
Impacts of Emergency Treatments on Sediment Microbial Communities Following Sudden Thallium Contamination Events: A Microcosm Study


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Microcosm Setup
2.2. Sequencing and Data Processing
2.3. Statistics and Visualization
3. Results
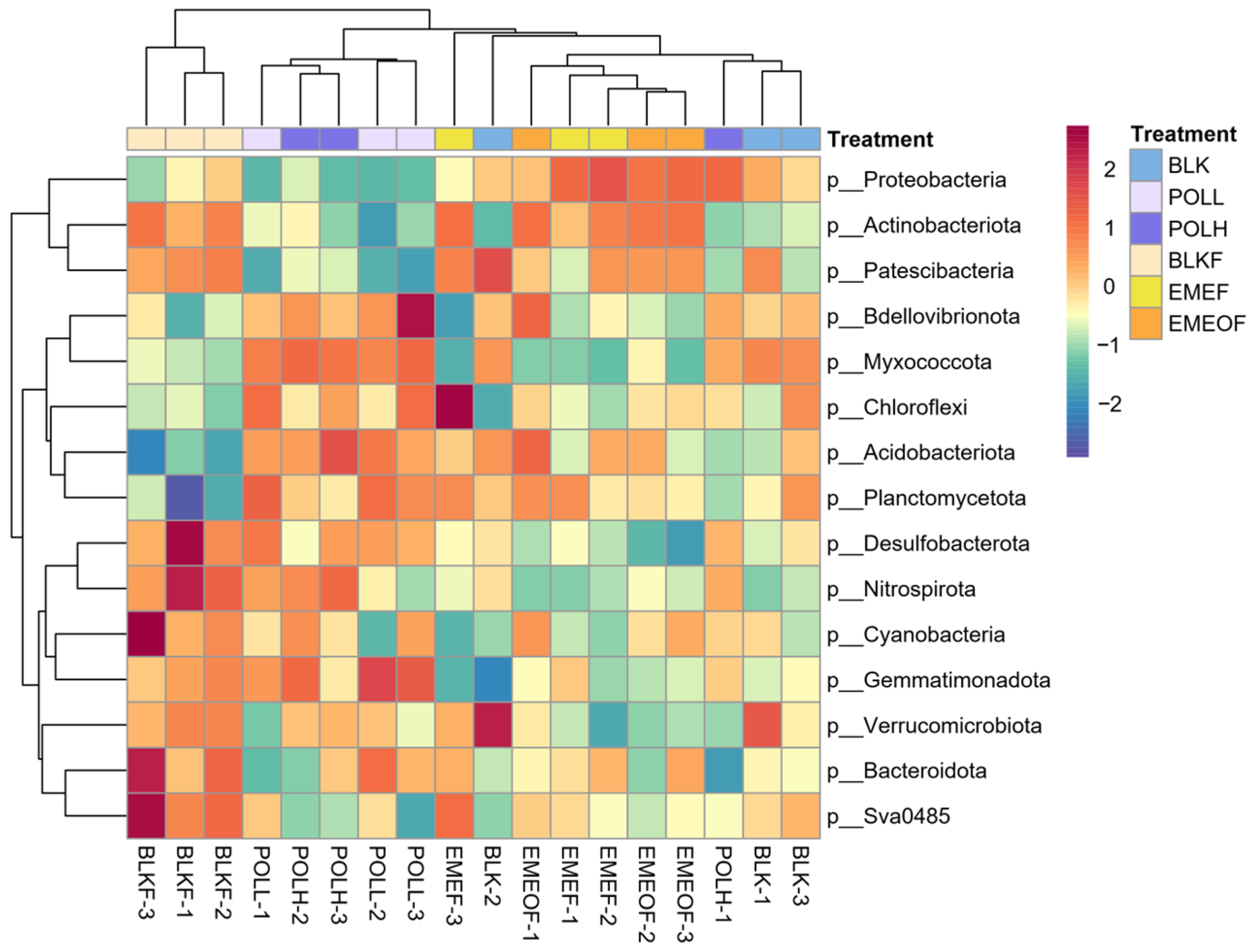
3.1. Microbial Community Composition During Incubation
3.2. Microbial Community Diversity
3.3. Potential Function Shift in Microbial Community
4. Discussion
4.1. Impacts of Thallium Shock Loading on Microbial Communities
4.2. Effects of Thallium-Bearing Flocs on Microbial Communities
4.3. Influence of Thallium-Free Flocs on Microbial Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cheam, V. Thallium contamination of water in Canada. Water Qual. Res. J. Canada 2001, 36, 851–878. [Google Scholar] [CrossRef]
- Peter, A.L.J.; Viraraghavan, T. Thallium: A review of public health and environmental concerns. Environ. Int. 2005, 31, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Lis, J.; Pasieczna, A.; Karbowska, B.; Zembrzuski, W.; Lukaszewski, Z. Thallium in soils and stream sediments of a Zn-Pb mining and smelting area. Environ. Sci. Technol. 2003, 37, 4569–4572. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.F.; Yang, F.; Li, S.H.; Zheng, B.S.; Ning, Z.P. Thallium pollution in China: A geo-environmental perspective. Sci. Total Environ. 2012, 421, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yi, X.; Dang, Z.; Yu, H.; Zeng, T.; We, C.; Feng, C. Fate of Fe and Cd upon microbial reduction of Cd-loaded polyferric flocs by Shewanella oneidensis MR-1. Chemosphere 2016, 144, 2065–2072. [Google Scholar] [CrossRef]
- Liu, J.; Ouyang, Q.; Wang, L.; Wang, J.; Zhang, Q.; Wei, X.; Lin, Y.; Zhou, Y.; Yuan, W.; Xiao, T. Quantification of smelter-derived contributions to thallium contamination in river sediments: Novel insights from thallium isotope evidence. J. Hazard. Mater. 2022, 424, 127594. [Google Scholar] [CrossRef]
- Zhou, Y.T.; He, H.P.; Wang, J.; Liu, J.; Lippold, H.; Bao, Z.A.; Wang, L.L.; Lin, Y.Y.; Fang, F.; Huang, Y.L.; et al. Stable isotope fractionation of thallium as novel evidence for its geochemical transfer during lead-zinc smelting activities. Sci. Total Environ. 2022, 803, 8. [Google Scholar] [CrossRef]
- D’Orazio, M.; Campanella, B.; Bramanti, E.; Ghezzi, L.; Onor, M.; Vianello, G.; Vittori-Antisari, L.; Petrini, R. Thallium pollution in water, soils and plants from a past-mining site of Tuscany: Sources, transfer processes and toxicity. J. Geochem. Explor. 2020, 209, 14. [Google Scholar] [CrossRef]
