


Impact of Chemotherapy Alone and in Combination with Immunotherapy on Oral Microbiota in Cancer Patients—A Pilot Study
 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
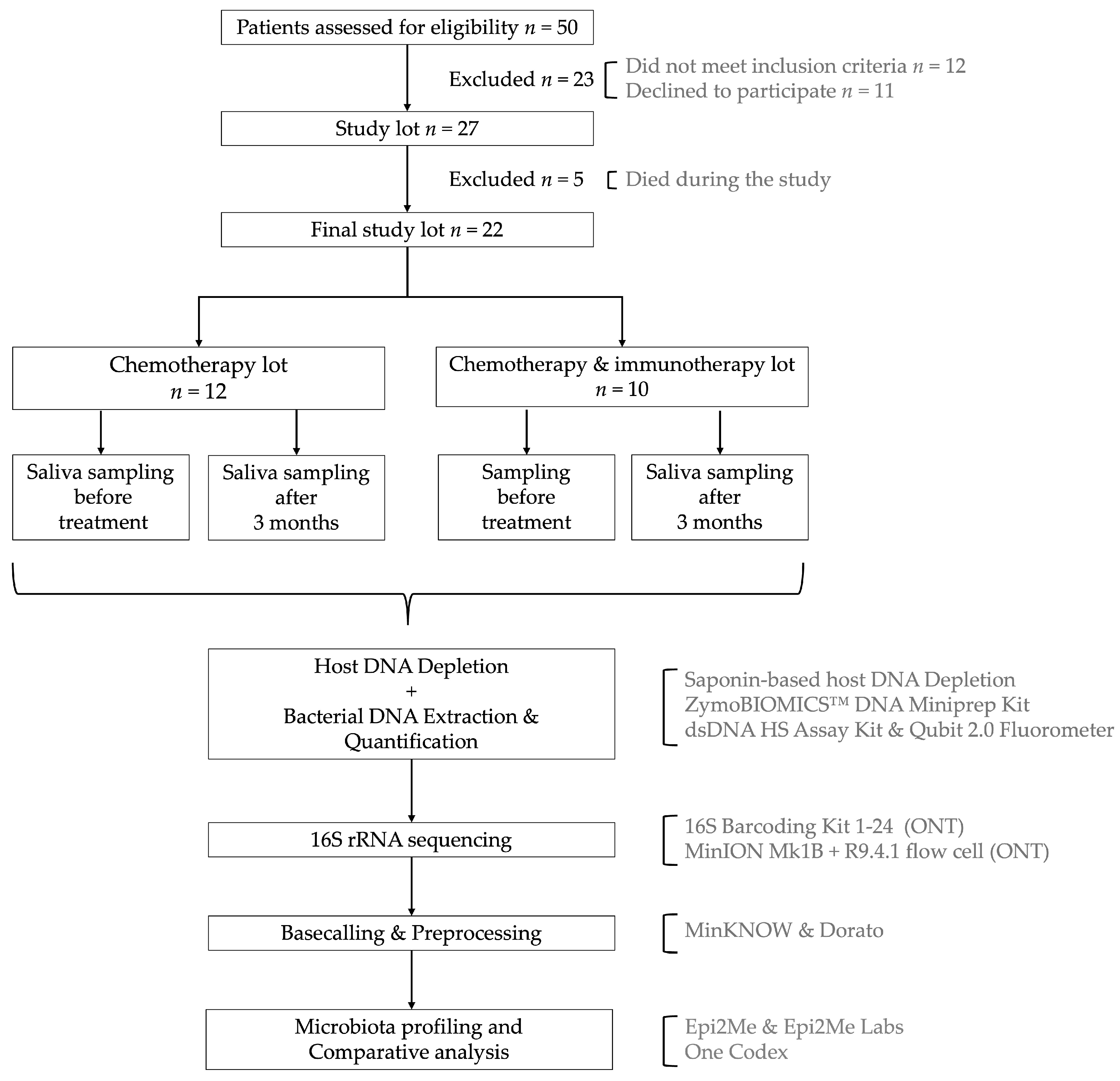
2.1. Study Design and Sample Collection
2.2. Bacterial DNA Extraction and Purification
- Centrifugation: 2 mL of saliva preserved in DNA/RNA Shield (Zymo Research, Irvine, CA, US) was centrifuged at 10,000× g for 7 min.
- Supernatant Removal and Resuspension: The supernatant was discarded, and the pellet was resuspended in 250 µL phosphate-buffered saline (PBS) (Thermo Fisher, Waltham, MA, USA).
- Saponin Treatment: 200 µL of 5% saponin (Sigma-Aldrich, St. Louis, MO, USA) solution was added, followed by 10 min of incubation at room temperature.
- Cell Lysis:
- ○
- 350 µL of nuclease free water was added and incubated for 30 s at room temperature.
- ○
- 12 µL of 5M NaCl was introduced and the mixture was centrifuged at 6000× g for 5 min at room temperature.
- Heat-Labile Salt Active Nuclease (HL-SAN) Treatment: The pellet was resuspended in:
- ○
- 100 µL PBS (Thermo Fisher, Waltham, MA, USA),
- ○
- 100 µL HL-SAN Buffer (5.5 M NaCl + 100 mM MgCl2) (Sigma-Aldrich, St. Louis, MO, USA),
- ○
- 10 µL HL-SAN DNase (Articzymes Technologies ASA, Tromsø, Norway),
- ○
- Followed by incubation at 37 °C with shaking at 800 rpm for 15 min.
- Pellet Washing: After another centrifugation (6000× g for 3 min), the pellet was washed with 800 µL of PBS (Thermo Fisher, Waltham, MA, USA) and then 1000 µL of PBS (Thermo Fisher, Waltham, MA, USA), followed by a final centrifugation (6000× g for 3 min).
- Bacterial DNA Extraction and Purification: The final pellet was subjected to DNA extraction and purification using a ZymoBIOMICSTM DNA Miniprep Kit (Zymo Research, Irvine, CA, USA), following the manufacturer’s protocol.
