
The Impact of Tank Disinfectants on the Development of Microbiota in Gilthead Seabream (Sparus aurata) Larviculture Systems

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Setup and Sample Collection
2.2. DNA Extraction, and 16S rRNA Gene Amplification
2.3. Libraries Preparation and Sequencing
2.3.1. Illumina MiSeq Sequencing of 16S rRNA Amplicons (V3–V4 Region)
2.3.2. ONT MinION Sequencing of 16S rRNA Amplicons (V1–V9 Region)
2.4. Bioinformatic Data Processing
2.4.1. Illumina Sequencing Data
2.4.2. Nanopore Sequencing Data
2.5. Bacterial Composition and Diversity Analysis
3. Results
3.1. Dataset Overview
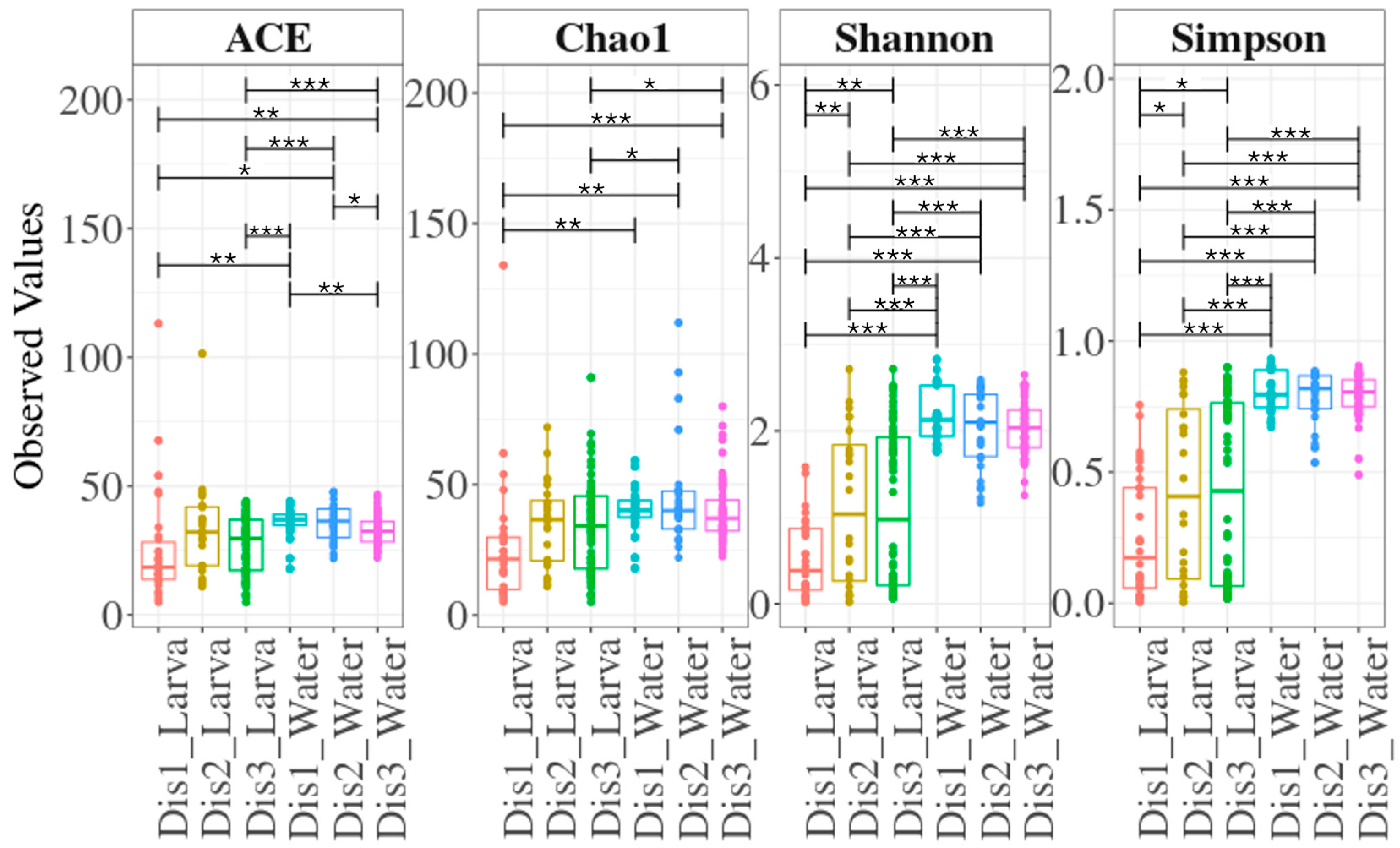
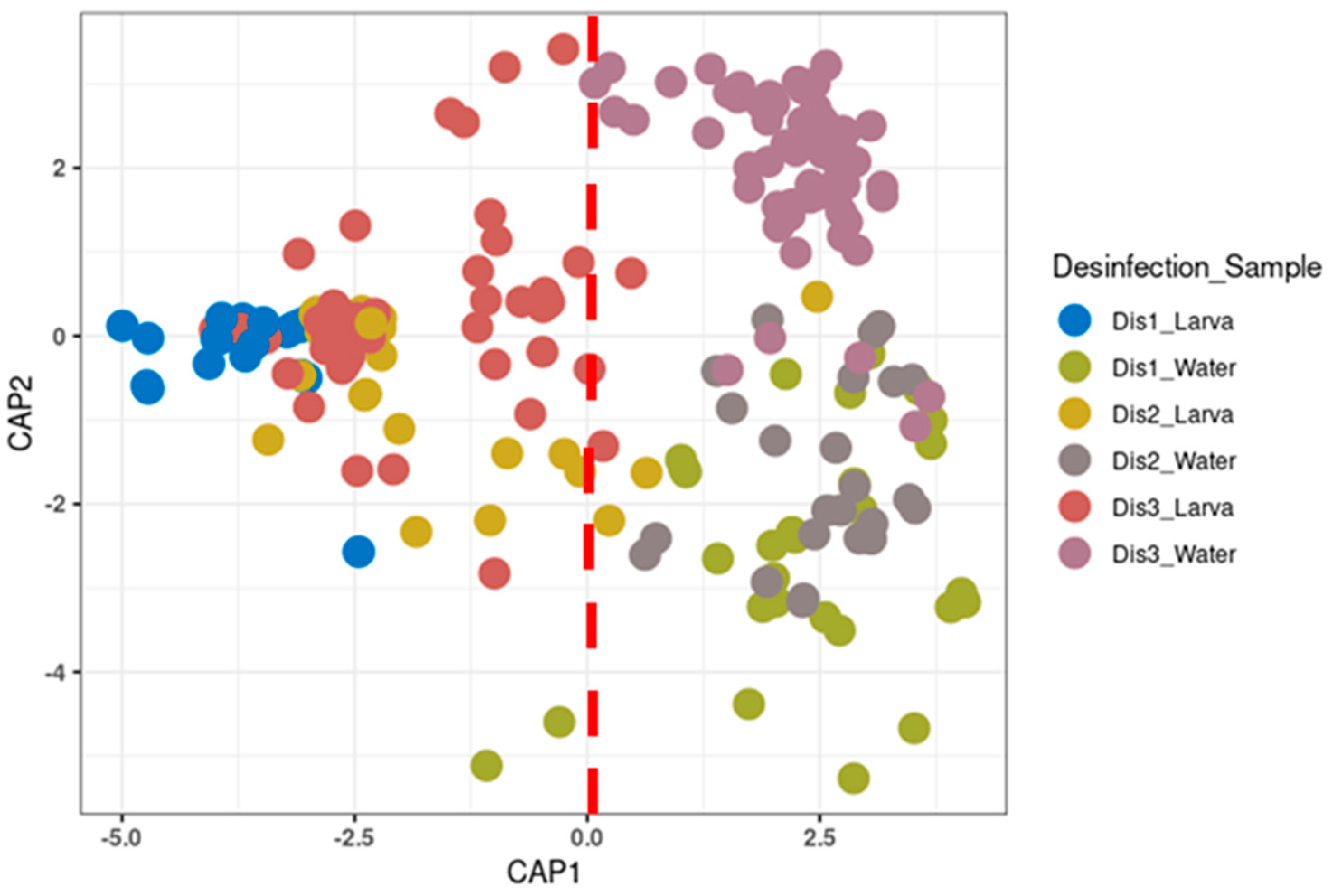
3.2. The Diversity of the Bacterial Communities Based on the Disinfection Method
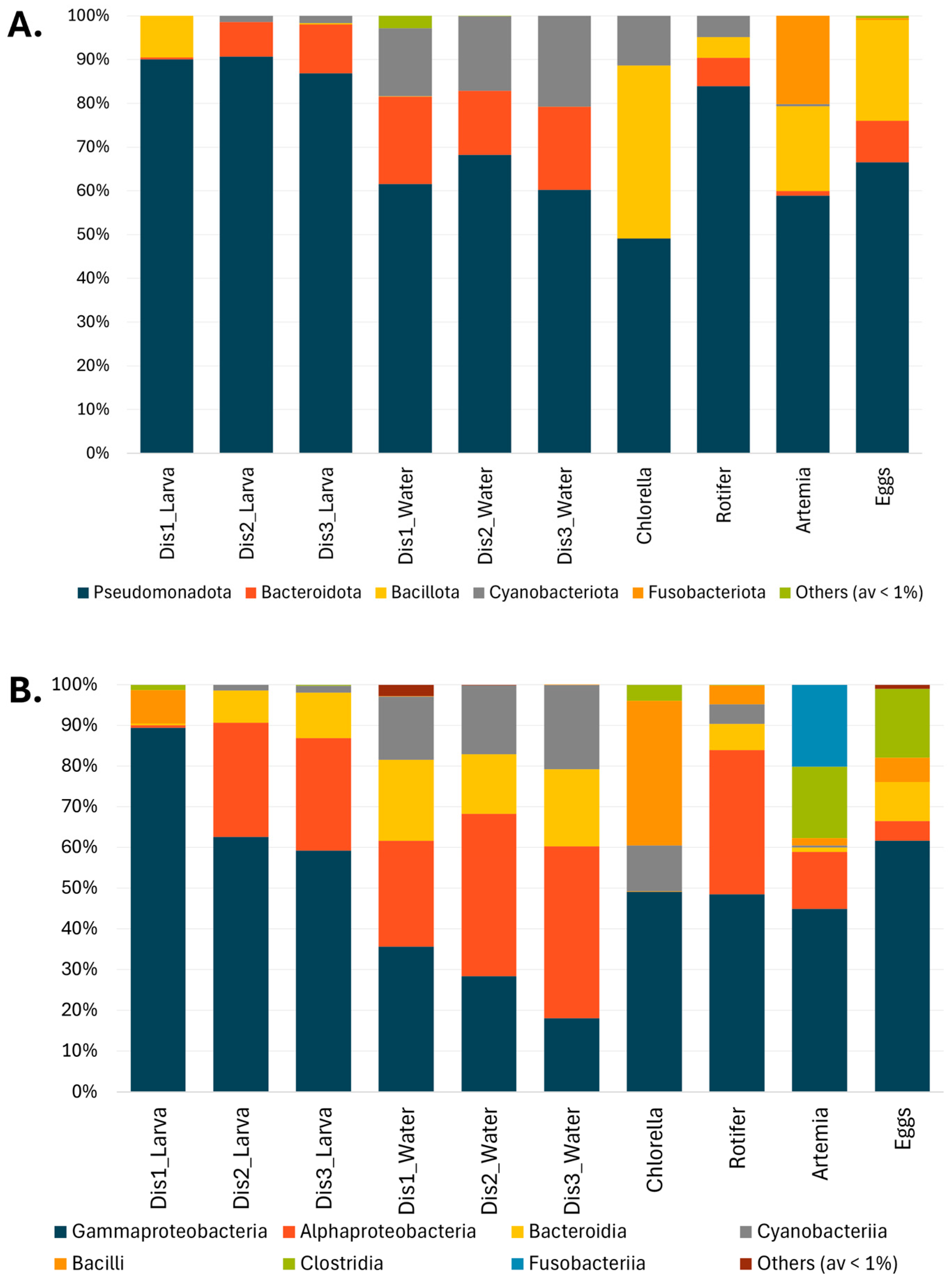
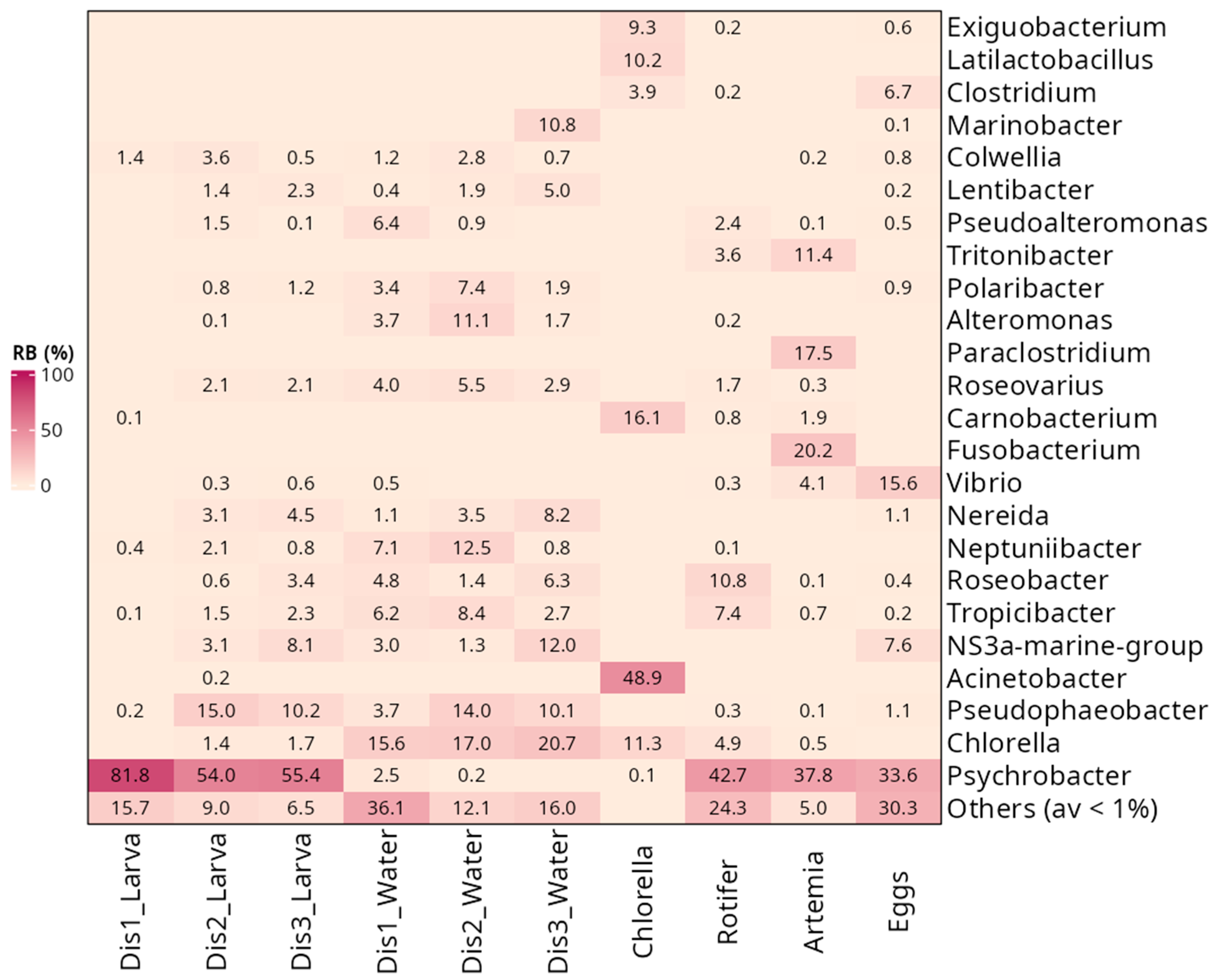
3.3. Taxonomic Composition of Bacterial Communities Based on the Type of Disinfection
4. Discussion
4.1. Type of Disinfection and Microbial Communities
4.2. Factors Influencing the Composition of the Larval Microbiota
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hellenic Aquaculture Producers Organization. HAPO Aquaculture in Greece, Annual Report 2024; Hellenic Aquaculture Producers Organization: Attica, Greece, 2024. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2022; FAO: Rome, Italy, 2022; ISBN 978-92-5-136364-5. [Google Scholar]
- Freitas, J.; Vaz-Pires, P.; Câmara, J.S. From Aquaculture Production to Consumption: Freshness, Safety, Traceability and Authentication, the Four Pillars of Quality. Aquaculture 2020, 518, 734857. [Google Scholar] [CrossRef]
- Ottinger, M.; Clauss, K.; Kuenzer, C. Aquaculture: Relevance, Distribution, Impacts and Spatial Assessments—A Review. Ocean Coast. Manag. 2016, 119, 244–266. [Google Scholar] [CrossRef]
- Araujo, G.S.; da Silva, J.W.A.; Cotas, J.; Pereira, L. Fish Farming Techniques: Current Situation and Trends. J. Mar. Sci. Eng. 2022, 10, 1598. [Google Scholar] [CrossRef]
- Gopakumar, G.; Madhu, K.; Jayasankar, R.; Madhu, R.; Kizhakudan, J.K.; Josileen, J.; Ignatius, B.; Vijayagopal, P.; Joseph, S.; Mohamed, G.; et al. Live Feed Research for Larviculture of Marine Finfish and Shellfish. Mar. Fish. Inf. Serv. T E Ser. 2008, 197, 1–6. [Google Scholar]
- Shields, R.J. Larviculture of Marine Finfish in Europe. Aquaculture 2001, 200, 55–88. [Google Scholar] [CrossRef]
- Muniesa, A.; Basurco, B.; Aguilera, C.; Furones, D.; Reverté, C.; Sanjuan-Vilaplana, A.; Jansen, M.D.; Brun, E.; Tavornpanich, S. Mapping the Knowledge of the Main Diseases Affecting Sea Bass and Sea Bream in Mediterranean. Transbound. Emerg. Dis. 2020, 67, 1089–1100. [Google Scholar] [CrossRef]