- Houmei, Z. Guangyuan: Promptly address the thallium pollution incident in the Jialing River. Environ. Superv. China 2017, 2. [Google Scholar]
- Zhang, X.; Chen, C.; Mi, Z.; Wang, C. Emergency Cadmium Removal and Water Purification Technologies for Drinking Water and Emergency Response to Sudden Environmental Incidents in the Longjiang River, Guangxi. Water Wastewater Eng. 2013, 39, 9. [Google Scholar]
- Karbowska, B. Presence of thallium in the environment: Sources of contaminations, distribution and monitoring methods. Environ. Monit. Assess. 2016, 188, 19. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Luo, X.W.; Sun, Y.Q.; Tsang, D.C.W.; Qi, J.Y.; Zhang, W.L.; Li, N.; Yin, M.L.; Wang, J.; Lippold, H.; et al. Thallium pollution in China and removal technologies for waters: A review. Environ. Int. 2019, 126, 771–790. [Google Scholar] [CrossRef]
- Liu, J.; Luo, X.W.; Wang, J.; Xiao, T.F.; Chen, D.Y.; Sheng, G.D.; Yin, M.L.; Lippold, H.; Wang, C.L.; Chen, Y.H. Thallium contamination in arable soils and vegetables around a steel plant A newly-found significant source of Tl pollution in South China. Environ. Pollut. 2017, 224, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Huang, D.; Zhang, Z.; Wang, J.; Bing, Y.; Chang, S.; Guo, Q. Pollution reduction technologies for emergent water pollution disposal in river basins. Chin. J. Environ. Eng. 2021, 15, 2233–2238. [Google Scholar]
- Fu, L.; Song, Y.; Wang, P.; Zhao, M.; Huang, Q.; Xi, H.; Yu, Y.; Wu, C. Emergency removal technologies and case study of typical water pollutants in abrupt environmental accidents. J. Environ. Eng. Technol. 2022, 12, 322–328. [Google Scholar]
- Bai, Y.; Zhang, H.; He, X.; Zhu, Z.; Tan, Z.; Sun, W.; Huang, D.; Bing, Y.; Guo, P. Emergency treatment strategies and influencing factors of thallium pollution in small watershed: A case study of Xiangshuihe Reservoir and its upstream in Qujing City. Chin. J. Environ. Eng. 2022, 16, 2558–2565. [Google Scholar]
- Schreier, H.; Erdos, G.; Reimer, K.; König, B.; König, W.; Fleischer, W. Molecular Effects of Povidone-Iodine on Relevant Microorganisms: An Electron-Microscopic and Biochemical Study. Dermatology 1998, 200, 111–116. [Google Scholar] [CrossRef]
- Yang, Z.H.; Ma, X.M.; Zhou, H.B.; Ma, J.X.; Ding, Y.X.; He, D. Chelated heavy metals removal by in-situ formed Fe(II) and Fe(III) iron (oxy)hydroxides: Mechanism and performance. J. Environ. Chem. Eng. 2023, 11, 8. [Google Scholar] [CrossRef]