2.3. Library Preparation for 16S rRNA Sequencing
2.4. Sequencing Data Analysis
2.5. Statistical Analysis
3. Results
3.1. Sequencing Quality Control
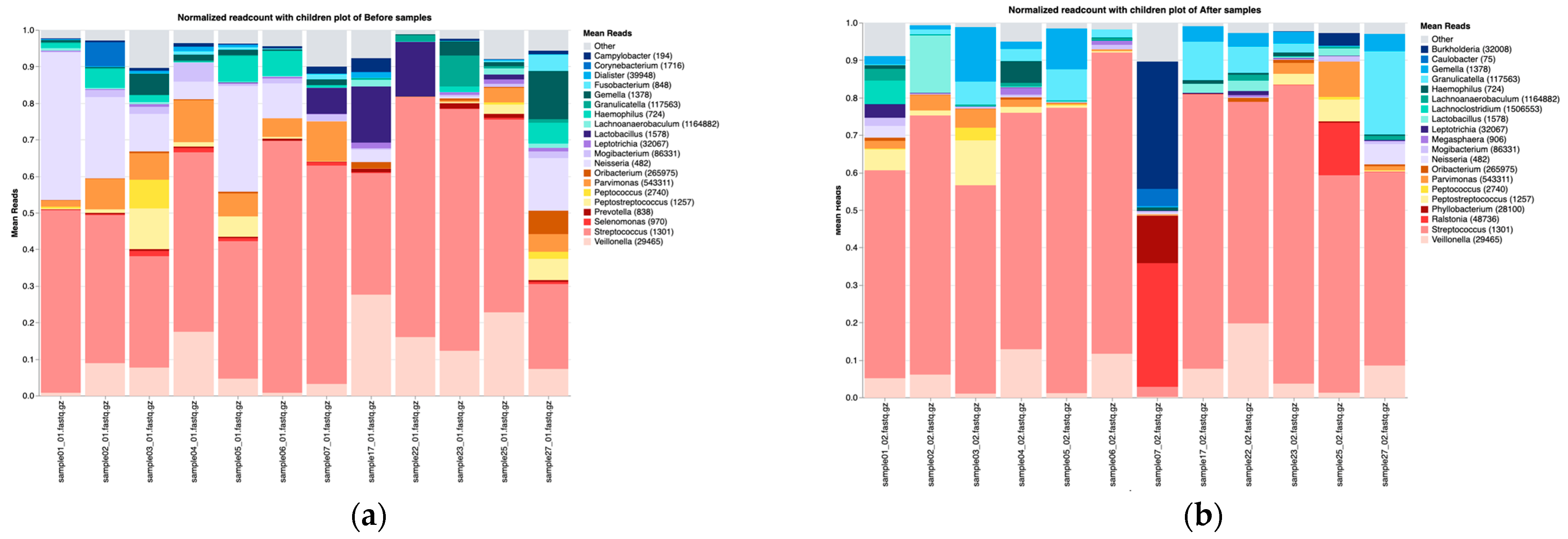
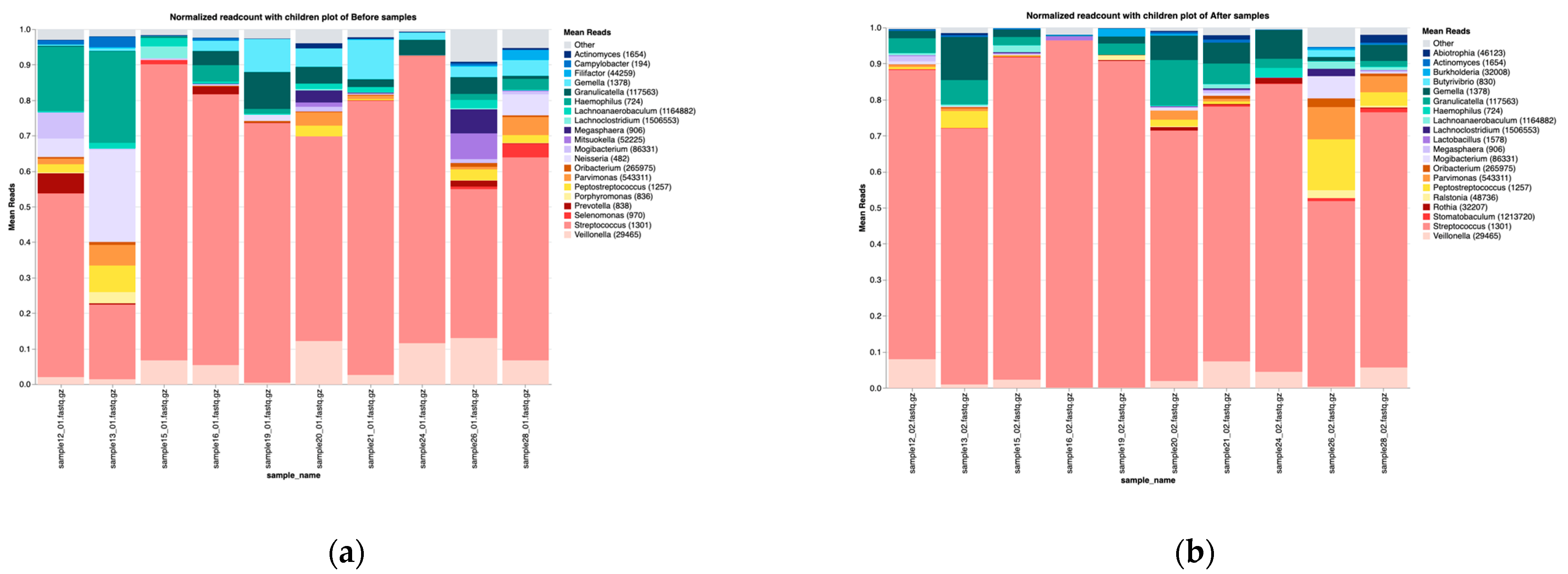
3.2. Overall Microbiota Composition Before and After Treatment
3.3. Microbiota Changes Following Chemotherapy Treatment
3.3.1. Taxonomic Composition
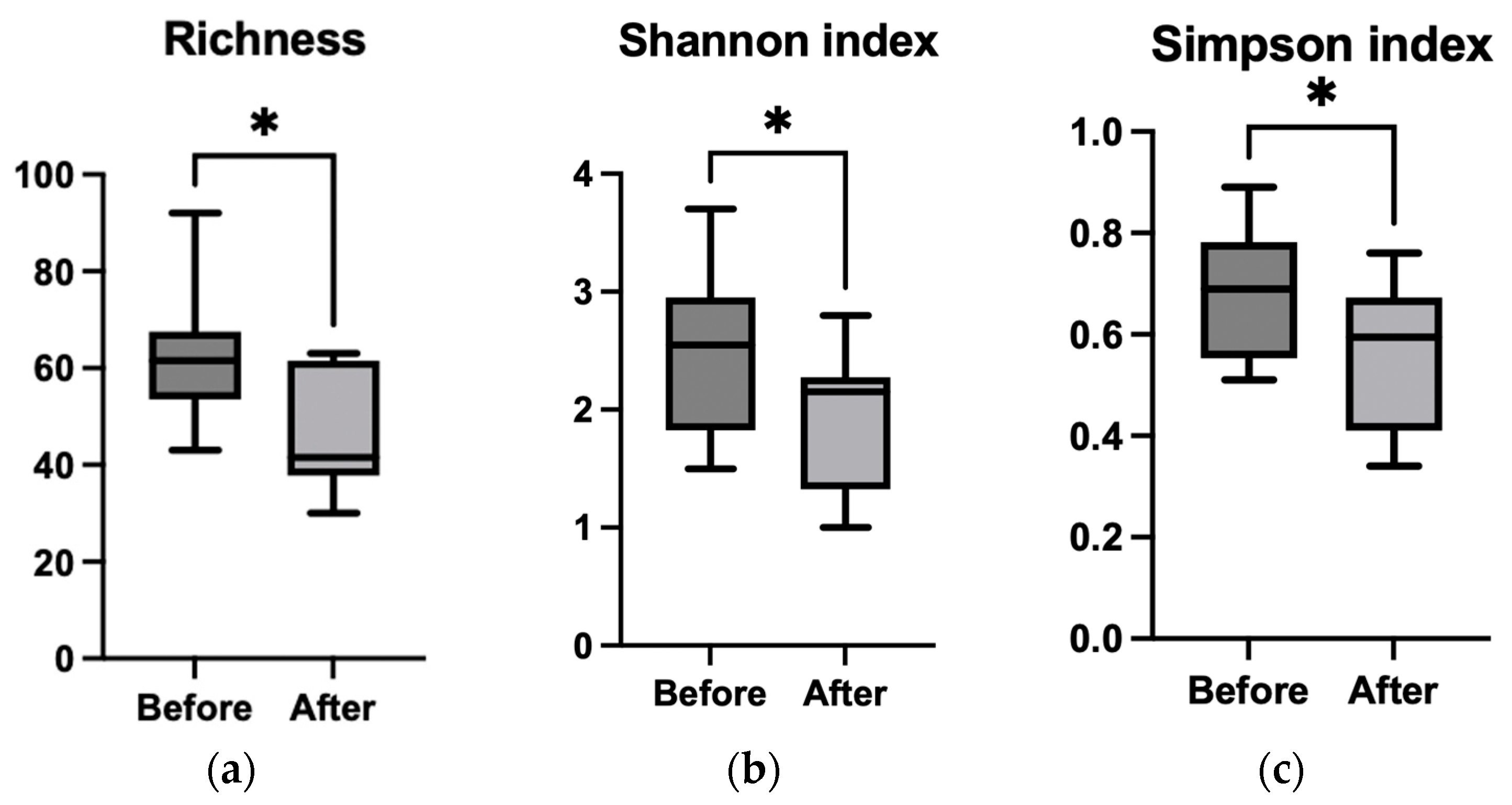
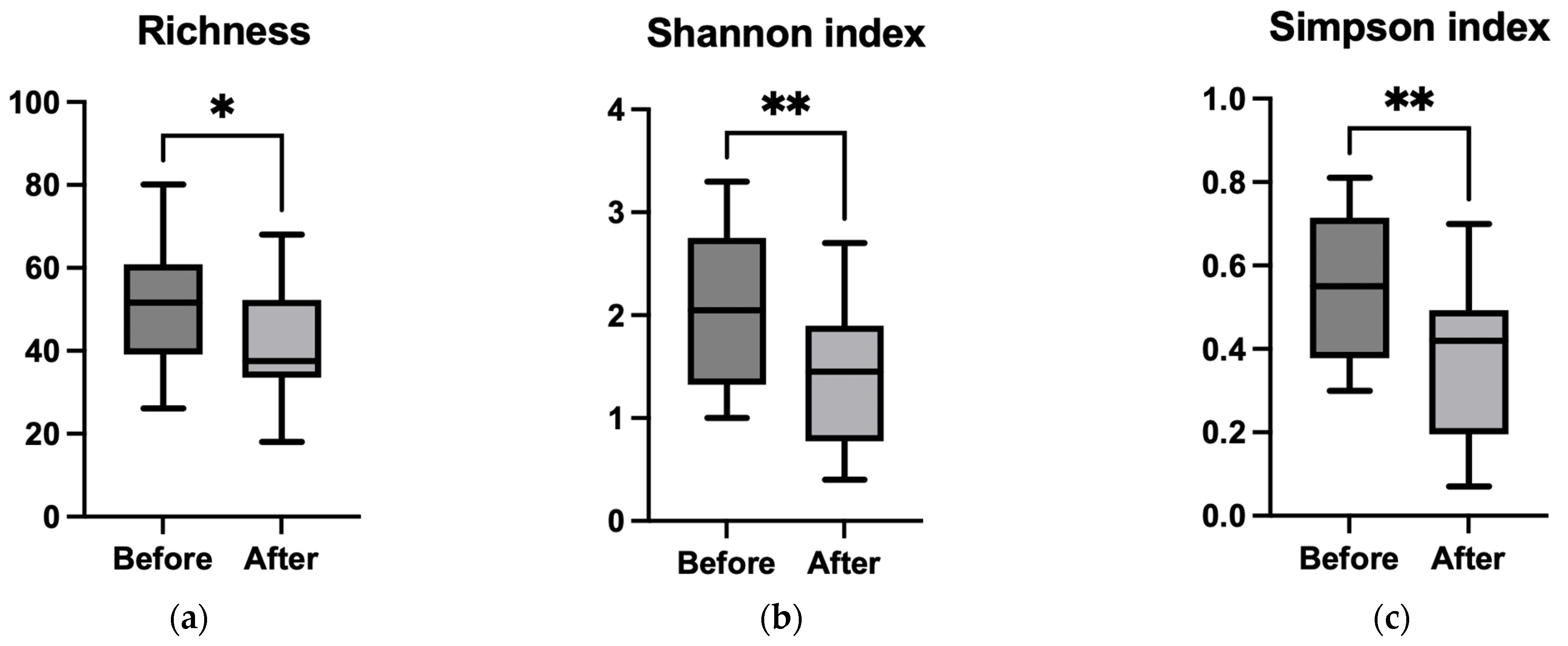
3.3.2. Alpha Diversity
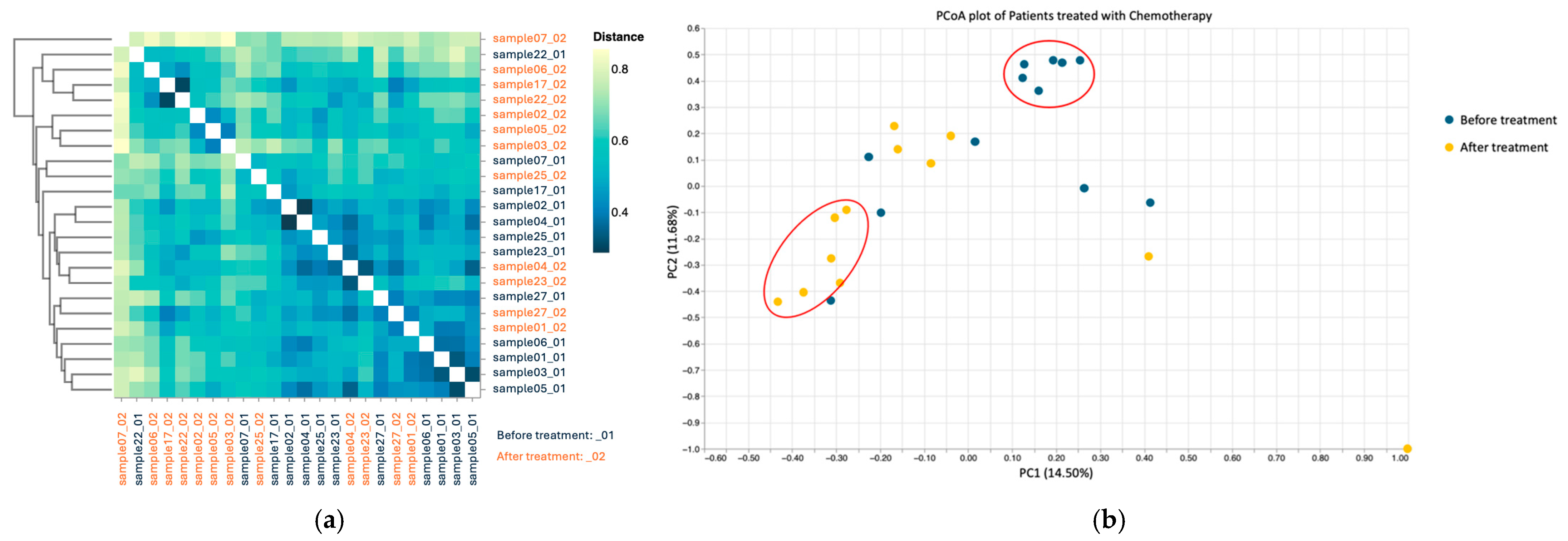
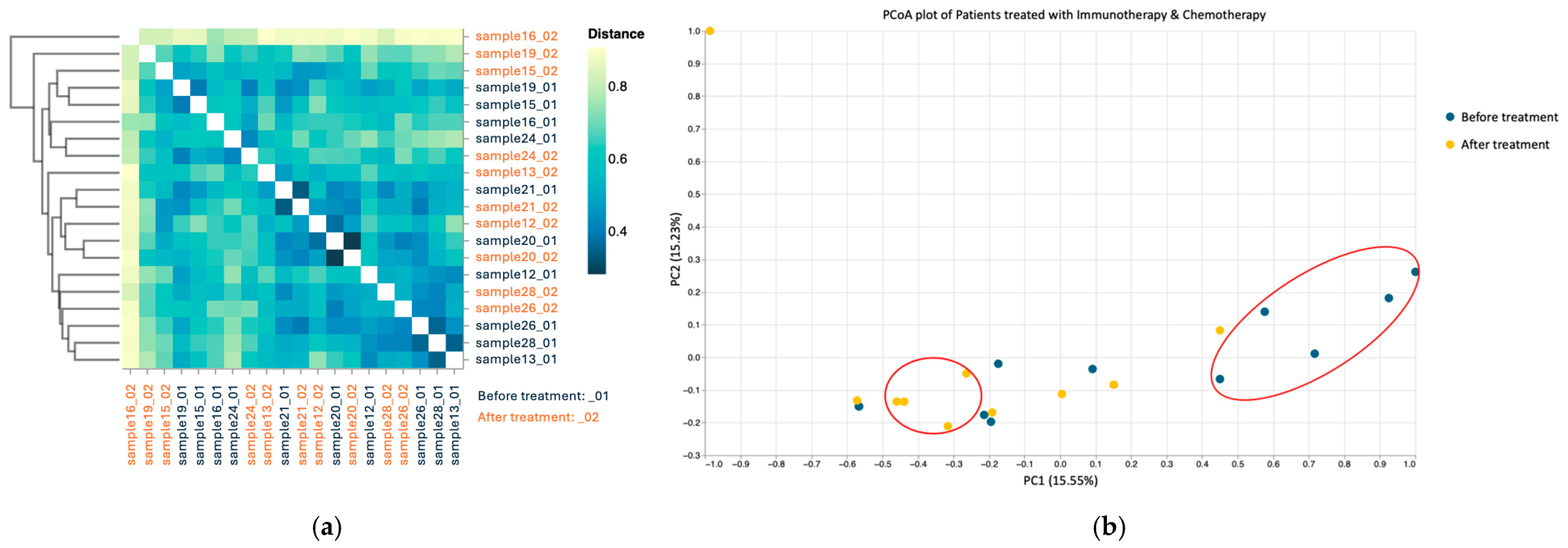
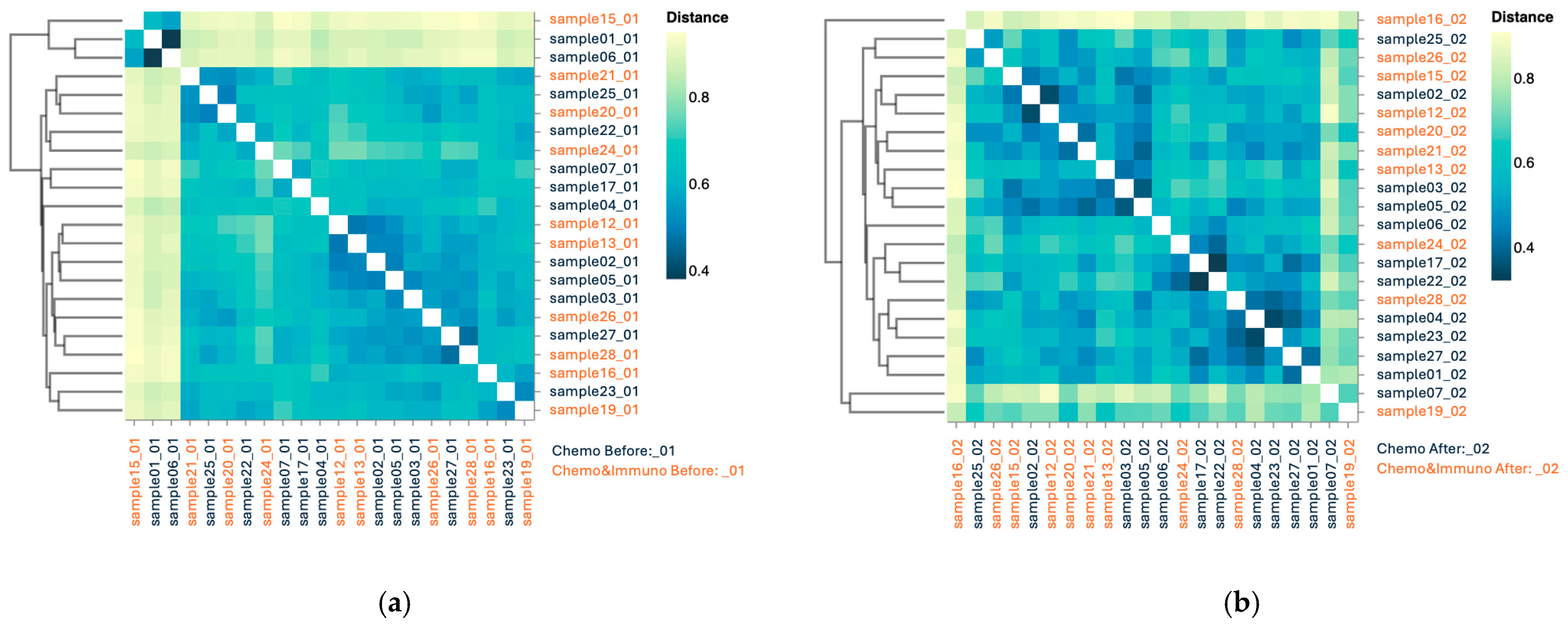
3.3.3. Beta Diversity
3.4. Microbiota Changes in Patients Treated with Combined Chemo–Immunotherapy
3.4.1. Taxonomic Composition
3.4.2. Alpha Diversity
3.4.3. Beta Diversity
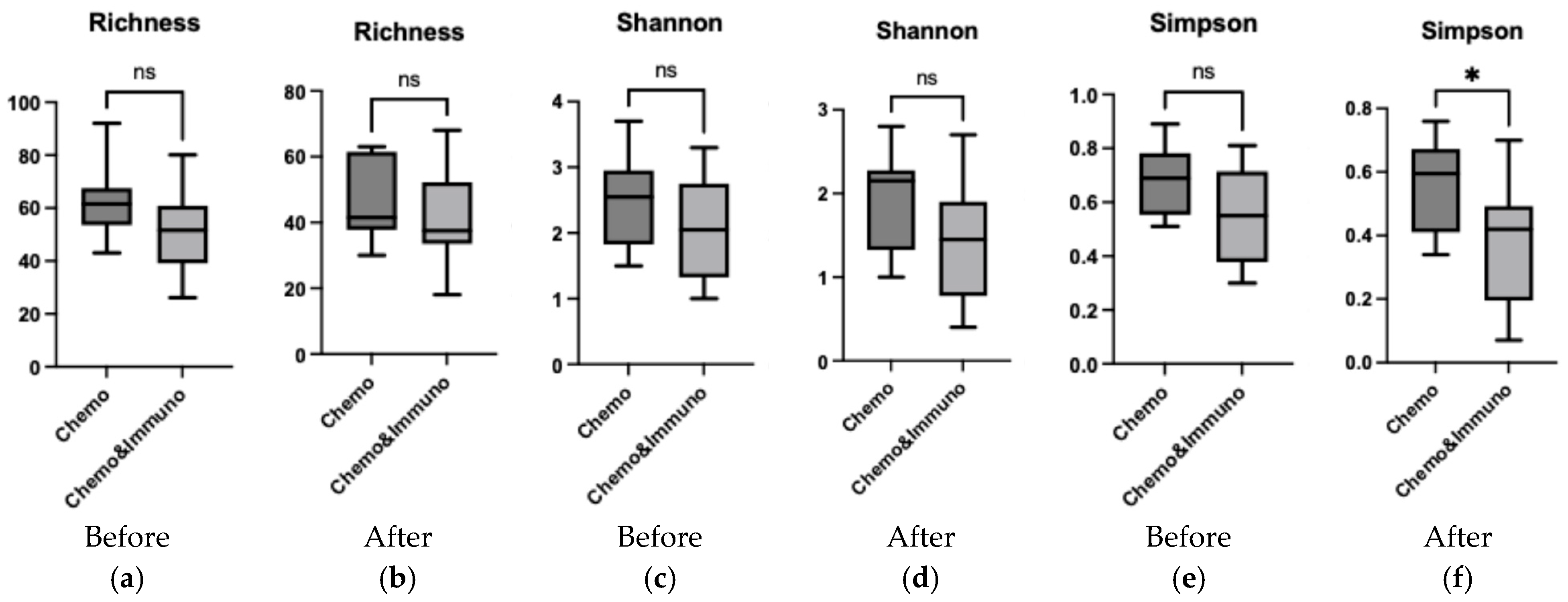
3.5. Comparison Between Group Treated with Chemotherapy and Group Treated with Combined Therapy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Wade, W.G. The oral microbiome in health and disease. Pharmacol. Res. 2013, 69, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.; Yu, W.H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef]
- Rajasekaran, J.J.; Krishnamurthy, H.K.; Bosco, J.; Jayaraman, V.; Krishna, K.; Wang, T.; Bei, K. Oral microbiome: A review of its impact on oral and systemic health. Microorganisms 2024, 12, 1797–1833. [Google Scholar] [CrossRef] [PubMed]
- Sedghi, L.; DiMassa, V.; Harrington, A.; Lynch, S.V.; Kapila, Y.L. The oral microbiome: Role of key organisms and complex networks in oral health and disease. Periodontology 2000 2021, 87, 107–131. [Google Scholar] [CrossRef]
- Maier, T. Oral microbiome in health and disease: Maintaining a healthy, balanced ecosystem and reversing dysbiosis. Microorganisms 2023, 11, 1453. [Google Scholar] [CrossRef]
- Santacroce, L.; Passarelli, P.C.; Azzolino, D.; Bottalico, L.; Charitos, I.A.; Cazzolla, A.P.; Colella, M.; Topi, S.; Godoy, F.G.; D’Addona, A. Oral microbiota in human health and disease: A perspective. Exp. Biol. Med. 2023, 248, 1288–1301. [Google Scholar] [CrossRef]
- Singhal, S.; Dian, D.; Keshavarzian, A.; Fogg, L.; Fields, J.Z.; Farhadi, A. The role of oral hygiene in inflammatory bowel disease. Dig. Dis. Sci. 2011, 56, 170–175. [Google Scholar] [CrossRef]
- Matsuoka, M.; Soria, S.A.; Pires, J.R.; Sant’Ana, A.C.P.; Freire, M. Natural and induced immune responses in oral cavity and saliva. BMC Immunol. 2025, 26, 34–61. [Google Scholar] [CrossRef]
- D’Agostino, S.; Valentini, G.; Dolci, M.; Ferrara, E. Potential relationship between poor oral hygiene and MRONJ: An observational retrospective study. Int. J. Environ. Res. Public Health 2023, 20, 5402. [Google Scholar] [CrossRef]