- Subasinghe, R.; Bernoth, E.-M. Disease Control and Health Management: Aquaculture Development, Health and Wealth; FAO: Rome, Italy, 2000. [Google Scholar]
- Borges, N.; Keller-Costa, T.; Sanches-Fernandes, G.M.M.; Louvado, A.; Gomes, N.C.M.; Costa, R. Bacteriome Structure, Function, and Probiotics in Fish Larviculture: The Good, the Bad, and the Gaps. Annu. Rev. Anim. Biosci. 2021, 9, 423–452. [Google Scholar] [CrossRef]
- Savaş, S.; Kubilay, A.; Basmaz, N. Effect of Bacterial Load in Feeds on Intestinal Microflora of Seabream (Sparus aurata) Larvae and Juveniles. Isr. J. Aquac.—Bamidgeh 2005, 57, 3–9. [Google Scholar] [CrossRef]
- Vadstein, O.; Attramadal, K.J.K.; Bakke, I.; Forberg, T.; Olsen, Y.; Verdegem, M.; Giatsis, C.; Skjermo, J.; Aasen, I.M.; Gatesoupe, F.-J.; et al. Managing the Microbial Community of Marine Fish Larvae: A Holistic Perspective for Larviculture. Front. Microbiol. 2018, 9, 1820. [Google Scholar] [CrossRef]
- Banerjee, G.; Ray, A.K. Bacterial Symbiosis in the Fish Gut and Its Role in Health and Metabolism. Symbiosis 2017, 72, 1–11. [Google Scholar] [CrossRef]
- Luan, Y.; Li, M.; Zhou, W.; Yao, Y.; Yang, Y.; Zhang, Z.; Ringø, E.; Erik Olsen, R.; Liu Clarke, J.; Xie, S.; et al. The Fish Microbiota: Research Progress and Potential Applications. Engineering 2023, 29, 137–146. [Google Scholar] [CrossRef]
- Sehnal, L.; Brammer-Robbins, E.; Wormington, A.M.; Blaha, L.; Bisesi, J.; Larkin, I.; Martyniuk, C.J.; Simonin, M.; Adamovsky, O. Microbiome Composition and Function in Aquatic Vertebrates: Small Organisms Making Big Impacts on Aquatic Animal Health. Front. Microbiol. 2021, 12, 567408. [Google Scholar] [CrossRef] [PubMed]
- Brugman, S.; Ikeda-Ohtsubo, W.; Braber, S.; Folkerts, G.; Pieterse, C.M.; Bakker, P.A. A Comparative Review on Microbiota Manipulation: Lessons from Fish, Plants, Livestock, and Human Research. Front. Nutr. 2018, 5, 80. [Google Scholar] [CrossRef] [PubMed]
- Wynne, J.W.; Thakur, K.K.; Slinger, J.; Samsing, F.; Milligan, B.; Powell, J.F.F.; McKinnon, A.; Nekouei, O.; New, D.; Richmond, Z.; et al. Microbiome Profiling Reveals a Microbial Dysbiosis During a Natural Outbreak of Tenacibaculosis (Yellow Mouth) in Atlantic Salmon. Front. Microbiol. 2020, 11, 586387. [Google Scholar] [CrossRef] [PubMed]
- Assefa, A.; Abunna, F. Maintenance of Fish Health in Aquaculture: Review of Epidemiological Approaches for Prevention and Control of Infectious Disease of Fish. Vet. Med. Int. 2018, 2018, 5432497. [Google Scholar] [CrossRef]
- American Fisheries Society. Guide to Using Drugs, Biologics & Other Chemicals in Aquaculture; American Fisheries Society—Fish Culture Section: Bethesda, MD, USA, 2019. [Google Scholar]
