

Boosting Broiler Health and Productivity: The Impact of in ovo Probiotics and Early Posthatch Feeding with Bacillus subtilis, Lactobacillus fermentum, and Enterococcus faecium



Abstract
1. Introduction
2. Materials and Methods
2.1. Probiotics
2.2. Experimental Design and Probiotic Embryo Intake Effectiveness Control Tests
2.3. Poultry Farming Conditions
2.4. Materials for Histological Studies
2.5. Immunohistochemical Staining
2.6. Morphometry
2.7. Isolation and Identification of Probiotic and Campylobacter Strains from the Intestinal Tract
2.8. Statistical Analysis
3. Results
3.1. Probiotics
3.2. Experimental Design and Probiotic Embryo Intake Effectiveness Control Tests
3.3. Histology
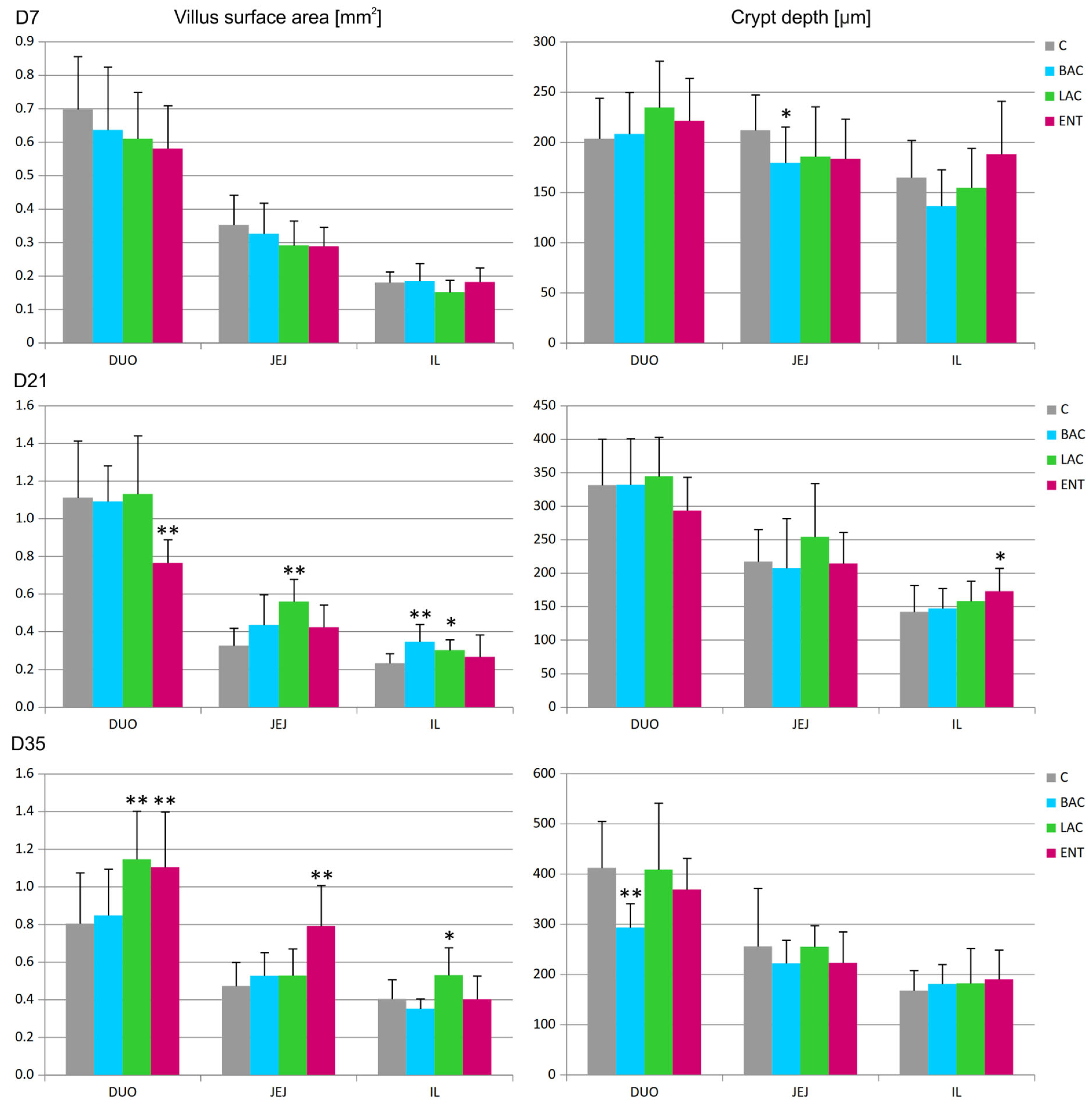
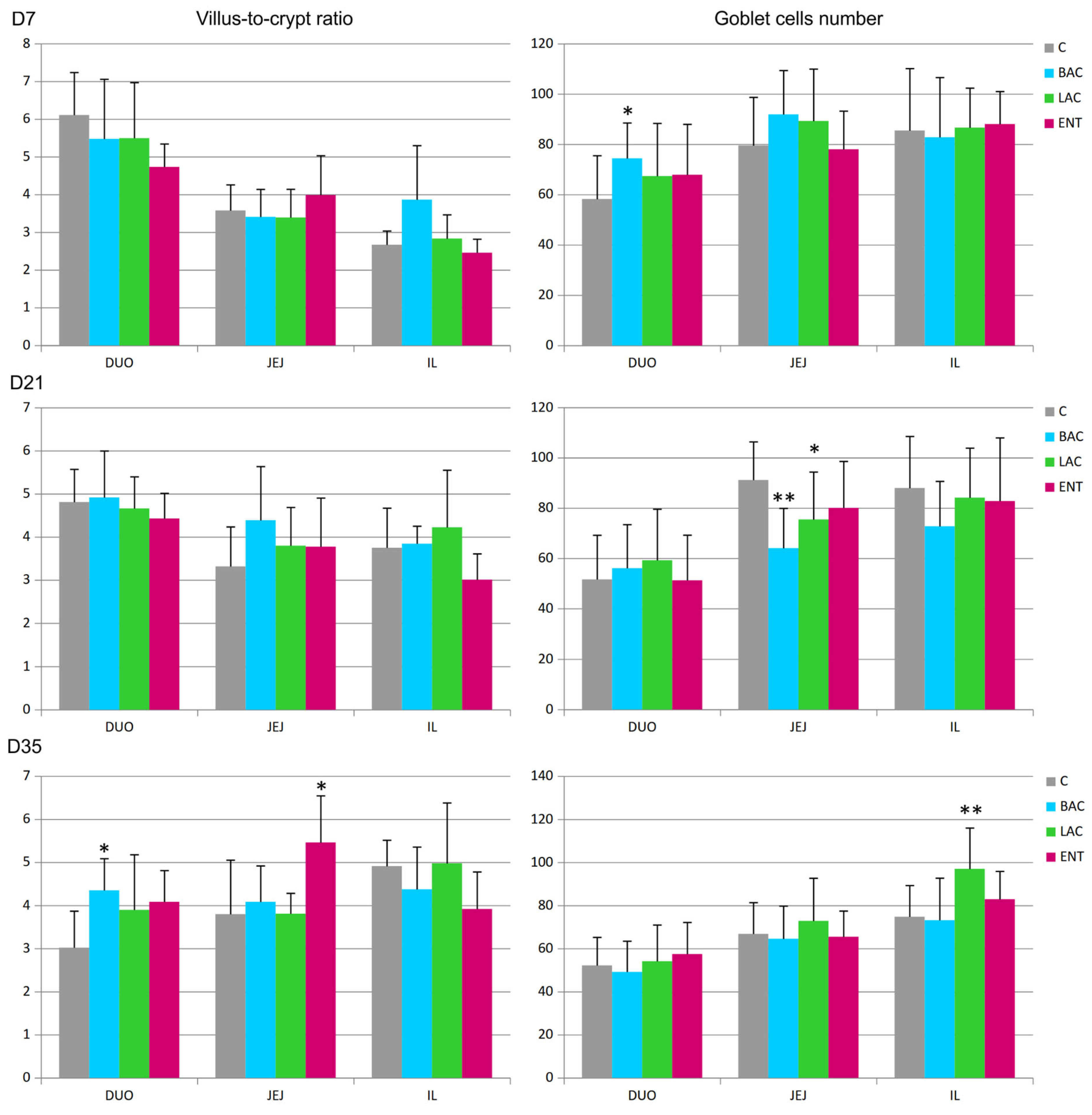
3.4. Morphometry of the Intestines
3.5. Morphometry of Immune Organs
3.6. Isolation and Identification of the Culture of Campylobacter spp. from the Intestinal Tract
3.7. Hatchability and Final Production Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. WHO Guidelines on Use of Medically Important Antimicrobials in Food-Producing Animals; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global Trends in Antimicrobial Use in Food Animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [PubMed]
- Woźniak-Biel, A.; Bugla-Płoskońska, G.; Kielsznia, A.; Korzekwa, K.; Tobiasz, A.; Korzeniowska-Kowal, A.; Wieliczko, A. High Prevalence of Resistance to Fluoroquinolones and Tetracycline Campylobacter Spp. Isolated from Poultry in Poland. Microb. Drug Resist. 2018, 24, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Woźniak-Biel, A.; Bugla-Płoskońska, G.; Burdzy, J.; Korzekwa, K.; Ploch, S.; Wieliczko, A. Antimicrobial Resistance and Biofilm Formation in Enterococcus Spp. Isolated from Humans and Turkeys in Poland. Microb. Drug Resist. 2019, 25, 277–286. [Google Scholar] [CrossRef]
- Muaz, K.; Riaz, M.; Akhtar, S.; Park, S.; Ismail, A. Antibiotic Residues in Chicken Meat: Global Prevalence, Threats, and Decontamination Strategies: A Review. J. Food Prot. 2018, 81, 619–627. [Google Scholar] [CrossRef]
- Kober, A.K.M.H.; Riaz Rajoka, M.S.; Mehwish, H.M.; Villena, J.; Kitazawa, H. Immunomodulation Potential of Probiotics: A novel strategy for improving livestock health, immunity, and productivity. Microorganisms 2022, 10, 388. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.S.; Kim, I.H. Effect of Bacillus Subtilis C-3102 Spores as a Probiotic Feed Supplement on Growth Performance, Noxious Gas Emission, and Intestinal Microflora in Broilers. Poult. Sci. 2014, 93, 3097–3103. [Google Scholar] [CrossRef]
