
Soil Microbiome Drives Depth-Specific Priming Effects in Picea schrenkiana Forests Following Labile Carbon Input

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Location and Soil Sample Collection
2.2. Incubation Experiment Design and CO2 Analysis
2.3. Determination of Soil Physicochemical Properties and Analysis of PLFAs
2.4. Index Calculation
2.5. Statistical Analysis
3. Results
3.1. Soil Organic Carbon Mineralization
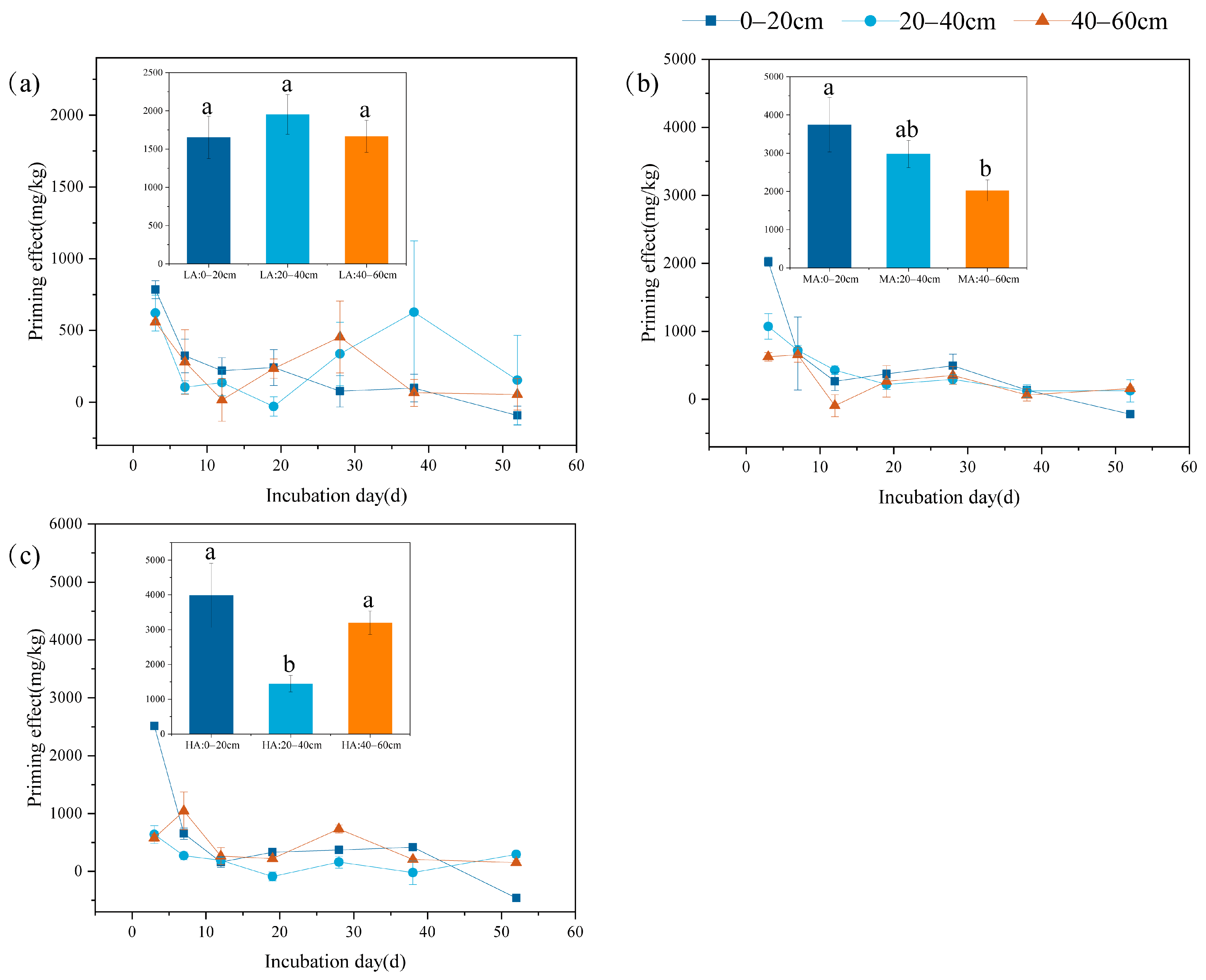
3.2. The Priming Effect of Soil Organic Carbon
3.3. Soil Microbial Communities
3.4. Driving Factors Affecting the PE of Labile Carbon Inputs on Soil Organic Carbon
4. Discussion
4.1. Effects of Soil Depth and Glucose Input Amount on Organic Carbon Mineralization and Priming Effect
4.2. Microbial and Abiotic Drivers of Soil Organic Carbon Priming Effects Under Labile Carbon Inputs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kendall, R.A.; Harper, K.A.; Burton, D.; Hamdan, K. The role of temperate treed swamps as a carbon sink in southwestern Nova Scotia. Can. J. Forest Res. 2021, 51, 78–88. [Google Scholar] [CrossRef]
- Sayer, E.J.; Heard, M.S.; Grant, H.K.; Marthews, T.R.; Tanner, E.V. Soil carbon release enhanced by increased tropical forest litterfall. Nat. Clim. Change 2011, 1, 304–307. [Google Scholar] [CrossRef]
- De Graaff, M.A.; Classen, A.T.; Castro, H.F.; Schadt, C.W. Labile soil carbon inputs mediate the soil microbial community composition and plant residue decomposition rates. New Phytol. 2010, 188, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Day, F.P.; Schroeder, R.E.; Stover, D.B.; Brown, A.L.; Butnor, J.R.; Dilustro, J.; Hungate, B.A.; Dijkstra, P.; Duval, B.D.; Seiler, T.J.; et al. The effects of 11 yr of CO2 enrichment on roots in a Florida scrub-oak ecosystem. New Phytol. 2013, 200, 778–787. [Google Scholar] [CrossRef] [PubMed]
- Kuzyakov, Y.; Friedel, J.K.; Stahr, K. Review of mechanisms and quantification of priming effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Guenet, B.; Camino-Serrano, M.; Ciais, P.; Tifafi, M.; Maignan, F.; Soong, J.L.; Janssens, I.A. Impact of priming on global soil carbon stocks. Glob. Change Biol. 2018, 24, 1873–1883. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.J.A.; Sun, J.R.; Mau, R.L.; Finley, B.K.; Compson, Z.G.; Van Gestel, N.; Brown, J.R.; Sehwarz, E.; Dijkstra, P.; Hungate, B.A. Labile carbon input determines the direction and magnitude of the priming effect. Appl. Soil Ecol. 2017, 109, 7–13. [Google Scholar] [CrossRef]
- Qiao, N.A.; Schaefer, D.; Blagodatskaya, E.; Zou, X.; Xu, X.; Kuzyakov, Y. Labile carbon retention compensates for CO2 released by priming in forest soils. Glob. Change Biol. 2014, 20, 1943–1954. [Google Scholar] [CrossRef] [PubMed]