- Russell Danner, G.; Merrill, P. Disinfectants, Disinfection, and Biosecurity in Aquaculture. In Aquaculture Biosecurity; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2005; pp. 91–128. ISBN 978-0-470-37685-0. [Google Scholar]
- Eleraky, N.Z.; Potgieter, L.N.D.; Kennedy, M.A. Virucidal Efficacy of Four New Disinfectants. J. Am. Anim. Hosp. Assoc. 2002, 38, 231–234. [Google Scholar] [CrossRef]
- Kelkar, V.P.; Rolsky, C.B.; Pant, A.; Green, M.D.; Tongay, S.; Halden, R.U. Chemical and Physical Changes of Microplastics during Sterilization by Chlorination. Water Res. 2019, 163, 114871. [Google Scholar] [CrossRef]
- Sanawar, H.; Xiong, Y.; Alam, A.; Croué, J.-P.; Hong, P.-Y. Chlorination or Monochloramination: Balancing the Regulated Trihalomethane Formation and Microbial Inactivation in Marine Aquaculture Waters. Aquaculture 2017, 480, 94–102. [Google Scholar] [CrossRef]
- Duan, Y.; Tang, Y.; Huang, J.; Zhang, J.; Lin, H.; Jiang, S.; Wang, R.; Wang, G. Changes in the Microbial Community of Litopenaeus vannamei Larvae and Rearing Water during Different Growth Stages after Disinfection Treatment of Hatchery Water. J. Microbiol. 2020, 58, 741–749. [Google Scholar] [CrossRef]
- Gallard, H.; von Gunten, U. Chlorination of Natural Organic Matter: Kinetics of Chlorination and of THM Formation. Water Res. 2002, 36, 65–74. [Google Scholar] [CrossRef]
- Maapea, A.D.; Vine, N.G.; Macey, B.M. Bacterial Microbiome of Dusky Kob Argyrosomus japonicus Eggs and Rearing Water and the Bacteriostatic Effect of Selected Disinfectants. Aquaculture 2021, 542, 736882. [Google Scholar] [CrossRef]
- Dawson, V.K.; Meinertz, J.R.; Schmidt, L.J.; Gingerich, W.H. A Simple Analytical Procedure to Replace HPLC for Monitoring Treatment Concentrations of Chloramine-T on Fish Culture Facilities. Aquaculture 2003, 217, 61–72. [Google Scholar] [CrossRef]
- Arvin, E.; Pedersen, L.-F. Hydrogen Peroxide Decomposition Kinetics in Aquaculture Water. Aquac. Eng. 2015, 64, 1–7. [Google Scholar] [CrossRef]
- Avendaño-Herrera, R.; Magariños, B.; Irgang, R.; Toranzo, A.E. Use of Hydrogen Peroxide against the Fish Pathogen Tenacibaculum maritimum and Its Effect on Infected Turbot (Scophthalmus maximus). Aquaculture 2006, 257, 104–110. [Google Scholar] [CrossRef]
- Bögner, D.; Bögner, M.; Schmachtl, F.; Bill, N.; Halfer, J.; Slater, M.J. Hydrogen Peroxide Oxygenation and Disinfection Capacity in Recirculating Aquaculture Systems. Aquac. Eng. 2021, 92, 102140. [Google Scholar] [CrossRef]
- Giménez-Papiol, G.; Padrós, F.; Roque, A.; Estévez, A.; Furones, D. Effects of a Peroxide-Based Commercial Product on Bacterial Load of Larval Rearing Water and on Larval Survival of Two Species of Sparidae under Intensive Culture: Preliminary Study. Aquac. Res. 2009, 40, 504–508. [Google Scholar] [CrossRef]