- Shuaib, M.; Azam, N.; Bahadur, S.; Romman, M.; Yu, Q. Variation and succession of microbial communities under the conditions of persistent heavy metal and their survival mechanism. Microb. Pathog. 2021, 150, 104713. [Google Scholar] [CrossRef]
- Wahsha, M.; Nadimi-Goki, M.; Fornasier, F.; Al-Jawasreh, R.; Hussein, E.I.; Bini, C. Microbial enzymes as an early warning management tool for monitoring mining site soils. Catena 2017, 148, 40–45. [Google Scholar] [CrossRef]
- Chen, X.L.; Wang, J.; Pan, C.Y.; Feng, L.S.; Chen, S.L.; Xie, S.G. Metagenomic insights into the influence of thallium spill on sediment microbial community. Environ. Pollut. 2023, 317, 11. [Google Scholar] [CrossRef]
- Liu, J.; Wan, Y.B.; Wei, X.D.; She, J.Y.; Ouyang, Q.E.; Deng, P.Y.; Hu, H.Y.; Zhang, X.Y.; Fang, M.Y.; Wei, X.L.; et al. Microbial diversity in paddy rhizospheric soils around a large industrial thallium sulfide utilization zone. Environ. Res. 2023, 216, 114627. [Google Scholar] [CrossRef]
- She, J.Y.; Liu, J.; He, H.P.; Zhang, Q.; Lin, Y.Y.; Wang, J.; Yin, M.L.; Wang, L.L.; Wei, X.D.; Huang, Y.L.; et al. Microbial response and adaption to thallium contamination in soil profiles. J. Hazard. Mater. 2022, 423, 127080. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Wang, Q.; Lv, M.; Chen, L. Microorganism remediation strategies towards heavy metals. Chem. Eng. J. 2019, 360, 1553–1563. [Google Scholar] [CrossRef]
- Tang, H.; Xiang, G.; Xiao, W.; Yang, Z.; Zhao, B. Microbial mediated remediation of heavy metals toxicity: Mechanisms and future prospects. Front. Plant Sci. 2024, 15, 23. [Google Scholar] [CrossRef]
- Yan, S.; Zhang, Z.; Wang, J.; Xia, Y.; Chen, S.; Xie, S. River sediment microbial community composition and function impacted by thallium spill. Sci. Total Environ. 2023, 880, 9. [Google Scholar] [CrossRef]
- Peng, J.P.; He, Y.; Ding, X.L. Green Waters Bathed in Spring—Observations on the River and Lake Chief System in Yichun City. China Water Resour. 2020, 12–15. [Google Scholar]
- Zhu, Y.; Yan, S.; Chen, X.; Li, Y.; Xie, S. Thallium spill shifts the structural and functional characteristics of viral communities with different lifestyles in river sediments. Sci. Total Environ. 2024, 947, 174531. [Google Scholar] [CrossRef]
- GB 3838-2002; Environmental Quality Standards for Surface Water. National Standard of the People’s Republic of China: Beijing, China, 2002.
