
Grapevine Responses to the Entomopathogenic Fungi Beauveria bassiana and Isaria fumosorosea and the Effects of Salicylic Acid on Their Virulence Against the European Grapevine Moth, Lobesia botrana

 , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Material (Plants, Fungal Isolates, Insects)
2.2. Chlorophyll a Fluorescence and Physiological Parameter Measurements
2.3. Spore Germination Assays
2.4. Larval Mortality Bioassay
2.5. Statistical Analysis
3. Results
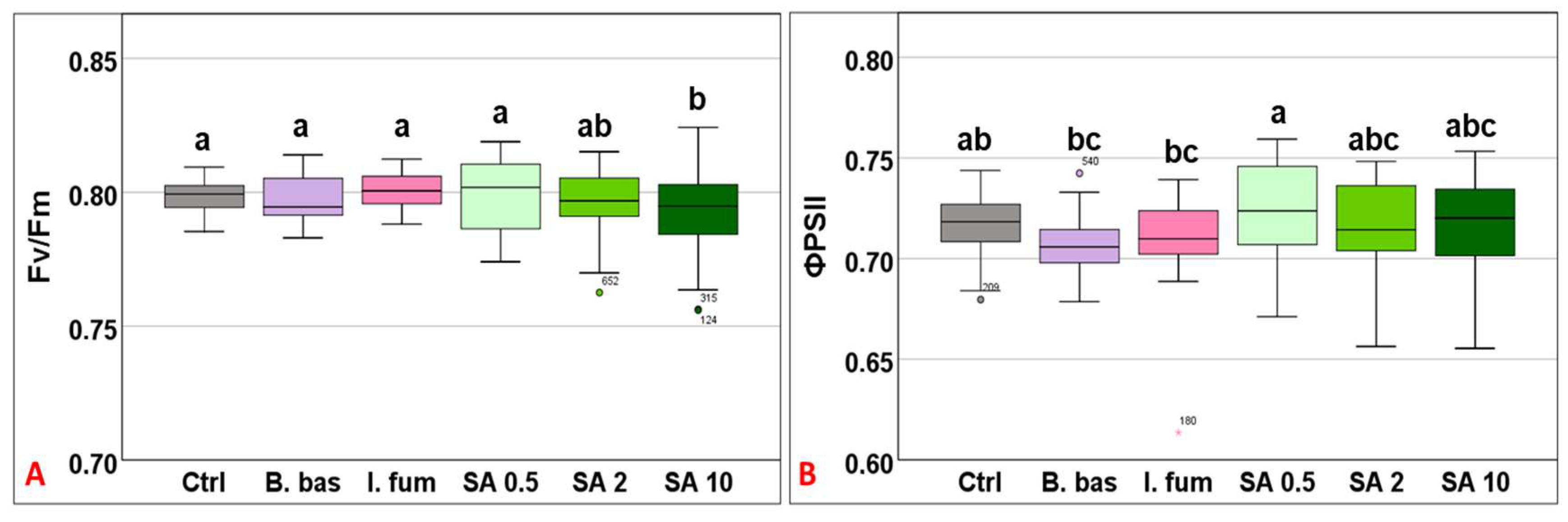
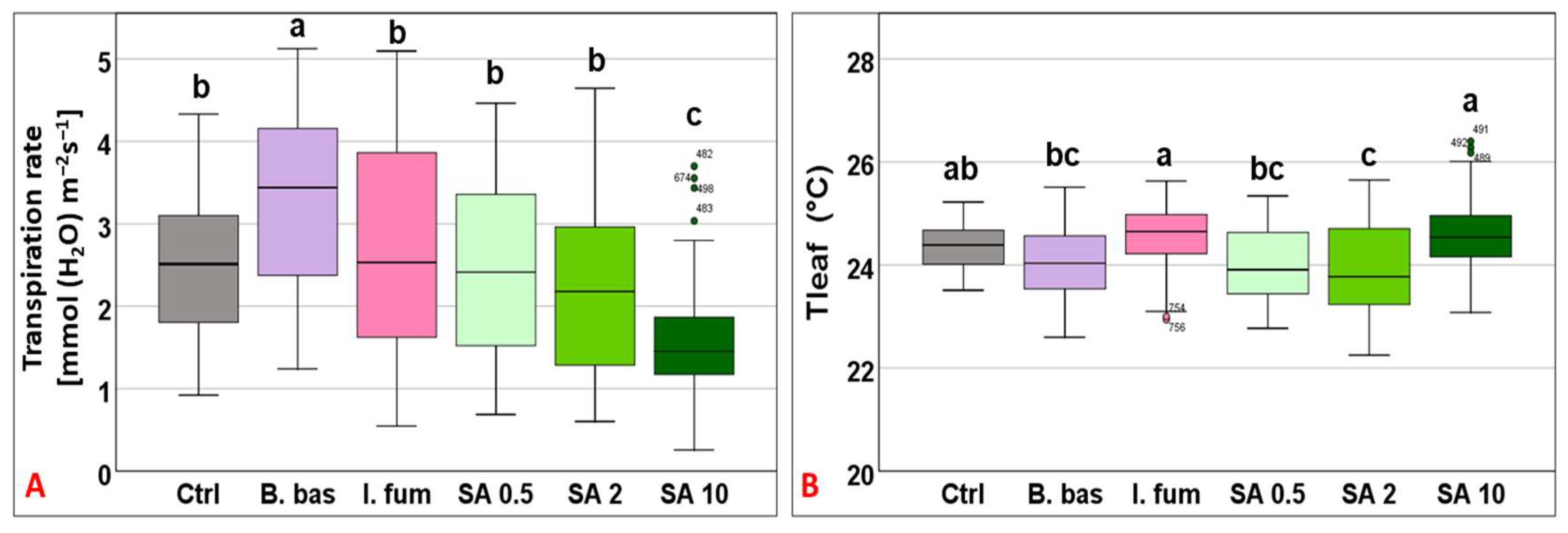
3.1. Evaluation of Grapevine Physiological Responses upon Foliar Application of Entomopathogenic Fungi and Salicylic Acid
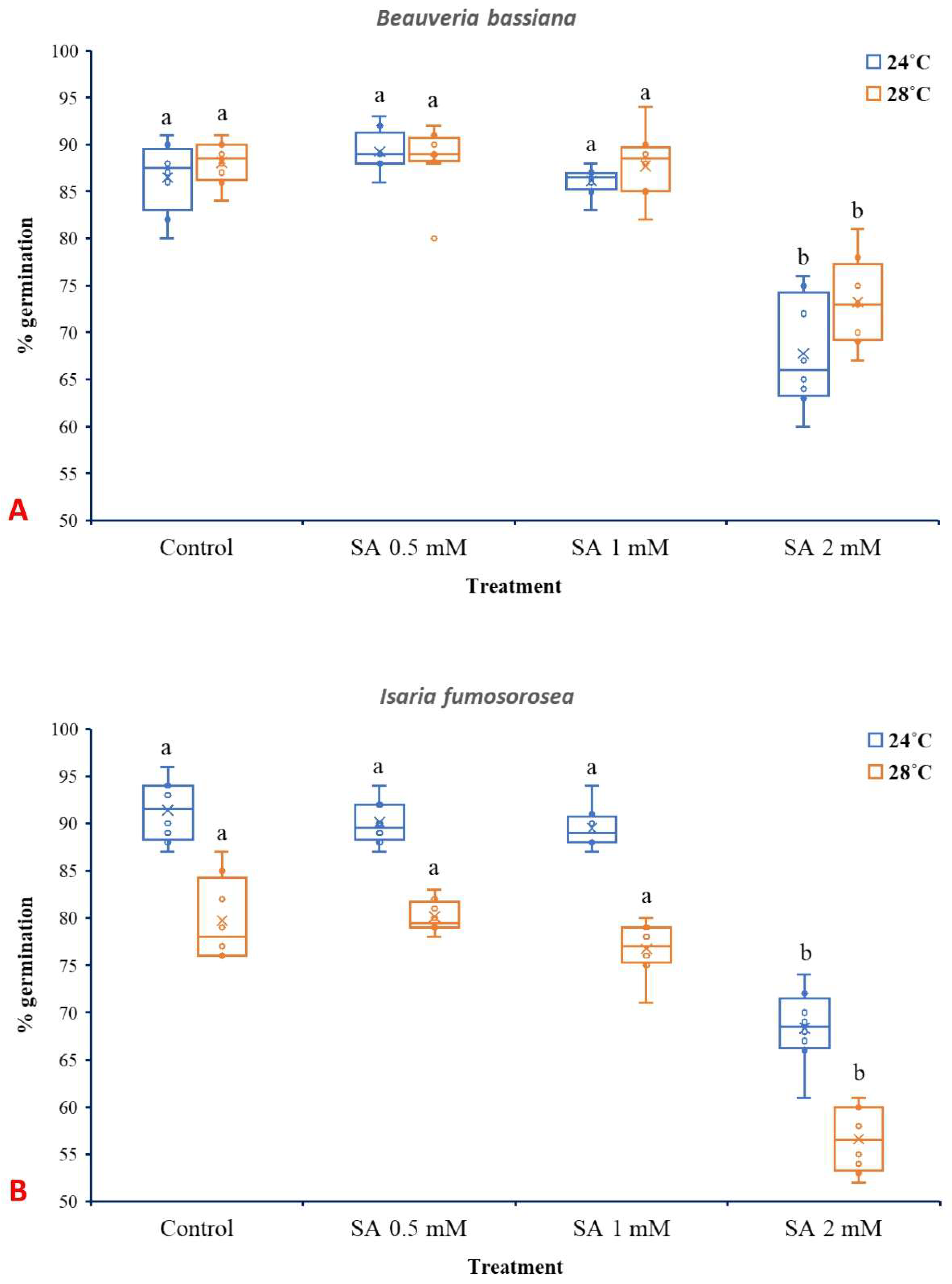
3.2. Effects of SA on Spore Germination of EPF
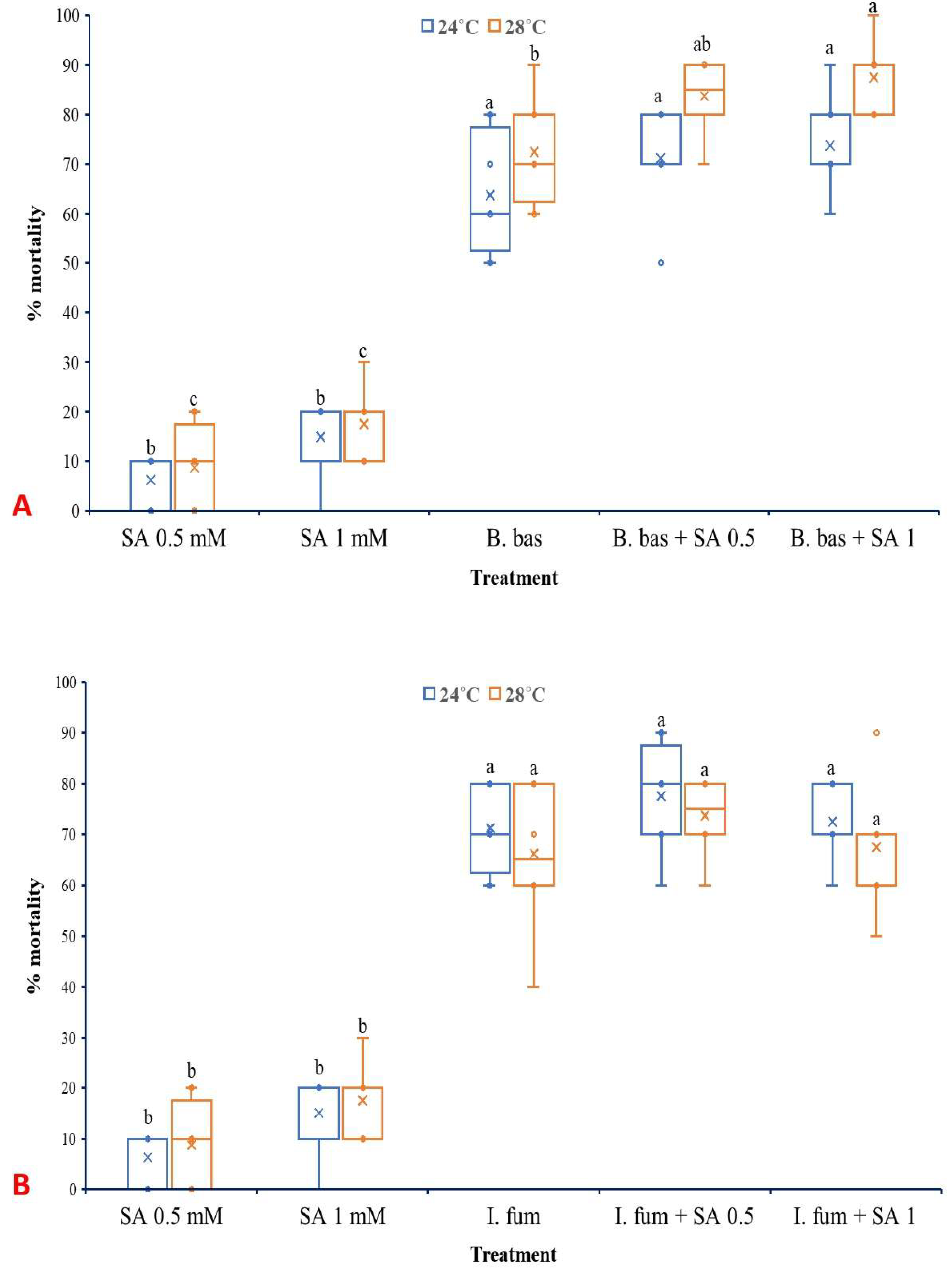
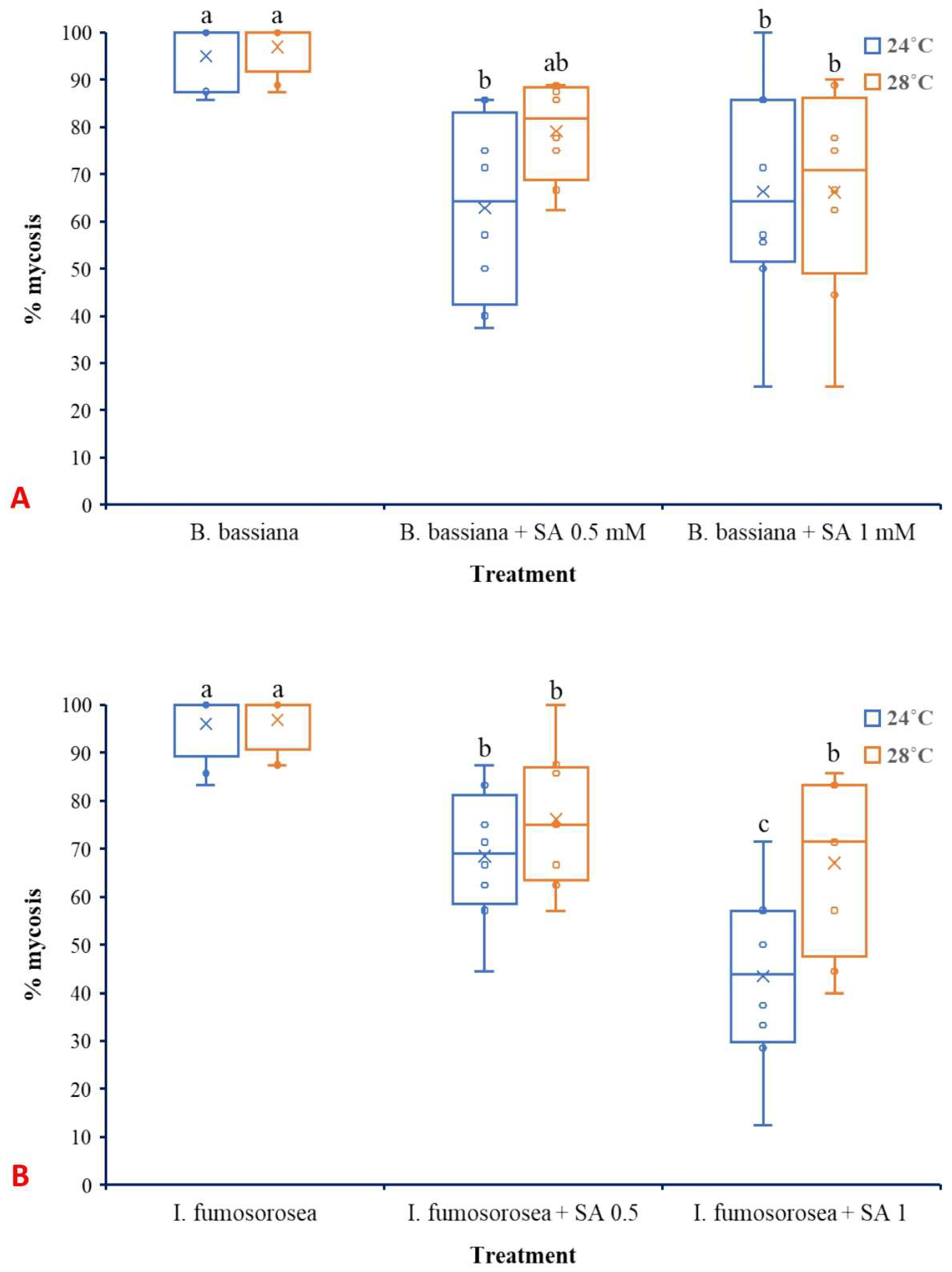
3.3. Effects of SA on Larval Mortality of L. botrana Caused by EPF
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| EPF | Entomopathogenic fungi |
| SA | Salicylic acid |
| ETR | Electron transport rate |
| PSII | Photosystem II |
| PSI | Photosystem I |
| E | Transpiration rate |
| gs | Stomatal conductance |
| Fv/Fm | Maximum quantum efficiency of PSII |
| IPM | Integrated pest management |
| JA | Jasmonic acid |
| Fs | steady-state fluorescence |
| Fm′ | maximum fluorescence in the light-adapted state |
| ΦPSII | actual quantum efficiency of PSII |
| Qamb | Ambient light |
| abs | Leaf light absorptance |
| PS2/1 | Ratio of PSII to PSII absorptance |
| Fm | maximum fluorescence |
| Fo | minimum fluorescence |
| Tleaf | Leaf temperature |
| IRT | non-contact infrared thermometer |
| S | leaf area |
| Mr | molar flow rate into the leaf cuvette |
| Wr | water vapor mole fractions into the leaf cuvette |
| Ws | water vapor mole fractions out of the leaf cuvette |
| gtw | total conductance to water vapor |
| gblw | boundary layer conductance |
| CEF | cyclic electron flow |
References
- Roy, H.E.; Brodie, E.L.; Chandler, D.; Goettel, M.S.; Pell, J.K.; Wajnberg, E.; Vega, F.E. Deep Space and Hidden Depths: Understanding the Evolution and Ecology of Fungal Entomopathogens. BioControl 2010, 55, 1–6. [Google Scholar] [CrossRef]
- Khun, K.K.; Wilson, B.A.L.; Stevens, M.M.; Huwer, R.K.; Ash, G.J. Integration of Entomopathogenic Fungi into IPM Programs: Studies Involving Weevils (Coleoptera: Curculionoidea) Affecting Horticultural Crops. Insects 2020, 11, 659. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.W.; Shin, T.Y.; Bae, S.M.; Woo, S.D. Screening and Evaluation of Entomopathogenic Fungi against the Green Peach Aphid, Myzus persicae, Using Multiple Tools. J. Asia Pac. Entomol. 2015, 18, 607–615. [Google Scholar] [CrossRef]