- Vallés, R.; Roque, A.; Caballero, A.; Estévez, A. Use of Ox-Aquaculture© for Disinfection of Live Prey and Meagre Larvae, Argyrosomus regius (Asso, 1801). Aquac. Res. 2015, 46, 413–419. [Google Scholar] [CrossRef]
- Rach, J.J.; Gaikowski, M.P.; Ramsay, R.T. Efficacy of Hydrogen Peroxide to Control Parasitic Infestations on Hatchery-Reared Fish. J. Aquat. Anim. Health 2000, 12, 267–273. [Google Scholar] [CrossRef]
- Kitis, M. Disinfection of Wastewater with Peracetic Acid: A Review. Environ. Int. 2004, 30, 47–55. [Google Scholar] [CrossRef]
- Suurnäkki, S.; Pulkkinen, J.T.; Lindholm-Lehto, P.C.; Tiirola, M.; Aalto, S.L. The Effect of Peracetic Acid on Microbial Community, Water Quality, Nitrification and Rainbow Trout (Oncorhynchus mykiss) Performance in Recirculating Aquaculture Systems. Aquaculture 2020, 516, 734534. [Google Scholar] [CrossRef]
- Francis-Floyd, R.; Klinger, R. Use of Potassium Permanganate to Control External Infections of Ornamental Fish; University of Florida: Florida, FL, USA, 2002. [Google Scholar]
- Attramadal, K.J.K.; Øien, J.V.; Kristensen, E.; Evjemo, J.O.; Kjørsvik, E.; Vadstein, O.; Bakke, I. UV Treatment in RAS Influences the Rearing Water Microbiota and Reduces the Survival of European Lobster Larvae (Homarus gammarus). Aquac. Eng. 2021, 94, 102176. [Google Scholar] [CrossRef]
- Attramadal, K.J.K.; Øie, G.; Størseth, T.R.; Alver, M.O.; Vadstein, O.; Olsen, Y. The Effects of Moderate Ozonation or High Intensity UV-Irradiation on the Microbial Environment in RAS for Marine Larvae. Aquaculture 2012, 330–333, 121–129. [Google Scholar] [CrossRef]
- Llewellyn, S.; Inpankaew, T.; Nery, S.V.; Gray, D.J.; Verweij, J.J.; Clements, A.C.A.; Gomes, S.J.; Traub, R.; McCarthy, J.S. Application of a Multiplex Quantitative PCR to Assess Prevalence and Intensity of Intestinal Parasite Infections in a Controlled Clinical Trial. PLoS Neglected Trop. Dis. 2016, 10, e0004380. [Google Scholar] [CrossRef] [PubMed]
- World Organization for Animal Health. Aquatic Code. Available online: https://www.woah.org/en/what-we-do/standards/codes-and-manuals/aquatic-code-online-access/ (accessed on 15 May 2024).
- Bel Mokhtar, N.; Apostolopoulou, G.; Koumoundouros, G.; Tzokas, K.; Toskas, K.; Gourzioti, E.; Stathopoulou, P.; Tsiamis, G. Bacterial Community Structures and Dynamics Associated with Rotated Positioning Syndrome in Gilthead Sea Bream (Sparus aurata) Larviculture. Front. Aquac. 2024, 2, 1270932. [Google Scholar] [CrossRef]
- Califano, G.; Castanho, S.; Soares, F.; Ribeiro, L.; Cox, C.J.; Mata, L.; Costa, R. Molecular Taxonomic Profiling of Bacterial Communities in a Gilthead Seabream (Sparus aurata) Hatchery. Front. Microbiol. 2017, 8, 204. [Google Scholar] [CrossRef]
