

Unveiling Species Diversity Within Early-Diverging Fungi from China VIII: Four New Species in Mortierellaceae (Mortierellomycota)


Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Morphology
2.1.1. Isolation
2.1.2. Morphology
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Phylogenetic Analyses
3. Results
3.1. Molecular Phylogeny
3.2. Taxonomy
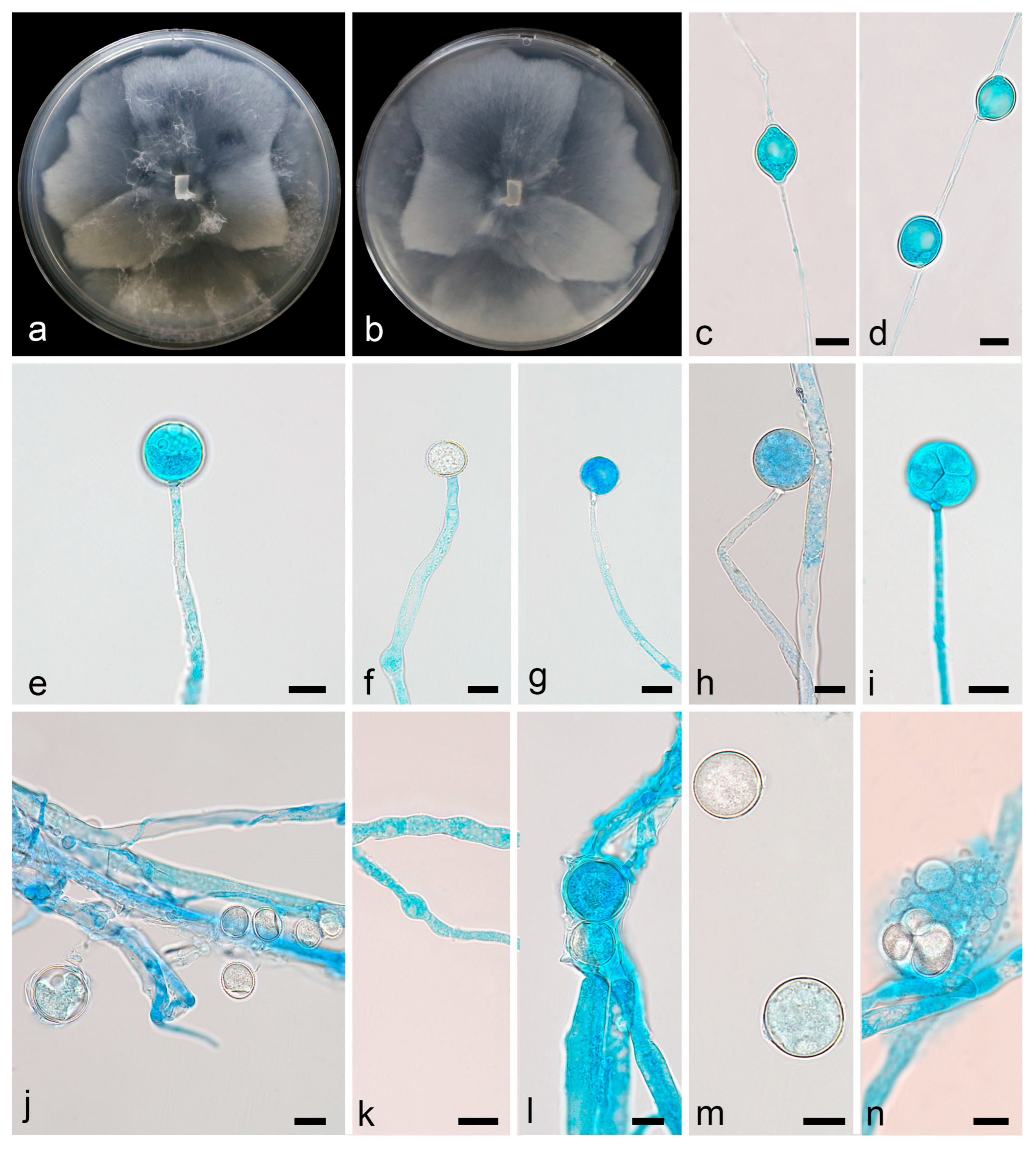
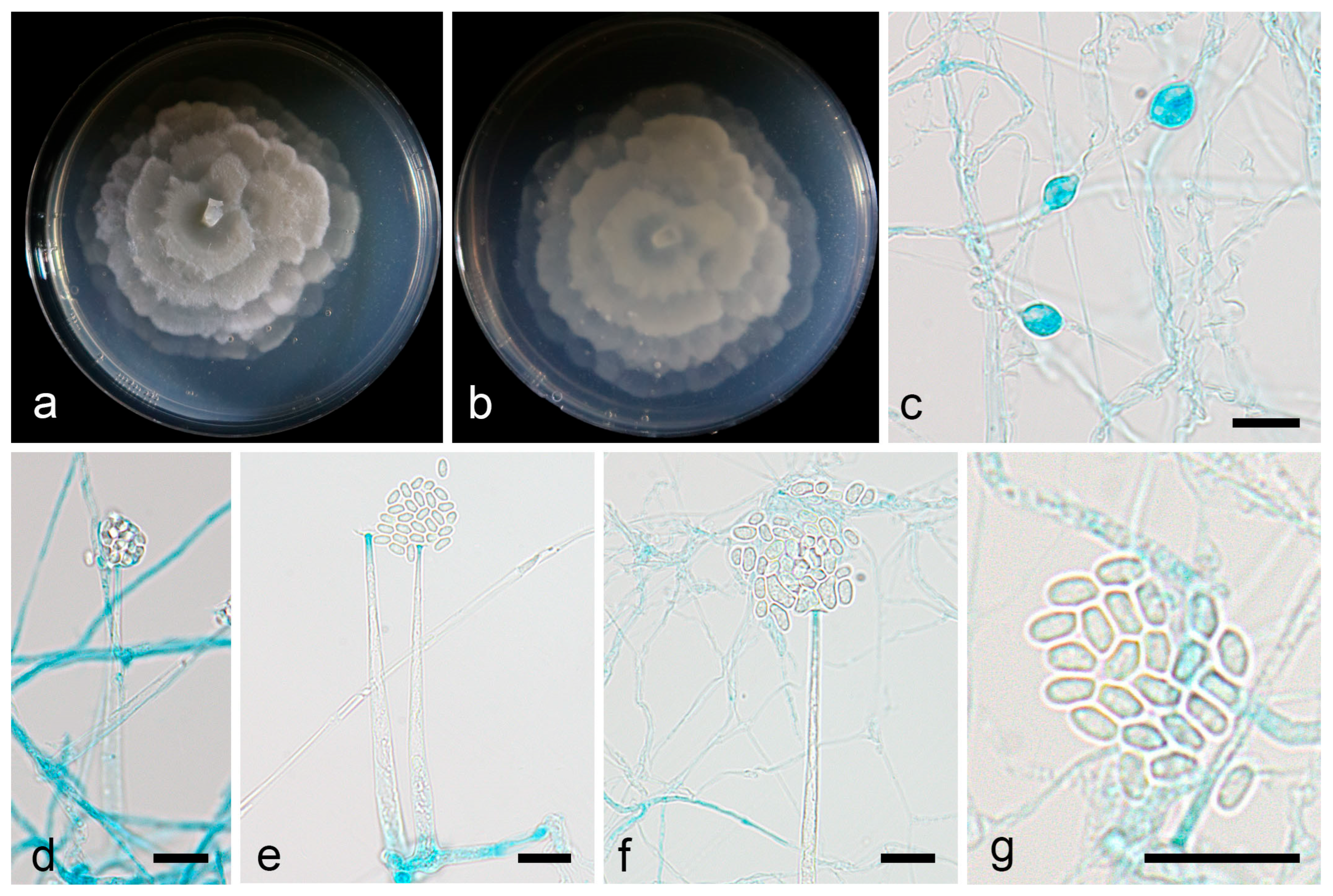
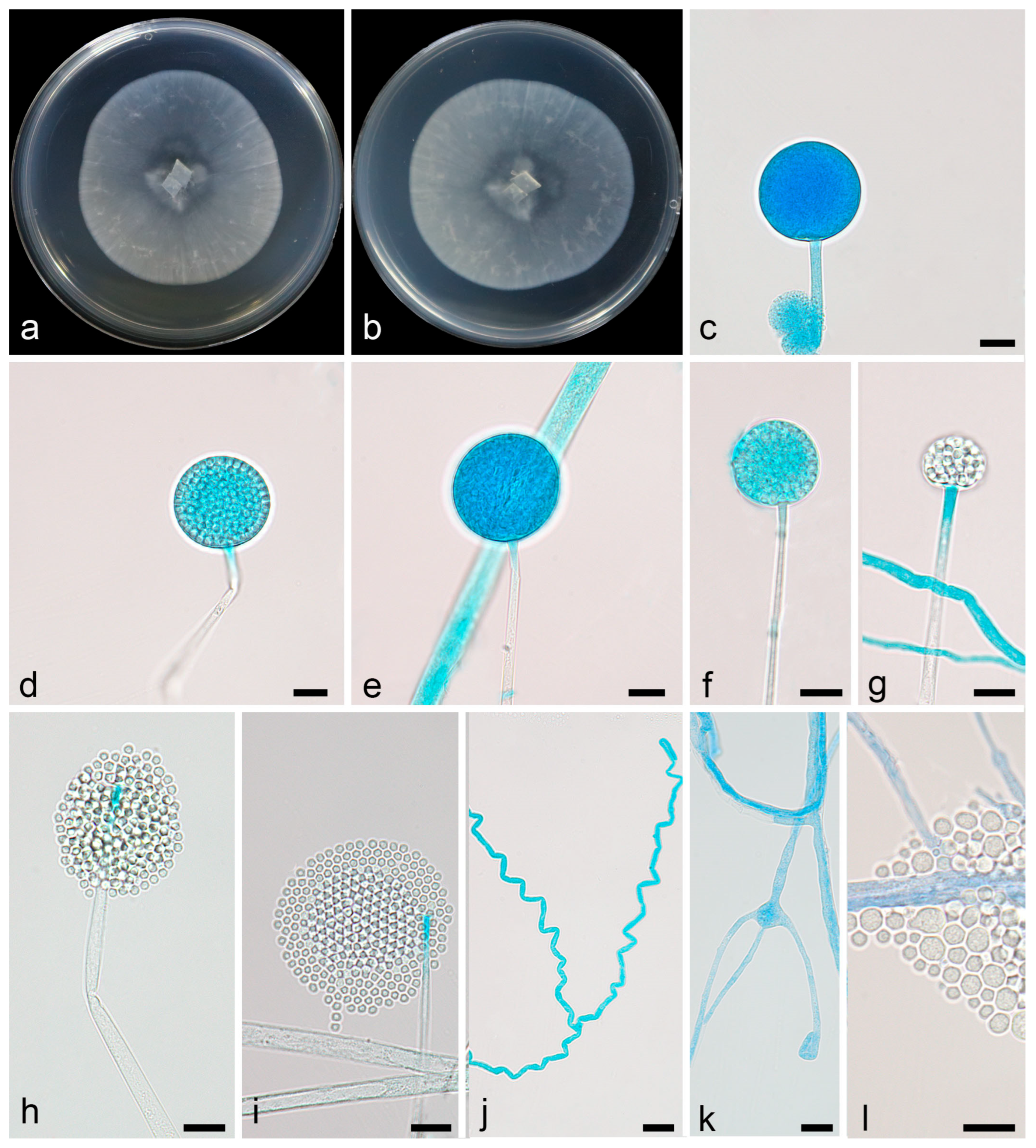
3.2.1. Linnemannia rotunda X.Y. Ji, Y. Jiang & X.Y. Liu, sp. nov.
3.2.2. Mortierella acuta X.Y. Ji, Y. Jiang, and X.Y. Liu, sp. nov.
3.2.3. Mortierella oedema X.Y. Ji, Y. Jiang, and X.Y. Liu, sp. nov.
3.2.4. Mortierella tibetensis X.Y. Ji, Y. Jiang, and X.Y. Liu, sp. nov.
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wijayawardene, N.N.; Hyde, K.D.; Mikhailov, K.V.; Péter, G.; Aptroot, A.; Pires-Zottarelli, C.L.A.; Goto, B.T.; Tokarev, Y.S.; Haelewaters, D.; Karunarathna, S.C.; et al. Classes and phyla of the kingdom Fungi. Fungal Divers. 2024, 128, 1–165. [Google Scholar] [CrossRef]
- Smith, M.E.; Gryganskyi, A.; Bonito, G.; Nouhra, E.; Moreno-Arroyo, B.; Benny, G. Phylogenetic analysis of the genus Modicella reveals an independent evolutionary origin of sporocarp-forming fungi in the Mortierellales. Fungal Genet. Biol. 2013, 61, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Linnemann, G. Die Mucorineen-Gattung Mortierella Coemans. Pflanzenforschung 1941, 23, 1–64. [Google Scholar]
- Gams, W. A key to the species of Mortierella. Persoonia 1977, 9, 381–391. [Google Scholar]
- Petkovits, T.; Nagy, L.G.; Hoffmann, K.; Wagner, L.; Nyilasi, I.; Griebel, T.; Schnabelrauch, D.; Vogel, H.; Voigt, K.; Vágvölgyi, C.; et al. Data partitions, Bayesian analysis and phylogeny of the zygomycetous fungal family Mortierellaceae, inferred from nuclear ribosomal DNA sequences. PLoS ONE 2011, 6, e27507. [Google Scholar] [CrossRef]
