
Host-Associated Biofilms: Vibrio fischeri and Other Symbiotic Bacteria Within the Vibrionaceae


Abstract
1. Introduction
2. Biofilms: Essential Structures in Symbiosis
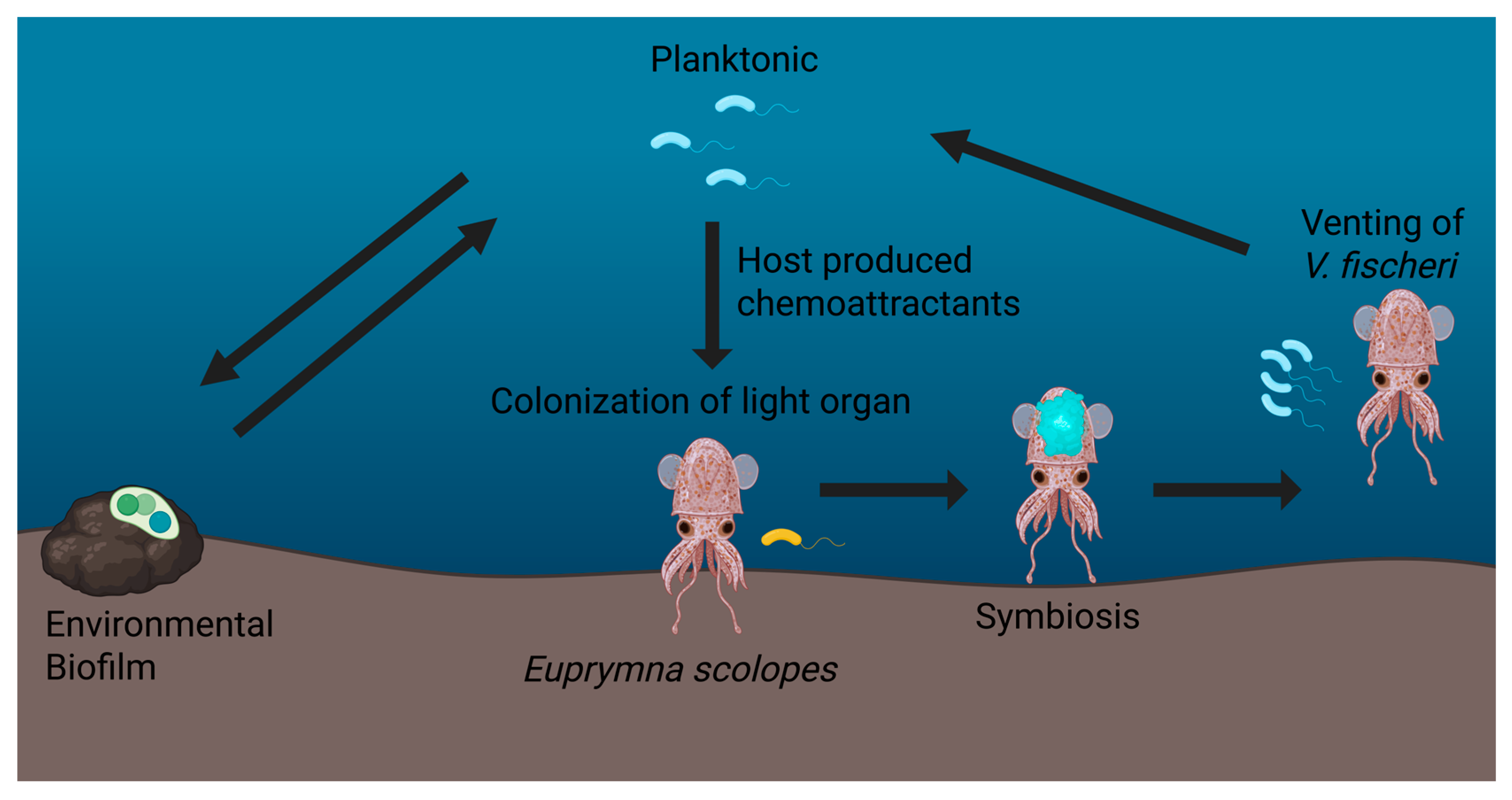
3. Vibrio fischeri as a Model for the Study of Host-Associated Biofilms
4. Expanding the Lens: Other Vibrio and Photobacterium Species
5. Adaptive and Functional Roles of Biofilms in Symbiosis
6. Advances in Understanding Marine Biofilms
7. Conclusions and Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nyholm, S.V.; McFall-Ngai, M.J. A lasting symbiosis: How the Hawaiian bobtail squid finds and keeps its bioluminescent bacterial partner. Nat. Rev. Microbiol. 2021, 19, 666–679. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.W.; Nishiguchi, M.K. Counterillumination in the Hawaiian bobtail squid, Euprymna scolopes Berry (Mollusca: Cephalopoda). Mar. Biol. 2004, 144, 1151–1155. [Google Scholar] [CrossRef]
- Fung, B.L.; Esin, J.J.; Visick, K.L. Vibrio fischeri: A model for host-associated biofilm formation. J. Bacteriol. 2024, 206, e00370-23. [Google Scholar] [CrossRef]
- Nyholm, S.V.; McFall-Ngai, M. The winnowing: Establishing the squid–Vibrio symbiosis. Nat. Rev. Microbiol. 2004, 2, 632–642. [Google Scholar] [CrossRef]
- Soto, W.; Nishiguchi, M.K. Microbial experimental evolution as a novel research approach in the Vibrionaceae and squid-Vibrio symbiosis. Front. Microbiol. 2014, 5, 593. [Google Scholar] [CrossRef] [PubMed]
- Soto, W.; Nishiguchi, M.K. Environmental Stress Selects for Innovations That Drive Vibrio Symbiont Diversity. Front. Ecol. Evol. 2021, 9, 616973. [Google Scholar] [CrossRef]
- Chavez-Dozal, A.; Hogan, D.; Gorman, C.; Quintanal-Villalonga, A.; Nishiguchi, M.K. Multiple Vibrio fischeri genes are involved in biofilm formation and host colonization. FEMS Microbiol. Ecol. 2012, 81, 562–573. [Google Scholar] [CrossRef]
- Nourabadi, N.; Nishiguchi, M.K. PH Adaptation Drives Diverse Phenotypes in a Beneficial Bacterium-Host Mutualism. Front. Ecol. Evol. 2021, 9, 611411. [Google Scholar] [CrossRef]
- Chavez-Dozal, A.A.; Gorman, C.; Lostroh, C.P.; Nishiguchi, M.K. Gene-Swapping Mediates Host Specificity among Symbiotic Bacteria in a Beneficial Symbiosis. PLoS ONE 2014, 9, e101691. [Google Scholar] [CrossRef]
- Visick, K.L.; Schembri, M.A.; Yildiz, F.; Ghigo, J.-M. Biofilms 2015: Multidisciplinary Approaches Shed Light into Microbial Life on Surfaces. J. Bacteriol. 2016, 198, 2553–2563. [Google Scholar] [CrossRef]
- Yildiz, F.H.; Visick, K.L. Vibrio biofilms: So much the same yet so different. Trends. Microbiol. 2009, 17, 109–118. [Google Scholar] [CrossRef]
- Dang, H.; Lovell, C.R. Microbial Surface Colonization and Biofilm Development in Marine Environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef] [PubMed]
- Takemura, A.F.; Chien, D.M.; Polz, M.F. Associations and dynamics of Vibrionaceae in the environment, from the genus to the population level. Front. Microbiol. 2014, 5, 38. [Google Scholar] [CrossRef]
- Zamborsky, D.J.; Nishiguchi, M.K. Phylogeographical Patterns among Mediterranean Sepiolid Squids and Their Vibrio Symbionts: Environment Drives Specificity among Sympatric Species. Appl. Environ. Microbiol. 2011, 77, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.J.; Fullmer, M.S.; Gogarten, J.P.; Nyholm, S. V Comparative genomics of Roseobacter clade bacteria isolated from the accessory nidamental gland of Euprymna scolopes. Front. Microbiol. 2015, 6, 123. [Google Scholar] [CrossRef]
