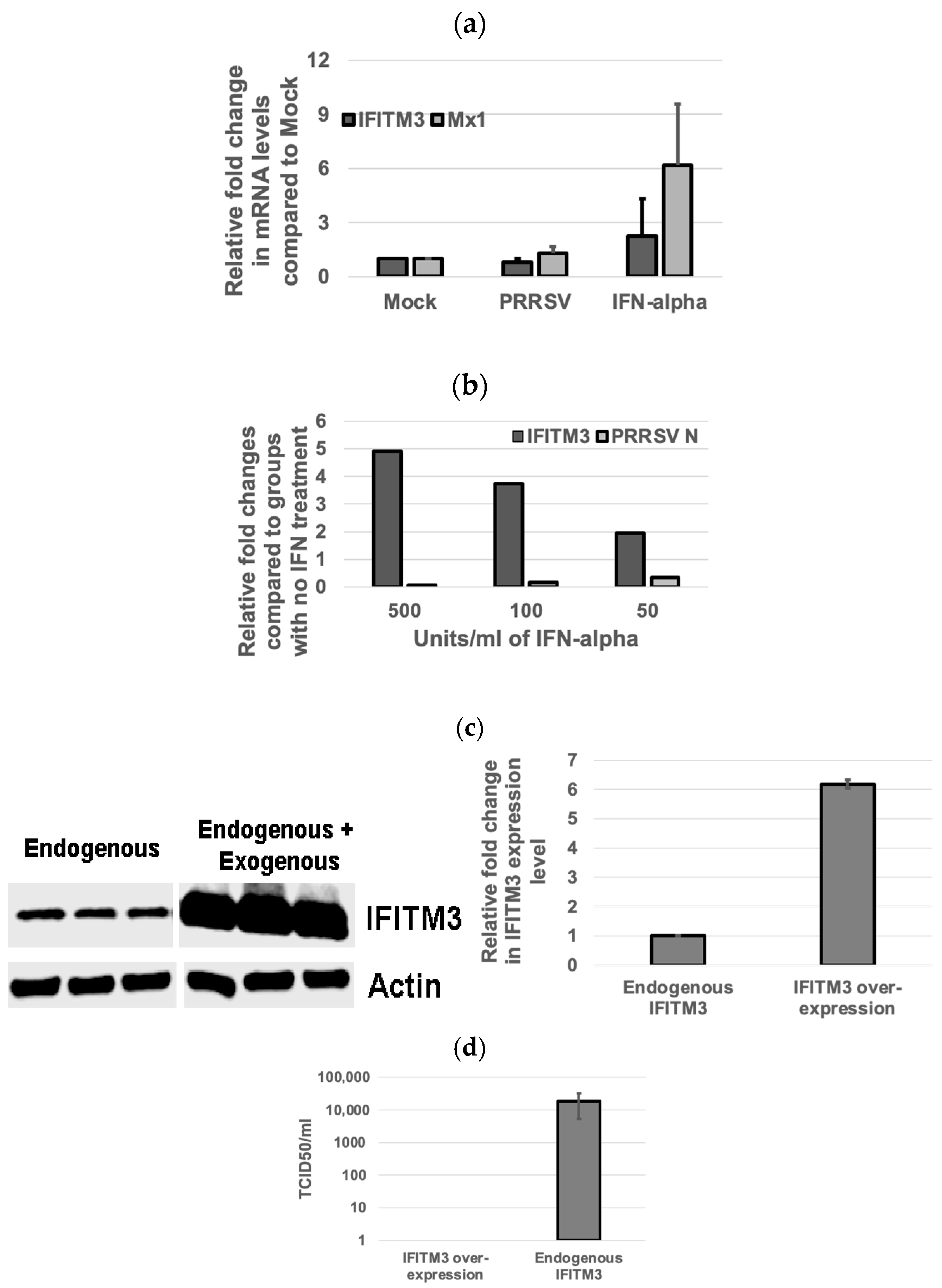
3.3. Inverse Correlation Between IFITM3 Expression Level and Virus Replication Efficiency
3.4. Amphotericin B Partially Restores PRRSV Replication in IFITM3-over-Expressing Cells
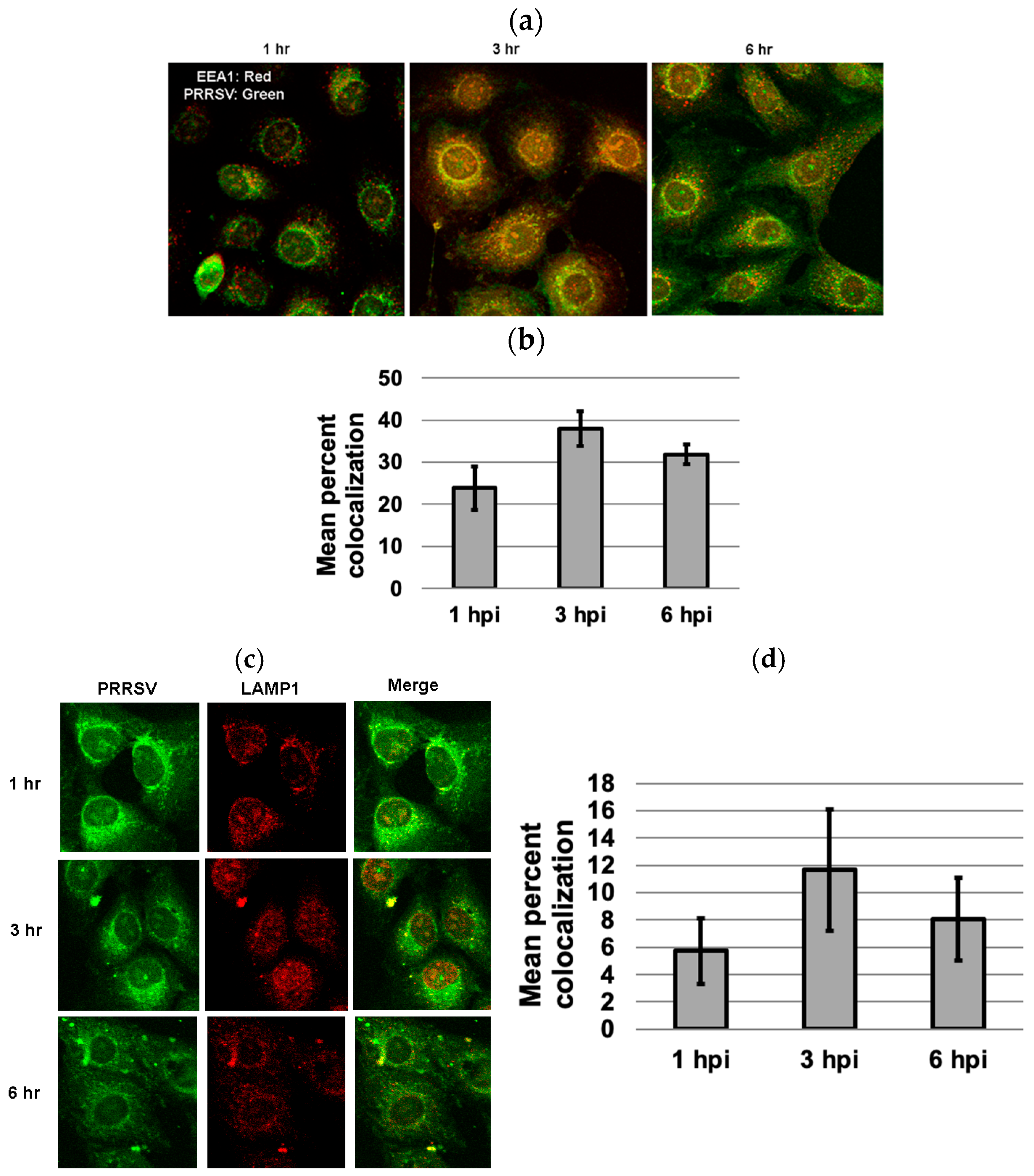
3.5. Colocalization of PRRSV with Early and Late Endosome/Lysosome Markers at 3 Hpi
3.6. Over-Expression of IFITM3 Does Not Significantly Impact Virus Entry
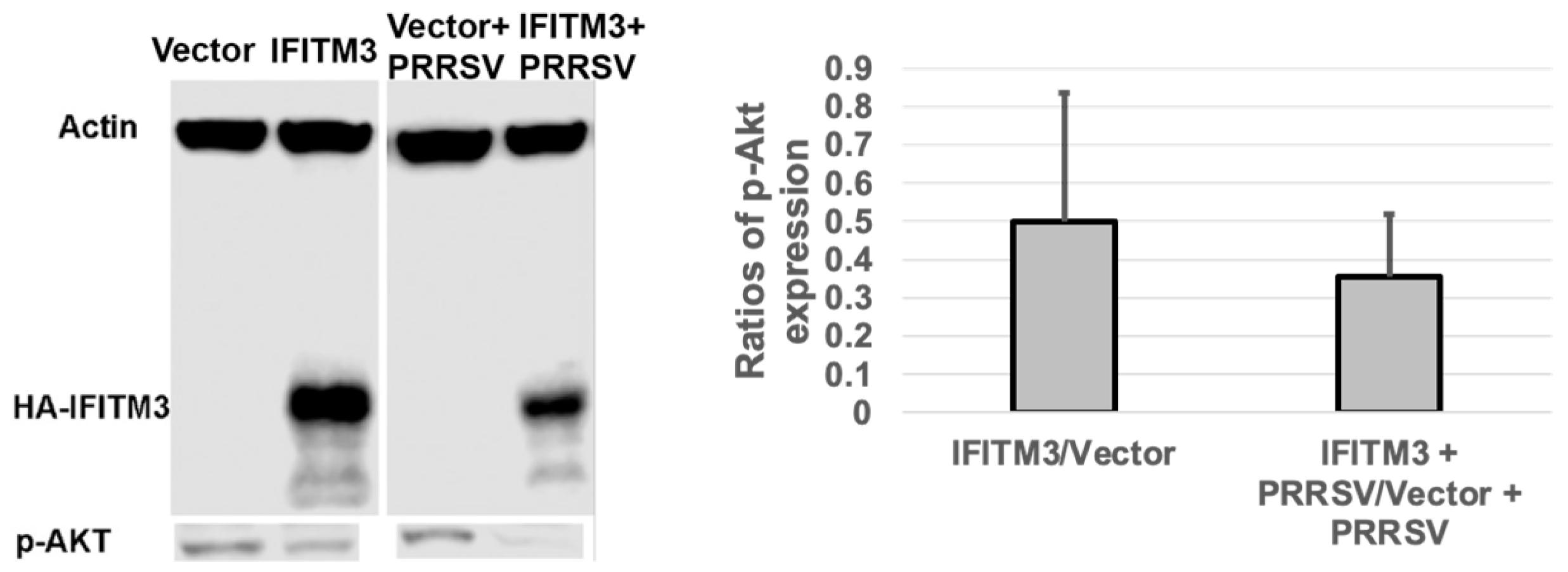
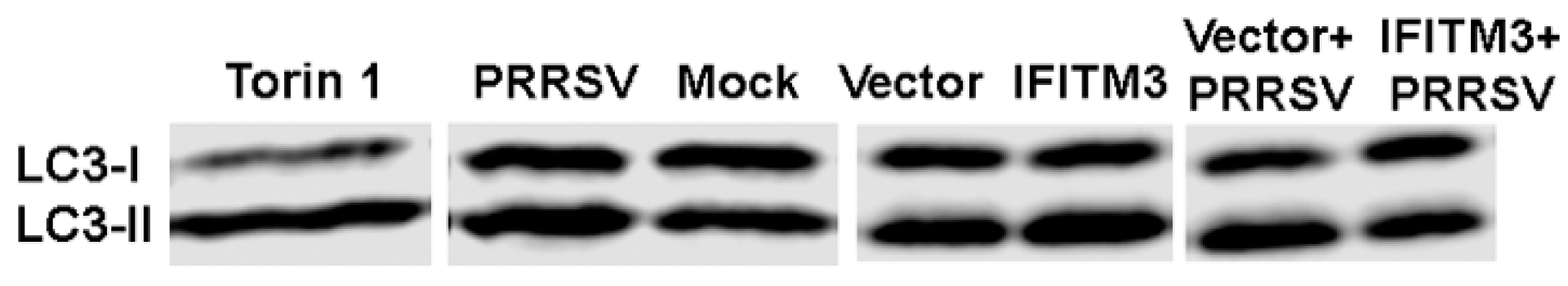
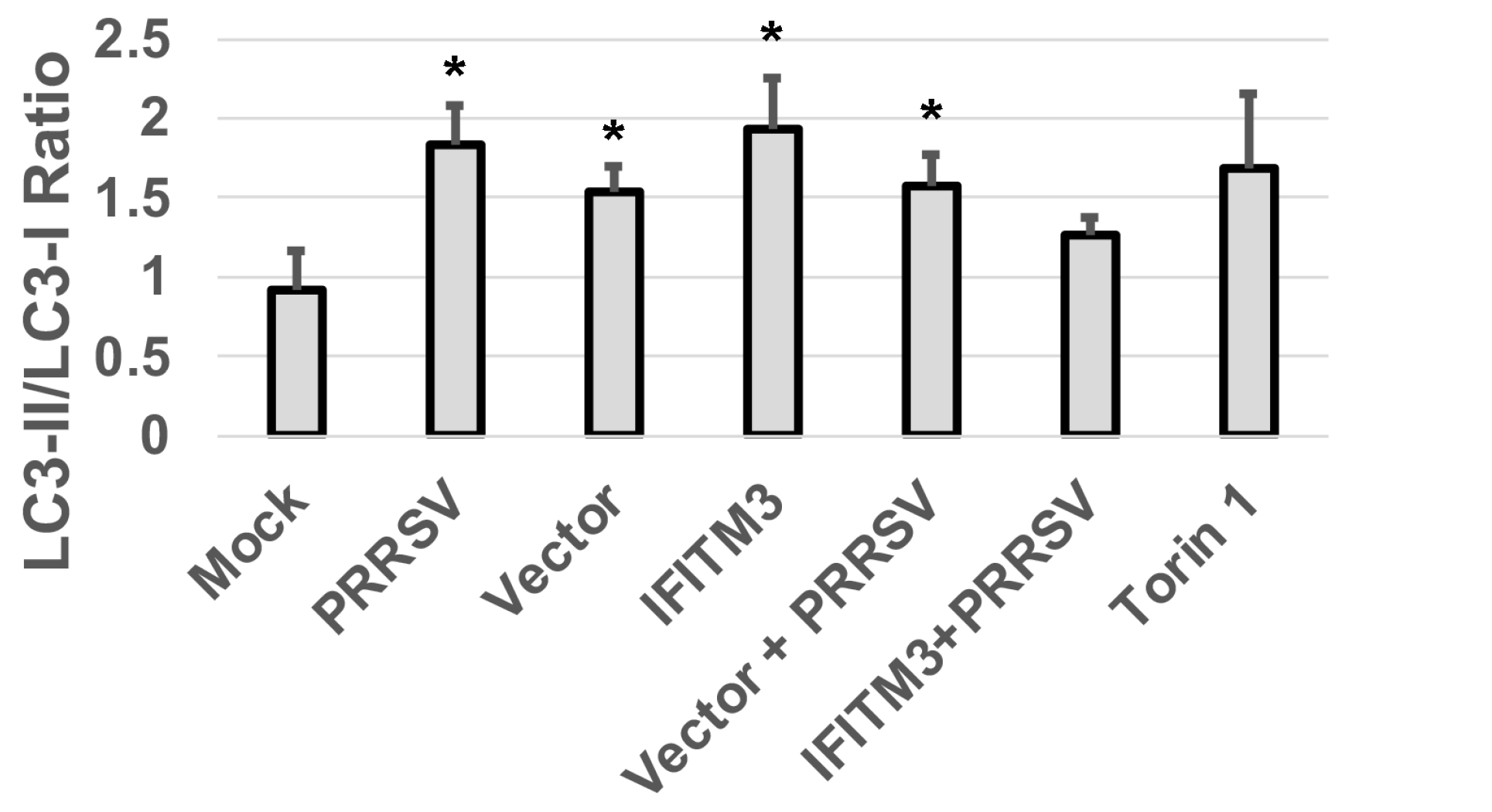
3.7. Over-Expression of IFITM3 Reduces the Level of P-Akt and Increases the Ratios of LC3-II/LC3-I
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IFITM3 | Interferon-induced transmembrane protein 3 |
| PRRSV | Porcine reproductive and respiratory syndrome virus |
| ISGs | Interferon-stimulated genes |
| ZMPSTE24 | Zinc metallopeptidase STE24 |
| IAV | Influenza A virus |
| MEFs | Mouse embryonic fibroblasts |
| vRNPs | Viral ribonucleoproteins |
| SARS-CoV | Severe acute respiratory syndrome coronavirus |
| LASVpp | Lassa virus pseudovirus |
| PI3K | Phosphatidylinositol 3-kinase |
| GBP1 | Guanylate binding protein 1 |
| TNTs | Tunneling nanotubes |
References
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-Stimulated Genes: A Complex Web of Host Defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef] [PubMed]
- Feeley, E.M.; Sims, J.S.; John, S.P.; Chin, C.R.; Pertel, T.; Chen, L.-M.; Gaiha, G.D.; Ryan, B.J.; Donis, R.O.; Elledge, S.J.; et al. IFITM3 Inhibits Influenza A Virus Infection by Preventing Cytosolic Entry. PLoS Pathog. 2011, 7, e1002337. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.L.; Manly, S.P.; McMahon, M.; Kerr, I.M.; Stark, G.R. Transcriptional and Posttranscriptional Regulation of Interferon-Induced Gene Expression in Human Cells. Cell 1984, 38, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.C.; Zhong, G.; Huang, I.-C.; Farzan, M. IFITM-Family Proteins: The Cell’s First Line of Antiviral Defense. Annu. Rev. Virol. 2014, 1, 261–283. [Google Scholar] [CrossRef] [PubMed]
- Brass, A.L.; Huang, I.-C.; Benita, Y.; John, S.P.; Krishnan, M.N.; Feeley, E.M.; Ryan, B.J.; Weyer, J.L.; van der Weyden, L.; Fikrig, E.; et al. The IFITM Proteins Mediate Cellular Resistance to Influenza A H1N1 Virus, West Nile Virus, and Dengue Virus. Cell 2009, 139, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Perreira, J.M.; Chin, C.R.; Feeley, E.M.; Brass, A.L. IFITMs Restrict the Replication of Multiple Pathogenic Viruses. J. Mol. Biol. 2013, 425, 4937–4955. [Google Scholar] [CrossRef] [PubMed]
- Haller, O.; Kochs, G. Human MxA Protein: An Interferon-Induced Dynamin-like GTPase with Broad Antiviral Activity. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 2011, 31, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Okumura, F.; Okumura, A.J.; Uematsu, K.; Hatakeyama, S.; Zhang, D.-E.; Kamura, T. Activation of Double-Stranded RNA-Activated Protein Kinase (PKR) by Interferon-Stimulated Gene 15 (ISG15) Modification down-Regulates Protein Translation. J. Biol. Chem. 2013, 288, 2839–2847. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J.W.; Rice, C.M. Interferon-Stimulated Genes and Their Antiviral Effector Functions. Curr. Opin. Virol. 2011, 1, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.C.; Huang, I.