- Guenet, B.; Juarez, S.; Bardoux, G.; Abbadie, L.; Chenu, C. Evidence that stable C is as vulnerable to priming elfect as is more labile C in soil. Soil Biol. Biochem. 2012, 52, 43–48. [Google Scholar] [CrossRef]
- Xiao, C.W.; Guenet, B.; Zhou, Y.; Su, J.W.; Janssens, I.A. Priming of soil organie matter deomposition seales linearly with microbial biomass responseto litter input in steppe vegetation. Oikos 2015, 124, 649–657. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Review: Factors affecting rhizosphere priming effects. J. Plant Nutr. Soil Sci. 2002, 165, 382–396. [Google Scholar] [CrossRef]
- Di Lonardo, D.P.; De Boer, W.; Zweers, H.; van der Wal, A. Effect of the amount of organic trigger compounds, nitrogen and soil microbial biomass on the magnitude of priming of soil organic matter. PLoS ONE 2019, 14, e0216730. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.J.A.; Finley, B.K.; Mau, R.L.; Schwartz, E.; Dijkstra, P.; Bowker, M.A.; Hungate, B.A. The soil priming effect: Consistent across ecosystems, elusive mechanisms. Soil Biol. Biochem. 2020, 140, 107617. [Google Scholar] [CrossRef]
- Feng, J.; Tang, M.; Zhu, B. Soil priming effect and its responses to nutrient addition along a tropical forest elevation gradient. Glob. Change Biol. 2021, 27, 2793–2806. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.J.; Mo, F.; Zhou, Z.H.; Bastida, F.; Delgado-Baquerizo, M.; Wang, J.Y.; Zhang, X.Y.; Luo, Y.Q.; Griffis, T.J.; Han, X.H.; et al. The global biogeography of soil priming effect intensity. Global Ecol. Biogeogr. 2022, 31, 1679–1687. [Google Scholar] [CrossRef]
- Gaudel, G.; Poudel, M.; Mosongo, P.S.; Li, X.; Oljira, M.A.; Zhang, Y.M.; Bizimana, F.; Liu, B.B.; Wang, Y.Y.; Dong, W.X.; et al. Meta-analysis of the priming effect on native soil organic carbon in response to glucose amendment across soil depths. Plant Soil 2022, 479, 107–124. [Google Scholar] [CrossRef]
- Luo, Z.; Wang, E.; Sun, O.J. A meta-analysis of the temporal dynamics of priming soil carbon decomposition by fresh carbon inputs across ecosystems. Soil Biol. Biochem. 2016, 101, 96–103. [Google Scholar] [CrossRef]
- Wang, C.; Kuzyakov, Y. Soil organic matter priming: The pH effects. Glob. Change Biol. 2024, 30, e17349. [Google Scholar] [CrossRef] [PubMed]
- Mehnaz, K.R.; Corneo, P.E.; Keitel, C.; Dijkstra, F.A. Carbon and phosphorus addition effects on microbial carbon use efficiency, soil organic matter priming, gross nitrogen mineralization and nitrous oxide emission from soil. Soil Biol. Biochem. 2019, 134, 175–186. [Google Scholar] [CrossRef]