- Wagner, L.; Stielow, B.; Hoffmann, K.; Petkovits, T.; Papp, T.; Vágvölgyi, C.; de Hoog, G.S.; Verkley, G.; Voigt, K. A comprehensive molecular phylogeny of the Mortierellales (Mortierellomycotina) based on nuclear ribosomal DNA. Persoonia 2013, 30, 77–93. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.V.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 6213. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, Z.; Lv, X.; Li, Y.; Li, Z. High-throughput sequencing reveals the diversity and community structure of rhizosphere fungi of Ferula sinkiangensis at different soil depths. Sci. Rep. 2019, 9, 6558. [Google Scholar] [CrossRef]
- Kuhnert, R.; Oberkofler, I.; Peintner, U. Fungal growth and biomass development is boosted by plants in snow-covered soil. Microb. Ecol. 2012, 64, 79–90. [Google Scholar] [CrossRef]
- Holland, H.L. Biotransformation of organic sulfides. Nat. Prod. Rep. 2001, 18, 171–181. [Google Scholar] [CrossRef]
- Higashiyama, K.; Fujikawa, S.; Park, E.Y.; Shimizu, S. Production of arachidonic acid by Mortierella fungi. Biotechnol. Bioprocess Eng. 2002, 7, 252–262. [Google Scholar] [CrossRef]
- Yadav, D.R.; Kim, S.W.; Babu, A.G.; Adhikari, M.; Kim, C.; Lee, H.B.; Lee, L.S. First report of Mortierella alpina (Mortierellaceae, Zygomycota) isolated from crop field soil in Korea. Mycobiology 2014, 42, 401–404. [Google Scholar] [CrossRef]
- Singh, A.; Ward, O.P. Production of high yields of arachidonic acid in a fed-batch system by Mortierella alpina ATCC 32222. Appl. Microbiol. Biotechnol. 1997, 48, 1–5. [Google Scholar] [CrossRef]
- Du, Z.Y.; Alvaro, J.; Hyden, B.; Zienkiewicz, K.; Benning, N.; Zienkiewicz, A.; Bonito, G.; Benning, C. Enhancing oil production and harvest by combining the marine alga Nannochloropsis oceanica and the oleaginous fungus Mortierella elongata. Biotechnol. Biofuels 2018, 11, 174. [Google Scholar] [CrossRef]
- Li, F.; Zhang, S.; Wang, Y.; Li, Y.; Li, P.; Chen, L.; Jie, X.; Hu, D.; Feng, B.; Yue, K.; et al. Rare fungus, Mortierella capitata, promotes crop growth by stimulating primary metabolisms related genes and reshaping rhizosphere bacterial community. Soil Biol. Biochem. 2020, 151, 108017. [Google Scholar] [CrossRef]