- Kerwin, A.H.; Gromek, S.M.; Suria, A.M.; Samples, R.M.; Deoss, D.J.; O’Donnell, K.; Frasca, S.; Sutton, D.A.; Wiederhold, N.P.; Balunas, M.J.; et al. Shielding the Next Generation: Symbiotic Bacteria from a Reproductive Organ Protect Bobtail Squid Eggs from Fungal Fouling. mBio 2019, 10, e02376-19. [Google Scholar] [CrossRef]
- Chavez-Dozal, A.; Gorman, C.; Erken, M.; Steinberg, P.D.; McDougald, D.; Nishiguchi, M.K. Predation Response of Vibrio fischeri Biofilms to Bacterivorus Protists. Appl. Environ. Microbiol. 2013, 79, 553–558. [Google Scholar] [CrossRef]
- Chavez-Dozal, A.; Gorman, C.; Nishiguchi, M.K. Proteomic and metabolomic profiles demonstrate variation among free-living and symbiotic Vibrio fischeri biofilms. BMC Microbiol. 2015, 15, 226. [Google Scholar] [CrossRef]
- Chavez-Dozal, A.; Soto, W.; Nishiguchi, M.K. Identification of a Transcriptomic Network Underlying the Wrinkly and Smooth Phenotypes of Vibrio fischeri. J. Bacteriol. 2021, 203, e00259-20. [Google Scholar] [CrossRef] [PubMed]
- Yanovski, R.; Barak, H.; Brickner, I.; Kushmaro, A.; Abelson, A. The microbial community of coral reefs: Biofilm composition on artificial substrates under different environmental conditions. Mar. Biol. 2024, 171, 74. [Google Scholar] [CrossRef]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The Microbial “Protective Clothing” in Extreme Environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, C.C.C.R. Marine Biofilms: A Successful Microbial Strategy with Economic Implications. Front. Mar. Sci. 2018, 5, 126. [Google Scholar] [CrossRef]
- Mao-Jones, J.; Ritchie, K.B.; Jones, L.E.; Ellner, S.P. How Microbial Community Composition Regulates Coral Disease Development. PLoS Biol. 2010, 8, e1000345. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Qin, P.; Lu, J.; Wang, S.; Zhang, J.; Wang, Y.; Cheng, A.; Cao, Y.; Ding, W.; Zhang, W. Bioprospecting of culturable marine biofilm bacteria for novel antimicrobial peptides. iMeta 2024, 3, e244. [Google Scholar] [CrossRef]
- Chavez-Dozal, A.; Nishiguchi, M.K. Variation in biofilm formation among symbiotic and free-living strains of Vibrio fischeri. J. Basic Microbiol. 2011, 51, 452–458. [Google Scholar] [CrossRef]
- Waters, C.M.; Lu, W.; Rabinowitz, J.D.; Bassler, B.L. Quorum Sensing Controls Biofilm Formation in Vibrio cholerae through Modulation of Cyclic Di-GMP Levels and Repression of vpsT. J. Bacteriol. 2008, 190, 2527–2536. [Google Scholar] [CrossRef]
- Urbanczyk, H.; Ast, J.C.; Dunlap, P.V. Phylogeny, genomics, and symbiosis of Photobacterium. FEMS Microbiol. Rev. 2011, 35, 324–342. [Google Scholar] [CrossRef]
- Battin, T.J.; Kaplan, L.A.; Denis Newbold, J.; Hansen, C.M.E. Contributions of microbial biofilms to ecosystem processes in stream mesocosms. Nature 2003, 426, 439–442. [Google Scholar] [CrossRef]
- Ghazvinian, M.; Asgharzadeh Marghmalek, S.; Gholami, M.; Amir Gholami, S.; Amiri, E.; Goli, H.R. Antimicrobial resistance patterns, virulence genes, and biofilm formation in enterococci strains collected from different sources. BMC Infect. Dis. 2024, 24, 274. [Google Scholar] [CrossRef]
- Soto, W.; Gutierrez, J.; Remmenga, M.D.; Nishiguchi, M.K. Salinity and Temperature Effects on Physiological Responses of Vibrio fischeri from Diverse Ecological Niches. Microb. Ecol. 2009, 57, 140–150. [Google Scholar] [CrossRef]
- Koch, E.J.; Moriano-Gutierrez, S.; Ruby, E.G.; McFall-Ngai, M.; Liebeke, M. The impact of persistent colonization by Vibrio fischeri on the metabolome of the host squid Euprymna scolopes. J. Exp. Biol. 2020, 223, jeb212860. [Google Scholar] [CrossRef] [PubMed]