- Lin, Q.; De Vrieze, J.; Fang, X.Y.; Li, L.J.; Li, X.Z. Labile carbon feedstocks trigger a priming effect in anaerobic digestion: An insight into microbial mechanisms. Bioresour. Technol. 2022, 344, 126243. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wu, C.; Wei, L.; Wang, S.; Deng, Y.W.; Ling, W.L.; Xiang, W.; Kuzyakov, Y.; Zhu, Z.K.; Ge, T.D. Microbial mechanisms of organic matter mineralization induced by straw in biochar-amended paddy soil. Biochar 2024, 6, 18. [Google Scholar] [CrossRef]
- Shahbaz, M.; Kuzyakov, Y.; Sanaullah, M.; Heitkamp, F.; Zelenev, V.; Kumar, A.; Blagodatskaya, E. Microbial decomposition of soil organic matter is mediated by quality and quantity of crop residues: Mechanisms and thresholds. Biol. Fertil. Soils 2017, 53, 287–301. [Google Scholar] [CrossRef]
- Yu, Z.; Chen, L.; Pan, S.; Li, Y.; Kuzyakov, Y.; Xu, J.; Luo, Y. Feedstock determines biochar-induced soil priming effects by stimulating the activity of specific microorganisms. Eur. J. Soil Sci. 2018, 69, 521–534. [Google Scholar] [CrossRef]
- Zhu, Z.K.; Fang, Y.Y.; Liang, Y.L.; Li, Y.H.; Liu, S.L.; Li, Y.F.; Li, B.Z.; Gao, W.; Yuan, H.Z.; Kuzyakov, Y.; et al. Stoichiometric regulation of priming effects and soil carbon balance by microbial life strategies. Soil Biol. Biochem. 2022, 169, 108669. [Google Scholar] [CrossRef]
- Fontaine, S.; Mariotti, A.; Abbadie, L. The priming effect of organic matter: A question of microbial competition? Soil Biol. Biochem. 2003, 35, 837–843. [Google Scholar] [CrossRef]
- Jackson, O.; Quilliam, R.S.; Stott, A.; Grant, H.; Subke, J.A.V. Rhizosphere carbon supply accelerates soil organic matter decomposition in the presenceof fresh organic substrates. Plant Soil 2019, 440, 473–490. [Google Scholar] [CrossRef]
- Fontaine, S.; Barot, S.; Barré, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of organiccarbon in deep soil layers controlled by fresh carbonsupply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.K.; Xie, J.; Wang, C.; Guo, J.F.; Wang, M.H.; Liu, X.F.; Chen, Y.H.; Chen, G.S.; Yang, Y.S. Forest conversion stimulated deep soil C losses and decreased C recalcitrance through priming effect in subtropical China. Biol. Fertil. Soils 2015, 51, 857–867. [Google Scholar] [CrossRef]
- Meyer, N.; Welp, G.; Rodionov, A.; Borchard, N.; Martius, C.; Amelung, W. Nitrogen and phosphorus supply controls soil organic carbon mineralization in tropical topsoil and subsoil. Soil Biol. Biochem. 2018, 119, 152–161. [Google Scholar] [CrossRef]
- Liang, Z.; Olesen, J.E.; Jensen, J.L.; Elsgaard, L. Nutrient availability affects carbon turnover and microbial physiology differently in topsoil and subsoil under a temperate grassland. Geoderma 2019, 336, 22–30. [Google Scholar] [CrossRef]
- Hicks, P.C.E.; Ryals, R.; Zhu, B.; Min, K.; Cooper, A.; Goldsmith, S.; Pett-Ridge, J.; Torn, M.; Berhe, A.A. The deep soil organic carbon response to global change. Annu. Rev. Ecol. Evol. Syst. 2023, 54, 375–401. [Google Scholar] [CrossRef]