- Marsh, P.D.; Zaura, E. Dental biofilm: Ecological interactions in health and disease. J. Clin. Periodontol. 2017, 44, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Sheetal, A.; Hiremath, V.K.; Patil, A.G.; Sajjansetty, S.; Kumar, S.R. Malnutrition and its oral outcome–a review. J. Clin. Diagn. Res. JCDR 2013, 7, 178–180. [Google Scholar] [CrossRef]
- Genco, R.J.; Sanz, M. Clinical and public health implications of periodontal and systemic diseases: An overview. Periodontology 2000 2020, 83, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Diaz, P.I.; Hong, B.Y.; Frias-Lopez, J.; Dupuy, A.K.; Angeloni, M.; Abusleme, L.; Terzi, E.; Ioannidou, E.; Strausbaugh, L.D.; Dongari-Bagtzoglou, A. Transplantation-associated long-term immunosuppression promotes oral colonization by potentially opportunistic pathogens without impacting other members of the salivary bacteriome. Clin. Vaccine Immunol. 2013, 20, 920–930. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Helmink, B.A.; Spencer, C.N.; Reuben, A.; Wargo, J.A. The influence of the gut microbiome on cancer, immunity, and cancer immunotherapy. Cancer Cell 2018, 33, 570–580. [Google Scholar] [CrossRef]
- Sepich-Poore, G.D.; Zitvogel, L.; Straussman, R.; Hasty, J.; Wargo, J.; Knight, R. The microbiome and human cancer. Science 2021, 371, 4552–4587. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhao, X.; Sheng, T.; Peng, B.; Meng, S.; Liu, W.; Ge, L.; Wang, L.; Pathak, J.L.; Jiang, Q.; et al. Investigation of dental health and salivary microbiota characteristics of children with visual impairment in Guangzhou, China. BMC Oral Health 2025, 25, 408. [Google Scholar]
- Yamasaki, K.; Kawanami, T.; Yatera, K.; Fukuda, K.; Noguchi, S.; Nagata, S.; Nishida, C.; Kido, T.; Ishimoto, H.; Taniguchi, H.; et al. Significance of anaerobes and oral bacteria in community-acquired pneumonia. PLoS ONE 2013, 8, 63103–63111. [Google Scholar] [CrossRef]
- Rathbun, K.P.; Bourgault, A.M.; Sole, M.L. Oral microbes in hospital-acquired pneumonia: Practice and research implications. Crit. Care Nurse 2022, 42, 47–54. [Google Scholar] [CrossRef]
- Ren, Y.; Liang, J.; Li, X.; Deng, Y.; Cheng, S.; Wu, Q.; Song, W.; He, Y.; Zhu, J.; Zhang, X.; et al. Association between oral microbial dysbiosis and poor functional outcomes in stroke-associated pneumonia patients. BMC Microbiol. 2023, 23, 305–321. [Google Scholar] [CrossRef]
- Contaldo, M.; Itro, A.; Lajolo, C.; Gioco, G.; Inchingolo, F.; Serpico, R. Overview on osteoporosis, periodontitis and oral dysbiosis: The emerging role of oral microbiota. Appl. Sci. 2020, 17, 6000. [Google Scholar] [CrossRef]
- Stefura, T.; Zapała, B.; Gosiewski, T.; Skomarovska, O.; Dudek, A.; Pędziwiatr, M.; Major, P. Differences in compositions of oral and fecal microbiota between patients with obesity and controls. Medicina 2021, 57, 678. [Google Scholar] [CrossRef]