- Younas, S.; Bukhari, D.A.; Bibi, Z.; Ullah, A.; Rehman, A. Impact of Multistrain Probiotics on Growth Performance, Immune Response, and Gut Morphometry in Broiler Chicken Gallus Gallus Domesticus. Poult. Sci. 2025, 104, 105026. [Google Scholar] [CrossRef]
- Castañeda, C.D.; McDaniel, C.D.; Abdelhamed, H.; Karsi, A.; Kiess, A.S. Evaluating bacterial colonization of a developing broiler embryo after in ovo injection with a bioluminescent bacteria. Poult. Sci. 2019, 98, 2997–3006. [Google Scholar] [CrossRef]
- Zhai, W.; Neuman, S.; Latour, M.A.; Hester, P.Y. The Effect of in Ovo Injection of L-Carnitine on Hatchability of White Leghorns. Poult. Sci. 2008, 87, 569–572. [Google Scholar] [CrossRef]
- Hong, H.A.; Duc, L.H.; Cutting, S.M. The Use of Bacterial Spore Formers as Probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef]
- Mohamed, T.M.; Sun, W.; Bumbie, G.Z.; Elokil, A.A.; Mohammed, K.A.F.; Zebin, R.; Hu, P.; Wu, L.; Tang, Z. Feeding Bacillus subtilis atcc19659 to broiler chickens enhances growth performance and immune function by modulating intestinal morphology and cecum microbiota. Front. Microbiol. 2021, 12, 798350. [Google Scholar] [CrossRef] [PubMed]
- Penha Filho, R.A.C.; Díaz, S.J.A.; Fernando, F.S.; Chang, Y.-F.; Andreatti Filho, R.L.; Berchieri Junior, A. immunomodulatory activity and control of Salmonella enteritidis colonization in the intestinal tract of chickens by lactobacillus based probiotic. Vet. Immunol. Immunopathol. 2015, 167, 64–69. [Google Scholar] [CrossRef]
- Castañeda, C.D.; Dittoe, D.K.; Wamsley, K.G.S.; McDaniel, C.D.; Blanch, A.; Sandvang, D.; Kiess, A.S. In ovo inoculation of an Enterococcus faecium-based product to enhance broiler hatchability, live performance, and intestinal morphology. Poult. Sci. 2020, 99, 6163–6172. [Google Scholar] [CrossRef] [PubMed]
- Al-Surrayai, T.; Al-Khalaifah, H.; Al-Mansour, H.; Kishk, M.; Al-Mutairi, A.; Sultan, H.; Al-Saleem, H. Evaluation of the Lactic Acid Bacteria Based Formulated Probiotic Product for Poultry. Front. Anim. Sci. 2022, 3, 1026958. [Google Scholar] [CrossRef]
- Pourabedin, M.; Zhao, X. Prebiotics and Gut Microbiota in Chickens. FEMS Microbiol. Lett. 2015, 362, fnv122. [Google Scholar] [CrossRef] [PubMed]