- Chen, X.L.; Wang, J.; Pan, C.Y.; Feng, L.S.; Guo, Q.W.; Chen, S.L.; Xie, S.G. Metagenomic analysis reveals the response of microbial community in river sediment to accidental antimony contamination. Sci. Total Environ. 2022, 813, 10. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Hall, M.; Beiko, R.G. 16S rRNA Gene Analysis with QIIME2. Methods Mol. Biol. 2018, 1849, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Xiao, E.Z.; Krumins, V.; Xiao, T.F.; Dong, Y.R.; Tang, S.; Ning, Z.P.; Huang, Z.Y.; Sun, W.M. Depth-resolved microbial community analyses in two contrasting soil cores contaminated by antimony and arsenic. Environ. Pollut. 2017, 221, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Heisi, H.D.; Nkuna, R.; Matambo, T. Rhizosphere microbial community structure and PICRUSt2 predicted metagenomes function in heavy metal contaminated sites: A case study of the Blesbokspruit wetland. Sci. Total Environ. 2025, 959, 178147. [Google Scholar] [CrossRef] [PubMed]
- Chambers, J.M. S, R, and Data Science. R J. 2020, 12, 462–476. [Google Scholar] [CrossRef]
- Sakai, T.; Association for Computing Machinery. Two Sample T-tests for IR Evaluation: Student or Welch? In Proceedings of the 39th International ACM SIGIR Conference on Research and Development in Information Retrieval, Pisa, Italy, 17–21 July 2016; pp. 1045–1048. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.6-4. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 9 August 2024).
- Anderson, M.J.; Walsh, D.C.I. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? Ecol. Monogr. 2013, 83, 557–574. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Kelly, B.J.; Robert, G.; Kyle, B.; Scott, S.M.; Lewis, J.D.; Collman, R.G.; Bushman, F.D.; Hongzhe, L. Power and sample-size estimation for microbiome studies using pairwise distances and PERMANOVA. Bioinformatics 2015, 31, 2461–2468. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis. 2016. Available online: https://ggplot2.tidyverse.org (accessed on 9 August 2024).
- Kolde, R. Pheatmap: Pretty Heatmaps. R Package Version 1.0.12. 2019. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 9 August 2024).
- Rasool, A.; Xiao, T.F. Response of microbial communities to elevated thallium contamination in river sediments. Geomicrobiol. J. 2018, 35, 854–868. [Google Scholar] [CrossRef]
- Cotto, I.; Vilardi, K.J.; Huo, L.X.; Fogarty, E.C.; Khunjar, W.; Wilson, C.; De Clippeleir, H.; Gilmore, K.; Bailey, E.; Lücker, S.; et al. Low diversity and microdiversity of comammox bacteria in wastewater systems suggest specific adaptations within the Ca. Nitrospira nitrosa cluster. Water Res. 2023, 229, 13. [Google Scholar] [CrossRef]
- Daims, H.; Lebedeva, E.V.; Pjevac, P.; Han, P.; Herbold, C.; Albertsen, M.; Jehmlich, N.; Palatinszky, M.; Vierheilig, J.; Bulaev, A.; et al. Complete nitrification by Nitrospira bacteria. Nature 2015, 528, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Wang, L.Y.; Xu, K.; Li, K.; Ren, H.Q. Revealing taxon-specific heavy metal-resistance mechanisms in denitrifying phosphorus removal sludge using genome-centric metaproteomics. Microbiome 2021, 9, 67. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.C.; Tran, P.Q.; Cowley, E.S.; Trembath-Reichert, E.; Anantharaman, K. Diversity and ecology of microbial sulfur metabolism. Nat. Rev. Microbiol. 2025, 23, 122–140. [Google Scholar] [CrossRef]