- Qasim, M.; Su, J.; Noman, A.; Ma, T.; Islam, W.; Hussain, D.; Rizwan, M.; Hameed, M.S.; Khan, K.A.; Ghramh, H.A.; et al. Citrus Psyllid Management by Collective Involvement of Plant Resistance, Natural Enemies and Entomopathogenic Fungi. Microb. Pathog. 2024, 197, 107047. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; González-Mas, N.; Yousef-Yousef, M.; Garrido-Jurado, I.; Fernández-Bravo, M. Key Role of Environmental Competence in Successful Use of Entomopathogenic Fungi in Microbial Pest Control. J. Pest Sci. 2024, 97, 1–15. [Google Scholar] [CrossRef]
- Vivekanandhan, P.; Kannan, S.; Pittarate, S.; Krutmuang, P. Classification, Biology and Entomopathogenic Fungi-Based Management and Their Mode of Action against Drosophila Species (Diptera: Drosophilidae): A Review. Front. Microbiol. 2024, 15, 1443651. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on Safety of the Entomopathogenic Fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci. Technol. 2007, 17, 553–596. [Google Scholar] [CrossRef]
- Shahid, A.; Rao, Q.; Bakhsh, A.; Husnain, T. Entomopathogenic Fungi as Biological Controllers: New Insights into Their Virulence and Pathogenicity. Arch. Biol. Sci. 2012, 64, 21–42. [Google Scholar] [CrossRef]
- Vega, F.E.; Goettel, M.S.; Blackwell, M.; Chandler, D.; Jackson, M.A.; Keller, S.; Koike, M.; Maniania, N.K.; Monzón, A.; Ownley, B.H.; et al. Fungal Entomopathogens: New Insights on Their Ecology. Fung. Ecol. 2009, 2, 149–159. [Google Scholar] [CrossRef]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect Pathogens as Biological Control Agents: Back to the Future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef]
- Kumar, P.; Joshi, A.; Sharma, N.; Lata, S.; Mehmood, S.; Ahlawat, Y.K.; Malik, A.; Moussa, I.M.; Kerketta, A.; Soni, P. Integrative Approaches to Improve Litchi (Litchi chinensis Sonn.) Plant Health Using Bio-Transformations and Entomopathogenic Fungi. BMC Plant Biol. 2024, 24, 902. [Google Scholar] [CrossRef] [PubMed]
- Vivekanandhan, P.; Alford, L.; Krutmuang, P. Editorial: Role of Entomopathogenic Fungi in Sustainable Agriculture. Front. Microbiol. 2024, 15, 1504175. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Siddiqui, J.A.; Akutse, K.S.; Ramos Aguila, L.C.; Xu, Y. General Limitations to Endophytic Entomopathogenic Fungi Use as Plant Growth Promoters, Pests and Pathogens Biocontrol Agents. Plants 2021, 10, 2119. [Google Scholar] [CrossRef] [PubMed]
- Parsa, S.; Ortiz, V.; Vega, F.E. Establishing Fungal Entomopathogens as Endophytes: Towards Endophytic Biological Control. J. Vis. Exp. 2013, 74, 50360. [Google Scholar] [CrossRef]
- Martins, J.L.A.; Franzin, M.L.; Ferreira, D.d.S.; Magina, L.C.R.; Martins, E.F.; Mendonça, L.V.P.; Neves, W.d.S.; Pallini, A.; Valicente, F.H.; Schmidt, J.M.; et al. Metarhizium-Inoculated Coffee Seeds Promote Plant Growth and Biocontrol of Coffee Leaf Miner. Microorganisms 2024, 12, 1845. [Google Scholar] [CrossRef] [PubMed]