- Ozimek, E.; Hanaka, A. Mortierella species as the plant growth-prmoting fungi present in the agricultural soils. Agriculture 2021, 11, 7. [Google Scholar] [CrossRef]
- Shemshura, O.N.; Shemsheyeva, Z.N.; Sadanov, A.K.; Lozovicka, B.; Kamzolova, S.V.; Morgunov, L.V. Antifungal potential of organic acids produced by Mortierella alpina. Int. J. Eng. Technol. 2018, 7, 1218–1221. [Google Scholar] [CrossRef]
- Telagathoti, A.; Probst, M.; Mandolini, E.; Peintner, U. Mortierellaceae from subalpine and alpine habitats: New species of Entomortierella, Linnemannia, Mortierella, Podila and Tyroliella gen. nov. Stud. Mycol. 2022, 103, 25–58. [Google Scholar] [CrossRef]
- Zhao, H.; Nie, Y.; Huang, B.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China I: Three new species of Backusella (Backusellaceae, Mucoromycota). MycoKeys 2024, 109, 285–304. [Google Scholar] [CrossRef]
- Tao, M.F.; Ding, Z.Y.; Wang, Y.X.; Zhang, Z.X.; Zhao, H.; Meng, Z.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China II: Three new species of Absidia (Cunninghamellaceae, Mucoromycota) from Hainan Province. MycoKeys 2024, 110, 255–272. [Google Scholar] [CrossRef]
- Wang, Y.X.; Zhao, H.; Jiang, Y.; Liu, X.Y.; Tao, M.F.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China III: Six new species and a new record of Gongronella (Cunninghamellaceae, Mucoromycota). MycoKeys 2024, 110, 287–317. [Google Scholar] [CrossRef]
- Ding, Z.Y.; Ji, X.Y.; Tao, M.F.; Liu, W.X.; Jiang, Y.; Zhao, H.; Meng, Z.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China IV: Four new species of Absidia (Cunninghamellaceae, Mucoromycota). MycoKeys 2025, 117, 267–288. [Google Scholar]
- Ji, X.Y.; Ding, Z.Y.; Nie, Y.; Zhao, H.; Wang, S.; Huang, B.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China V: Five new species of Absidia (Cunninghamellaceae, Mucoromycota). MycoKeys 2025, 116, 91–124. [Google Scholar] [CrossRef]