- Prince, Y.; Davison, G.M.; Davids, S.F.; Erasmus, R.T.; Kengne, A.P.; Graham, L.M.; Raghubeer, S.; Matsha, T.E. The relationship between the oral microbiota and metabolic syndrome. Biomedicines 2022, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Xie, G.; Chen, M.; He, Y.; Yu, W.; Chen, X.; Mao, W.; Liu, N.; Zhang, Y.; Chang, Q.; et al. Oral microbial dysbiosis in patients with periodontitis and chronic obstructive pulmonary disease. Front. Cell. Infect. Microbiol. 2023, 13, 1121399. [Google Scholar] [CrossRef] [PubMed]
- Garrett, W.S. Cancer and the microbiota. Science 2015, 348, 80–86. [Google Scholar] [CrossRef]
- Raza, M.H.; Gul, K.; Arshad, A.; Riaz, N.; Waheed, U.; Rauf, A.; Aldakheel, F.; Alduraywish, S.; Rehman, M.U.; Abdullah, M.; et al. Microbiota in cancer development and treatment. J. Cancer Res. Clin. Oncol. 2019, 145, 49–63. [Google Scholar] [CrossRef]
- Rajagopala, S.V.; Vashee, S.; Oldfield, L.M.; Suzuki, Y.; Venter, J.C.; Telenti, A.; Nelson, K.E. The human microbiome and cancer. Cancer Prev. Res. 2017, 10, 226–234. [Google Scholar] [CrossRef]
- Joyce, J.A.; Fearon, D.T. T cell exclusion, immune privilege, and the tumor microenvironment. Science 2015, 348, 74–80. [Google Scholar] [CrossRef]
- Perera, M.; Al-Hebshi, N.N.; Perera, I.; Ipe, D.; Ulett, G.C.; Speicher, D.J.; Chen, T.; Johnson, N.W. A dysbiotic mycobiome dominated by Candida albicans is identified within oral squamous-cell carcinomas. J. Oral Microbiol. 2017, 9, 1385369–1385378. [Google Scholar] [CrossRef]
- Sonis, S.T. The pathobiology of mucositis. Nat. Rev. Cancer 2004, 4, 277–284. [Google Scholar] [CrossRef]
- Hong, B.Y.; Sobue, T.; Choquette, L.; Dupuy, A.K.; Thompson, A.; Burleson, J.A.; Salner, A.L.; Schauer, P.K.; Joshi, P.; Fox, E.; et al. Chemotherapy-induced oral mucositis is associated with detrimental bacterial dysbiosis. Microbiome 2019, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Klymiuk, I.; Bilgilier, C.; Mahnert, A.; Prokesch, A.; Heininger, C.; Brandl, I.; Sahbegovic, H.; Singer, C.; Fuereder, T.; Steininger, C. Chemotherapy-associated oral microbiome changes in breast cancer patients. Front. Oncol. 2022, 12, 949071. [Google Scholar] [CrossRef]
- Zhou, P.; Manoil, D.; Belibasakis, G.N.; Kotsakis, G.A. Veillonellae: Beyond bridging species in oral biofilm ecology. Front. Oral Health. 2021, 2, 774115–774126. [Google Scholar] [CrossRef]
- Irfan, M.; Delgado, R.Z.; Frias-Lopez, J. The oral microbiome and cancer. Front. Immunol. 2020, 11, 591088–591106. [Google Scholar] [CrossRef] [PubMed]
- Cremonesi, E.; Governa, V.; Garzon, J.F.; Mele, V.; Amicarella, F.; Muraro, M.G.; Trella, E.; Galati-Fournier, V.; Oertli, D.; Däster, S.R.; et al. Gut microbiota modulate T cell trafficking into human colorectal cancer. Gut 2018, 67, 1984–1994. [Google Scholar] [CrossRef]