- Chu, H.; Sun, H.; Tripathi, B.M.; Adams, J.M.; Huang, R.; Zhang, Y.; Shi, Y. Bacterial community dissimilarity between the surface and subsurface soils equals horizontal differences over several kilometers in the western Tibetan Plateau. Environ. Microbiol. 2016, 18, 1523–1533. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.T.; Chen, X.; Wang, Q.Q.; Zhang, Y.L.; Yao, S.H.; Zhang, B. The priming effect dynamics are driven by microbial activation and growth and constrained by the relative availability of input C and soil N. Biol. Fertil. Soils 2022, 58, 745–760. [Google Scholar] [CrossRef]
- Bai, X.J.; Zhai, G.G.; Yan, Z.F.; An, S.; Liu, J.; Huo, L.G.; Dippold, M.A.; Kuzyakov, Y. Effects of microbial groups on soil organic carbon accrual and mineralization during high-and low-quality litter decomposition. Catena 2024, 241, 108051. [Google Scholar] [CrossRef]
- Fu, K.X.; Chen, L.X.; Yu, X.X.; Jia, G.D. How has carbon storage changed in the Yili-Tianshan region over the past three decades and into the future? What has driven it to change? Sci. Total Environ. 2024, 945, 174005. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Bao, A.M.; Xu, W.Q.; Zheng, G.X.; Nzabarinda, V.; Yu, T.; Huang, X.R.; Long, G.; Naibi, S. Dynamics of forest net primary productivity based on tree ring reconstruction in the Tianshan Mountains. Ecol. Indic. 2023, 146, 109713. [Google Scholar] [CrossRef]
- Xu, F.J.; Xu, Z.L.; Xu, C.C.; Yu, T.T. Automatic Extraction of the Spatial Distribution of Picea schrenkiana in the Tianshan Mountains Based on Google Earth Engine and the Jeffries-Matusita Distance. Forests 2023, 14, 1373. [Google Scholar] [CrossRef]
- Li, X.C.; Gong, L.; Ding, Z.L.; Abudesiyiti, K.; Wang, X.F.; Ma, X.Y.; Li, H. Negative effects of canopy gaps on soil physicochemical properties and microbial community structure and functions in Picea Schrenkiana pure forests. Plant Soil 2024, 509, 83–103. [Google Scholar] [CrossRef]
- Wu, L.; Xu, H.; Xiao, Q.; Huang, Y.P.; Suleman, M.M.; Zhu, P.; Kuzyakov, Y.; Xu, X.L.; Xu, M.G.; Zhang, W.J. Soil carbon balance by priming differs with single versus repeated addition of glucose and soil fertility level. Soil Biol. Biochem. 2020, 148, 107913. [Google Scholar] [CrossRef]
- Liu, G.; Feng, Y.; Yin, Z.; Yan, G.; Wang, Q.; Xing, Y. Exogenous organic C inputs profit soil C sequestration under different long-term N addition levels in a boreal forest. J. Soil Sci. Plant Nutr. 2023, 23, 1740–1750. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Joergensen, R.G. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the KEC value. Soil Biol. Biochem. 1996, 28, 25–31. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purifcation. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Bossio, D.A.; Scow, K.M.; Gunapala, N.; Graham, K.J. Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profles. Microb. Ecol. 1998, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Thornton, B.; Zhang, Z.; Mayes, R.W.; Hogberg, M.N.; Midwood, A.J. Can gas chromatography combustion isotope ratio mass spectrometry be used to quantify organic compound abundance? Rapid Commun. Mass Spectrom. 2011, 25, 2433–2438. [Google Scholar] [CrossRef] [PubMed]
- Buyer, J.S.; Teasdale, J.R.; Roberts, D.P.; Zasada, I.A.; Maul, J.E. Factors affecting soil microbial community structure in tomato cropping systems. Soil Biol. Biochem. 2010, 42, 831–841. [Google Scholar] [CrossRef]
- Joergensen, R.G. Phospholipid fatty acids in soil-drawbacks and future prospects. Biol. Fertil. Soils 2022, 58, 1–6. [Google Scholar] [CrossRef]
- Williams, M.A.; Myrold, D.D.; Bottomley, P.J. Carbon fow from C-13-labeled straw and root residues into the phospholipid fatty acids of a soil microbial community under feld conditions. Soil Biol. Biochem. 2006, 38, 759–768. [Google Scholar] [CrossRef]
- Liang, Z.; Elsgaard, L.; Nicolaisen, M.H.; Lyhne-Kjærbye, A.; Olesen, J.E. Carbon mineralization and microbial activity in agricultural topsoil and subsoil as regulated by root nitrogen and recalcitrant carbon concentrations. Plant Soil 2018, 433, 65–82. [Google Scholar] [CrossRef]
- Liao, C.; Tian, Q.X.; Wang, D.Y.; Qu, L.Y.; Wu, Y.; Liu, F. Effects of labile carbon addition on organic carbon mineralization and microbial growth strategies in subtropical forest soils. J. Appl. Ecol. 2016, 27, 2848–2854. [Google Scholar]
- Oertel, C.; Matschullat, J.; Zurba, K.; Zimmermann, F.; Erasmi, S. Greenhouse gas emissions from soils-A review. Geochemistry 2016, 76, 327–352. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Li, P.; Wang, X.; Xiao, L.; Zhou, S. Surface soil microbiome changes in Grain for Green Project accelerates organic carbon mineralization on the Loess Plateau in China. Earth Surf. Process. Landf. 2024, 49, 1621–1631. [Google Scholar] [CrossRef]
- Endress, M.G.; Dehghani, F.; Blagodatsky, S.; Reitz, T.; Schlüter, S.; Blagodatskaya, E. Spatial substrate heterogeneity limits microbial growth as revealed by the joint experimental quantification and modeling of carbon and heat fluxes. Soil Biol. Biochem. 2024, 197, 109509. [Google Scholar] [CrossRef]
- Kleber, M.; Bourg, I.C.; Coward, E.K.; Hansel, C.M.; Myneni, S.C.B.; Nunan, N. Dynamic interactions at the mineral-organic matter interface. Nat. Rev. Earth Environ. 2021, 2, 402–421. [Google Scholar] [CrossRef]