- Wang, Y.X.; Ding, Z.Y.; Ji, X.Y.; Meng, Z.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China VI: Four Absidia sp. nov. (Mucorales) in Guizhou and Hainan. Microorganisms 2025, 13, 1315. [Google Scholar] [CrossRef]
- Ding, Z.Y.; Tao, M.F.; Ji, X.Y.; Jiang, Y.; Liu, W.X.; Wang, S.; Liu, X.Y. Unveiling species diversity within early-diverging fungi from China VII: Seven new species of Cunninghamella (Cunninghamellaceae, Mucoromycota). J. Fungi 2025, 11, 417. [Google Scholar] [CrossRef]
- Zou, Y.; Hou, J.; Guo, S.; Li, Z.; Stephenson, S.L.; Pavlov, I.N.; Liu, P.; Li, Y. Diversity of dictyostelid cellular slime molds, including two species new to science, in forest soils of Changbai Mountain, China. Microbiol. Spectr. 2022, 10, e02402-22. [Google Scholar] [CrossRef]
- Liu, P.; Zou, Y.; Li, S.; Stephenson, S.L.; Wang, Q.; Li, Y. Two new species of dictyostelid cellular slime molds in high-elevation habitats on the Qinghai-Tibet Plateau, China. Sci. Rep. 2019, 9, 5. [Google Scholar] [CrossRef]
- Zhao, H.; Lv, M.L.; Liu, Z.; Zhang, M.Z.; Wang, Y.N.; Ju, X.; Song, Z.; Ren, L.Y.; Jia, B.S.; Qiao, M.; et al. High-yield oleaginous fungi and high-value microbial lipid resources from Mucoromycota. BioEnerg. Res. 2021, 14, 1196–1206. [Google Scholar] [CrossRef]
- Corry, J.E.L. Rose bengal chloramphenicol (RBC) agar. In Progress in Industrial Microbiology; Elsevier: Amsterdam, The Netherlands, 1995; pp. 431–433. [Google Scholar]
- Jiang, Y.; Zhang, Z.; Zhang, J.; Wang, S.; Zhang, X. Morphological and phylogenetic analyses reveal three new species of Phyllosticta (Botryosphaeriales, Phyllostictaceae) in China. J. Fungi 2024, 10, 7. [Google Scholar] [CrossRef]
- Zhang, Z.X.; Liu, R.Y.; Liu, S.B.; Mu, T.C.; Zhang, X.G.; Xia, J.W. Morphological and phylogenetic analyses reveal two new species of Sporocadaceae from Hainan, China. MycoKeys 2022, 88, 171–192. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, F.J.R.M.; Lee, S.H.; Taylor, L.; Shawe-Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 31, 315–322. [Google Scholar] [CrossRef]