- Iida, N.; Dzutsev, A.; Stewart, C.A.; Smith, L.; Bouladoux, N.; Weingarten, R.A.; Molina, D.A.; Salcedo, R.; Back, T.; Cramer, S.; et al. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science 2013, 6161, 967–970. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhao, X.; Cui, L.; Huang, S. Metagenomic and metatranscriptomic insight into oral biofilms in periodontitis and related systemic diseases. Front. Microbiol. 2021, 12, 728585–728597. [Google Scholar] [CrossRef]
- Bars-Cortina, D.; Ramon, E.; Rius-Sansalvador, B.; Guinó, E.; Garcia-Serrano, A.; Mach, N.; Khannous-Lleiffe, O.; Saus, E.; Gabaldón, T.; Ibáñez-Sanz, G.; et al. Comparison between 16S rRNA and shotgun sequencing in colorectal cancer, advanced colorectal lesions, and healthy human gut microbiota. BMC Genom. 2024, 25, 730–750. [Google Scholar] [CrossRef]
- Augustine, D.; Sowmya, S.V.; Haragannavar, V.C.; Yousef, A.; Patil, S.; Gujjar, N.; Kashyap, S. Nanopore sequencing technology in oral oncology: A comprehensive insight. Insight 2022, 23, 268–275. [Google Scholar]
- Butler, I.; Turner, O.; Mohammed, K.; Akhtar, M.; Evans, D.; Lambourne, J.; Harris, K.; O’sullivan, D.M.; Sergaki, C. Standardization of 16S rRNA gene sequencing using nanopore long read sequencing technology for clinical diagnosis of culture negative infections. Front. Cell. Infect. Microbiol 2025, 15, 1517208–1517222. [Google Scholar] [CrossRef]
- Lee, A.W.T.; Chan, C.T.M.; Wong, L.L.Y.; Yip, C.Y.; Lui, W.T.; Cheng, K.C.; Leung, J.S.L.; Lee, L.K.; Wong, I.T.F.; Ng, T.T.L.; et al. Identification of microbial community in the urban environment: The concordance between conventional culture and nanopore 16S rRNA sequencing. Front. Microbiol. 2023, 14, 1164632–1164642. [Google Scholar] [CrossRef]
- Turco, L.; Della Monica, R.; Giordano, P.; Cuomo, M.; Biazzo, M.; Mateu, B.; Di Liello, R.; Daniele, B.; Normanno, N.; De Luca, A.; et al. Case report: Tracing in parallel the salivary and gut microbiota profiles to assist Larotrectinib anticancer treatment for NTRK fusion–positive glioblastoma. Front. Oncol. 2024, 14, 1458990–1458997. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Chen, Y.; Li, C.; Yang, S.; Lin, L.; Zhang, X.; Su, X.; Liu, L.; Zhao, H.; Luo, T.; et al. Variations in salivary microbiome and metabolites are associated with immunotherapy efficacy in patients with advanced NSCLC. mSystems 2025, 10, e01115-24. [Google Scholar] [CrossRef] [PubMed]
- Marotz, C.A.; Sanders, J.G.; Zuniga, C.; Zaramela, L.S.; Knight, R.; Zengler, K. Improving saliva shotgun metagenomics by chemical host DNA depletion. Microbiome 2018, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Charalampous, T.; Kay, G.L.; Richardson, H.; Aydin, A.; Baldan, R.; Jeanes, C.; Rae, D.; Grundy, S.; Turner, D.J.; Wain, J.; et al. Nanopore metagenomics enables rapid clinical diagnosis of bacterial lower respiratory infection. Nat. Biotechnol. 2019, 37, 783–792. [Google Scholar] [CrossRef]