-C.; Kam, C.; Farzan, M. Ifitm3 Limits the Severity of Acute Influenza in Mice. PLoS Pathog. 2012, 8, e1002909. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; Weston, S.; Kellam, P.; Marsh, M. IFITM Proteins-Cellular Inhibitors of Viral Entry. Curr. Opin. Virol. 2014, 4, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Gorman, M.J.; Poddar, S.; Farzan, M.; Diamond, M.S. The Interferon-Stimulated Gene Ifitm3 Restricts West Nile Virus Infection and Pathogenesis. J. Virol. 2016, 90, 8212–8225. [Google Scholar] [CrossRef] [PubMed]
- Weidner, J.M.; Jiang, D.; Pan, X.-B.; Chang, J.; Block, T.M.; Guo, J.-T. Interferon-Induced Cell Membrane Proteins, IFITM3 and Tetherin, Inhibit Vesicular Stomatitis Virus Infection via Distinct Mechanisms. J. Virol. 2010, 84, 12646–12657. [Google Scholar] [CrossRef] [PubMed]
- Savidis, G.; Perreira, J.M.; Portmann, J.M.; Meraner, P.; Guo, Z.; Green, S.; Brass, A.L. The IFITMs Inhibit Zika Virus Replication. Cell Rep. 2016, 15, 2323–2330. [Google Scholar] [CrossRef] [PubMed]
- Done, S.H.; Paton, D.J.; White, M.E. Porcine Reproductive and Respiratory Syndrome (PRRS): A Review, with Emphasis on Pathological, Virological and Diagnostic Aspects. Br. Vet. J. 1996, 152, 153–174. [Google Scholar] [CrossRef] [PubMed]
- Nauwynck, H.J.; Duan, X.; Favoreel, H.W.; Van Oostveldt, P.; Pensaert, M.B. Entry of Porcine Reproductive and Respiratory Syndrome Virus into Porcine Alveolar Macrophages via Receptor-Mediated Endocytosis. J. Gen. Virol. 1999, 80 Pt 2, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yoo, D. PRRS Virus Receptors and Their Role for Pathogenesis. Vet. Microbiol. 2015, 177, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Calvert, J.G.; Slade, D.E.; Shields, S.L.; Jolie, R.; Mannan, R.M.; Ankenbauer, R.G.; Welch, S.-K.W. CD163 Expression Confers Susceptibility to Porcine Reproductive and Respiratory Syndrome Viruses. J. Virol. 2007, 81, 7371–7379. [Google Scholar] [CrossRef] [PubMed]