- Fung, B.L.; Visick, K.L. LitR and its quorum-sensing regulators modulate biofilm formation by Vibrio fischeri. J. Bacteriol. 2025, 207, e00476-24. [Google Scholar] [CrossRef] [PubMed]
- Septer, A.N.; Visick, K.L. Lighting the way: How the Vibrio fischeri model microbe reveals the complexity of Earth’s “simplest” life forms. J. Bacteriol. 2024, 206, e00035-24. [Google Scholar] [CrossRef]
- Pan, M.; Schwartzman, J.A.; Dunn, A.K.; Lu, Z.; Ruby, E.G. A Single Host-Derived Glycan Impacts Key Regulatory Nodes of Symbiont Metabolism in a Coevolved Mutualism. mBio 2015, 6, e00811. [Google Scholar] [CrossRef]
- Wang, Y.; Dufour, Y.S.; Carlson, H.K.; Donohue, T.J.; Marletta, M.A.; Ruby, E.G. H-NOX–mediated nitric oxide sensing modulates symbiotic colonization by Vibrio fischeri. Proc. Natl. Acad. Sci. USA 2010, 107, 8375–8380. [Google Scholar] [CrossRef]
- Verma, S.C.; Miyashiro, T. Quorum sensing in the squid-Vibrio symbiosis. Int. J. Mol. Sci. 2013, 14, 16386–16401. [Google Scholar] [CrossRef]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef]
- Lade, H.; Paul, D.; Kweon, J.H. N-acyl homoserine lactone-mediated quorum sensing with special reference to use of quorum quenching bacteria in membrane biofouling control. Biomed. Res. Int. 2014, 2014, 162584. [Google Scholar] [CrossRef] [PubMed]
- Shibata, S.; Yip, E.S.; Quirke, K.P.; Ondrey, J.M.; Visick, K.L. Roles of the structural symbiosis polysaccharide (syp) genes in host colonization, biofilm formation, and polysaccharide biosynthesis in Vibrio fischeri. J. Bacteriol. 2012, 194, 6736–6747. [Google Scholar] [CrossRef]
- Yip, E.S.; Geszvain, K.; DeLoney-Marino, C.R.; Visick, K.L. The symbiosis regulator rscS controls the syp gene locus, biofilm formation and symbiotic aggregation by Vibrio fischeri. Mol. Microbiol. 2006, 62, 1586–1600. [Google Scholar] [CrossRef]
- Browne-Silva, J.; Nishiguchi, M.K. Gene sequences of the pil operon reveal relationships between symbiotic strains of Vibrio fischeri. Int. J. Syst. Evol. Microbiol. 2008, 58, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Darnell, C.L.; Hussa, E.A.; Visick, K.L. The Putative Hybrid Sensor Kinase SypF Coordinates Biofilm Formation in Vibrio fischeri by Acting Upstream of Two Response Regulators, SypG and VpsR. J. Bacteriol. 2008, 190, 4941–4950. [Google Scholar] [CrossRef] [PubMed]
- Millikan, D.S.; Ruby, E.G. Vibrio fischeri flagellin A is essential for normal motility and for symbiotic competence during initial squid light organ colonization. J. Bacteriol. 2004, 186, 4315–4325. [Google Scholar] [CrossRef]
- Colin, R.; Ni, B.; Laganenka, L.; Sourjik, V. Multiple functions of flagellar motility and chemotaxis in bacterial physiology. FEMS Microbiol. Rev. 2021, 45, fuab038. [Google Scholar] [CrossRef]
- Ariyakumar, D.S.; Nishiguchi, M.K. Characterization of two host-specific genes, mannose-sensitive hemagglutinin (mshA) and uridyl phosphate dehydrogenase (UDPDH) that are involved in the Vibrio fischeri-Euprymna tasmanica mutualism. FEMS Microbiol. Lett. 2009, 299, 65–73. [Google Scholar] [CrossRef]
- Aschtgen, M.-S.; Brennan, C.A.; Nikolakakis, K.; Cohen, S.; McFall-Ngai, M.; Ruby, E.G. Insights into flagellar function and mechanism from the squid–Vibrio symbiosis. npj Biofilms Microbiomes 2019, 5, 32. [Google Scholar] [CrossRef]