- Zhang, X.B.; Pei, G.T.; Zhang, T.Y.; Fan, X.L.; Liu, Z.P.; Bai, E. Erosion effects on soil microbial carbon use efficiency in the mollisol cropland in northeast China. Soil Ecol. Lett. 2023, 5, 230176. [Google Scholar] [CrossRef]
- Hu, Y.L.; Feng, J.G.; Zhang, S.; Luo, Z.K.; Zhu, B. Depth-dependent regulations of soil priming effects along a 2000 km grassland transect. Soil Biol. Biochem. 2025, 202, 109696. [Google Scholar] [CrossRef]
- Blagodatskaya, E.V.; Blagodatsky, S.A.; Anderson, T.H.; Kuzyakov, Y. Priming effects in Chernozem induced by glucose and N in relation to microbial growth strategies. Appl. Soil Ecol. 2007, 37, 95–105. [Google Scholar] [CrossRef]
- Cui, J.; Zhu, Z.K.; Xu, X.L.; Liu, S.L.; Jones, D.L.; Kuzyakov, Y.; Shibistova, O.; Wu, J.S.; Ge, T.D. Carbon and nitrogen recycling from microbial necromass to cope with C: N stoichiometric imbalance by priming. Soil Biol. Biochem. 2020, 142, 107720. [Google Scholar] [CrossRef]
- Chang, Q.; Liu, Z.P.; Zhang, T.Y.; Liu, S.S.; Liu, B.; Fan, X.L.; Meng, D.; Zhang, K.; Bai, E. Soil priming effect in the organic and mineral layers regulated by nitrogen mining mechanism in a temperate forest. Land Degrad. Dev. 2024, 35, 4961–4971. [Google Scholar] [CrossRef]
- Zhang, Q.F.; Cheng, L.; Feng, J.G.; Mei, K.C.; Zeng, Q.X.; Zhu, B.; Chen, Y. Nitrogen addition stimulates priming effect in a subtropical forest soil. Soil Biol. Biochem. 2021, 160, 108339. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhang, S.; Tang, C.; Xiao, M.L.; Zhou, J.S.; Luo, Y.; Ge, T.D.; Yu, B.; White, J.C.; Li, Y.F. Divergent effects of straw and biochar on soil carbon priming are depth-dependent in subtropical Moso bamboo forests. Biol. Fertil. Soils 2025. [Google Scholar] [CrossRef]
- Jagadamma, S.; Mayes, M.A.; Steinweg, J.M.; Schaeffer, S.M. Substrate quality alters the microbial mineralization of added substrate and soil organic carbon. Biogeosciences 2014, 11, 4665–4678. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, B.R.; Shen, J.K.; Xu, F.J.; Li, N.; Jia, P.H.; Jia, Y.J.; An, S.S.; Amoah, I.D.; Huang, Y.M. Shifts in C-degradation genes and microbial metabolic activity with vegetation types affected the surface soil organic carbon pool. Soil Biol. Biochem. 2024, 28, 10937. [Google Scholar] [CrossRef]
- Butler, J.L.; Williams, M.A.; Bottomley, P.J.; Myrold, D.D. Microbial community dynamics associated with rhizosphere carbon flow. Appl. Environ. Microb. 2003, 69, 6793–6800. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.Q.; Gong, L.; Luo, Y.; Tang, J.; Ding, Z.; Li, X.Z. Effects of litter and root manipulations on soil bacterial and fungal community structure and function in a schrenk’s spruce (Picea schrenkiana) forest. Front. Plant Sci. 2022, 13, 849483. [Google Scholar] [CrossRef] [PubMed]