- Hurdeal, V.G.; Jones, E.B.G.; Gentekaki, E. Absidia zygospore (Mucoromycetes), a new species from Nan province, Thailand. Stud. Fungi 2023, 8, 15. [Google Scholar] [CrossRef]
- Stiller, J.W.; Hall, B.D. The origin of red algae: Implications for plastid evolution. Proc. Natl. Acad. Sci. USA 1997, 94, 4520–4525. [Google Scholar] [CrossRef] [PubMed]
- Voigt, K.; Wöstemeyer, J. Reliable amplification of actin genes facilitates deep-level phylogeny. Microbiol. Res. 2000, 155, 179–195. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Nie, Y.; Cai, Y.; Gao, Y.; Yu, D.S.; Wang, Z.M.; Liu, X.Y.; Huang, B. Three new species of Conidiobolus sensu stricto from plant debris in eastern China. MycoKeys 2020, 73, 133–149. [Google Scholar] [CrossRef]
- Nie, Y.; Yu, D.S.; Wang, C.F.; Liu, X.Y.; Huang, B. A taxonomic revision of the genus Conidiobolus (Ancylistaceae, Entomophthorales): Four clades including three new genera. MycoKeys 2020, 66, 55–81. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: New York, NY, USA, 2010; pp. 1–8. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.J. Mortierella species in China. Mycosystema 1992, 5, 23–64. [Google Scholar]
- Vandepol, N.; Liber, J.; Desirò, A.; Na, H.; Kennedy, M.; Barry, K.; Grigoriev, I.V.; Miller, A.N.; O’Donnell, K.; Stajich, J.E.; et al. Resolving the Mortierellaceae phylogeny through synthesis of multi-gene phylogenetics and phylogenomics. Fungal Divers. 2020, 104, 267–289. [Google Scholar] [CrossRef]
- Wu, F.; Yuan, H.S.; Zhou, L.W.; Yuan, Y.; Cui, B.K.; Dai, Y.C. Polypore diversity in south China. Mycosystema 2020, 39, 653–682. [Google Scholar] [CrossRef]
- Wang, K.; Chen, S.L.; Dai, Y.C.; Jia, Z.F.; Li, T.H.; Liu, T.Z.; Phurbu, D.; Mamut, R.; Sun, G.Y.; Bau, T.; et al. Overview of China’s nomenclature novelties of fungi in the new century (2000–2020). Mycosystema 2021, 40, 822–833. [Google Scholar] [CrossRef]