- Thompson, C.M.; Tischler, A.H.; Tarnowski, D.A.; Mandel, M.J.; Visick, K.L. Nitric oxide inhibits biofilm formation by Vibrio fischeri via the nitric oxide sensor HnoX. Mol. Microbiol. 2019, 111, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Nyholm, S.V.; Deplancke, B.; Gaskins, H.R.; Apicella, M.A.; McFall-Ngai, M.J. Roles of Vibrio fischeri and Nonsymbiotic Bacteria in the Dynamics of Mucus Secretion during Symbiont Colonization of the Euprymna scolopes Light Organ. Appl. Environ. Microbiol. 2002, 68, 5113–5122. [Google Scholar] [CrossRef]
- Cohen, M.L.; Mashanova, E.V.; Rosen, N.M.; Soto, W. Adaptation to temperature stress by Vibrio fischeri facilitates this microbe’s symbiosis with the Hawaiian bobtail squid (Euprymna scolopes). Evolution 2019, 73, 1885–1897. [Google Scholar] [CrossRef]
- Hussa, E.A.; Darnell, C.L.; Visick, K.L. RscS Functions Upstream of SypG to Control the syp Locus and Biofilm Formation in Vibrio fischeri. J. Bacteriol. 2008, 190, 4576–4583. [Google Scholar] [CrossRef]
- Ray, V.A.; Eddy, J.L.; Hussa, E.A.; Misale, M.; Visick, K.L. The syp Enhancer Sequence Plays a Key Role in Transcriptional Activation by the 54-Dependent Response Regulator SypG and in Biofilm Formation and Host Colonization by Vibrio fischeri. J. Bacteriol. 2013, 195, 5402–5412. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.R.; Visick, K.L. The response regulator SypE controls biofilm formation and colonization through phosphorylation of the syp-encoded regulator SypA in Vibrio fischeri. Mol. Microbiol. 2013, 87, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Brooks, J.F.; Mandel, M.J. The Histidine Kinase BinK Is a Negative Regulator of Biofilm Formation and Squid Colonization. J. Bacteriol. 2016, 198, 2596–2607. [Google Scholar] [CrossRef] [PubMed]
- Stabb, E.V.; Ruby, E.G. Contribution of pilA to competitive colonization of the squid Euprymna scolopes by Vibrio fischeri. Appl. Environ. Microbiol. 2003, 69, 820–826. [Google Scholar] [CrossRef]
- Millikan, D.S.; Ruby, E.G. FlrA, a σ 54-Dependent Transcriptional Activator in Vibrio fischeri, Is Required for Motility and Symbiotic Light-Organ Colonization. J. Bacteriol. 2003, 185, 3547–3557. [Google Scholar] [CrossRef]
- Isenberg, R.Y.; Holschbach, C.S.; Gao, J.; Mandel, M.J. Functional analysis of cyclic diguanylate-modulating proteins in Vibrio fischeri. mSystems 2024, 9, e0095624. [Google Scholar] [CrossRef]
- Visick, K.L.; Foster, J.; Doino, J.; McFall-Ngai, M.; Ruby, E.G. Vibrio fischeri lux Genes Play an Important Role in Colonization and Development of the Host Light Organ. J. Bacteriol. 2000, 182, 4578–4586. [Google Scholar] [CrossRef]
- Nishiguchi, M.K.; Hirsch, A.M.; Devinney, R.; Vedantam, G.; Riley, M.A.; Mansky, L.M. Deciphering Evolutionary Mechanisms Between Mutualistic and Pathogenic Symbioses. Vie Milieu 2008, 58, 87–106. [Google Scholar]
- Bellissimo, K.A.; Septer, A.N.; Whistler, C.A.; Rodríguez, C.; Stabb, E.V. Deletion of luxI increases luminescence of Vibrio fischeri. mBio 2024, 15, e02446-24. [Google Scholar] [CrossRef]
- Plate, L.; Marletta, M.A. Nitric Oxide Modulates Bacterial Biofilm Formation through a Multicomponent Cyclic-di-GMP Signaling Network. Mol. Cell 2012, 46, 449–460. [Google Scholar] [CrossRef]
- Silva, A.J.; Benitez, J.A. Vibrio cholerae Biofilms and Cholera Pathogenesis. PLoS Negl. Trop. Dis. 2016, 10, e0004330. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Cooper, J.N.; Mishra, A.; Raskin, D.M. Stringent Response Regulation of Biofilm Formation in Vibrio cholerae. J. Bacteriol. 2012, 194, 2962–2972. [Google Scholar] [CrossRef] [PubMed]
- Lutz, C.; Erken, M.; Noorian, P.; Sun, S.; McDougald, D. Environmental reservoirs and mechanisms of persistence of Vibrio cholerae. Front. Microbiol. 2013, 4, 375. [Google Scholar] [CrossRef]