- Nikouli, E.; Meziti, A.; Antonopoulou, E.; Mente, E.; Kormas, K.A. Host-Associated Bacterial Succession during the Early Embryonic Stages and First Feeding in Farmed Gilthead Sea Bream (Sparus aurata). Genes 2019, 10, 483. [Google Scholar] [CrossRef]
- Al-Ashhab, A.; Alexander-Shani, R.; Avrahami, Y.; Ehrlich, R.; Strem, R.I.; Meshner, S.; Shental, N.; Sharon, G. Sparus aurata and Lates calcarifer Skin Microbiota under Healthy and Diseased Conditions in UV and Non-UV Treated Water. Anim. Microbiome 2022, 4, 42. [Google Scholar] [CrossRef]
- Moretti, A.; Pedini Fernandez-Criado, M.; Vetillart, R. Manual on Hatchery Production of Seabass and Gilthead Seabream; FAO: Rome, Italy, 2005; Volume 2. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Hartley, J.; Bowen, H. PEG Precipitation for Selective Removal of Small DNA Fragments. Focus 1996, 18, 27. [Google Scholar]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.C. UNCROSS2: Identification of Cross-Talk in 16S rRNA OTU Tables. bioRxiv 2018, 400762. [Google Scholar]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A Comprehensive Online Resource for Quality Checked and Aligned Ribosomal RNA Sequence Data Compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing Large Minimum Evolution Trees with Profiles Instead of a Distance Matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Completing bacterial genome assemblies with multiplex MinION sequencing. Microb. Genom. 2017, 3, e000132. [Google Scholar] [CrossRef]
- De Coster, W.; D’Hert, S.; Schultz, D.T.; Cruts, M.; Van Broeckhoven, C. NanoPack: Visualizing and Processing Long-Read Sequencing Data. Bioinformatics 2018, 34, 2666–2669. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, H.; Ciuffreda, L.; Flores, C. NanoCLUST: A Species-Level Analysis of 16S rRNA Nanopore Sequencing Data. Bioinformatics 2021, 37, 1600–1601. [Google Scholar] [CrossRef]
- McInnes, L.; Healy, J.; Melville, J. UMAP: Uniform Manifold Approximation and Projection for Dimension Reduction. arXiv 2018, arXiv:1802.03426. [Google Scholar]
- McInnes, L.; Healy, J.; Astels, S. Hdbscan: Hierarchical Density Based Clustering. J. Open Source Softw. 2017, 2, 205. [Google Scholar] [CrossRef]
- Vaser, R.; Sovic, I.; Nagarajan, N.; Sikic, M. Fast and Accurate de Novo Genome Assembly from Long Uncorrected Reads. Genome Res. 2017, 27, 737–746. [Google Scholar] [CrossRef]
- Medaka, Version 2.1.0. Sequence Correction Provided by ONT Research. Oxford Nanopore Technologies Ltd.: Oxford, UK, 2020. Available online: https://github.com/nanoporetech/medaka (accessed on 1 February 2025).
