Abstract
This study evaluated the oenological potential of two non-Saccharomyces yeast strains, Hanseniaspora opuntiae TR-5 and Zygosaccharomyces bailii L-25, isolated from extreme winemaking environments in southern Spain. Out of 156 yeast isolates screened from high-sugar musts and flor yeast biofilms, strains were selected based on their β-glucosidase activity, killer phenotype, and ethanol production, traits associated with aroma release and microbial competition. Fermentation trials on sugar-rich synthetic medium showed that both H. opuntiae and Z. bailii achieved ethanol yields of 10% v/v and residual sugars at 4 g/L. Co-culture and sequential inoculation, with H. opuntiae introduced first and Z. bailii added on day four, resulted in complete alcoholic fermentation and a reduction in undesirable acetoin levels compared to single-strain fermentations. These findings highlight the practical potential of using selected non-Saccharomyces strains in sequential fermentations to improve aroma complexity, fermentation reliability, and sensory quality in wines, even in the absence of Saccharomyces cerevisiae. The application of these strains offers a novel approach for precision oenology and varietal expression in challenging musts.
1. Introduction
The variability of the microbiota involved in wine fermentation can influence the stability and quality of the wine [1]. Yeasts have an important role during the fermentation of the must by carrying out alcoholic fermentation. However, not all species that make up the wine microbiome contribute pleasant aromas, and this poses a challenge for oenology. The participation of non-Saccharomyces yeasts in winemaking can produce a complexity of compounds, including esters, alcohols, and acids, compared to pure Saccharomyces fermentations [2,3]. Therefore, the selection of yeast strains used in the fermentation process is crucial to improve the fermentation conditions. This is why the need for precision in yeast selection is so important. In fact, non-Saccharomyces yeasts have traditionally been regarded as undesirable in winemaking due to their limited fermentative capacity and association with spoilage, with Brettanomyces and Pichia serving as classic examples. Brettanomyces is notorious for producing off-flavors such as barnyard, medicinal, or phenolic notes, which can compromise wine quality even at low concentrations. Similarly, certain Pichia species are linked to the development of surface films and the production of volatile phenols and ethyl acetate, leading to sensory defects and diminished wine stability [4]. However, the role of non-Saccharomyces yeasts in oenology is increasingly recognized as dual and context-dependent. While some species and strains can cause spoilage, others contribute positively to fermentation and wine complexity. Recent research has demonstrated that selected non-Saccharomyces yeasts can enhance the aromatic profile, mouthfeel, and microbial stability of wines. For example, genera such as Hanseniaspora, Lachancea, and Zygosaccharomyces have been shown to release varietal aroma compounds, produce desirable esters, and even inhibit spoilage organisms through killer activity or competition for nutrients. However, there are now many studies that have shown that non-conventional yeasts have positive effects during winemaking from different grape musts. Furthermore, wine obtained from spontaneous fermentation is superior to simple Saccharomyces cerevisiae fermentation in complexity, body, aroma, and terroir characteristics [5]. Aroma plays a fundamental role in wine selection and consumer acceptance. The aromatic profile or volatilome of wine is composed of several odorants, influencing its flavor [6].
Studies have confirmed that the genus Hanseniaspora can positively influence the color, flavor, aroma, and stability of wines [7,8,9]. Within this genus, Hanseniaspora uvarum is one of the most predominant species, which is one of the most predominant non-Saccharomyces yeast species found in grapes [10]. This species contributes to the natural aroma of wine when used in mixed fermentations. However, if this yeast is not present during fermentation, the resulting wine will lack aromatic complexity, as observed in the study conducted by Rementeria et al. [11]. On the other hand, the Hanseniaspora opuntiae species has a fermentative power of less than 6% v/v; however, it has been proposed that this yeast be used in mixed fermentations with Lachancea thermotolerans, as it may improve the freshness and fruitiness of wines [12].
Zygosaccharomyces bailii is recognized as an osmotolerant yeast with a high fermentative power. This yeast strain can grow under conditions of ≥18% v/v ethanol. The study carried out by Santos et al. [13] showed that this strain was able to metabolize 25 g/L of fructose, a characteristic that would be advantageous to restart or terminate fermentation [14]. Some selected strains of Zygosaccharomyces spp. could be useful because they can produce high levels of ethyl esters [15], such as in the study carried out by Garavaglia et al. [16] in Chardonnay wine. Furthermore, these strains can produce high levels of 2-phenylethyl acetate, isoamyl acetate, ethyl acetate, and ethyl esters, the latter being very important, since they contribute fruity and floral aromatic notes to the wine [16]. Solieri [17] describes this species as promising to produce lactic acid and ethanol. Furthermore, Z. bailii strains that have killer activity produce a toxin known as “zygocin”, a toxin encoded by dsRNA. This study aimed to characterize the oenological potential of H. opuntiae and Z. bailii yeast strains, evaluating their contributions to improving wine quality during fermentation processes. By analyzing their metabolic profiles and fermentative behavior, the research aimed to discover how these yeast strains could improve key oenological attributes such as aroma complexity, microbial stability, and overall sensory characteristics of wines. Our initial hypothesis is that strains adapted to extreme environments (high ethanol and sugar content) may exhibit greater fermentation and a better contribution to aroma. To this end, strains from extremophile environments were isolated and characterized. From the strains selected based on the enzyme parameters analyzed, the fermentative potential of different non-Saccharomyces yeasts present in hostile natural environments was evaluated. These include very sugary musts from the Montilla-Moriles Protected Designation of Origin (PDO), a warm-climate region, characterized by high sugar concentrations and where the predominant non-Saccharomyces yeast species are in a sugar-rich environment, and therefore adapted to these, allowing them to survive the initial concentrations of the medium used for this oenological characterization. Yeasts from the biologically aged biofilms/veils wines from the Jerez–Xérès–Sherry PDO, characterized by high alcohol concentrations, where the predominant species can complete the fermentation of these sugars, and, therefore, complete the fermentation. The Montilla-Moriles and Sherry PDOs were chosen for this study because they represent extreme and contrasting oenological environments—Montilla-Moriles is characterized by musts with high sugar concentrations, while Jerez–Xérès is known for biologically aged wines with high ethanol content and unique flor yeast biofilms—offering exceptional microbial biodiversity and presenting distinct fermentation challenges that are ideal for isolating and evaluating robust, adaptive non-Saccharomyces yeast strains.
To our knowledge, this is one of the few studies evaluating β-glucosidase and killing the activity of wild yeasts from both extremophile oenological environments, based on two PDOs in Andalusia (Spain), in addition to performing a fermentation analysis of pure and mixed culture strains of the selected species: H. opuntiae TR-5 and Z. baili L-25.
The duality underscores the importance of strain selection and controlled application. Rather than being dismissed solely as spoilage agents, non-Saccharomyces yeasts should be evaluated for both their risks and their potential technological benefits in precision oenology. The present study builds on this perspective by exploring the oenological potential of H. opuntiae and Z. bailii, two non-Saccharomyces species with promising enzymatic activities and fermentative traits, aiming to harness their positive contributions while managing possible drawbacks.
2. Materials and Methods
2.1. Isolation of Autochthonous Wine Yeasts
Wine yeasts were isolated from the two aforementioned wine-growing regions in southern Spain. These yeasts were harvested and isolated using aseptic techniques from natural musts from the Montilla-Moriles PDO region in Córdoba, Spain. In this study, yeasts were isolated during the spontaneous fermentation of these musts with more than 250 g/L sugars, using serial dilutions and different screening methods in the laboratory. The natural must is made from the Pedro Ximénez grape variety. However, the yeasts isolated from the Jerez–Xérès–Sherry PDO (Cádiz, Spain) were harvested from flor yeast biofilms during the biological ageing of Fino wine using a sterile spatula. For this study, the yeasts were isolated using serial dilution and enrichment methods. Sherry PDO Fino wine is made from a different grape variety known as Palomino. The must and the flor yeast were independent sample types.