- Mishra, A.; Aggarwal, A.; Khan, F. Medical Device-Associated Infections Caused by Biofilm-Forming Microbial Pathogens and Controlling Strategies. Antibiotics 2024, 13, 623. [Google Scholar] [CrossRef]
- Haygood, M.G.; Distel, D.L. Bioluminescent symbionts of flashlight fishes and deep-sea anglerfishes form unique lineages related to the genus Vibrio. Nature 1993, 363, 154–156. [Google Scholar] [CrossRef]
- Tanet, L.; Tamburini, C.; Baumas, C.; Garel, M.; Simon, G.; Casalot, L. Bacterial Bioluminescence: Light Emission in Photobacterium phosphoreum Is Not Under Quorum-Sensing Control. Front. Microbiol. 2019, 10, 365. [Google Scholar] [CrossRef] [PubMed]
- Machineni, L. Effects of biotic and abiotic factors on biofilm growth dynamics and their heterogeneous response to antibiotic challenge. J. Biosci. 2020, 45, 25. [Google Scholar] [CrossRef]
- Kitts, G.; Rogers, A.; Teschler, J.K.; Park, J.H.; Trebino, M.A.; Chaudry, I.; Erill, I.; Yildiz, F.H. The Rvv two-component regulatory system regulates biofilm formation and colonization in Vibrio cholerae. PLoS Pathog. 2023, 19, e1011415. [Google Scholar] [CrossRef]
- Marsden, A.E.; Grudzinski, K.; Ondrey, J.M.; DeLoney-Marino, C.R.; Visick, K.L. Impact of Salt and Nutrient Content on Biofilm Formation by Vibrio fischeri. PLoS ONE 2017, 12, e0169521. [Google Scholar] [CrossRef]
- Ramos-Vivas, J.; Acosta, F. Editorial: Host-bacteria interactions in fish pathogens. Front. Cell. Infect. Microbiol. 2024, 14, 1515641. [Google Scholar] [CrossRef]
- Mass, S.; Cohen, H.; Podicheti, R.; Rusch, D.B.; Gerlic, M.; Ushijima, B.; van Kessel, J.C.; Bosis, E.; Salomon, D. The coral pathogen Vibrio coralliilyticus uses a T6SS to secrete a group of novel anti-eukaryotic effectors that contribute to virulence. PLoS Biol. 2024, 22, e3002734. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Chen, X.; Liu, R.; Tian, P.; Niu, W.; Zhang, X.-H.; Liu, J.; Wang, X. Distinct coral environments shape the dynamic of planktonic Vibrio spp. Environ. Microbiome 2023, 18, 77. [Google Scholar] [CrossRef]
- Mincer, T.J.; Zettler, E.R.; Amaral-Zettler, L.A. Biofilms on Plastic Debris and Their Influence on Marine Nutrient Cycling, Productivity, and Hazardous Chemical Mobility. In Hazardous Chemicals Associated with Plastics in the Marine Environment; Springer: Berlin/Heidelberg, Germany, 2016; pp. 221–233. [Google Scholar]
- Zhang, X.; Lin, H.; Wang, X.; Austin, B. Significance of Vibrio species in the marine organic carbon cycle—A review. Sci. China Earth Sci. 2018, 61, 1357–1368. [Google Scholar] [CrossRef]
- Alotaibi, G.F. Factors Influencing Bacterial Biofilm Formation and Development. Am. J. Biomed. Sci. Res. 2021, 12, 617–626. [Google Scholar] [CrossRef]
- Filippini, G.; Bugnot, A.B.; Varkey, D.R.; Siboni, N.; Ferguson, A.; Gribben, P.E.; Erickson, K.; Palmer, J.; Dafforn, K.A. Nitrogen-cycling genes in oyster reefs and surrounding sediments: Relationships with environmental factors and respective nitrogen rates. Mar. Pollut. Bull. 2023, 197, 115710. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.E.; O’toole, G.A. Microbial Biofilms: From Ecology to Molecular Genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef]
- McCaughey, C.S.; Trebino, M.A.; McAtamney, A.; Isenberg, R.Y.; Mandel, M.J.; Yildiz, F.H.; Sanchez, L.M. A Label-Free Approach for Relative Spatial Quantitation of c-di-GMP in Microbial Biofilms. Anal. Chem. 2024, 96, 8308–8316. [Google Scholar] [CrossRef]