- Mantzoukas, S.; Daskalaki, E.; Kitsiou, F.; Papantzikos, V.; Servis, D.; Bitivanos, S.; Patakioutas, G.; Eliopoulos, P.A. Dual Action of Beauveria bassiana (Hypocreales; Cordycipitaceae) Endophytic Stains as Biocontrol Agents against Sucking Pests and Plant Growth Biostimulants on Melon and Strawberry Field Plants. Microorganisms 2022, 10, 2306. [Google Scholar] [CrossRef]
- Ponchon, M.; Reineke, A.; Massot, M.; Bidochka, M.J.; Thiéry, D.; Papura, D. Three Methods Assessing the Association of the Endophytic Entomopathogenic Fungus Metarhizium robertsii with Non-Grafted Grapevine Vitis vinifera. Microorganisms 2022, 10, 2437. [Google Scholar] [CrossRef]
- Rondot, Y.; Reineke, A. Endophytic Beauveria bassiana in Grapevine Vitis vinifera (L.) Reduces Infestation with Piercing-Sucking Insects. Biol. Control 2018, 116, 82–89. [Google Scholar] [CrossRef]
- Galland, C.D.; Lalaymia, I.; Declerck, S.; Verheggen, F. Efficacy of Entomopathogenic Fungi against the Fruit Fly Drosophila Suzukii and Their Side Effects on Predator (Orius laevigatus) and Pollinator (Bombus terrestris) Insects. Entomol. Gen. 2023, 43, 1203–1210. [Google Scholar] [CrossRef]
- Pope, T.W.; Hough, G.; Arbona, C.; Roberts, H.; Bennison, J.; Buxton, J.; Prince, G.; Chandler, D. Investigating the Potential of an Autodissemination System for Managing Populations of Vine Weevil, Otiorhynchus sulcatus (Coleoptera: Curculionidae) with Entomopathogenic Fungi. J. Invertebr. Pathol. 2018, 154, 79–84. [Google Scholar] [CrossRef]
- Aguilera Sammaritano, J.; Deymié, M.; Herrera, M.; Vazquez, F.; Cuthbertson, A.G.S.; López-Lastra, C.; Lechner, B. The Entomopathogenic Fungus, Metarhizium anisopliae for the European Grapevine Moth, Lobesia botrana Den. & Schiff. (Lepidoptera: Tortricidae) and Its Effect to the Phytopathogenic Fungus, Botrytis Cinerea. Egypt J. Biol. Pest Control 2018, 28, 83. [Google Scholar] [CrossRef]
- Altimira, F.; De La Barra, N.; Rebufel, P.; Soto, S.; Soto, R.; Estay, P.; Vitta, N.; Tapia, E. Potential Biological Control of the Pupal Stage of the European Grapevine Moth Lobesia Botrana by the Entomopathogenic Fungus Beauveria pseudobassiana in the Winter Season in Chile. BMC Res. Notes 2019, 12, 548. [Google Scholar] [CrossRef]
- López Plantey, R.; Papura, D.; Couture, C.; Thiéry, D.; Pizzuolo, P.H.; Bertoldi, M.V.; Lucero, G.S. Characterization of Entomopathogenic Fungi from Vineyards in Argentina with Potential as Biological Control Agents against the European Grapevine Moth Lobesia botrana. BioControl 2019, 64, 501–511. [Google Scholar] [CrossRef]