A total of 156 isolates were obtained from the two areas studied. During the spontaneous fermentation of must from vineyards in the Montilla-Moriles area, a total of 85 yeasts were obtained from the Coop La Union (Montilla, Córdoba, Spain) and the Coop Vitivinícola Aguilar de la Fra (Aguilar de la Frontera, Córdoba, Spain). A total of 71 isolates were obtained from two of the most prominent sherry producers in Jerez: the Williams & Humbert (Jerez, Cádiz, Spain) and Gonzalez Byass wineries (Jerez, Cádiz, Spain). The selection of yeast species from these environments was aimed at comparing strains from opposing difficult environments. The strains isolated from Córdoba came from musts with a high sugar concentration, while the strains from Cadiz came from an environment with a high ethanol concentration (over 15.5% v/v), no fermentable sugars, and a low nitrogen source.
2.2. Culture Media and Enzymatic Screening Procedures
2.2.1. WL Nutrient Agar (Wallerstein Laboratory Medium)
According to the manufacturer’s instructions (Oxoid, Basingstoke, UK), WL and a specific medium were prepared from its dehydrated formulation by dissolving 6% (w/v) of the preparation in distilled water, together with 2% (w/v) of agar. The mixture was autoclaved at 120 °C for 15 min. After being poured onto Petri plates, these were incubated at 28 °C for a period of three days. The resulting medium allowed effective distinction of the different yeast genera, thanks to the colony shape and colorimetric variations observed, thus providing a consistent visual tool for their identification [18]. The WL medium is a selective medium, and thanks to the green-bromocresol indicator, the yeasts, depending on their acidification, acquired a different colorimetry ranging from white to different shades of green, allowing different yeast strains to be selected.
2.2.2. Lysine Medium
For the confirmation of isolates belonging to non-Saccharomyces yeasts, a selective medium prepared according to the procedure described by Alcalá-Jiménez et al. [18] was used. Positive identification was established when microbial growth was observed after two successive subcultures, which was considered a potential verification criterion.
2.2.3. Determination of Yeast Killer Activity
Killer activity was measured using the method described by Ramírez et al. [19]. Yeast was analyzed on methylene blue plates at pH 4.0 according to the method described by Kaiser et al. [20]. Plates were seeded with 100 μL of a culture of the sensitive strain previously grown for 48 h. The strains to be studied for their killer activity were patched from solid cultures and seeded in triplicate on the seeded MB plates. Subsequently, the plates were incubated for 7 days at 21 °C.
2.2.4. Medium for Detecting β-Glucosidase Activity
The enzymatic activity of β-glucosidase was measured qualitatively. It was evaluated as a medium containing 0.5% (w/v) arbutin (Sigma–Aldrich Chemie GmbH, Taufkirchen, Germany), 0.1% (w/v) yeast extract (Oxoid, Basingstoke, United Kingdom), 1% (v/v) ferric chloride solution, and 2% (w/v) agar (PanReac Castellar del Vallès, Barcelona, Spain) [21]. This medium was autoclaved at 120 °C for 15 min. The cultures were incubated at 28 °C for 14 days. This medium is based on the hydrolysis of arbutin through the action of β-glucosidase activity.
2.2.5. YPD Agar Medium
Yeast extract peptone dextrose (YPD) agar medium is composed of 1% (w/v) yeast extract (Oxoid, Basingstoke, UK), 2% (w/v) peptone (Oxoid, Basingstoke, UK), 2% (w/v) glucose (Oxoid, Basingstoke, United Kingdom), and 2% (PanReac Castellar del Vallès, Barcelona, Spain) (w/v) agar. It is the most widely used medium for developing and maintaining yeast growth, since it is rich in amino acids, nucleotide precursors, vitamins, and essential metabolites necessary for optimal cell growth.
In the alcoholic fermentation experiments, a modified version of the YPD medium was employed, but features a notable adjustment: it incorporates a high sugar concentration of 20%, equally divided between glucose and fructose, to simulate the high sugar concentration of must.
2.3. Yeast Identification
Yeast identification was carried out using Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry (MALDI TOF) (Bruker Daltonics, ultraflextreme, Bremen, Germany) at the Central Research Support Service (SCAI) of the University of Córdoba (UCO).
In the process of preparing the samples for analysis using MALDI-TOF-MS, the yeast was grown in the YPD agar medium used for their short-term preservation, and was dispensed into Eppendorf (Hamburg, Germany) vials, in which 300 μL of Milli-Q water and 900 μL of absolute ethanol (Merck, Darmstadt, Germany) were added.
2.4. Alcoholic Fermentation Tests
As the final phase of the oenological characterization process for wine yeasts, an alcoholic fermentation test was included, the objective of which was to determine their fermentative power. The basis for the selection of these non-Saccharomyces yeasts is their potential to ferment, modulate, and improve the flavor and aroma of wine, due to the presence of β-glucosidase activity. Furthermore, these yeasts could also be used as biological control yeasts, as they exhibit a killer phenotype.
In this study, fermentation tests were conducted with strains of different non-Saccharomyces yeast species, specifically Hanseniaspora opuntiae and Zygosaccharomyces bailii.
The conditions used here were monoculture to determine how they performed alone in a medium with optimal conditions; in co-culture, to determine how they behaved together; and, finally, in sequence, to determine if there was a reduction in ethanol content as in the study carried out by Urtubia et al. [22].
2.4.1. Preparation of Pre-Inocula
The yeast strains were seeded in 100 mL flasks containing liquid YPD. These flasks underwent shaking for two days at 100 rpm to promote the growth of the yeasts selected for the alcoholic fermentation trials. The total number of cells present in the pre-inocula was then calculated using the Neubauer chamber (Paul Marienfeld GmbH&Co.KG, Lauda-Königshofen, Germany) to establish the inoculation volume necessary to correctly inoculate the different yeast strains in the different cultures during the alcoholic fermentation tests.
2.4.2. Fermentation Monitoring
Fermentations were carried out in 250 mL flasks with 150 mL of modified YPD medium; each flask was inoculated with a total of 2 × 106 cells/mL. Three different conditions were used in the fermentation tests, with three biological replicates of each. The first condition was in monoculture of both selected strains, where the fermentative efficiency of each of these strains was tested. The second condition was in co-culture (CO), inoculated together at the beginning of the fermentation, in the same proportion (1:1), and in sequential culture (SQ). In the latter case, H. opuntiae was inoculated first, since this genus of yeast is the first to appear during spontaneous fermentation, and on the fourth day, the Z. bailii strain was inoculated. After inoculation, the flasks were kept in the culture chamber at 21 °C and under constant and slight agitation. Fermentation kinetics were monitored via the weight loss of the different flasks, following the method described by Spedding [23]. Fermentation was considered complete when no weight changes were observed for a couple of days.
2.5. Determination of Microbiological Parameters
The total number of cells present was calculated in the corresponding dilution using the Neubauer chamber (Paul Marienfeld GmbH&Co.KG, Lauda-Königshofen, Germany). To monitor the yeast population present in the different cultures daily, the number of viable cells was analyzed. To perform the study, one mL was first removed from each replicate, and serial dilutions were made. These cells were plated on WL plates to perform viable cell counts. This count was performed after 48 h of incubation at 28 °C.
2.6. Determination of Oenological Parameters
Oenological parameters such as pH, titratable acidity, volatile acidity, and residual sugar content were determined according to the Organisation Internationale de la Vigne et du Vin (OIV), following official European Economic Community (EEC) methods [24]. Ethanol was measured via dichromate oxidation [25].
2.7. Analytical Determinations of Major Aroma Compounds
Major volatile compounds were quantified with a gas chromatograph HP 6890 Series II equipped with a capillary column with molten silica CP-WAX 57 CB (50 m in length, 0.25 mm in internal diameter, and 0.4 μm in coating thickness) and a Flame ionization detector. Chromatographic conditions and sample preparation were those described by Peinado et al. [26]. The identification and quantification of the major volatile compounds were carried out by using standards submitted to the same treatment as the analyzed samples.