- Welch, S.-K.W.; Calvert, J.G. A Brief Review of CD163 and Its Role in PRRSV Infection. Virus Res. 2010, 154, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Kreutz, L.C.; Ackermann, M.R. Porcine Reproductive and Respiratory Syndrome Virus Enters Cells through a Low pH-Dependent Endocytic Pathway. Virus Res. 1996, 42, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Van Gorp, H.; Van Breedam, W.; Delputte, P.L.; Nauwynck, H.J. The Porcine Reproductive and Respiratory Syndrome Virus Requires Trafficking through CD163-Positive Early Endosomes, but Not Late Endosomes, for Productive Infection. Arch. Virol. 2009, 154, 1939–1943. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Duan, H.; Zhao, H.; Liao, H.; Du, Y.; Li, L.; Jiang, D.; Wan, B.; Wu, Y.; Ji, P.; et al. Interferon-Induced Transmembrane Protein 3 Is a Virus-Associated Protein Which Suppresses Porcine Reproductive and Respiratory Syndrome Virus Replication by Blocking Viral Membrane Fusion. J. Virol. 2020, 94, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Waheed, A.A.; Ablan, S.D.; Soheilian, F.; Nagashima, K.; Ono, A.; Schaffner, C.P.; Freed, E.O. Inhibition of Human Immunodeficiency Virus Type 1 Assembly and Release by the Cholesterol-Binding Compound Amphotericin B Methyl Ester: Evidence for Vpu Dependence. J. Virol. 2008, 82, 9776–9781. [Google Scholar] [CrossRef] [PubMed]