- Matsuo, Y.; Komiya, S.; Yasumizu, Y.; Yasuoka, Y.; Mizushima, K.; Takagi, T.; Kryukov, K.; Fukuda, A.; Morimoto, Y.; Naito, Y.; et al. Full-length 16S rRNA gene amplicon analysis of human gut microbiota using MinION™ nanopore sequencing confers species-level resolution. BMC Microbiol. 2021, 21, 35. [Google Scholar] [CrossRef]
- Minot, S.S.; Krumm, N.; Greenfield, N.B. One codex: A sensitive and accurate data platform for genomic microbial identification. bioRxiv 2015, 027607. [Google Scholar] [CrossRef]
- Torres, P.J.; Fletcher, E.M.; Gibbons, S.M.; Bouvet, M.; Doran, K.S.; Kelley, S.T. Characterization of the salivary microbiome in patients with pancreatic cancer. PeerJ 2015, 3, 1373–1389. [Google Scholar] [CrossRef]
- Heshiki, Y.; Vazquez-Uribe, R.; Li, J.; Ni, Y.; Quainoo, S.; Imamovic, L.; Li, J.; Sørensen, M.; Chow, B.K.; Weiss, G.J.; et al. Predictable modulation of cancer treatment outcomes by the gut microbiota. Microbiome 2020, 8, 1–4. [Google Scholar] [CrossRef]
- Medeiros, M.C.; The, S.; Bellile, E.; Russo, N.; Schmitd, L.; Danella, E.; Singh, P.; Banerjee, R.; Bassis, C.; Murphy III, G.R.; et al. Salivary microbiome changes distinguish response to chemoradiotherapy in patients with oral cancer. Microbiome 2023, 11, 268. [Google Scholar] [CrossRef]
- Bourgeois, D.; Inquimbert, C.; Ottolenghi, L.; Carrouel, F. Periodontal pathogens as risk factors of cardiovascular diseases, diabetes, rheumatoid arthritis, cancer, and chronic obstructive pulmonary disease—Is there cause for consideration? Microorganisms 2019, 7, 424. [Google Scholar] [CrossRef] [PubMed]
- Otto-Dobos, L.D.; Grant, C.V.; Lahoud, A.A.; Wilcox, O.R.; Strehle, L.D.; Loman, B.R.; Yiadom, S.A.; Seng, M.M.; Halloy, N.R.; Russart, K.L.; et al. Chemotherapy-induced gut microbiome disruption, inflammation, and cognitive decline in female patients with breast cancer. Brain Behav. Immun. 2024, 120, 208–220. [Google Scholar] [CrossRef]
- Ludwig, H.; Hausmann, B.; Schreder, M.; Pönisch, W.; Zojer, N.; Knop, S.; Gunsilius, E.; Egle, A.; Petzer, A.; Einsele, H.; et al. Reduced alpha diversity of the oral microbiome correlates with short progression-free survival in patients with relapsed/refractory multiple myeloma treated with ixazomib-based therapy (AGMT MM 1, phase II trial). EJHaem 2021, 2, 99–103. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Álvarez-Mercado, A.I. The interplay between microbiota and chemotherapy-derived metabolites in breast cancer. Metabolites 2023, 13, 703–725. [Google Scholar] [CrossRef]
- Bloch, S.; Hager-Mair, F.F.; Andrukhov, O.; Schäffer, C. Oral streptococci: Modulators of health and disease. Front. Cell. Infect. Microbiol. 2024, 14, 1357631. [Google Scholar] [CrossRef]