2.8. Statistical Analysis
To identify differences between the different strains used, the parameters and aromatic compounds analyzed were subjected to statistical analysis using the Statgraphics Centurion statistical package (v. 16.1.11). All experiments were performed in triplicate. All quantified compounds were subjected to simple ANOVA, and P tests were used to establish homogeneous groups (different letters indicate significant differences at a 95% confidence level); moreover, principal component analysis was conducted.
3. Results and Discussion
3.1. Microbiological Analysis
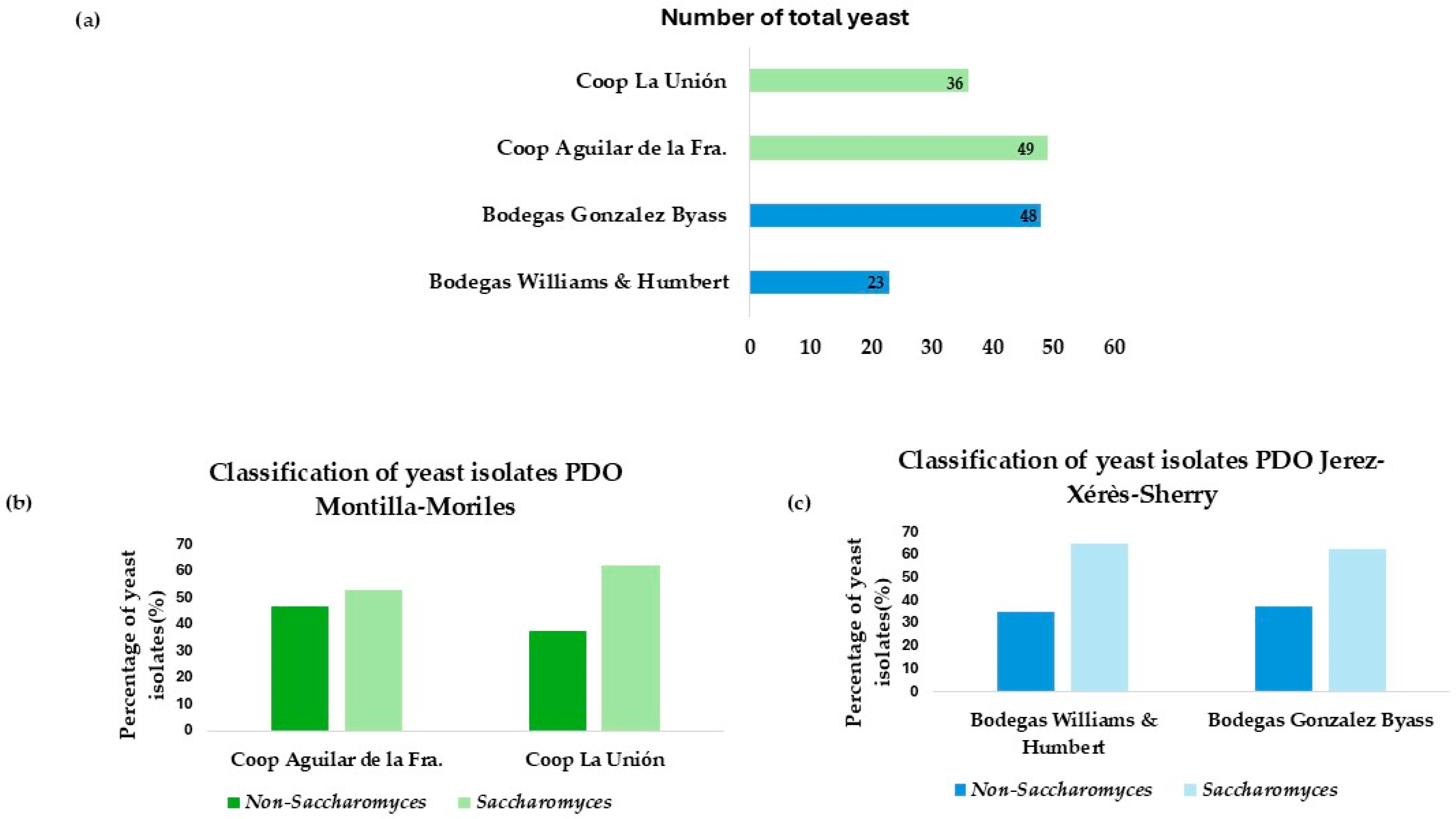
Based on the appearance of the colony in WL and lysine medium, a total of 156 yeasts were isolated from the two sources. These isolates are distributed as shown in Figure 1a.
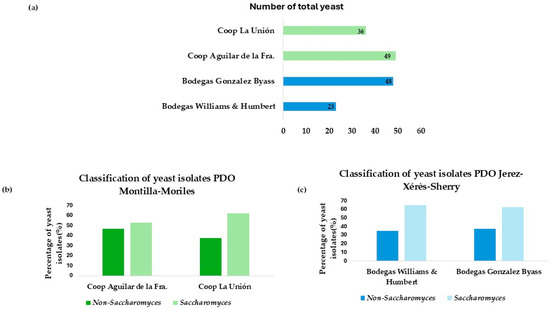
Figure 1.
(a) Total number of isolates differentiated by wineries and cooperatives. (b) Number of isolates from each winery, differentiating between the percentage of Saccharomyces and non-Saccharomyces. (c) Number of isolates from each cooperative, differentiating between the percentage of Saccharomyces and non-Saccharomyces.
A percentage differentiation was made between Saccharomyces and non-Saccharomyces among the different winemaking environments. A higher number of isolates corresponding to the non-Saccharomyces genus was observed in vineyards in the Montilla-Moriles area, as shown in Figure 1b. However, Figure 1c, corresponding to the flor veil isolates from the wineries of Jerez de la Fra., shows that the highest percentage of isolates obtained belong to the Saccharomyces genus. This is because non-Saccharomyces yeasts are those that appear first during spontaneous fermentation, as shown in the study by Miranda et al. [27]. Therefore, there is a higher percentage of this type of yeast in the Montilla-Moriles area, as they are isolated from fermented musts. The isolations obtained in Jerez belong to biologically aged yeasts, where the predominant species is Saccharomyces cerevisiae, as reported in the study by Ruiz-Muñoz et al. [28].