- Teschler, J.K.; Nadell, C.D.; Drescher, K.; Yildiz, F.H. Mechanisms Underlying Vibrio cholerae Biofilm Formation and Dispersion. Annu. Rev. Microbiol. 2022, 76, 503–532. [Google Scholar] [CrossRef]
- Shrestha, P.; Razvi, A.; Fung, B.L.; Eichinger, S.J.; Visick, K.L. Mutational Analysis of Vibrio fischeri c-di-GMP-Modulating Genes Reveals Complex Regulation of Motility. J. Bacteriol. 2022, 204, e00109-22. [Google Scholar] [CrossRef]
- Ragupathi, H.; Pushparaj, M.M.; Gopi, S.M.; Govindarajan, D.K.; Kandaswamy, K. Biofilm matrix: A multifaceted layer of biomolecules and a defensive barrier against antimicrobials. Arch. Microbiol. 2024, 206, 432. [Google Scholar] [CrossRef]
- Almatroudi, A. Biofilm Resilience: Molecular Mechanisms Driving Antibiotic Resistance in Clinical Contexts. Biology 2025, 14, 165. [Google Scholar] [CrossRef] [PubMed]
- Roilides, E.; Simitsopoulou, M.; Katragkou, A.; Walsh, T.J. How Biofilms Evade Host Defenses. Microb. Biofilms 2015, 3, 287–300. [Google Scholar] [CrossRef]
- Hitzler, S.U.J.; Fernández-Fernández, C.; Montaño, D.E.; Dietschmann, A.; Gresnigt, M.S. Microbial adaptive pathogenicity strategies to the host inflammatory environment. FEMS Microbiol. Rev. 2025, 49, fuae032. [Google Scholar] [CrossRef] [PubMed]
- Tanveer, M.; Ntakiyisumba, E.; Won, G. Revealing antimicrobial resistance profile and associated factors of Vibrio vulnificus isolated from clinical, environmental, and seafood samples across asia: A systematic review and meta-analysis. Heliyon 2024, 10, e40334. [Google Scholar] [CrossRef]
- Azeem, K.; Fatima, S.; Ali, A.; Ubaid, A.; Husain, F.M.; Abid, M. Biochemistry of Bacterial Biofilm: Insights into Antibiotic Resistance Mechanisms and Therapeutic Intervention. Life 2025, 15, 49. [Google Scholar] [CrossRef]
- Mandel, M.J.; Dunn, A.K. Impact and Influence of the Natural Vibrio-Squid Symbiosis in Understanding Bacterial–Animal Interactions. Front. Microbiol. 2016, 7, 1982. [Google Scholar] [CrossRef] [PubMed]
- Muthulakshmi, T.; Mothadaka, M.P. Vibrio Vulnificus and Its Antimicrobial Resistance. In Handbook on Antimicrobial Resistance; Springer Nature: Singapore, 2023; pp. 1–18. [Google Scholar]
- Yang, Y.; Yan, J.; Olson, R.; Jiang, X. Comprehensive Genomic and Evolutionary Analysis of Biofilm Matrix Clusters and Proteins in the Vibrio Genus 2024. mSystems 2025, 10, e00060-25. [Google Scholar] [CrossRef]
- Pang, R.; Xie, T.; Wu, Q.; Li, Y.; Lei, T.; Zhang, J.; Ding, Y.; Wang, J.; Xue, L.; Chen, M.; et al. Comparative Genomic Analysis Reveals the Potential Risk of Vibrio parahaemolyticus Isolated from Ready-To-Eat Foods in China. Front. Microbiol. 2019, 10, 186. [Google Scholar] [CrossRef]
- Steenackers, H.P.; Parijs, I.; Foster, K.R.; Vanderleyden, J. Experimental evolution in biofilm populations. FEMS Microbiol. Rev. 2016, 40, 373–397. [Google Scholar] [CrossRef]
- Norsworthy, A.N.; Visick, K.L. Gimme shelter: How Vibrio fischeri successfully navigates an animal’s multiple environments. Front. Microbiol. 2013, 4, 356. [Google Scholar] [CrossRef]
- Kuper, T.J.; Islam, M.M.; Peirce-Cottler, S.M.; Papin, J.A.; Ford, R.M. Spatial transcriptome-guided multi-scale framework connects P. aeruginosa metabolic states to oxidative stress biofilm microenvironment. PLoS Comput. Biol. 2024, 20, e1012031. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Ostrov, N.; Wong, B.G.; Gold, M.A.; Khalil, A.S.; Church, G.M. Functional genomics of the rapidly replicating bacterium Vibrio natriegens by CRISPRi. Nat. Microbiol. 2019, 4, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Pipes, B.L.; Nishiguchi, M.K. Nocturnal Acidification: A Coordinating Cue in the Euprymna scolopes–Vibrio fischeri Symbiosis. Int. J. Mol. Sci. 2022, 23, 3743. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, A.; Silva, V.; Poeta, P.; Aonofriesei, F. Vibrio spp.: Life Strategies, Ecology, and Risks in a Changing Environment. Diversity 2022, 14, 97. [Google Scholar] [CrossRef]