- Ocejo, M.; Oporto, B.; Juste, R.A.; Hurtado, A. Effects of Dry Whey Powder and Calcium Butyrate Supplementation of Corn/Soybean-Based Diets on Productive Performance, Duodenal Histological Integrity, and Campylobacter Colonization in Broilers. BMC Vet. Res. 2017, 13, 199. [Google Scholar] [CrossRef]
- Deng, W.; Dittoe, D.K.; Pavilidis, H.O.; Chaney, W.E.; Yang, Y.; Ricke, S.C. Current Perspectives and Potential of Probiotics to Limit Foodborne Campylobacter in Poultry. Front. Microbiol. 2020, 11, 583429. [Google Scholar] [CrossRef]
- Connerton, P.L.; Richards, P.J.; Lafontaine, G.M.; O’Kane, P.M.; Ghaffar, N.; Cummings, N.J.; Smith, D.L.; Fish, N.M.; Connerton, I.F. The Effect of the Timing of Exposure to Campylobacter Jejuni on the Gut Microbiome and Inflammatory Responses of Broiler Chickens. Microbiome 2018, 6, 88. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global epidemiology of Campylobacter infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef]
- Heimesaat, M.M.; Backert, S.; Alter, T.; Bereswill, S. Human campylobacteriosis-A serious infectious threat in a one health perspective. Curr. Top Microbiol. Immunol. 2021, 431, 1–23. [Google Scholar]
- Burnham, P.M.; Hendrixson, D.R. Campylobacter Jejuni: Collective components promoting a successful enteric lifestyle. Nat. Rev. Microbiol. 2018, 16, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Madej, J.P.; Graczyk, S.; Bobrek, K.; Bajzert, J.; Gaweł, A. Impact of early posthatch feeding on the immune system and selected hematological, biochemical, and hormonal parameters in broiler chickens. Poult. Sci. 2024, 103, 103366. [Google Scholar] [CrossRef] [PubMed]
- Proszkowiec-Weglarz, M.; Miska, K.B.; Ellestad, L.E.; Schreier, L.L.; Kahl, S.; Darwish, N.; Campos, P.; Shao, J. Delayed access to feed early post-hatch affects the development and maturation of gastrointestinal tract microbiota in broiler chickens. BMC Microbiol. 2022, 22, 206. [Google Scholar] [CrossRef]
- Gaweł, A.; Madej, J.P.; Kozak, B.; Bobrek, K. Early Post-Hatch Nutrition Influences Performance and Muscle Growth in Broiler Chickens. Animals 2022, 12, 3281. [Google Scholar] [CrossRef]
- Kariyama, R.; Mitsuhata, R.; Chow, J.W.; Clewell, D.B.; Kumon, H. Simple and reliable multiplex PCR assay for surveillance isolates of vancomycin-resistant enterococci. J. Clin. Microbiol. 2000, 38, 3092–3095. [Google Scholar] [CrossRef]