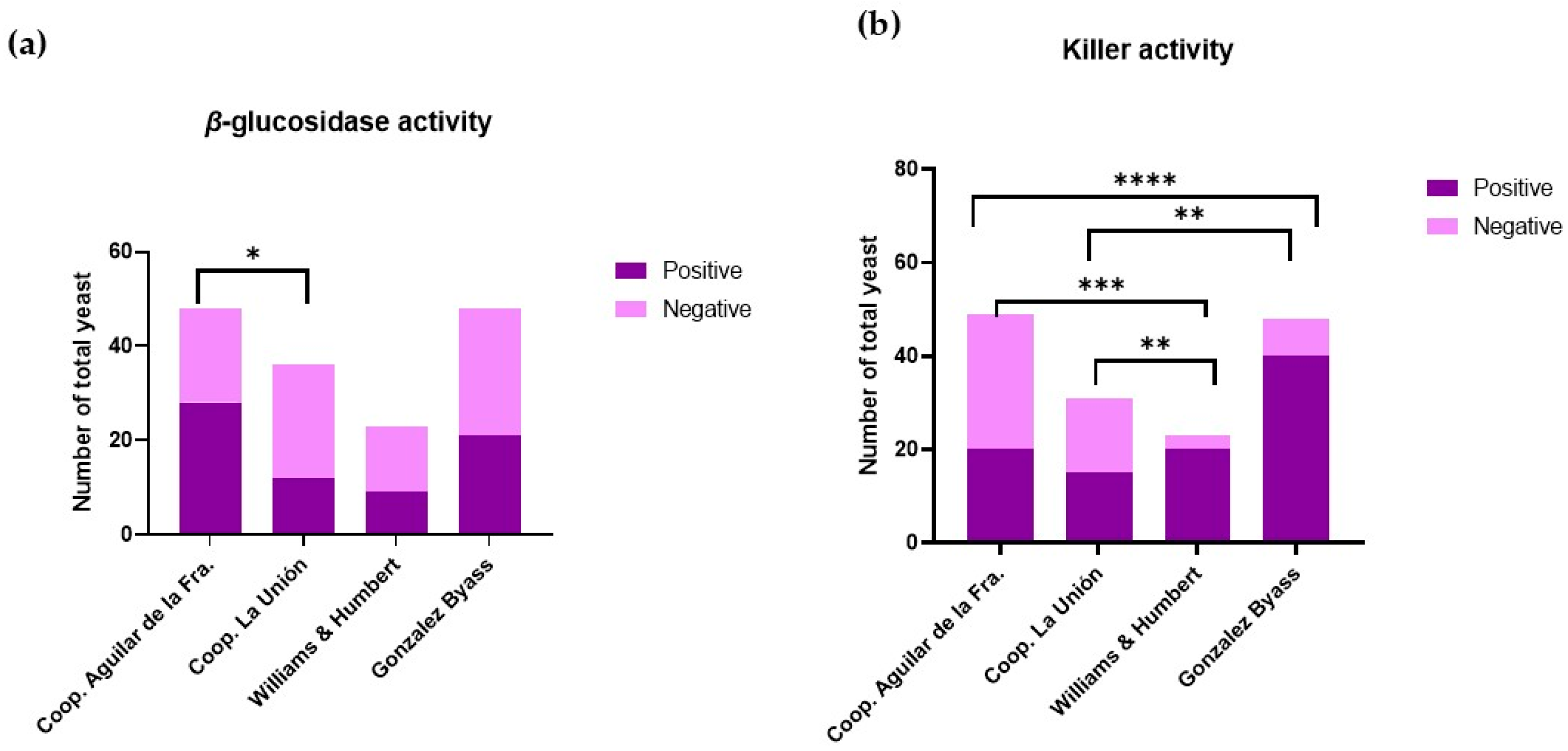
Figure 2a shows the qualitative activity of the β-glucosidase enzyme observed in different viticultural environments. In grapes, some of the aromatic compounds are present as non-volatile glycosidic precursors [29]. β-glucosidase is an enzyme involved in the hydrolysis of substrates with glycosidic bonds, thus releasing glucose and aglycones. This enzyme is responsible for the cleavage of glycosidic bonds, releasing free terpenes, phenylpropenes, and aliphatic esters into the medium [30]. Because of this catalytic activity carried out by β-glucosidase, these glycosides release volatile aromatic compounds, improving the organoleptic properties [31]. This contributes to the development of the varietal aroma and the typicality of wine [32]. Consequently, as can be seen in Figure 2a, it can be concluded that yeasts from both wine-growing areas have a high potential to improve the aromas associated with compounds derived from glycosides.
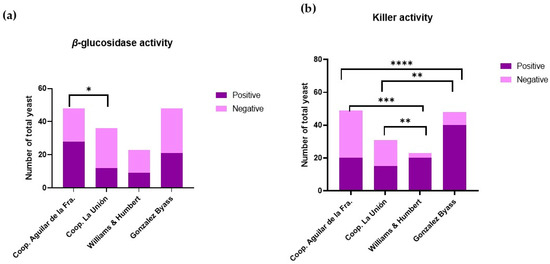
Figure 2.
(a) Absence and presence of β-glucosidase activity in Montilla-Moriles yeast (Córdoba, Spain) and POD Jerez–Xérès–Sherry yeast (Cádiz, Spain). (b) Absence and presence of the killer phenotype in Montilla-Moriles yeast (Córdoba, Spain) and POD Jerez–Xérès–Sherry yeast (Cádiz, Spain). Bar plot with significant differences: * means q < 0.05; ** means q < 0.01; *** means q < 0.001; and **** means q < 0.0001.
Figure 2b shows that yeasts found in different wine-growing environments exhibit a killer phenotype, implying the presence of natural biological control. This activity is associated with changes in population patterns, which influence wine fermentation, as suggested by Englezos et al. [33]. Killer activity is carried out by yeasts that produce “killer substances”. These substances are of protein origin and have an antimicrobial function, inhibiting the growth and development of yeast strains susceptible to these toxins, but not their own producers [34].
3.2. Yeast Identification and Selection
Non-Saccharomyces yeast species found and identified in this study were Hanseniaspora opuntiae (four strains), Lachancea thermotolerans (four strains), Rhodotorula mucilaginosa (one strain), Torulaspora delbrueckii (three strains), Kluyveromyces marxianus (one strain), Hanseniaspora uvarum (one strain), Pichia kudriavzevii (two strains), and Pichia terricola (two strains) in Montilla-Moriles musts and, in Jerez veils, Zygosaccharomyces bailii (two strains) and Torulaspora delbrueckii (five strains). These strains were selected over the rest of the identified species because they presented both microbial properties described in Section 2. For this study, strains were selected that exhibited positive killer and β-glucosidase activity and produced high levels of ethanol in a synthetic medium at concentrations greater than 10% v/v. The selected strains were H. opuntiae TR-5 from the Montilla-Moriles Protected Designation of Origin (PDO) and Z. bailii L-25 from the Sherry PDO.
It should be noted that non-Saccharomyces strains have great potential in winemaking and should not be discarded prematurely. As some characteristics are strain-specific, isolates should not be discarded at the initial stage of research. While some strains can be selected for further studies, the potential value of others can be recognized for future research.
3.3. Fermentation Kinetics
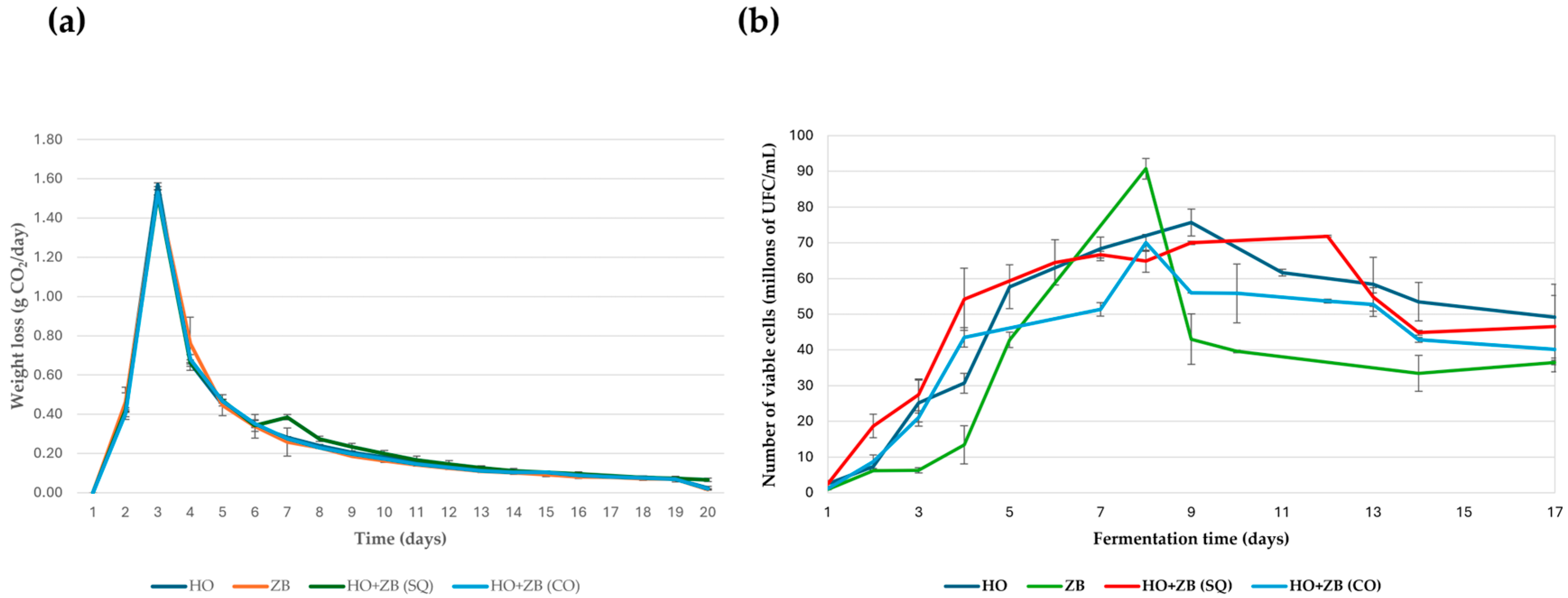
Figure 3a shows the kinetic profiles obtained during the daily monitoring of each of the fermentations carried out with the yeasts H. opuntiae in monoculture (HO) and Z. bailii in monoculture (ZB), as was carried out in the study by Ansanay-Galeote et al. [35] with Saccharomyces cerevisiae. This graph also shows the kinetics of the co-inoculation (CO) and sequential (SQ) fermentations. These presented similar behavior to monoculture fermentations. It is noteworthy that the maximum CO2 release was obtained approximately 24 h after the start of fermentation in both cases. From that moment on, the fermentation rate gradually decreased until it was practically zero, marking the end of the fermentation process for the yeasts used in the trial. In the case of the sequential fermentation, a small change in the kinetic profile was observed on the fourth day due to the inoculation of Z. bailii. In addition to daily monitoring of the kinetic profiles of the different fermentations, the population growth of the different yeast strains was monitored daily during fermentation (Figure 3b). This study of population growth allows us to determine whether there is a relationship between fermentation kinetics and the yeast population present in the medium. In monoculture fermentations inoculated with one of the two yeast strains tested, the Z. bailii strain showed a significantly slower growth and survival rate than the H. opuntiae strain as fermentation progressed, according to the daily total cell count. Similarly, H. opuntiae exhibited higher viability than Z. bailii. On the other hand, in the mixed fermentations, the evolution of the yeast population, expressed in terms of the number of live cells, was very similar in both cases, as was the case in the individual fermentation with H. opuntiae. However, in the sequential fermentation, maximum viability lasted for a longer period rather than showing a prominent peak on a specific day, compared to the other fermentations performed with the H. opuntiae and Z. bailii strains. These results seem very interesting, since they suggest that a sequential fermentation, in which the H. opuntiae strain is inoculated first and then the Z. bailii strain, could be considered a strategy to complete alcoholic fermentation in the absence of Saccharomyces cerevisiae.