- Altimira, F.; De La Barra, N.; Godoy, P.; Roa, J.; Godoy, S.; Vitta, N.; Tapia, E. Lobesia botrana: A Biological Control Approach with a Biopesticide Based on Entomopathogenic Fungi in the Winter Season in Chile. Insects 2021, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Arias-Aravena, M.; Altimira, F.; Gutiérrez, D.; Ling, J.; Tapia, E. Identification of Exoenzymes Secreted by Entomopathogenic Fungus Beauveria pseudobassiana RGM 2184 and Their Effect on the Degradation of Cocoons and Pupae of Quarantine Pest Lobesia botrana. JoF 2022, 8, 1083. [Google Scholar] [CrossRef]
- Beris, E.; Papachristos, D.; Ponchon, M.; Caca, D.; Kontodimas, D.; Reineke, A. The Effects of Temperature on Pathogenicity of Entomopathogenic Fungi for Controlling Larval Populations of the European Grapevine Moth (Lobesia botrana) (Lepidoptera: Tortricidae). Crop Prot. 2024, 177, 106542. [Google Scholar] [CrossRef]
- Aguilera-Sammaritano, J.; Caballero, J.; Deymié, M.; Rosa, M.; Vazquez, F.; Pappano, D.; Lechner, B.; González-Teuber, M. Dual Effects of Entomopathogenic Fungi on Control of the Pest Lobesia botrana and the Pathogenic Fungus Eutypella microtheca on Grapevine. Biol. Res. 2021, 54, 44. [Google Scholar] [CrossRef] [PubMed]
- Beris, E.; Korkas, E. Additive and Synergistic Interactions of Entomopathogenic Fungi with Bacillus thuringiensis for the Control of the European Grapevine Moth Lobesia botrana (Denis and Schiffermüller) (Lepidoptera: Tortricidae). Egypt J. Biol. Pest Control 2021, 31, 109. [Google Scholar] [CrossRef]
- Andreadis, S.S.; Milonas, P.G.; Savopoulou-Soultani, M. Cold Hardiness of Diapausing and Non-diapausing Pupae of the European Grapevine Moth, Lobesia botrana. Entomol. Exp. Appl. 2005, 117, 113–118. [Google Scholar] [CrossRef]
- Moreau, J.; Villemant, C.; Benrey, B.; Thiéry, D. Species Diversity of Larval Parasitoids of the European Grapevine Moth (Lobesia botrana, Lepidoptera: Tortricidae): The Influence of Region and Cultivar. Biol. Control 2010, 54, 300–306. [Google Scholar] [CrossRef]
- Benelli, G.; Lucchi, A.; Anfora, G.; Bagnoli, B.; Botton, M.; Campos-Herrera, R.; Carlos, C.; Daugherty, M.P.; Gemeno, C.; Harari, A.R.; et al. European Grapevine Moth, Lobesia botrana Part I: Biology and ecology. Entomol. Gen. 2023, 43, 261–280. [Google Scholar] [CrossRef]
- Iltis, C.; Moreau, J.; Pecharová, K.; Thiéry, D.; Louâpre, P. Reproductive Performance of the European Grapevine Moth Lobesia botrana (Tortricidae) Is Adversely Affected by Warming Scenario. J. Pest Sci. 2020, 93, 679–689. [Google Scholar] [CrossRef]
- Pasquini, S.; Haxaire-Lutun, M.O.; Rison, J.-L.; Flier, W.G.; Teixeira, L.A. Susceptibility of Lobesia botrana (Lepidoptera: Tortricidae) to Chlorantraniliprole in the Emilia Romagna Region of Northeast Italy. J. Econ. Entomol. 2018, 111, 369–374. [Google Scholar] [CrossRef]