- Wattiau, P.; Renard, M.E.; Ledent, P.; Debois, V.; Blackman, G.; Agathos, S.N. A PCR test to identify Bacillus subtilis and closely related species and its application to the monitoring of wastewater biotreatment. Appl. Microbiol. Biotechnol. 2001, 56, 816–819. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Yang, S.-M.; Lim, B.; Park, S.H.; Rackerby, B.; Kim, H.-Y. Design of PCR assays to specifically detect and identify 37 Lactobacillus species in a single 96 well plate. BMC Microbiol. 2020, 20, 96. [Google Scholar] [CrossRef]
- Bazireh, H.; Shariati, P.; Azimzadeh Jamalkandi, S.; Ahmadi, A.; Boroumand, M.A. Isolation of novel probiotic Lactobacillus and Enterococcus strains from human salivary and fecal sources. Front. Microbiol. 2020, 11, 597946. [Google Scholar] [CrossRef]
- Sakamoto, K.; Hirose, H.; Onizuka, A.; Hayashi, M.; Futamura, N.; Kawamura, Y.; Ezaki, T. Quantitative study of changes in intestinal morphology and mucus gel on total parenteral nutrition in rats. J. Surg. Res. 2000, 94, 99–106. [Google Scholar] [CrossRef]
- Uni, Z.; Platin, R.; Sklan, D. Cell proliferation in chicken intestinal epithelium occurs both in the crypt and along the villus. J. Comp. Physiol. B 1998, 168, 241–247. [Google Scholar] [CrossRef]
- Moreno, Y.; Herńandez, M.; Ferrús, M.A.; Alonso, J.L.; Botella, S.; Montes, R.; Hernández, J. Direct detection of thermotolerant Campylobacters in chicken products by PCR and in situ hybridization. Res. Microbiol. 2001, 152, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Clark, C.G.; Taylor, T.M.; Pucknell, C.; Barton, C.; Price, L.; Woodward, D.L.; Rodgers, F.G. Colony multiplex PCR assay for identification and differentiation of Campylobacter jejuni, C. coli, C. lari, C. upsaliensis, and C. fetus subsp. fetus. J. Clin. Microbiol. 2002, 40, 4744–4747. [Google Scholar] [CrossRef]
- Siwek, M.; Slawinska, A.; Stadnicka, K.; Bogucka, J.; Dunislawska, A.; Bednarczyk, M. Prebiotics and synbiotics—In ovo delivery for improved lifespan condition in chicken. BMC Vet. Res. 2018, 14, 402. [Google Scholar] [CrossRef] [PubMed]
- Uni, Z.; Farket, P.R. Enhancement of Development of Oviparous Species by in Ovo Feeding. U.S. Patent No. 6,592,878, 15 July 2003. [Google Scholar]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Shafi, M.E.; Qattan, S.Y.A.; Batiha, G.E.; Khafaga, A.F.; Abdel-Moneim, A.-M.E.; Alagawany, M. Probiotics in poultry feed: A comprehensive review. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1835–1850. [Google Scholar] [CrossRef]