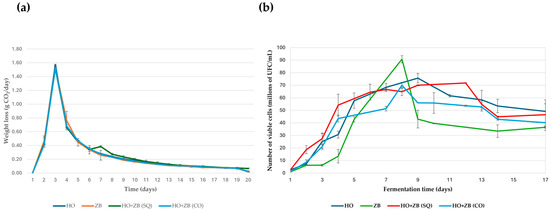
Figure 3.
(a) Kinetic profiles of different fermentations performed with Hanseniaspora opuntiae (HO) and Zygosaccharomyces bailii (ZB) in monoculture. In sequential (SQ), where H. opuntiae was added first, and on the fourth day, Z. bailii was introduced and co-cultured (CO). (b) Viable cell count during the different fermentation conditions performed with Hanseniaspora opuntiae (HO) and Zygosaccharomyces bailii (ZB), as described in the previous figure. The means and standard deviations are represented.
3.4. Oenological Parameters
In Table 1, the values shown represent averages of triplicate samples (data are mean ± standard deviation (SD)).

Table 1.
Oenological parameters.
The two non-Saccharomyces yeast species tested showed similar ethanol contents (≈10–11% v/v) in both single and mixed fermentations. However, the residual sugar concentration was lower in the mixed fermentations, although it never exceeded 5 g/L, which is characteristic of dry wines.
The fact that both yeasts are capable of consuming virtually all the sugars initially present in the culture medium (200 g/L), together with density values below the reference density of water, indicates that both non-Saccharomyces yeast strains are capable of complete alcoholic fermentation, achieving similar and even higher ethanol levels compared to various strains belonging to the Saccharomyces genus [36]. The study conducted by Del Fresno et al. [37] on the use of different yeasts for the production of rosé wines describes a strain of Hanseniaspora vineae capable of achieving, in a single fermentation, an ethanol content very similar to another strain of Saccharomyces cerevisiae (≈13% v/v) and, in addition, it achieves better results in terms of a more attractive color and the organoleptic characteristics of the final product.
Wine quality depends on multiple factors, and one of the most relevant aspects is volatile acidity, whose legal limit is 1.2 g/L for most wines. However, it is advisable to keep this as low as possible because acetic acid can be sensorially detected by the consumer at concentrations above 0.8 g/L and impart an undesirable vinegary flavor and aroma to the final product. Thus, it could be said that, in a winemaking process, it would be ideal to carry out a single fermentation with Z. bailii, mainly due to the fact that the volatile acidity value is below the sensory threshold, contrary to what is observed with H. opuntiae or in mixed fermentations with both yeasts, and the remaining oenological parameters do not differ much in either case.
3.5. Major Aroma and Statistical Analysis
The determined alcohol, except propanol, which has α-ketobutyric acid as its precursor, is derived from the degradation of sugars or amino acids through the decarboxylation of keto acids. These products, except 2-phenylethanol, which possesses a floral aroma, usually have solvent aromas and a higher perception threshold. Their overall concentration must not exceed 400–500 mg/L. Below, these amounts contribute to the aroma complexity of the wine [38].
As can be seen in Table 2, isoamyl alcohols show significant differences under the different conditions assayed. The highest values were observed with H. opuntiae and in sequential culture, whereas the lowest was reached with Z. bailii. The same behavior is observed with isobutanol, and the opposite is true in the case of propanol. 2-Phenylethanol provides sweet and floral aromas, as observed in the study by Gómez-Míguez et al. [39]. In our study, it was observed to exceed the perception threshold (10 mg/L) in all conditions. The lowest values were found in co-inoculation, and the highest were with H. opuntiae and in the sequential assay. The results obtained by Martin et al. [40] show that the concentration of 2-phenylethanol increased more in co-culture, intensifying the floral and sweet attributes of the wine.
Table 2.
Major aroma compounds. The values shown represent averages of triplicate samples (data are mean ± SD). Values with different superscript letters in the same row are significantly different test (p < 0.05).
Among the carbonyl compounds, acetaldehyde is formed by yeasts as an intermediary metabolite in the conversion of glucose into ethanol. Acetaldehyde has a pleasant, fruity aroma at minimal concentrations; however, at high levels, its odor becomes irritating and penetrating [41,42]. Usually, its concentration does not exceed 50 to 70 mg/L [38]. In this study, the amount was higher in all the cases. However, it was observed that the compounds 2,3-butanediol (meso and levo) and glycerol were produced at lower levels in the presence of Z. bailii. This may be due to the lower activity of glyceropyruvic fermentation. On the other hand, acetoin contributes to the development of lactic odors, such as butter. However, it only exceeds the threshold in monocultures; therefore, its use in axenic cultures would have this unique characteristic. However, in mixed cultures, a significant reduction in this compound was observed. This result is supported by the study by Capece et al. [43]. Elevated levels of acetaldehyde and acetoin are likely due to the shift in their metabolism toward glyceropyruvic fermentation to regenerate the NAD+ that could not be obtained through alcoholic fermentation. This would result in a balance in redox potential. At these concentrations, acetaldehyde can contribute to pungent and oxidized apple aromas. The elevated acetaldehyde levels in the different cultures are probably the result of high pyruvate decarboxylase activity in these non-Saccharomyces strains. Acetoin exceeded its perception threshold only in monoculture conditions (36–44 mg/L), contributing buttery notes, whereas mixed cultures showed significantly reduced levels (25–28 mg/L) below sensory relevance. This reduction in mixed cultures represents a potential advantage for the use of combined inoculation strategies.