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 February 2025).
- Chen, J.; Zhang, X.; Yang, L.; Zhang, L. GUniFrac: Generalized UniFrac Distances, Distance-Based Multivariate Methods and Feature-Based Univariate Methods for Microbiome Data Analysis. 2023. Available online: https://CRAN.R-project.org/package=GUniFrac (accessed on 1 February 2025).
- Bel Mokhtar, N.; Asimakis, E.; Galiatsatos, I.; Maurady, A.; Stathopoulou, P.; Tsiamis, G. Development of MetaXplore: An Interactive Tool for Targeted Metagenomic Analysis. Curr. Issues Mol. Biol. 2024, 46, 4803–4814. [Google Scholar] [CrossRef] [PubMed]
- de Bruijn, I.; Liu, Y.; Wiegertjes, G.F.; Raaijmakers, J.M. Exploring Fish Microbial Communities to Mitigate Emerging Diseases in Aquaculture. FEMS Microbiol. Ecol. 2018, 94, fix161. [Google Scholar] [CrossRef] [PubMed]
- Perry, W.B.; Lindsay, E.; Payne, C.J.; Brodie, C.; Kazlauskaite, R. The Role of the Gut Microbiome in Sustainable Teleost Aquaculture. Proc. R. Soc. B Biol. Sci. 2020, 287, 20200184. [Google Scholar] [CrossRef]
- Raulo, A.; Ruokolainen, L.; Lane, A.; Amato, K.; Knight, R.; Leigh, S.; Stumpf, R.; White, B.; Nelson, K.E.; Baden, A.L.; et al. Social Behaviour and Gut Microbiota in Red-Bellied Lemurs (Eulemur rubriventer): In Search of the Role of Immunity in the Evolution of Sociality. J. Anim. Ecol. 2018, 87, 388–399. [Google Scholar] [CrossRef]
- Karlsen, C.; Tzimorotas, D.; Robertsen, E.M.; Kirste, K.H.; Bogevik, A.S.; Rud, I. Feed Microbiome: Confounding Factor Affecting Fish Gut Microbiome Studies. ISME Commun. 2022, 2, 14. [Google Scholar] [CrossRef]
- Krotman, Y.; Yergaliyev, T.M.; Alexander Shani, R.; Avrahami, Y.; Szitenberg, A. Dissecting the Factors Shaping Fish Skin Microbiomes in a Heterogeneous Inland Water System. Microbiome 2020, 8, 9. [Google Scholar] [CrossRef]
- Nikouli, E.; Meziti, A.; Smeti, E.; Antonopoulou, E.; Mente, E.; Kormas, K. Gut Microbiota of Five Sympatrically Farmed Marine Fish Species in the Aegean Sea. Microb. Ecol. 2021, 81, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Quero, G.M.; Piredda, R.; Basili, M.; Maricchiolo, G.; Mirto, S.; Manini, E.; Seyfarth, A.M.; Candela, M.; Luna, G.M. Host-Associated and Environmental Microbiomes in an Open-Sea Mediterranean Gilthead Sea Bream Fish Farm. Microb. Ecol. 2022, 86, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Rabelo-Ruiz, M.; Newman-Portela, A.M.; Peralta-Sánchez, J.M.; Martín-Platero, A.M.; del Mar Agraso, M.; Bermúdez, L.; Aguinaga, M.A.; Baños, A.; Maqueda, M.; Valdivia, E.; et al. Beneficial Shifts in the Gut Bacterial Community of Gilthead Seabream (Sparus aurata) Juveniles Supplemented with Allium-Derived Compound Propyl Propane Thiosulfonate (PTSO). Animals 2022, 12, 1821. [Google Scholar] [CrossRef]
- Roquigny, R.; Mougin, J.; Le Bris, C.; Bonnin-Jusserand, M.; Doyen, P.; Grard, T. Characterization of the Marine Aquaculture Microbiome: A Seasonal Survey in a Seabass Farm. Aquaculture 2021, 531, 735987. [Google Scholar] [CrossRef]
- Wang, A.R.; Ran, C.; Ringø, E.; Zhou, Z.G. Progress in Fish Gastrointestinal Microbiota Research. Rev. Aquac. 2018, 10, 626–640. [Google Scholar] [CrossRef]
- Zeng, A.; Tan, K.; Gong, P.; Lei, P.; Guo, Z.; Wang, S.; Gao, S.; Zhou, Y.; Shu, Y.; Zhou, X.; et al. Correlation of Microbiota in the Gut of Fish Species and Water. 3 Biotech 2020, 10, 472. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Ellakany, H.F.; Elbestawy, A.R.; Abaza, S.S.; Geneedy, A.M.; Khafaga, A.F.; Salem, H.M.; Abd El-Aziz, A.H.; Selim, S.; et al. Inhibition of Microbial Pathogens in Farmed Fish. Mar. Pollut. Bull. 2022, 183, 114003. [Google Scholar] [CrossRef]
- Teitge, F.; Peppler, C.; Steinhagen, D.; Jung-Schroers, V. Effect of Disinfection with Peracetic Acid on the Microbial Community of a Seawater Aquaculture Recirculation System for Pacific White Shrimp (Litopenaeus vannamei). J. Fish Dis. 2020, 43, 991–1017. [Google Scholar] [CrossRef]
- De Schryver, P.; Vadstein, O. Ecological Theory as a Foundation to Control Pathogenic Invasion in Aquaculture. ISME J. 2014, 8, 2360–2368. [Google Scholar] [CrossRef]
- Vestrum, R.I.; Attramadal, K.J.K.; Winge, P.; Li, K.; Olsen, Y.; Bones, A.M.; Vadstein, O.; Bakke, I. Rearing Water Treatment Induces Microbial Selection Influencing the Microbiota and Pathogen Associated Transcripts of Cod (Gadus morhua) Larvae. Front. Microbiol. 2018, 9, 851. [Google Scholar] [CrossRef]