- Vezzulli, L.; Pezzati, E.; Brettar, I.; Höfle, M.; Pruzzo, C. Effects of Global Warming on Vibrio Ecology. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef]
- Sauer, K. Cyclic Di-GMP and the Regulation of Biofilm Dispersion. In Microbial Cyclic Di-Nucleotide Signaling; Springer International Publishing: Cham, Switzerland, 2020; pp. 545–560. [Google Scholar]
- Griend, J.A.V.; Isenberg, R.Y.; Kotla, K.R.; Mandel, M.J. Transcriptional Pathways across Colony Biofilm Models in the Symbiont Vibrio fischeri. bioRxiv 2023. [Google Scholar] [CrossRef]
- Youngblom, M.A.; Smith, T.M.; Murray, H.J.; Pepperell, C.S. Adaptation of the Mycobacterium tuberculosis transcriptome to biofilm growth. PLoS Pathog. 2024, 20, e1012124. [Google Scholar] [CrossRef]
- Madsen, J.S.; Burmølle, M.; Hansen, L.H.; Sørensen, S.J. The interconnection between biofilm formation and horizontal gene transfer. FEMS Immunol. Med. Microbiol. 2012, 65, 183–195. [Google Scholar] [CrossRef]
- León, M.; Kokkari, C.; García, K.; Castillo, D.; Katharios, P.; Bastías, R. Diversification of Vibrio anguillarum Driven by the Bacteriophage CHOED. Front. Microbiol. 2019, 10, 1396. [Google Scholar] [CrossRef]
- Martin, M.; Hölscher, T.; Dragoš, A.; Cooper, V.S.; Kovács, Á.T. Laboratory Evolution of Microbial Interactions in Bacterial Biofilms. J. Bacteriol. 2016, 198, 2564–2571. [Google Scholar] [CrossRef]
- Soto, W.; Rivera, F.M.; Nishiguchi, M.K. Ecological Diversification of Vibrio fischeri Serially Passaged for 500 Generations in Novel Squid Host Euprymna tasmanica. Microb. Ecol. 2014, 67, 700–721. [Google Scholar] [CrossRef] [PubMed]
- Barraud, N.; Kjelleberg, S.; Rice, S.A. Dispersal from Microbial Biofilms. Microb. Biofilms 2015, 3, 343–362. [Google Scholar] [CrossRef]
- Edel, M.; Horn, H.; Gescher, J. Biofilm systems as tools in biotechnological production. Appl. Microbiol. Biotechnol. 2019, 103, 5095–5103. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, N.S.; da Silva Ramos, R.C.P.; de Paula, R.C.; da Costa Pereira, M.G.; Rosa, R.T.; Bianchini, L.F.; Rosa, E.A.R. Advantages of using biofilms to obtain high-value molecules by microbial biotransformations. Explor. Drug Sci. 2025, 3, 100884. [Google Scholar] [CrossRef]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting microbial biofilms: Current and prospective therapeutic strategies. Nat. Rev. Microbiol. 2017, 15, 740–755. [Google Scholar] [CrossRef]
- Ruby, E.G.; Lee, K.-H. The Vibrio fischeri-Euprymna scolopes Light Organ Association: Current Ecological Paradigms. Appl. Environ. Microbiol. 1998, 64, 805–812. [Google Scholar] [CrossRef]
- Prentice, J.A.; Bridges, A.A.; Bassler, B.L. Synergy between c-di-GMP and Quorum-Sensing Signaling in Vibrio cholerae Biofilm Morphogenesis. J. Bacteriol. 2022, 204, e00249-22. [Google Scholar] [CrossRef]
- Conner, J.G.; Teschler, J.K.; Jones, C.J.; Yildiz, F.H. Staying Alive: Vibrio cholerae ’s Cycle of Environmental Survival, Transmission, and Dissemination. Virulence Mech. Bact. Pathog. 2016, 4, 593–633. [Google Scholar] [CrossRef]
- Kilic, T. Factors Affecting Biofilm Formation and the Effects of These Factors on Bacteria. In Exploring Bacterial Biofilms [Working Title]; Intech Open: Rijeka, Croatia, 2025. [Google Scholar]