From the sugars (glucose and fructose) present in the synthetic medium, two different pathways can be carried out. The first, through which glycerol precursors, dihydroxyacetone phosphate (dihydroxyacetone-P), are formed. This is converted into glycerol-3-phosphate (G3P) through the reduction of a molecule of NADH, through the action of the enzyme triose phosphate isomerase. Once this compound is obtained, it is converted into glycerol through the enzymatic action of glycerol-1-phosphatase, which can hydrolyze the phosphate group of G3P, forming glycerol. On the other hand, glyceraldehyde-3-phosphate is formed from glucose. There is a balance between the formation of dihydroxyacetone-P and glyceraldehyde-3-phosphate; therefore, the yeast regulates the pathway it uses to maintain this balance. From glyceraldehyde-3-phosphate, pyruvate is formed through glycolysis, producing one molecule of NADH. From pyruvate, two different pathways can occur simultaneously: First, pyruvate is transformed into acetaldehyde through decarboxylation thanks to pyruvate decarboxylase. Finally, acetaldehyde is reduced by the consumption of NADH and the action of alcohol dehydrogenase to form ethanol. The second option in the pyruvate production pathway involves the condensation of two pyruvate molecules to form α-ketolactate, catalyzed by acetolactate synthase. α-Ketolactate can be converted into several compounds, including the amino acid valine, by the action of acetolactate synthase and diacetyl, and by the action of alpha-acetolactate decarboxylase, thus preventing the formation of acetoin (a compound that imparts an undesirable flavor). When diacetyl is reduced by diacetyl reductase, acetoin (3-hydroxybutanone) is formed, which is subsequently transformed into 2,3-butanediol through a reduction reaction catalyzed by the action of the enzyme butanediol dehydrogenase [44].
This fermentation produces lactic acid, which reacts with the ethanol produced during alcoholic fermentation, where an esterification reaction occurs, forming ethyl lactate. This is also due to the acidity of the medium. As observed in Table 2, the highest values of this compound are found in the monoculture of H. opuntiae and in the co-culture. It is also observed that, in the presence of Z. bailii, the values of this ester tend to decrease, as observed in the sequential culture or monoculture of this strain, which presents the lowest values in all conditions. On the other hand, yeasts are capable of producing acetic acid as a byproduct of their metabolism, which, as mentioned above, reacts with the ethanol and triggers another esterification reaction, forming ethyl acetate Z. bailii, which is an osmotolerant and highly fermentative yeast; little is known about its metabolism under culture conditions or its ability to produce esters. Ethyl esters are very interesting because they have a pleasant fruity and floral aromatic note [45]. In the study conducted by Garavaglia et al. [16], it was shown that the Z. bailii BCV 08 strain was able to increase ester production. However, in our study, the strain used did not increase the production of major esters; rather, in the presence of this yeast, its production decreased. A greater production of this compound was observed in the monoculture of H. opuntiae, this is because apiculated yeasts are good producers of acetate esters, as observed in the work of Rojas et al. [45] and Filippousi et al. [46]. It was concluded that this compound decreased in the presence of Z. bailii, both in co-culture and sequentially. This compound provides fruity aromas, such as pineapple.
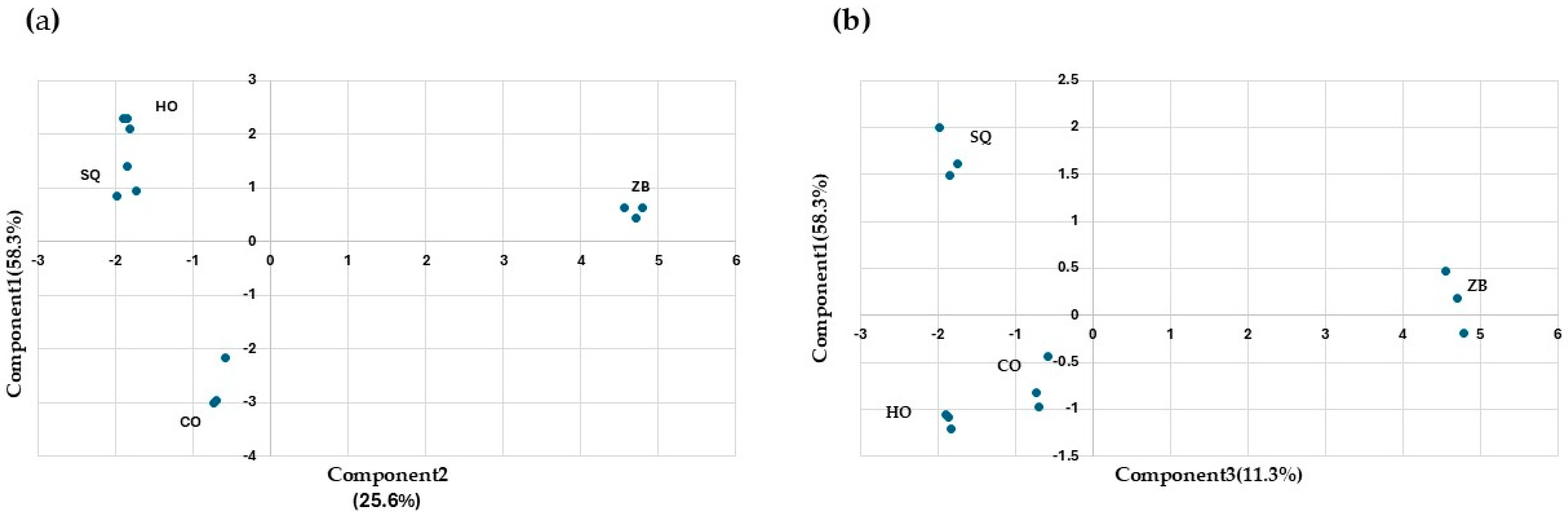
With the aim of reducing the number of variables, a principal component analysis was carried out (Figure 4). Components are a linear combination of the input variables, and the interpretation of the components obtained must be performed by the analyst. Here, the aroma compounds were selected as input variables. The first three components explain 83.4% of the observed variability (Figure 4a,b). Component 1 differentiates the ZB assay from the rest. The compounds with the greatest influence are propanol, isobutanol, and ethyl acetate. Component 2 differentiates co-inoculation from SQ and HO and is mainly influenced by 2,3-butanediol (levo), 2-phenylethanol, and diethyl succinate. Lastly, component 3 differentiates SQ from HO, and the most influential compounds are 2,3-butanediol (meso), acetaldehyde, and ethyl lactate.
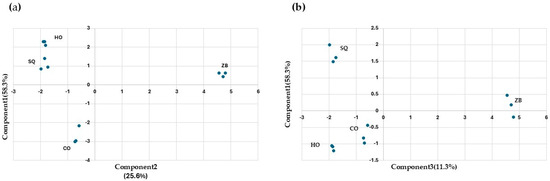
Figure 4.
Principal component analysis of the “wines” obtained after fermentations under different conditions: Hanseniaspora opuntiae (HO) and Zygosaccharomyces bailii (ZB) in monoculture. In sequential (SQ), where H. opuntiae was added first, and on the fourth day, Z. bailii was introduced and co-cultured (CO). The analysis was performed on the major volatile compounds. (a) Principal component analysis between components one and two. (b) Principal component analysis between components one and three.
4. Conclusions
This study demonstrates that non-Saccharomyces yeasts, specifically Hanseniaspora opuntiae and Zygosaccharomyces bailii, isolated from challenging oenological environments, possess significant potential to improve wine quality through their dual β-glucosidase activity and killer phenotype. These traits are crucial for the release of aroma-enhancing compounds and for providing a natural form of microbial competition during fermentation, positioning these strains as valuable alternatives to traditional fermentative yeasts. Our results show that H. opuntiae is particularly effective when applied during the early stages of fermentation, where its β-glucosidase activity enhances the release of fruity and floral aromas, contributing to a more complex and appealing sensory profile. In contrast, Z. bailii excels in completing fermentation, even under high-sugar conditions, and is associated with lower volatile acidity production, which is essential for maintaining wine freshness and avoiding undesirable vinegary notes. Sequential inoculation—introducing H. opuntiae first, followed by Z. bailii—proved especially beneficial. This strategy achieved complete fermentation, reduced residual sugar, and resulted in wines with enhanced aromatic complexity and improved mouthfeel, even in the absence of Saccharomyces cerevisiae. These findings provide practical guidance for winemakers: employing H. opuntiae at the start of fermentation and introducing Z. bailii after several days can optimize both fermentation kinetics and sensory outcomes. Fermentation temperatures around 21 °C and careful timing of inoculation are recommended to maximize the performance of these strains. Importantly, this work challenges the traditional view of these non-Saccharomyces species as spoilage organisms. Instead, it highlights their suitability as bio-tools for precision oenology, capable of enhancing varietal expression and contributing to the development of wines with distinctive sensory attributes. The application of these strains in commercial winemaking could offer new avenues for aroma enhancement, microbial control, and adaptation to musts with high sugar or ethanol content.