- Ingerslev, H.-C.; von Gersdorff Jørgensen, L.; Lenz Strube, M.; Larsen, N.; Dalsgaard, I.; Boye, M.; Madsen, L. The Development of the Gut Microbiota in Rainbow Trout (Oncorhynchus mykiss) Is Affected by First Feeding and Diet Type. Aquaculture 2014, 424–425, 24–34. [Google Scholar] [CrossRef]
- Wilkes Walburn, J.; Wemheuer, B.; Thomas, T.; Copeland, E.; O’Connor, W.; Booth, M.; Fielder, S.; Egan, S. Diet and Diet-Associated Bacteria Shape Early Microbiome Development in Yellowtail Kingfish (Seriola lalandi). Microb. Biotechnol. 2019, 12, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Wennberg, A.C.; Martins, S.E.; Furseth, K.; Tobiesen, A.E.D.; Hess-Erga, O.-K. Evaluation of Factors Influencing Disinfection Efficacy for Aquaculture; Norsk Institutt for Vannforskning: Oslo, Norway, 2022; ISBN 978-82-577-7537-7. [Google Scholar]
- Kormas, K.A.; Meziti, A.; Mente, E.; Frentzos, A. Dietary Differences Are Reflected on the Gut Prokaryotic Community Structure of Wild and Commercially Reared Sea Bream (Sparus aurata). MicrobiologyOpen 2014, 3, 718–728. [Google Scholar] [CrossRef] [PubMed]
- Najafpour, B.; Pinto, P.I.S.; Moutou, K.A.; Canario, A.V.M.; Power, D.M. Factors Driving Bacterial Microbiota of Eggs from Commercial Hatcheries of European Seabass and Gilthead Seabream. Microorganisms 2021, 9, 2275. [Google Scholar] [CrossRef]
- Bowman, J.P. The Genus Psychrobacter. In The Prokaryotes: A Handbook on the Biology of Bacteria Volume 6: Proteobacteria: Gamma Subclass; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 920–930. ISBN 978-0-387-30746-6. [Google Scholar]
- Denner, E.B.; Mark, B.; Busse, H.J.; Turkiewicz, M.; Lubitz, W. Psychrobacter proteolyticus sp. nov., a Psychrotrophic, Halotolerant Bacterium Isolated from the Antarctic Krill Euphausia superba Dana, Excreting a Cold-Adapted Metalloprotease. Syst. Appl. Microbiol. 2001, 24, 44–53. [Google Scholar] [CrossRef]
- Pukall, R.; Kramer, I.; Rohde, M.; Stackebrandt, E. Microbial Diversity of Cultivatable Bacteria Associated with the North Sea Bryozoan Flustra foliacea. Syst. Appl. Microbiol. 2001, 24, 623–633. [Google Scholar] [CrossRef]
- Baldry, M.G.C. The Bactericidal, Fungicidal and Sporicidal Properties of Hydrogen Peroxide and Peracetic Acid. J. Appl. Bacteriol. 1983, 54, 417–423. [Google Scholar] [CrossRef]
- Niu, T.; Zhou, Z.; Ren, W.; Jiang, L.-M.; Li, B.; Wei, H.; Li, J.; Wang, L. Effects of Potassium Peroxymonosulfate on Disintegration of Waste Sludge and Properties of Extracellular Polymeric Substances. Int. Biodeterior. Biodegrad. 2016, 106, 170–177. [Google Scholar] [CrossRef]
- Pedersen, L.-F.; Pedersen, P.B.; Nielsen, J.L.; Nielsen, P.H. Peracetic Acid Degradation and Effects on Nitrification in Recirculating Aquaculture Systems. Aquaculture 2009, 296, 246–254. [Google Scholar] [CrossRef]
- Acosta, F.; Montero, D.; Izquierdo, M.; Galindo-Villegas, J. High-Level Biocidal Products Effectively Eradicate Pathogenic γ-Proteobacteria Biofilms from Aquaculture Facilities. Aquaculture 2021, 532, 736004. [Google Scholar] [CrossRef]
- Kunanusont, N.; Punyadarsaniya, D.; Jantafong, T.; Pojprasath, T.; Takehara, K.; Ruenphet, S. Bactericidal Efficacy of Potassium Peroxymonosulfate under Various Concentrations, Organic Material Conditions, Exposure Timing and Its Application on Various Surface Carriers. J. Vet. Med. Sci. 2020, 82, 320–324. [Google Scholar] [CrossRef]
- Wu, G.; Wang, J.; Wan, Q.; Cao, S.; Huang, T.; Lu, J.; Ma, J.; Wen, G. Kinetics and Mechanism of Sulfate Radical-and Hydroxyl Radical-Induced Disinfection of Bacteria and Fungal Spores by Transition Metal Ions-Activated Peroxymonosulfate. Water Res. 2023, 243, 120378. [Google Scholar] [CrossRef]
- Marchand, P.-A.; Phan, T.-M.; Straus, D.L.; Farmer, B.D.; Stüber, A.; Meinelt, T. Reduction of In Vitro Growth in Flavobacterium columnare and Saprolegnia parasitica by Products Containing Peracetic Acid. Aquac. Res. 2012, 43, 1861–1866. [Google Scholar] [CrossRef]
- Jakob, H.; Leininger, S.; Lehmann, T.; Jacobi, S.; Gutewort, S. Peroxo Compounds, Inorganic. In Ullmann’s Encyclopedia of Industrial Chemistry; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2007; ISBN 978-3-527-30673-2. [Google Scholar]