- Matera, G.; Muto, V.; Vinci, M.; Zicca, E.; Abdollahi-Roodsaz, S.; van de Veerdonk, F.L.; Kullberg, B.J.; Liberto, M.C.; van der Meer, J.W.; Foca, A.; et al. Receptor recognition of and immune intracellular pathways for Veillonella parvula lipopolysaccharide. Clin. Vaccine Immunol. 2009, 16, 1804–1809. [Google Scholar] [CrossRef]
- Rai, A.K.; Panda, M.; Das, A.K.; Rahman, T.; Das, R.; Das, K.; Sarma, A.; Kataki, A.C.; Chattopadhyay, I. Dysbiosis of salivary microbiome and cytokines influence oral squamous cell carcinoma through inflammation. Arch. Microbiol. 2021, 203, 137–152. [Google Scholar] [CrossRef]
- Rosier, B.T.; Takahashi, N.; Zaura, E.; Krom, B.P.; MartÍnez-Espinosa, R.M.; van Breda, S.G.; Marsh, P.D.; Mira, A. The importance of nitrate reduction for oral health. J. Dent. Res. 2022, 101, 887–897. [Google Scholar] [CrossRef]
- Almståhl, A.; Finizia, C.; Carlén, A.; Fagerberg-Mohlin, B.; Alstad, T. Mucosal microflora in head and neck cancer patients. Int. J. Dent. Hyg. 2018, 16, 459–466. [Google Scholar] [CrossRef]
- Chai, X.; Liu, L.; Chen, F. Oral nitrate-reducing bacteria as potential probiotics for blood pressure homeostasis. Front. Cardiovasc. med. 2024, 11, 1337281–1337287. [Google Scholar] [CrossRef]
- Miyoshi, T.; Oge, S.; Nakata, S.; Ueno, Y.; Ukita, H.; Kousaka, R.; Miura, Y.; Yoshinari, N.; Yoshida, A. Gemella haemolysans inhibits the growth of the periodontal pathogen Porphyromonas gingivalis. Sci. Rep. 2021, 11, 11742–11754. [Google Scholar] [CrossRef]
- Havel, J.J.; Chowell, D.; Chan, T.A. The evolving landscape of biomarkers for checkpoint inhibitor immunotherapy. Nat. Rev. Cancer 2019, 19, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Han, J.; Peng, X.; Zhou, X.; Gong, T.; Zheng, X. The role of Fusobacterium nucleatum in cancer and its implications for clinical applications. Mol. Oral Microbiol. 2024, 39, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Spilker, M.E.; Goetz, L.; Peterson, S.N.; Siuzdak, G. Metabolite and microbiome interplay in cancer immunotherapy. Cancer Res. 2016, 76, 6146–6152. [Google Scholar] [CrossRef] [PubMed]
- Matson, V.; Fessler, J.; Bao, R.; Chongsuwat, T.; Zha, Y.; Alegre, M.L.; Luke, J.J.; Gajewski, T.F. The commensal microbiome is associated with anti–PD-1 efficacy in metastatic melanoma patients. Science 2018, 359, 104–108. [Google Scholar] [CrossRef]
- Lu, M.; Xuan, S.; Wang, Z. Oral microbiota: A new view of body health. Food Sci. Hum. Wellness 2019, 8, 8–15. [Google Scholar] [CrossRef]
- Yi, M.; Jiao, D.; Qin, S.; Chu, Q.; Li, A.; Wu, K. Manipulating gut microbiota composition to enhance the therapeutic effect of cancer immunotherapy. Integr. Cancer Ther. 2019, 18, 1534735419876351–1534735419876365. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All n = 22 | Chemotherapy n = 12 | Chemotherapy and Immunotherapy n = 10 | p | |
|---|---|---|---|---|