- Karunarathna, S.C.; Dong, Y.; Karasaki, S.; Tibpromma, S.; Hyde, K.D.; Lumyong, S.; Xu, J.; Sheng, J.; Mortimer, P.E. Discovery of novel fungal species and pathogens on bat carcasses in a cave in Yunnan Province, China. Emerg. Microbes Infect. 2020, 9, 1554–1566. [Google Scholar] [CrossRef] [PubMed]
- Edgington, S.; Thompson, E.; Moore, D.; Hughes, K.A.; Bridge, P. Investigating the insecticidal potential of Geomyces (Myxotrichaceae: Helotiales) and Mortierella (Mortierellacea: Mortierellales) isolated from Antarctica. SpringerPlus 2014, 3, 289. [Google Scholar] [CrossRef]
- Nam, B.; Lee, H.B.; Choi, Y.J. Three unreported fungi isolated from reservoirs in Korea: Mortierella biramosa, Paraphoma radicina, and Sordaria macrospora. Korean J. Mycol. 2022, 50, 113. [Google Scholar] [CrossRef]
- Yuan, H.S.; Lu, X.; Dai, Y.C.; Hyde, K.D.; Kan, Y.H.; Kušan, I.; He, S.H.; Liu, N.G.; Sarma, V.V.; Zhao, C.L.; et al. Fungal diversity notes 1277–1386: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2020, 104, 1–266. [Google Scholar] [CrossRef]
- Gams, W.; Hooghiemstra, H. Mortierella turficola Ling Yong. Persoonia 1976, 9, 141–156. [Google Scholar]
- Yadav, D.R.; Adhikari, M.; Um, Y.H.; Kim, H.S.; Kim, C.; Lee, H.B.; Lee, Y.S.; Kim, S.W. Three New Records of Mortierella Species Isolated from Crop Field Soil in Korea. Mycobiology 2015, 43, 203–209. [Google Scholar] [CrossRef] [PubMed]
- University of Adelaide. Mortierella wolfii|Mycology. The University of Adelaide. Available online: https://www.adelaide.edu.au/mycology/fungal-descriptions-and-antifungal-susceptibility/zygomycota-pin-moulds/mortierella-wolfii (accessed on 30 May 2025).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Loci | PCR Primers | Primer Sequence (5′–3′) | PCR Cycles | References |
|---|---|---|---|---|
| ITS | ITS5 | GGA AGT AAA AGT CGT AAC AAG G | 95 °C 5 min; (95 °C: 30 s, 55 °C: 30 s, 72 °C: 1 min) × 35 cycles; 72 °C 10 min | [32] |