- Chan, W.Y.; Rudd, D.; van Oppen, M.J. Spatial metabolomics for symbiotic marine invertebrates. Life Sci. Alliance 2023, 6, e202301900. [Google Scholar] [CrossRef]
- Yun, M.S.; Sun, J.; Lovejoy, C.; Lee, S.H. Editorial: Microbial Response to a Rapidly Changing Marine Environment: Global Warming and Ocean Acidification. Front. Microbiol. 2021, 12, 731732. [Google Scholar] [CrossRef]
- Yeor-Davidi, E.; Zverzhinetsky, M.; Krivitsky, V.; Patolsky, F. Real-time monitoring of bacterial biofilms metabolic activity by a redox-reactive nanosensors array. J. Nanobiotechnol. 2020, 18, 81. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Liao, H.; Wang, C.; Huang, C.; Zhang, W.; Guo, C.; Pu, Y. An improved bacterial single-cell RNA-seq reveals biofilm heterogeneity. eLife 2024, 13, RP97543. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Category | Molecule/Gene | Type/Role | Function in Biofilm or Symbiosis | Reference |
|---|---|---|---|---|
| Host Cue | Chitin/chitobiose | Carbohydrate | Induces chemotaxis and activates syp genes to initiate biofilm formation. | [46] |
| Host Cue | Nitric oxide (NO) | Reactive species | Temporally inhibits biofilm formation; contributes to daily symbiont expulsion. | [47] |
| Host Cue | Mucus (mucins) | Glycoprotein | Provides scaffold for aggregation; supports early colonization. | [48] |
| Host Cue | Temperature/pH shift | Environmental | Signals transition from seawater to crypts; modulates V. fischeri gene expression. | [49] |
| Regulator | rscS | Sensor kinase | Master regulator that initiates syp biofilm gene expression. | [40,50] |
| Regulator | sypG | Response regulator | Activates transcription of syp locus for polysaccharide production. | [51] |
| Regulator | sypE | Dual-function regulator | Modulates sypA activity, balancing biofilm formation and dispersal. | [51] |
| Matrix Genes | sypA–sypR | Structural operon | Encodes exopolysaccharide synthesis and transport for biofilm matrix. | [52] |
| Negative Regulator | binK | Sensor kinase | Inhibits syp transcription and biofilm formation; responds to environmental cues. | [53] |
| Surface Interaction | pilA, pilT | Type IV pili | Essential for initial attachment to host mucus. | [54] |
| Motility Gene | flrA, motB | Flagellar regulators | Loss reduces motility and promotes biofilm formation. | [55] |
| Second Messenger | mifB | c-di-GMP synthase | Modulates biofilm via intracellular signaling pathways. | [56] |
| Quorum Sensing | luxR/luxO | Transcriptional regulators | Coordinates biofilm gene expression and light production in response to population density. | [57] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucero, J.; Nishiguchi, M.K. Host-Associated Biofilms: Vibrio fischeri and Other Symbiotic Bacteria Within the Vibrionaceae. Microorganisms 2025, 13, 1223. https://doi.org/10.3390/microorganisms13061223
Lucero J, Nishiguchi MK. Host-Associated Biofilms: Vibrio fischeri and Other Symbiotic Bacteria Within the Vibrionaceae. Microorganisms. 2025; 13(6):1223. https://doi.org/10.3390/microorganisms13061223
Chicago/Turabian StyleLucero, Joaquin, and Michele K. Nishiguchi. 2025. "Host-Associated Biofilms: Vibrio fischeri and Other Symbiotic Bacteria Within the Vibrionaceae" Microorganisms 13, no. 6: 1223. https://doi.org/10.3390/microorganisms13061223
APA StyleLucero, J., & Nishiguchi, M. K. (2025). Host-Associated Biofilms: Vibrio fischeri and Other Symbiotic Bacteria Within the Vibrionaceae. Microorganisms, 13(6), 1223. https://doi.org/10.3390/microorganisms13061223