- Civolani, S.; Boselli, M.; Butturini, A.; Chicca, M.; Fano, E.A.; Cassanelli, S. Assessment of Insecticide Resistance of Lobesia botrana (Lepidoptera: Tortricidae) in Emilia-Romagna Region. J. Econ. Entomol. 2014, 107, 1245–1249. [Google Scholar] [CrossRef]
- Caboni, P.; Cabras, P. Pesticides’ Influence on Wine Fermentation. In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2010; Volume 59, pp. 43–62. ISBN 978-0-12-380942-1. [Google Scholar]
- Ioriatti, C.; Anfora, G.; Tasin, M.; De Cristofaro, A.; Witzgall, P.; Lucchi, A. Chemical Ecology and Management of Lobesia botrana (Lepidoptera: Tortricidae). J. Econ. Entom. 2011, 104, 1125–1137. [Google Scholar] [CrossRef] [PubMed]
- Lucchi, A.; Sambado, P.; Juan Royo, A.B.; Bagnoli, B.; Conte, G.; Benelli, G. Disrupting Mating of Lobesia botrana Using Sex Pheromone Aerosol Devices. Environ. Sci. Pollut. Res. 2018, 25, 22196–22204. [Google Scholar] [CrossRef]
- Benelli, G.; Lucchi, A.; Anfora, G.; Bagnoli, B.; Botton, M.; Campos-Herrera, R.; Carlos, C.; Daugherty, M.P.; Gemeno, C.; Harari, A.R.; et al. European Grapevine Moth, Lobesia botrana Part II: Prevention and Management. Entomol. Gen. 2023, 43, 281–304. [Google Scholar] [CrossRef]
- Benelli, G.; Pavoni, L.; Zeni, V.; Ricciardi, R.; Cosci, F.; Cacopardo, G.; Gendusa, S.; Spinozzi, E.; Petrelli, R.; Cappellacci, L.; et al. Developing a Highly Stable Carlina acaulis Essential Oil Nanoemulsion for Managing Lobesia botrana. Nanomaterials 2020, 10, 1867. [Google Scholar] [CrossRef] [PubMed]
- Koo, Y.M.; Heo, A.Y.; Choi, H.W. Salicylic Acid as a Safe Plant Protector and Growth Regulator. Plant Pathol. J. 2020, 36, 1–10. [Google Scholar] [CrossRef]
- Liu, J.; Qiu, G.; Liu, C.; Li, H.; Chen, X.; Fu, Q.; Lin, Y.; Guo, B. Salicylic Acid, a Multifaceted Hormone, Combats Abiotic Stresses in Plants. Life 2022, 12, 886. [Google Scholar] [CrossRef]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-Related Proteins and Peptides as Promising Tools for Engineering Plants with Multiple Stress Tolerance. Microbiol. Res. 2018, 212–213, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Shao, H.; Zheng, A.; Zhao, L.; Xu, Y. Advances in Roles of Salicylic Acid in Plant Tolerance Responses to Biotic and Abiotic Stresses. Plants 2023, 12, 3475. [Google Scholar] [CrossRef]
- Rivas-San Vicente, M.; Plasencia, J. Salicylic Acid beyond Defence: Its Role in Plant Growth and Development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef] [PubMed]
- Dobón-Suárez, A.; Giménez, M.J.; García-Pastor, M.E.; Zapata, P.J. Salicylic Acid Foliar Application Increases Crop Yield and Quality Parameters of Green Pepper Fruit during Postharvest Storage. Agronomy 2021, 11, 2263. [Google Scholar] [CrossRef]
- Chen, C.; Sun, C.; Wang, Y.; Gong, H.; Zhang, A.; Yang, Y.; Guo, F.; Cui, K.; Fan, X.; Li, X. The Preharvest and Postharvest Application of Salicylic Acid and Its Derivatives on Storage of Fruit and Vegetables: A Review. Sci. Hortic. 2023, 312, 111858. [Google Scholar] [CrossRef]