| Age ± SD | 62.86 ± 9.57 | 64.33 ± 7.28 | 61.1 ± 11.95 | 0.467 * |
| Male/Female | 9/13 | 3/9 | 6/4 | 0.097 ** |
| Hypertension | 13 (59.09) | 7 (58.33) | 6 (60) | 0.936 ** |
| Diabetes mellitus | 6 (27.27) | 3 (25) | 3 (30) | 0.795 ** |
| Thyroid affection | 3 (13.64) | 2 (16.64) | 1 (10) | 0.653 ** |
| Smoking | 6 (27.27) | 3 (25) | 3 (30) | 0.795 ** |
| Allergies | 3 (13.64) | 1 (8.33) | 2 (20) | 0.430 ** |
| Cancer in family history | 7 (31.82) | 4 (33.33) | 3 (30) | 0.865 ** |
| Cancer relapse | 6 (27.27) | 3 (25) | 3 (30) | 0.795 ** |
| Removable dental prosthesis | 10 (45.45) | 5 (41.67) | 5 (50) | 0.697 ** |
| Oral thrush | 9 (40.91) | 7 (58.33) | 2 (20) | 0.069 ** |
| Mucositis | 5 (22.73) | 2 (16.64) | 3 (30) | 0.459 ** |
| Gingivitis | 8 (36.36) | 3 (25) | 5(50) | 0.023 ** |
| Cancer Type | ||||
| Breast cancer | 8 (36.36) | 6 (50) | 2 (20) | 0.144 ** |
| Liposarcoma | 1 (4.54) | 1 (8.33) | 0 (0) | 0.352 ** |
| Stomach cancer | 1 (4.54) | 1 (8.33) | 0 (0) | 0.352 ** |
| Melanoma | 2 (9.09) | 0 (0) | 2 (20) | 0.105 ** |
| Lung cancer | 5 (22.73) | 2 (16.64) | 3 (30) | 0.459 ** |
| Cervical cancer | 2 (9.09) | 0 (0) | 2 (20) | 0.105 ** |
| Gallbladder cancer | 1 (4.54) | 1 (8.33) | 0 (0) | 0.352 ** |
| Rectal cancer | 2 (9.09) | 1 (8.33) | 1 (10) | 0.889 ** |
| QC | Seq 1 | Seq 2 |
|---|---|---|
| Reads Analyzed | 1,388,589 | 2,199,826 |
| Total Yield | 2.1 Gbases | 3.4 Gbases |
| Avg Quality Score | 10.19 | 10.04 |
| Avg Sequence Length | 1542 | 1539 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Padure, A.; Talpos-Niculescu, I.C.; Ciordas, P.D.; Romanescu, M.; Chis, A.R.; Rusu, L.-C.; Sirbu, I.O. Impact of Chemotherapy Alone and in Combination with Immunotherapy on Oral Microbiota in Cancer Patients—A Pilot Study. Microorganisms 2025, 13, 1565. https://doi.org/10.3390/microorganisms13071565
Padure A, Talpos-Niculescu IC, Ciordas PD, Romanescu M, Chis AR, Rusu L-C, Sirbu IO. Impact of Chemotherapy Alone and in Combination with Immunotherapy on Oral Microbiota in Cancer Patients—A Pilot Study. Microorganisms. 2025; 13(7):1565. https://doi.org/10.3390/microorganisms13071565
Chicago/Turabian StylePadure, Adriana, Ioana Cristina Talpos-Niculescu, Paula Diana Ciordas, Mirabela Romanescu, Aimee Rodica Chis, Laura-Cristina Rusu, and Ioan Ovidiu Sirbu. 2025. "Impact of Chemotherapy Alone and in Combination with Immunotherapy on Oral Microbiota in Cancer Patients—A Pilot Study" Microorganisms 13, no. 7: 1565. https://doi.org/10.3390/microorganisms13071565
APA StylePadure, A., Talpos-Niculescu, I. C., Ciordas, P. D., Romanescu, M., Chis, A. R., Rusu, L.-C., & Sirbu, I. O. (2025). Impact of Chemotherapy Alone and in Combination with Immunotherapy on Oral Microbiota in Cancer Patients—A Pilot Study. Microorganisms, 13(7), 1565. https://doi.org/10.3390/microorganisms13071565