- Kraft, B.; Strous, M.; Tegetmeyer, H.E. Microbial nitrate respiration—Genes, enzymes and environmental distribution. J. Biotechnol. 2011, 155, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Simpson, P.J.L.; Richardson, D.J.; Codd, R. The periplasmic nitrate reductase in Shewanella: The resolution, distribution and functional implications of two NAP Isoforms, NapEDABC and NapDAGHB. Microbiology-Sgm 2010, 156, 302–312. [Google Scholar] [CrossRef]
- Rusmana, I.; Nedwell, D.B. Use of chlorate as a selective inhibitor to distinguish membrane-bound nitrate reductase (Nar) and periplasmic nitrate reductase (Nap) of dissimilative nitrate reducing bacteria in sediment. Fems Microbiol. Ecol. 2004, 48, 379–386. [Google Scholar] [CrossRef]
- Chovanec, P.; Sparacino-Watkins, C.; Zhang, N.; Basu, P.; Stolz, J.F. Microbial reduction of chromate in the presence of nitrate by three nitrate respiring organisms. Front. Microbiol. 2012, 3, 32684. [Google Scholar] [CrossRef]
- Chen, X.; Liu, C.M.; Zhu, B.L.; Wei, W.X.; Sheng, R. The Contribution of Nitrate Dissimilation to Nitrate Consumption in narG- and napA-Containing Nitrate Reducers with Various Oxygen and Nitrate Supplies. Microbiol. Spectr. 2022, 10, 12. [Google Scholar] [CrossRef]
- Bothe, H.; Schmitz, O.; Yates, M.G.; Newton, W.E. Nitrogen Fixation and Hydrogen Metabolism in Cyanobacteria. Microbiol. Mol. Biol. Rev. 2010, 74, 529–551. [Google Scholar] [CrossRef]
- Guo, Y.F.; Cheng, S.L.; Fang, H.J.; Geng, J.; Li, Y.A.; Shi, F.Y.; Wang, H.; Chen, L.; Zhou, Y. Copper and cadmium co-contamination increases the risk of nitrogen loss in red paddy soils. J. Hazard. Mater. 2024, 479, 10. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Hämmerle, I.; Fuchslueger, L.; Hofhansl, F.; Knoltsch, A.; Schnecker, J.; Takriti, M.; Watzka, M.; Wild, B.; et al. Adjustment of microbial nitrogen use efficiency to carbon: Nitrogen imbalances regulates soil nitrogen cycling. Nat. Commun. 2014, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Broman, E.; Izabel-Shen, D.; Rodríguez-Gijón, A.; Bonaglia, S.; Garcia, S.L.; Nascimento, F.J.A. Microbial functional genes are driven by gradients in sediment stoichiometry, oxygen, and salinity across the Baltic benthic ecosystem. Microbiome 2022, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Qu, G.; Xu, R.; Liu, X.; Jin, C. Iron-based materials for immobilization of heavy metals in contaminated soils: A critical review. J. Environ. Chem. Eng. 2024, 12, 113741. [Google Scholar] [CrossRef]
- Zhang, L.M.; Zeng, Q.; Liu, X.; Chen, P.; Guo, X.X.; Ma, L.Y.Z.; Dong, H.L.; Huang, Y. Iron reduction by diverse actinobacteria under oxic and pH-neutral conditions and the formation of secondary minerals. Chem. Geol. 2019, 525, 390–399. [Google Scholar] [CrossRef]
- Behera, S.; Das, S. Potential and prospects of Actinobacteria in the bioremediation of environmental pollutants: Cellular mechanisms and genetic regulations. Microbiol. Res. 2023, 273, 17. [Google Scholar] [CrossRef]
- Zhang, T.; Sun, Y.F.; Parikh, S.J.; Colinet, G.; Garland, G.; Huo, L.J.; Zhang, N.; Shan, H.; Zeng, X.B.; Su, S.M. Water-fertilizer regulation drives microorganisms to promote iron, nitrogen and manganese cycling: A solution for arsenic and cadmium pollution in paddy soils. J. Hazard. Mater. 2024, 477, 10. [Google Scholar] [CrossRef]