Future studies will focus on validating these findings in real grape must fermentations and further exploring the sensory and technological impacts of these yeasts under commercial winemaking conditions.
Author Contributions
Conceptualization, J.C.M., J.M., R.P. and T.G.-M.; methodology, M.T.A.-J., J.C.G.-G., R.P. and T.G.-M.; software, R.P.; validation, R.P. and T.G.-M.; formal analysis, M.T.A.-J. and J.C.G.-G. investigation, M.T.A.-J. and J.C.G.-G.; resources, J.C.M., J.M., R.P. and T.G.-M.; data curation, M.T.A.-J. and J.C.G.-G.; writing—original draft preparation, M.T.A.-J. and J.C.G.-G.; writing—review and editing, J.C.M., R.P. and T.G.-M.; visualization, J.C.M. and J.M.; supervision, J.M., R.P. and T.G.-M.; project administration, J.C.M. and J.M.; funding acquisition, J.C.M. and J.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Junta Andalucia (Spain) Program: Specialized Transfer Actions Universities–CEI-RIS3-FEDER. Singular Project AGROMIS-ceiA3, Subsector: Traditional Wines of Andalusia. Call 2020: Grants for R+D+I projects in the field of Innovation Ecosystems of the International Centers of Excellence (PY2020 ECOSIS INNOV CEIS)—grant number PYC20 RE 068 UCO. And also, this study was co-funded by the Spanish Ministry of Science, Innovation, and Universities (MICIU/European Union FEDER), Ref. PID 2021-127766OB-I00 (JCM).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors would like to thank the staff and management of the wineries involved. Gonzalez Byass, Williams & Humbert and Coop. La Unión and Coop. Aguilar de la Frontera. They would also like to thank the staff at the Central Research Support Service (SCAI) of the University of Córdoba for their help with identifying the yeast using the MALDI-Biotyper.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wu, Z.Q.; Jiao, M.L.; Li, D.M.; Zhang, H.X.; Xie, Y.H.; Li, H.J.; Yang, M.-M.; Duan, X.-R.; Pang, X.-N.; Fan, X.-M. Microbial diversity during wine fermentation in Beijing region and its influence on wine flavor. LWT 2025, 215, 117209. [Google Scholar] [CrossRef]
- Gao, J.; Wang, M.; Huang, W.; You, Y.; Zhan, J. Indigenous Saccharomyces cerevisiae could better adapt to the physicochemical conditions and natural microbial ecology of prince grape must compared with commercial Saccharomyces cerevisiae FX10. Molecules 2022, 27, 6892. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Liu, B.; Maimaitiyiming, R.; Wang, L.; Zhao, L.; Zhang, H.; Chen, K.; Aihaiti, A. The Effect of Non-Saccharomyces Cerevisiae Torulaspora delbrueckii on the aroma composition of munage grape base-wine and the mechanism of the effect. Fermentation 2024, 10, 266. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled mixed culture fermentation: A new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef]
- Tofalo, R.; Perpetuini, G.; Fasoli, G.; Schirone, M.; Corsetti, A.; Suzzi, G. Biodiversity study of wine yeasts belonging to the “terroir” of Montepulciano d’Abruzzo “Colline Teramane” revealed Saccharomyces cerevisiae strains exhibiting atypical and unique 5.8S-ITS restriction patterns. Food Microbiol. 2014, 39, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, G.; Guan, X.; Li, A.; Tao, Y. Volatile aroma compound-based decoding and prediction of sweet berry aromas in dry red wine. Food Chem. 2025, 463, 141248. [Google Scholar] [CrossRef]
- Lombardi, S.J.; Pannella, G.; Iorizzo, M.; Moreno-Arribas, M.V.; Tremonte, P.; Succi, M.; Sorrentino, E.; Macciola, V.; Di Renzo, M.; Coppola, R. Sequential inoculum of Hanseniaspora Guilliermondii and Saccharomyces Cerevisiae for winemaking campanino on an industrial Scale. World J. Microbiol. Biotechnol. 2018, 34, 161. [Google Scholar] [CrossRef]
- Wang, L.; Dou, G.; Guo, H.; Zhang, Q.; Qin, X.; Yu, W.; Jiang, C.; Xiao, H. Volatile organic compounds of Hanseniaspora Uvarum increase strawberry fruit flavor and defense during cold storage. Food Sci. Nutr. 2019, 7, 2625–2635. [Google Scholar] [CrossRef]
- Capozzi, V.; Berbegal, C.; Tufariello, M.; Grieco, F.; Spano, G. Impact of co-inoculation of Saccharomyces Cerevisiae, Hanseniaspora Uvarum and Oenococcus Oeni autochthonous strains in controlled multi starter grape must Fermentations. LWT 2019, 109, 241–249. [Google Scholar] [CrossRef]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Rementeria, A.; Rodriguez, J.A.; Cadaval, A.; Amenabar, R.; Muguruza, J.R.; Hernando, F.L.; Sevilla, M.J. Yeast associated with spontaneous fermentations of white wines from the “Txakoli de Bizkaia” region (Basque Country, North Spain). Int. J. Food Microbiol. 2003, 86, 201–207. [Google Scholar] [CrossRef]
- Morata, A.; Escott, C.; Loira, I.; Manuel Del Fresno, J.M.; Vaquero, C.; Antonia Bañuelos, M.; Palomero, F.; López, C.; González, C. pH Control and Aroma Improvement Using the Non-Saccharomyces Lachancea thermotolerans and Hanseniaspora spp. Yeasts to improve wine freshness in warm areas. In Grapes Wine; Intechopen: London, UK, 2022. [Google Scholar] [CrossRef]
- Santos, J.; Sousa, M.J.; Cardoso, H.; Inacio, J.; Silva, S.; Spencer-Martins, I.; Leao, C. Ethanol tolerance of sugar transport, and the rectification of stuck wine fermentations. Microbiology 2008, 154, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Zuehlke, J.M.; Childs, B.C.; Edwards, C.G. Evaluation of Zygosaccharomyces bailii to metabolize residual sugar present in partially-fermented red wines. Fermentation 2015, 1, 3–12. [Google Scholar] [CrossRef]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial resources and enological significance: Opportunities and benefits. Front. Microbiol 2017, 8, 995. [Google Scholar] [CrossRef] [PubMed]
- Garavaglia, J.; de Souza Schneider RD, C.; Mendes SD, C.; Welke, J.E.; Zini, C.A.; Caramão, E.B.; Valente, P. Evaluation of Zygosaccharomyces bailii BCV 08 as a co-starter in wine fermentation for the improvement of ethyl esters production. Microbial. Res. 2015, 173, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Solieri, L. The revenge of Zygosaccharomyces yeasts in food biotechnology and applied microbiology. World J. Microbiol. Biotechnol. 2021, 37, 96. [Google Scholar] [CrossRef]
- Alcalá-Jiménez, M.T.; García-Martínez, T.; Mauricio, J.C.; Moreno, J.; Peinado, R.A. Influence of terroir on microbial diversity and wine volatilome. Appl. Sci. 2025, 15, 3237. [Google Scholar] [CrossRef]
- Ramírez, M.; Vinagre, A.; Ambrona, J.; Molina, F.; Maqueda, M.; Rebollo, J.E. Genetic instability of heterozygous hybrid populations of natural wine yeasts. Appl. Environ. Microbiol. 2004, 70, 4686–4691. [Google Scholar] [CrossRef]
- Kaiser, C.; Michaelis, S.; Mitchell, A. Methods in Yeast Genetics; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1994. [Google Scholar]
- Lodder, J.; Kreger-van Rij, N.J.W. The Yeasts: A Taxonomic Study, 1st ed.; Elsevier: Cham, The Netherlands, 1952; pp. 84–85. [Google Scholar]
- Urtubia, A.; Franco, W.; De Lecco, C.C.; Benavides, S.; Durán, A. Impact of non-Saccharomyces autochthonous yeasts on ethanol reduction and chemical profile of Chilean Sauvignon Blanc wine. In Proceedings of the BIO Web of Conferences, Jember, Indonesia, 12–13 September 2023; EDP Sciences: Ulis, France, 2023; Volume 56, p. 02018. [Google Scholar]
- Spedding, G. Alcohol and its measurement. In Brewing Materials and Processes; Academic Press: Cambridge, MA, USA, 2016; pp. 123–149. [Google Scholar] [CrossRef]
- International Code of Oenological Practices; OIV International Organisation of Vine and Wine: Dijon, France, 2023.