- Fathima, S.; Shanmugasundaram, R.; Adams, D.; Selvaraj, R.K. Gastrointestinal microbiota and their manipulation for improved growth and performance in chickens. Foods 2022, 11, 1401. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.R.; Wilson, K.M.; Trombetta, M.; Briggs, W.N.; Duff, A.F.; Chasser, K.M.; Bottje, W.G.; Bielke, L. A proteomic view of the cross-talk between early intestinal microbiota and poultry immune system. Front. Physiol. 2020, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, J.E.; Van Der Hoeven-Hangoor, E.; Van De Linde, I.B.; Montijn, R.C.; Van Der Vossen, J.M.B.M. In ovo inoculation of chicken embryos with probiotic bacteria and its effect on posthatch Salmonella susceptibility. Poult. Sci. 2014, 93, 818–829. [Google Scholar] [CrossRef]
- Willemsen, H.; Debonne, M.; Swennen, Q.; Everaert, N.; Careghi, C.; Han, H.; Bruggeman, V.; Tona, K.; Decuypere, E. Delay in feed access and spread of hatch: Importance of early nutrition. World’s Poult. Sci. J. 2010, 66, 177–188. [Google Scholar] [CrossRef]
- Vieira, S.L.; Moran, E.T. Effect of egg origin and chick post-hatch nutrition on broiler live performance and meat yields. World’s Poult. Sci. J. 1999, 56, 125–142. [Google Scholar] [CrossRef]
- Wilson, F.D.; Cummings, T.S.; Barbosa, T.M.; Williams, C.J.; Gerard, P.D.; Peebles, E.D. Comparison of two methods for determination of intestinal villus to crypt ratios and documentation of early age-associated ratio changes in broiler chickens. Poult. Sci. 2018, 97, 1757–1761. [Google Scholar] [CrossRef]
- Wijtten, P.J.A.; Langhout, D.J.; Verstegen, M.W.A. Small intestine development in chicks after hatch and in pigs around the time of weaning and its relation with nutrition: A review. Acta Agric. Scand. Sect. A—Anim. Sci. 2012, 62, 1–12. [Google Scholar] [CrossRef]
- Willing, B.P.; Van Kessel, A.G. Enterocyte proliferation and apoptosis in the caudal small intestine is influenced by the composition of colonizing commensal bacteria in the neonatal gnotobiotic pig. J. Anim. Sci. 2007, 85, 3256–3266. [Google Scholar] [CrossRef]
- Ilina, L.A.; Yildirim, E.A.; Nikonov, I.N.; Filippova, V.A.; Laptev, G.Y.; Novikova, N.I.; Grozina, A.A.; Lenkova, T.N.; Manukyan, V.A.; Egorov, I.A.; et al. Metagenomic bacterial community profiles of chicken embryo gastrointestinal tract by using T-RFLP analysis. Dokl. Biochem. Biophys. 2016, 466, 47–51. [Google Scholar] [CrossRef]