- Walling, L.L. The Myriad Plant Responses to Herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef]
- Setotaw, Y.B.; Li, J.; Qi, J.; Ma, C.; Zhang, M.; Huang, C.; Wang, L.; Wu, J. Salicylic Acid Positively Regulates Maize Defenses against Lepidopteran Insects. Plant Divers. 2024, 46, 519–529. [Google Scholar] [CrossRef]
- Beris, E. Evaluation and Environmental Testing of Entomopathogenic Fungi for Their Effectiveness as Bio-Control Agents of Major Vineyard Pests. Plant Prot. 2021, 5, 13–23. [Google Scholar] [CrossRef]
- Giménez, M.J.; Valverde, J.M.; Valero, D.; Guillén, D.; Martínez-Romero, D.; Serrano, M.; Castillo, S. Quality and Antioxidant Properties on Sweet Cherries as Affected by Preharvest Salicylic and Acetylsalicylic Acids Treatments. Food Chem. 2014, 160, 226–232. [Google Scholar] [CrossRef]
- Vega, F.E.; Dowd, P.F.; McGuire, M.R.; Jackson, M.A.; Nelsen, T.C. In VitroEffects of Secondary Plant Compounds on Germination of Blastospores of the Entomopathogenic Fungus Paecilomyces fumosoroseus (Deuteromycotina: Hyphomycetes). J. Invertebr. Pathol. 1997, 70, 209–213. [Google Scholar] [CrossRef]
- Strobel, N.E.; Porter, L.A. Salicylate Inhibits Growth of Plant-Pathogenic Fungi and Synergistically Enhances the Activity of Other Antifungal Materials In Vitro. J. Ky. Acad. Sci. 2005, 66, 118–128. [Google Scholar]
- Onsongo, S.K.; Mohamed, S.A.; Akutse, K.S.; Gichimu, B.M.; Dubois, T. The Entomopathogenic Fungi Metarhizium anisopliae and Beauveria bassiana for Management of the Melon Fly Zeugodacus cucurbitae: Pathogenicity, Horizontal Transmission, and Compatability with Cuelure. Insects 2022, 13, 859. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Moreiras, A.M.; Graña, E.; Reigosa, M.J.; Araniti, F. Imaging of Chlorophyll a Fluorescence in Natural Compound-Induced Stress Detection. Front. Plant Sci. 2020, 11, 583590. [Google Scholar] [CrossRef] [PubMed]
- Moustaka, J.; Meyling, N.V.; Hauser, T.P. Root-Associated Entomopathogenic Fungi Modulate Their Host Plant’s Photosystem II Photochemistry and Response to Herbivorous Insects. Molecules 2021, 27, 207. [Google Scholar] [CrossRef] [PubMed]
- Oue, H. Comparisons of the Stomatal Conductance and Electron Transport Rate of Three Japanese Rice Cultivars Including Himenorin in Ehime Prefecture. J. Agric. Meteorol. 2023, 79, 77–84. [Google Scholar] [CrossRef]
- Ahsan, S.M.; Injamum-Ul-Hoque, M.; Das, A.K.; Rahman, M.M.; Mollah, M.M.I.; Paul, N.C.; Choi, H.W. Plant–Entomopathogenic Fungi Interaction: Recent Progress and Future Prospects on Endophytism-Mediated Growth Promotion and Biocontrol. Plants 2024, 13, 1420. [Google Scholar] [CrossRef]