- Waldrop, M.P.; Balser, T.C.; Firestone, M.K. Linking microbial community composition to function in a tropical soil. Soil Biol. Biochem. 2000, 32, 1837–1846. [Google Scholar] [CrossRef]
- Karhu, K.; Alaei, S.; Li, J.; Merilä, P.; Ostonen, I.; Bengtson, P. Microbial carbon use efficiency and priming of soil organic matter mineralization by glucose additions in boreal forest soils with different C: N ratios. Soil Biol. Biochem. 2022, 167, 108615. [Google Scholar] [CrossRef]
- Perveen, N.; Barot, S.; Maire, V.; Cotrufo, M.F.; Shahzad, T.; Blagodatskaya, E.; Stewart, C.E.; Ding, W.X.; Siddiq, M.R.; Dimassi, B.; et al. Universality of priming effect: An analysis using thirty-five soils with contrasted properties sampled from five continents. Soil Biol. Biochem. 2019, 134, 162–171. [Google Scholar] [CrossRef]
- Feng, J.; Zhu, B. Global patterns and associated drivers of priming effect in response to nutrient addition. Soil Biol. Biochem. 2021, 153, 108118. [Google Scholar] [CrossRef]
- Zhou, J.; Guillaume, T.; Wen, Y.; Blagodatskaya, E.; Shahbaz, M.; Zeng, Z.; Peixoto, L.; Zang, H.; Kuzyakov, Y. Frequent carbon input primes decomposition of decadal soil organic matter. Soil Biol. Biochem. 2022, 175, 108850. [Google Scholar] [CrossRef]
- Nazir, M.J.; Hussain, M.M.; Albasher, G.; Iqbal, B.; Khan, K.A.; Rahim, R.; Li, G.; Du, D. Glucose input profit soil organic carbon mineralization and nitrogen dynamics in relation to nitrogen amended soils. J. Environ. Manag. 2024, 351, 119715. [Google Scholar] [CrossRef] [PubMed]
- Sawada, K.; Inagaki, Y.; Toyota, K. Priming effects induced by C and N additions in relation to microbial biomass turnover in Japanese forest soils. Appl. Soil Ecol. 2021, 162, 103884. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, K.; Gong, L.; Ma, X.; Li, X.; Sun, X. Soil Microbiome Drives Depth-Specific Priming Effects in Picea schrenkiana Forests Following Labile Carbon Input. Microorganisms 2025, 13, 1729. https://doi.org/10.3390/microorganisms13081729
Yin K, Gong L, Ma X, Li X, Sun X. Soil Microbiome Drives Depth-Specific Priming Effects in Picea schrenkiana Forests Following Labile Carbon Input. Microorganisms. 2025; 13(8):1729. https://doi.org/10.3390/microorganisms13081729
Chicago/Turabian StyleYin, Kejie, Lu Gong, Xinyu Ma, Xiaochen Li, and Xiaonan Sun. 2025. "Soil Microbiome Drives Depth-Specific Priming Effects in Picea schrenkiana Forests Following Labile Carbon Input" Microorganisms 13, no. 8: 1729. https://doi.org/10.3390/microorganisms13081729
APA StyleYin, K., Gong, L., Ma, X., Li, X., & Sun, X. (2025). Soil Microbiome Drives Depth-Specific Priming Effects in Picea schrenkiana Forests Following Labile Carbon Input. Microorganisms, 13(8), 1729. https://doi.org/10.3390/microorganisms13081729