- Katwal, P.; Aftab, S.; Nelson, E.; Hildreth, M.; Li, S.; Wang, X. Role of Zinc Metalloprotease (ZMPSTE24) in Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) Replication in Vitro. Arch. Virol. 2022, 167, 2281–2286. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.; Wang, L.; Li, S.; Dorf, M.E. ZMPSTE24 Defends against Influenza and Other Pathogenic Viruses. J. Exp. Med. 2017, 214, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.A.; Christopher-Hennings, J.; Drew, T.; Wensvoort, G.; Collins, J.E.; Benfield, D.A. Differentiation of U.S. and European Isolates of Porcine Reproductive and Respiratory Syndrome Virus by Monoclonal Antibodies. J. Clin. Microbiol. 1993, 31, 3184–3189. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Franz, S.; Pott, F.; Zillinger, T.; Schüler, C.; Dapa, S.; Fischer, C.; Passos, V.; Stenzel, S.; Chen, F.; Döhner, K.; et al. Human IFITM3 Restricts Chikungunya Virus and Mayaro Virus Infection and Is Susceptible to Virus-Mediated Counteraction. Life Sci. Alliance 2021, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Denz, P.J.; Speaks, S.; Kenney, A.D.; Eddy, A.C.; Papa, J.L.; Roettger, J.; Scace, S.C.; Rubrum, A.; Hemann, E.A.; Forero, A.; et al. Innate Immune Control of Influenza Virus Interspecies Adaptation via IFITM3. Nat. Commun. 2024, 15, 9375. [Google Scholar] [CrossRef] [PubMed]
- Desai, T.M.; Marin, M.; Chin, C.R.; Savidis, G.; Brass, A.L.; Melikyan, G.B. IFITM3 Restricts Influenza A Virus Entry by Blocking the Formation of Fusion Pores Following Virus-Endosome Hemifusion. PLoS Pathog. 2014, 10, e1004048. [Google Scholar] [CrossRef] [PubMed]
- McMichael, T.M.; Zhang, Y.; Kenney, A.D.; Zhang, L.; Zani, A.; Lu, M.; Chemudupati, M.; Li, J.; Yount, J.S. IFITM3 Restricts Human Metapneumovirus Infection. J. Infect. Dis. 2018, 218, 1582–1591. [Google Scholar] [CrossRef] [PubMed]