- Wilberts, L.; Rojas-Preciado, N.; Jacquemyn, H.; Lievens, B. Fungal Strain and Crop Cultivar Affect Growth of Sweet Pepper Plants after Root Inoculation with Entomopathogenic Fungi. Front. Plant Sci. 2023, 14, 1196765. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, Y.; Wang, B. Entomopathogenic Fungi Beauveria bassiana and Metarhizium anisopliae Play Roles of Maize (Zea mays) Growth Promoter. Sci. Rep. 2022, 12, 15706. [Google Scholar] [CrossRef]
- Chaudhary, P.J.; B.L., R.; Patel, H.K.; Mehta, P.V.; Patel, N.B.; Sonth, B.; Dave, A.; Bagul, S.Y.; M., D.; Jain, D.; et al. Plant Growth-Promoting Potential of Entomopathogenic Fungus Metarhizium pinghaense AAUBC-M26 under Elevated Salt Stress in Tomato. Agronomy 2023, 13, 1577. [Google Scholar] [CrossRef]
- Wang, L.-J.; Fan, L.; Loescher, W.; Duan, W.; Liu, G.-J.; Cheng, J.-S.; Luo, H.-B.; Li, S.-H. Salicylic Acid Alleviates Decreases in Photosynthesis under Heat Stress and Accelerates Recovery in Grapevine Leaves. BMC Plant Biol. 2010, 10, 34. [Google Scholar] [CrossRef]
- Gonçalves, F.C.D.M.; Mantoan, L.P.B.; Corrêa, C.V.; Parreiras, N.D.S.; De Almeida, L.F.R.; Ono, E.O.; Rodrigues, J.D.; Prado, R.D.M.; Boaro, C.S.F. Effects of Salicylic Acid on Physiological Responses of Pepper Plants Pre-Subjected to Drought under Rehydration Conditions. Plants 2024, 13, 2805. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha Neto, A.C.; Maraschin, M.; Di Piero, R.M. Antifungal Activity of Salicylic Acid against Penicillium expansum and Its Possible Mechanisms of Action. Int. J. Food Microbiol. 2015, 215, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhu, T.; Song, Y.; Feng, L.; Kear, P.J.; Riseh, R.S.; Sitohy, M.; Datla, R.; Ren, M. Salicylic Acid Fights against Fusarium Wilt by Inhibiting Target of Rapamycin Signaling Pathway in Fusarium oxysporum. J. Adv. Res. 2022, 39, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gacnik, S.; Munda, A.; Veberic, R.; Hudina, M.; Mikulic-Petkovsek, M. Preventive and Curative Effects of Salicylic and Methyl Salicylic Acid Having Antifungal Potential against Monilinia laxa and the Development of Phenolic Response in Apple Peel. Plants 2023, 12, 1584. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Baldwin, L.A. Hormesis: The Dose-Response Revolution. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 175–197. [Google Scholar] [CrossRef]
- Mishra, S.; Kumar, P.; Malik, A. Effect of Temperature and Humidity on Pathogenicity of Native Beauveria bassiana Isolate against Musca domestica L. J. Parasit. Dis. 2015, 39, 697–704. [Google Scholar] [CrossRef]
- Athanassiou, C.G.; Kavallieratos, N.G.; Rumbos, C.I.; Kontodimas, D.C. Influence of Temperature and Relative Humidity on the Insecticidal Efficacy of Metarhizium anisopliae against Larvae of Ephestia kuehniella (Lepidoptera: Pyralidae) on Wheat. J. Insect Sci. 2017, 17, 22. [Google Scholar] [CrossRef]