- Pilone, G.J. Determination of ethanol in wine by titrimetric and spectrophotometric dichromate methods: Collaborative study. J. AOAC Int. 1985, 68, 188–190. [Google Scholar] [CrossRef]
- Peinado, R.A.; Moreno, J.A.; Muñoz, D.; Medina, M.; Moreno, J. Gas chromatographic quantification of major volatile compounds and polyols in wine by direct injection. J. Agric. Food Chem. 2004, 52, 6389–6393. [Google Scholar] [CrossRef]
- Miranda, A.; Pereira, V.; Jardim, H.; Malfeito-Ferreira, M.; Marques, J.C. Impact of Non-Saccharomyces yeast fermentation in madeira wine chemical composition. Processes 2023, 11, 482. [Google Scholar] [CrossRef]
- Ruiz-Muñoz, M.; Hernández-Fernández, M.; Cordero-Bueso, G.; Martínez-Verdugo, S.; Pérez, F.; Cantoral, J.M. Non-Saccharomyces are also forming the veil of flor insherry wines. Fermentation 2022, 8, 456. [Google Scholar] [CrossRef]
- de Ovalle, S.; Cavello, I.; Brena, B.M.; Cavalitto, S.; González-Pombo, P. Production and characterization of a β-glucosidase from Issatchenkia terricola and its use for hydrolysis of aromatic precursors in Cabernet Sauvignon wine. LWT 2018, 87, 515–522. [Google Scholar] [CrossRef]
- Michlmayr, H.; Kneifel, W.; Kneifel, W. β-Glucosidase activities of lactic acid bacteria: Mechanisms, impact on fermented food and human health. FEMS Microbiol. Lett. 2014, 352, 1–10. [Google Scholar] [CrossRef]
- Liang, Z.; Fang, Z.; Pai, A.; Luo, J.; Gan, R.; Gao, Y.; Lu, J.; Zhang, P. Glycosidically bound aroma precursors in fruits: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2022, 62, 215–243. [Google Scholar] [CrossRef] [PubMed]
- Romo-Sánchez, S.; Arévalo-Villena, M.; García Romero, E.; Ramirez, H.L.; Briones Pérez, A. Immobilization of β-glucosidase and its application for enhancement of aroma precursors in muscat wine. Food Bioprocess Technol. 2014, 7, 1381–1392. [Google Scholar] [CrossRef]
- Englezos, V.; Jolly, N.P.; Di Gianvito, P.; Rantsiou, K.; Cocolin, L. Microbial interactions in winemaking: Ecological aspects and effect on wine quality. Trends Food Sci. Technol. 2022, 127, 99–113. [Google Scholar] [CrossRef]
- Lowes, K.F.; Shearman, C.A.; Payne, J.; MacKenzie, D.; Archer, D.B.; Merry, R.J.; Gasson, M.J. Prevention of yeast spoilage in feed and food by the yeast mycocin HMK. Appl. Environ. Microbiol. 2000, 66, 1066–1076. [Google Scholar] [CrossRef]
- Ansanay-Galeote, V.; Blondin, B.; Dequin, S.; Sablayrolles, J.-M. Stress effect of ethanol on fermentation kinetics by stationary-phase cells of Saccharomyces cerevisiae. Biotechnol. Lett. 2001, 23, 677–681. [Google Scholar] [CrossRef]
- Sottil, C.; Salor-Torregrosa, J.M.; Moreno-Garcia, J.; Peinado, J.; Mauricio, J.C.; Moreno, J.; Garcia-Martinez, T. Using Torulaspora delbrueckii, Saccharomyces cerevisiae and Saccharomyces bayanus wine yeasts as starter cultures for fermentation and quality improvement of mead. Eur. Food Res. Technol. 2019, 245, 2705–2714. [Google Scholar] [CrossRef]
- Del Fresno, J.M.; Loira, I.; Escott, C.; Carrau, F.; González, C.; Cuerda, R.; Morata, A. Application of Hanseniaspora vineae yeast in the production of rosé wines from a blend of Tempranillo and Albillo grapes. Fermentation 2021, 7, 141. [Google Scholar] [CrossRef]
- Moreno, J.; Peinado, R. Enological Chemistry; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Gómez-Míguez, M.J.; Cacho, J.F.; Ferreira, V.; Vicario, I.M.; Heredia, F.J. Volatile components of zalema white wines. Food Chem. 2007, 100, 1464–1473. [Google Scholar] [CrossRef]
- Martin, V.; Valera, M.J.; Medina, K.; Boido, E.; Carrau, F. Oenological impact of the Hanseniaspora/Kloeckera yeast genus on wines—A review. Fermentation 2018, 4, 76. [Google Scholar] [CrossRef]
- Guittin, C.; Maçna, F.; Picou, C.; Perez, M.; Barreau, A.; Poitou, X.; Sablayrolles, J.-M.; Mouret, J.-R.; Farines, V. New online monitoring approaches to describe and understand the kinetics of acetaldehyde concentration during wine alcoholic fermentation: Access to production balances. Fermentation 2023, 9, 299. [Google Scholar] [CrossRef]
- Wang, X.; Fan, G.; Peng, Y.; Xu, N.; Xie, Y.; Zhou, H.; Matallana, E.; Siesto, G.; Aranda, A. Mechanisms and effects of non-Saccharomyces yeast fermentation on the aromatic profile of wine. J. Food Compos. Anal. 2023, 124, 105660. [Google Scholar] [CrossRef]
- Capece, A.; Pietrafesa, A.; Pietrafesa, R.; Garrigós, V.; Tedesco, F.; Romano, P.; Matallana, E.; Siesto, G.; Aranda, A. Impact of Starmerella bacillaris and Zygosaccharomyces bailii on ethanol reduction and Saccharomyces cerevisiae metabolism during mixed wine fermentations. Food Res. Int. 2022, 159, 111649. [Google Scholar] [CrossRef]
- Epifanio, S. La Influencia de la Tecnología de la Vinificación en la Microbiología y el Desarrollo de la Fermentación Alcohólica. Ph.D. Thesis, Universidad de la Rioja, Logroño, Spain, 2005. [Google Scholar]
- Rojas, V.; Gil, J.V.; Piñaga, F.; Manzanares, P. Acetate ester formation in wine by mixed cultures in laboratory fermentations. Int. J. Food Microbiol. 2003, 86, 181–188. [Google Scholar] [CrossRef]
- Filippousi, M.E.; Chalvantzi, I.; Mallouchos, A.; Marmaras, I.; Banilas, G.; Nisiotou, A. The use of Hanseniaspora opuntiae to improve ‘Sideritis’ wine quality, a late-ripening Greek grape variety. Foods 2024, 13, 1061. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).