- Amini-Bavil-Olyaee, S.; Choi, Y.J.; Lee, J.H.; Shi, M.; Huang, I.-C.; Farzan, M.; Jung, J.U. The Antiviral Effector IFITM3 Disrupts Intracellular Cholesterol Homeostasis to Block Viral Entry. Cell Host Microbe 2013, 13, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-Y.; Chin, C.R.; Everitt, A.R.; Clare, S.; Perreira, J.M.; Savidis, G.; Aker, A.M.; John, S.P.; Sarlah, D.; Carreira, E.M.; et al. Amphotericin B Increases Influenza A Virus Infection by Preventing IFITM3-Mediated Restriction. Cell Rep. 2013, 5, 895–908. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zheng, S.; Chen, D.; Zheng, M.; Li, X.; Li, G.; Lin, H.; Chang, J.; Zeng, H.; Guo, J.-T. LY6E Restricts Entry of Human Coronaviruses, Including Currently Pandemic SARS-CoV-2. J. Virol. 2020, 94, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Suddala, K.C.; Lee, C.C.; Meraner, P.; Marin, M.; Markosyan, R.M.; Desai, T.M.; Cohen, F.S.; Brass, A.L.; Melikyan, G.B. Interferon-Induced Transmembrane Protein 3 Blocks Fusion of Sensitive but Not Resistant Viruses by Partitioning into Virus-Carrying Endosomes. PLoS Pathog. 2019, 15, e1007532. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Katz, B.B.; Tomich, J.M.; Gallagher, T.; Fang, Y. Porcine Reproductive and Respiratory Syndrome Virus Utilizes Nanotubes for Intercellular Spread. J. Virol. 2016, 90, 5163–5175. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, H.; Abel, A.M.; Young, A.J.; Xie, L.; Xie, Z. Role of Phosphatidylinositol 3-Kinase (PI3K) and Akt1 Kinase in Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) Replication. Arch. Virol. 2014, 159, 2091–2096. [Google Scholar] [CrossRef] [PubMed]