- Lazzarini, G.M.J.; Rocha, L.F.N.; Luz, C. Impact of Moisture on in Vitro Germination of Metarhizium anisopliae and Beauveria bassiana and Their Activity on Triatoma infestans. Mycol. Res. 2006, 110, 485–492. [Google Scholar] [CrossRef]
- Bohatá, A.; Folorunso, E.A.; Lencová, J.; Osborne, L.S.; Mraz, J. Control of Sweet Potato Whitefly (Bemisia tabaci) Using Entomopathogenic Fungi under Optimal and Suboptimal Relative Humidity Conditions. Pest Manag. Sci. 2024, 80, 1065–1075. [Google Scholar] [CrossRef]
- Mollah, M.M.I.; Choi, H.W.; Yeam, I.; Lee, J.M.; Kim, Y. Salicylic Acid, a Plant Hormone, Suppresses Phytophagous Insect Immune Response by Interrupting HMG-Like DSP1. Front. Physiol. 2021, 12, 744272. [Google Scholar] [CrossRef]
- Yang, J.; Wang, Y.; Liu, L.; Liu, L.; Wang, C.; Wang, C.; Li, C. Effects of Exogenous Salicylic Acid and pH on Pathogenicity of Biotrophy-Associated Secreted Protein 1 (BAS1)-Overexpressing Strain, Magnaporthe oryzae. Environ. Sci. Pollut. Res. 2019, 26, 13725–13737. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beris, E.; Venios, X.; Papachristos, D.; Ponchon, M.; Kontodimas, D.; Korkas, E.; Banilas, G.; Reineke, A. Grapevine Responses to the Entomopathogenic Fungi Beauveria bassiana and Isaria fumosorosea and the Effects of Salicylic Acid on Their Virulence Against the European Grapevine Moth, Lobesia botrana. Microorganisms 2025, 13, 1630. https://doi.org/10.3390/microorganisms13071630
Beris E, Venios X, Papachristos D, Ponchon M, Kontodimas D, Korkas E, Banilas G, Reineke A. Grapevine Responses to the Entomopathogenic Fungi Beauveria bassiana and Isaria fumosorosea and the Effects of Salicylic Acid on Their Virulence Against the European Grapevine Moth, Lobesia botrana. Microorganisms. 2025; 13(7):1630. https://doi.org/10.3390/microorganisms13071630
Chicago/Turabian StyleBeris, Evangelos, Xenophon Venios, Dimitrios Papachristos, Mathilde Ponchon, Dimitrios Kontodimas, Elias Korkas, Georgios Banilas, and Annette Reineke. 2025. "Grapevine Responses to the Entomopathogenic Fungi Beauveria bassiana and Isaria fumosorosea and the Effects of Salicylic Acid on Their Virulence Against the European Grapevine Moth, Lobesia botrana" Microorganisms 13, no. 7: 1630. https://doi.org/10.3390/microorganisms13071630
APA StyleBeris, E., Venios, X., Papachristos, D., Ponchon, M., Kontodimas, D., Korkas, E., Banilas, G., & Reineke, A. (2025). Grapevine Responses to the Entomopathogenic Fungi Beauveria bassiana and Isaria fumosorosea and the Effects of Salicylic Acid on Their Virulence Against the European Grapevine Moth, Lobesia botrana. Microorganisms, 13(7), 1630. https://doi.org/10.3390/microorganisms13071630