- Awang, N.A.; Salleh, W.N.W.; Aziz, F.; Yusof, N.; Ismail, A.F. A review on preparation, surface enhancement and adsorption mechanism of biochar-supported nano zero-valent iron adsorbent for hazardous heavy metals. J. Chem. Technol. Biotechnol. 2023, 98, 22–44. [Google Scholar] [CrossRef]
- Chen, Z.; An, L.; Wei, H.; Zhang, J.; Zou, Q.; Sun, M.; Huang, L.; Liu, M. Nitrate alleviate dissimilatory iron reduction and arsenic mobilization by driving microbial community structure change. Surf. Interfaces 2021, 26, 101421. [Google Scholar] [CrossRef]
- Liu, Y.; Xue, C.; Yu, S.R.; Li, F. Variations of Abundance and Community Structure of Ammonia Oxidizers and Nitrification Activity in Two Paddy Soils Polluted by Heavy Metals. Geomicrobiol. J. 2019, 36, 1–10. [Google Scholar] [CrossRef]
- Zhang, X.W.; Feng, C.Y.; Wei, D.; Liu, X.L.; Luo, W.C. Optimization of “sulfur-iron-nitrogen” cycle in constructed wetlands by adjusting siderite/sulfur (Fe/S) ratio. J. Environ. Manag. 2024, 363, 11. [Google Scholar] [CrossRef]
- Wang, Z.; Ruan, X.; Li, R.; Zhang, Y. Microbial interaction patterns and nitrogen cycling regularities in lake sediments under different trophic conditions. Sci. Total Environ. 2024, 907, 12. [Google Scholar] [CrossRef]
- Tong, H.; Li, J.H.; Chen, M.J.; Fang, Y.J.; Yi, X.; Dong, L.H.; Jiang, Q.; Liu, C.S. Iron oxidation coupled with nitrate reduction affects the acetate-assimilating microbial community structure elucidated by stable isotope probing in flooded paddy soil. Soil Biol. Biochem. 2023, 183, 9. [Google Scholar] [CrossRef]
- Lin, J.F.; Yin, M.L.; Wang, J.; Liu, J.; Tsang, D.C.W.; Wang, Y.X.; Lin, M.; Li, H.C.; Zhou, Y.Y.; Song, G.; et al. Geochemical fractionation of thallium in contaminated soils near a large-scale Hg-Tl mineralised area. Chemosphere 2020, 239, 8. [Google Scholar] [CrossRef]
- Li, Q.; Yin, J.; Wu, L.; Li, S.; Chen, L. Effects of biochar and zero valent iron on the bioavailability and potential toxicity of heavy metals in contaminated soil at the field scale. Sci. Total Environ. 2023, 897, 9. [Google Scholar] [CrossRef]
- Ghezzi, L.; D’Orazio, M.; Doveri, M.; Lelli, M.; Petrini, R.; Giannecchini, R. Groundwater and potentially toxic elements in a dismissed mining area: Thallium contamination of drinking spring water in the Apuan Alps (Tuscany, Italy). J. Geochem. Explor. 2019, 197, 84–92. [Google Scholar] [CrossRef]
- Li, N.; Zhou, Y.C.; Liu, J.; Tsang, D.C.W.; Wang, J.; She, J.Y.; Zhou, Y.T.; Yin, M.L.; Chen, Z.R.; Chen, D.Y. Persistent thallium contamination in river sediments, source apportionment and environmental implications. Ecotoxicol. Environ. Saf. 2020, 202, 9. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.D.; Wang, J.; She, J.Y.; Sun, J.; Liu, J.; Wang, Y.X.; Yang, X.; Ouyang, Q.E.; Lin, Y.Y.; Xiao, T.F.; et al. Thallium geochemical fractionation and migration in Tl-As rich soils: The key controls. Sci. Total Environ. 2021, 784, 9. [Google Scholar] [CrossRef]
- Chan, C.S.; Fakra, S.C.; Edwards, D.C.; Emerson, D.; Banfield, J.F. Iron oxyhydroxide mineralization on microbial extracellular polysaccharides. Geochim. Cosmochim. Acta 2009, 73, 3807–3818. [Google Scholar] [CrossRef]
- Peng, J.; Feng, F.; Zhang, G.; Zou, L. Transcriptome Analysis Reveals the Inhibitory Effect of Cu2 on Polyferric Sulfate Floc Reduction by Shewanella putrefaciens CN32. Appl. Biochem. Biotechnol. 2024, 196, 4862–4873. [Google Scholar] [CrossRef]