- Kiejza, D.; Kotowska, U.; Polińska, W.; Karpińska, J. Peracids—New Oxidants in Advanced Oxidation Processes: The Use of Peracetic Acid, Peroxymonosulfate, and Persulfate Salts in the Removal of Organic Micropollutants of Emerging Concern—A Review. Sci. Total Environ. 2021, 790, 148195. [Google Scholar] [CrossRef] [PubMed]
- Welter, D.K.; Ruaud, A.; Henseler, Z.M.; De Jong, H.N.; van Coeverden de Groot, P.; Michaux, J.; Gormezano, L.; Waters, J.L.; Youngblut, N.D.; Ley, R.E. Free-Living, Psychrotrophic Bacteria of the Genus Psychrobacter Are Descendants of Pathobionts. mSystems 2021, 6, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Wang, R.; Zhao, P.; Chen, R.; Zhou, W.; Tang, L.; Tang, X. Interaction between Chlorella vulgaris and Bacteria: Interference and Resource Competition. Acta Oceanol. Sin. 2014, 33, 135–140. [Google Scholar] [CrossRef]
- Frans, I.; Michiels, C.W.; Bossier, P.; Willems, K.A.; Lievens, B.; Rediers, H. Vibrio Anguillarum as a Fish Pathogen: Virulence Factors, Diagnosis and Prevention. J. Fish Dis. 2011, 34, 643–661. [Google Scholar] [CrossRef]
- Hussein, M.A.; El-tahlawy, A.S.; Abdelmoneim, H.M.; Abdallah, K.M.E.; Bayomi, R.M.E. Pseudomonas aeruginosa in Fish and Fish Products: A Review on the Incidence, Public Health Significance, Virulence Factors, Antimicrobial Resistance, and Biofilm Formation. J. Adv. Vet. Res. 2023, 13, 1464–1468. [Google Scholar]
- Toxqui-Rodríguez, S.; Naya-Català, F.; Sitjà-Bobadilla, A.; Piazzon, M.C.; Pérez-Sánchez, J. Fish Microbiomics: Strengths and Limitations of MinION Sequencing of Gilthead Sea Bream (Sparus aurata) Intestinal Microbiota. Aquaculture 2023, 569, 739388. [Google Scholar] [CrossRef]
- Wuertz, S.; Beça, F.; Kreuz, E.; Wanka, K.M.; Azeredo, R.; Machado, M.; Costas, B. Two Probiotic Candidates of the Genus Psychrobacter Modulate the Immune Response and Disease Resistance after Experimental Infection in Turbot (Scophthalmus maximus, Linnaeus 1758). Fishes 2023, 8, 144. [Google Scholar] [CrossRef]
- Lahnsteiner, F.; Lahnsteiner, E.; Duenser, A. Suitability of Different Live Feed for First Feeding of Freshwater Fish Larvae. Aquac. J. 2023, 3, 107–120. [Google Scholar] [CrossRef]
- Ina-Salwany, M.Y.; Al-saari, N.; Mohamad, A.; Mursidi, F.-A.; Mohd-Aris, A.; Amal, M.N.A.; Kasai, H.; Mino, S.; Sawabe, T.; Zamri-Saad, M. Vibriosis in Fish: A Review on Disease Development and Prevention. J. Aquat. Anim. Health 2019, 31, 3–22. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disinfection | Active Ingredient | Application | Dry Period | No. of Tanks | No. of Samples | |
|---|---|---|---|---|---|---|
| Larva | Water | |||||
| Dis1 | PAA-HP | Nebulization | 0 days | 2 | 30 | 30 |
| Dis2 | PMS-SC | Wash | 7 days | 2 | 30 | 30 |
| Dis3 | PMS-SC | Wash | 50 days | 4 | 60 | 60 |
| Total | 8 | 120 | 120 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apostolopoulou, G.; Bel Mokhtar, N.; Asimakis, E.; Dionyssopoulou, E.; Toskas, K.; Koumoundouros, G.; Tsiamis, G.; Stathopoulou, P. The Impact of Tank Disinfectants on the Development of Microbiota in Gilthead Seabream (Sparus aurata) Larviculture Systems. Microorganisms 2025, 13, 1359. https://doi.org/10.3390/microorganisms13061359
Apostolopoulou G, Bel Mokhtar N, Asimakis E, Dionyssopoulou E, Toskas K, Koumoundouros G, Tsiamis G, Stathopoulou P. The Impact of Tank Disinfectants on the Development of Microbiota in Gilthead Seabream (Sparus aurata) Larviculture Systems. Microorganisms. 2025; 13(6):1359. https://doi.org/10.3390/microorganisms13061359
Chicago/Turabian StyleApostolopoulou, Georgia, Naima Bel Mokhtar, Elias Asimakis, Eva Dionyssopoulou, Kosmas Toskas, George Koumoundouros, George Tsiamis, and Panagiota Stathopoulou. 2025. "The Impact of Tank Disinfectants on the Development of Microbiota in Gilthead Seabream (Sparus aurata) Larviculture Systems" Microorganisms 13, no. 6: 1359. https://doi.org/10.3390/microorganisms13061359
APA StyleApostolopoulou, G., Bel Mokhtar, N., Asimakis, E., Dionyssopoulou, E., Toskas, K., Koumoundouros, G., Tsiamis, G., & Stathopoulou, P. (2025). The Impact of Tank Disinfectants on the Development of Microbiota in Gilthead Seabream (Sparus aurata) Larviculture Systems. Microorganisms, 13(6), 1359. https://doi.org/10.3390/microorganisms13061359