- Gu, T.; Yu, D.; Xu, L.; Yao, Y.-L.; Yao, Y.-G. Tupaia GBP1 Interacts with STING to Initiate Autophagy and Restrict Herpes Simplex Virus Type 1 Infection. J. Immunol. 2021, 207, 2673–2680. [Google Scholar] [CrossRef] [PubMed]
- Yount, J.S.; Karssemeijer, R.A.; Hang, H.C. S-Palmitoylation and Ubiquitination Differentially Regulate Interferon-Induced Transmembrane Protein 3 (IFITM3)-Mediated Resistance to Influenza Virus. J. Biol. Chem. 2012, 287, 19631–19641. [Google Scholar] [CrossRef] [PubMed]
- Aftab, S.; Nelson, E.; Hildreth, M.; Wang, X. Silencing RNA-Mediated Knockdown of IFITM3 Enhances Senecavirus A Replication. Pathogens 2024, 13, 290. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, M.; Hirata, N.; Suizu, F. The Links between AKT and Two Intracellular Proteolytic Cascades: Ubiquitination and Autophagy. Biochim. Biophys. Acta 2014, 1846, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Fang, L.; Wang, D.; Wang, S.; Li, P.; Li, M.; Luo, R.; Chen, H.; Xiao, S. Induction of Autophagy Enhances Porcine Reproductive and Respiratory Syndrome Virus Replication. Virus Res. 2012, 163, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Men, Y.; Wang, D.; Xu, D.; Liu, S.; Xiao, S.; Fang, L. Porcine Reproductive and Respiratory Syndrome Virus Infection Induces Endoplasmic Reticulum Stress, Facilitates Virus Replication, and Contributes to Autophagy and Apoptosis. Sci. Rep. 2020, 10, 13131. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Qin, Y.; Zhou, L.; Kou, Q.; Guo, X.; Ge, X.; Yang, H.; Hu, H. Autophagy Sustains the Replication of Porcine Reproductive and Respiratory Virus in Host Cells. Virology 2012, 429, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Bowman, J.W.; Jung, J.U. Autophagy during Viral Infection—A Double-Edged Sword. Nat. Rev. Microbiol. 2018, 16, 341–354. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | (Reverse Primer (5′-3′) |
|---|---|---|
| beta-actin | TTGCTGACAGGATGCAGAAGGAGA | ACTCCTGCTTGCTGATCCACATCT |
| IFITM3 | GGTCTTCGCTGGACACCAT | TGTCCCTAGACTTCACGGAGTA |
| PRRSV N | GTCAATCCAGACCGCCTTTA | GATCAGGCGCACAGTATGAT |
| Mx1 | TTTTCAAGAAGGAGGCCAGCAA | TCAGGAACTTCCGCTTGTCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katwal, P.; Aftab, S.; Nelson, E.; Hildreth, M.; Li, S.; Wang, X. Interferon-Induced Transmembrane Protein 3 (IFITM3) Restricts PRRSV Replication via Post-Entry Mechanisms. Microorganisms 2025, 13, 1737. https://doi.org/10.3390/microorganisms13081737
Katwal P, Aftab S, Nelson E, Hildreth M, Li S, Wang X. Interferon-Induced Transmembrane Protein 3 (IFITM3) Restricts PRRSV Replication via Post-Entry Mechanisms. Microorganisms. 2025; 13(8):1737. https://doi.org/10.3390/microorganisms13081737
Chicago/Turabian StyleKatwal, Pratik, Shamiq Aftab, Eric Nelson, Michael Hildreth, Shitao Li, and Xiuqing Wang. 2025. "Interferon-Induced Transmembrane Protein 3 (IFITM3) Restricts PRRSV Replication via Post-Entry Mechanisms" Microorganisms 13, no. 8: 1737. https://doi.org/10.3390/microorganisms13081737
APA StyleKatwal, P., Aftab, S., Nelson, E., Hildreth, M., Li, S., & Wang, X. (2025). Interferon-Induced Transmembrane Protein 3 (IFITM3) Restricts PRRSV Replication via Post-Entry Mechanisms. Microorganisms, 13(8), 1737. https://doi.org/10.3390/microorganisms13081737