| ITS4 | TCC TCC GCT TAT TGA TAT GC | |||
| LSU | LR0R | GTA CCC GCT GAA CTT AAG C | 95 °C 5 min; (94 °C: 30 s, 52 °C: 45 s, 72 °C: 1.5 min) × 30 cycles; 72 °C 10 min | [33] |
| LR5 | TCC TGA GGG AAA CTT CG | |||
| RPB1 | RPB1-Af | GAR TGY CCD GGD CAY TTY GG | 95 °C 3 min; (94 °C: 40 s, 60 °C: 40 s, 72 °C: 2 min) × 9 (94 °C: 45 s, 55 °C: 1.5 min, 72 °C: 2 min) × 37 cycles; 72 °C 10 min | [34] |
| RPB1-Cr | CCN GCD ATN TCR TTR TCC ATR TA | |||
| Act | ACT-1 | TGG GAC GAT ATG GAI AAI ATC TGG CA | 95 °C 3 min; (95 °C: 60 s, 55 °C: 60 s, 72 °C: 1 min) × 30 cycles; 72 °C 10 min | [35] |
| ACT-4R | TC ITC GTA TIC TIG CTI IGA IAT CCA CA T | |||
| SSU | NS1 | GTA GTC ATA TGC TTG TCT CC | 95 °C, 5 min; (94 °C: 60 s, 54 °C: 50 s, 72 °C: 1 min) × 37 cycles; 72 °C 10 min | [32] |
| NS4 | CTT CCG TCA ATT CCT TTA AG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, X.-Y.; Jiang, Y.; Li, F.; Ding, Z.-Y.; Meng, Z.; Liu, X.-Y. Unveiling Species Diversity Within Early-Diverging Fungi from China VIII: Four New Species in Mortierellaceae (Mortierellomycota). Microorganisms 2025, 13, 1330. https://doi.org/10.3390/microorganisms13061330
Ji X-Y, Jiang Y, Li F, Ding Z-Y, Meng Z, Liu X-Y. Unveiling Species Diversity Within Early-Diverging Fungi from China VIII: Four New Species in Mortierellaceae (Mortierellomycota). Microorganisms. 2025; 13(6):1330. https://doi.org/10.3390/microorganisms13061330
Chicago/Turabian StyleJi, Xin-Yu, Yang Jiang, Fei Li, Zi-Ying Ding, Zhe Meng, and Xiao-Yong Liu. 2025. "Unveiling Species Diversity Within Early-Diverging Fungi from China VIII: Four New Species in Mortierellaceae (Mortierellomycota)" Microorganisms 13, no. 6: 1330. https://doi.org/10.3390/microorganisms13061330
APA StyleJi, X.-Y., Jiang, Y., Li, F., Ding, Z.-Y., Meng, Z., & Liu, X.-Y. (2025). Unveiling Species Diversity Within Early-Diverging Fungi from China VIII: Four New Species in Mortierellaceae (Mortierellomycota). Microorganisms, 13(6), 1330. https://doi.org/10.3390/microorganisms13061330