- Lin, Y.; Ye, Y.; Hu, Y.; Shi, H. The variation in microbial community structure under different heavy metal contamination levels in paddy soils. Ecotoxicol. Environ. Saf. 2019, 180, 557–564. [Google Scholar] [CrossRef]
- Rasool, A.; Nasim, W.; Xiao, T.; Ali, W.; Chaudhary, H.J. Microbial diversity response in thallium polluted riverbank soils of the Lanmuchang. Ecotoxicol. Environ. Saf. 2019, 187, 109854. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhao, Y.; Zhang, B.; Wang, J.; Zhu, L.; Hu, B. Deterministic Effect of pH on Shaping Soil Resistome Revealed by Metagenomic Analysis. Environ. Sci. Technol. 2023, 57, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Nwaehiri, U.L.; Akwukwaegbu, P.I.; Nwoke, B.E.B. Bacterial remediation of heavy metal polluted soil and effluent from paper mill industry. Environ. Health Toxicol. 2020, 35, e2020009. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, Z.M.; Zhao, J.; Ma, J.W.; Yu, Q.G.; Zou, P.; Lin, H.; Ma, J.C. Fate of heavy metals and bacterial community composition following biogas slurry application in a single rice cropping system. J. Soils Sediments 2022, 22, 968–981. [Google Scholar] [CrossRef]
- Liu, H.W.; Yuan, R.L.; Sarkodie, E.K.; Tang, J.H.; Jiang, L.H.; Miao, B.; Liu, X.D.; Zhang, S.Y. Insight into functional microorganisms in wet-dry conversion to alleviate the toxicity of chromium fractions in red soil. Front. Microbiol. 2022, 13, 15. [Google Scholar] [CrossRef]
- Ke, Y.X.; Si, S.C.; Zhang, Z.Y.; Geng, P.Y.; Shen, Y.H.; Wang, J.Q.; Zhu, X.L. Synergistic passivation performance of cadmium pollution by biochar combined with sulfate reducing bacteria. Environ. Technol. Innov. 2023, 32, 12. [Google Scholar] [CrossRef]
- Zhao, Z.; Sun, C.; Li, Y.; Yu, Q.; Jin, Z.; Wang, M.; Liang, L.; Zhang, Y. Driving microbial sulfur cycle for phenol degradation coupled with Cr(VI) reduction via Fe(III)/Fe(II) transformation. Chem. Eng. J. 2020, 393, 124801. [Google Scholar] [CrossRef]
- Chen, H.; Zhao, X.H.; Cheng, Y.Y.; Jiang, M.J.; Li, X.; Xue, G. Iron Robustly Stimulates Simultaneous Nitrification and Denitrification Under Aerobic Conditions. Environ. Sci. Technol. 2018, 52, 1404–1412. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, X.; Huang, Z.; Chen, S.; Zhang, Z.; Wang, J.; Wen, X.; Yang, Y. Impacts of Emergency Treatments on Sediment Microbial Communities Following Sudden Thallium Contamination Events: A Microcosm Study. Microorganisms 2025, 13, 1336. https://doi.org/10.3390/microorganisms13061336
Cai X, Huang Z, Chen S, Zhang Z, Wang J, Wen X, Yang Y. Impacts of Emergency Treatments on Sediment Microbial Communities Following Sudden Thallium Contamination Events: A Microcosm Study. Microorganisms. 2025; 13(6):1336. https://doi.org/10.3390/microorganisms13061336
Chicago/Turabian StyleCai, Xiaodie, Zeqiang Huang, Sili Chen, Zhengke Zhang, Jingsong Wang, Xinyu Wen, and Yuyin Yang. 2025. "Impacts of Emergency Treatments on Sediment Microbial Communities Following Sudden Thallium Contamination Events: A Microcosm Study" Microorganisms 13, no. 6: 1336. https://doi.org/10.3390/microorganisms13061336
APA StyleCai, X., Huang, Z., Chen, S., Zhang, Z., Wang, J., Wen, X., & Yang, Y. (2025). Impacts of Emergency Treatments on Sediment Microbial Communities Following Sudden Thallium Contamination Events: A Microcosm Study. Microorganisms, 13(6), 1336. https://doi.org/10.3390/microorganisms13061336