- Khan, W.I. Physiological changes in the gastrointestinal tract and host protective immunity: Learning from the mouse-Trichinella spiralis model. Parasitology 2008, 135, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Khan, W.I. Goblet cells and mucins: Role in innate defense in enteric infections. Pathogens 2013, 2, 55–70. [Google Scholar] [CrossRef]
- Khaligh, F.; Hassanabadi, A.; Nassiri-Moghaddam, H.; Golian, A.; Kalidari, G. Effect of probiotic administration route and dietary nutrient density on growth performance, gut health, and some hematological variables in healthy or Eimeria infected broiler chickens. Iran. J. Appl. Anim. Sci. 2019, 5, 305–315. [Google Scholar]
- Awad, W.A.; Ghareeb, K.; Abdel-Raheem, S.; Böhm, J. Effects of dietary inclusion of probiotic and synbiotic on growth performance, organ weights, and intestinal histomorphology of broiler chickens. Poult. Sci. 2009, 88, 49–56. [Google Scholar] [CrossRef]
- Caballero-Franco, C.; Keller, K.; De Simone, C.; Chadee, K. The VSL#3 probiotic formula induces mucin gene expression and secretion in colonic epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G315–G322. [Google Scholar]
- Ariyadi, B.; Harimurti, S. Effect of indigenous probiotics lactic acid bacteria on the small intestinal histology structure and the expression of mucins in the ileum of broiler chickens. Int. J. Poult. Sci. 2015, 14, 276–278. [Google Scholar] [CrossRef]
- Smirnov, A.; Perez, R.; Amit-Romach, E.; Sklan, D.; Uni, Z. Mucin dynamics and microbial populations in chicken small intestine are changed by dietary probiotic and antibiotic growth promoter supplementation. J. Nutr. 2005, 135, 187–192. [Google Scholar] [CrossRef]
- Rantala, M.; Nurmi, E. Prevention of the growth of Salmonella infantis in chicks by the flora of the alimentary tract of chickens. Br. Poult. Sci. 1973, 14, 627–630. [Google Scholar] [CrossRef]
- Roto, S.M.; Kwon, Y.M.; Ricke, S.C. Applications of in ovo technique for the optimal development of the gastrointestinal tract and the potential influence on the establishment of its microbiome in poultry. Front. Vet. Sci. 2016, 17, 63. [Google Scholar] [CrossRef]
- Bielke, L.R.; Elwood, A.L.; Donoghue, D.J.; Donoghue, A.M.; Newberry, L.A.; Neighbor, N.K.; Hargis, B.M. Approach for selection of individual enteric bacteria for competitive exclusion in turkey poults. Poult. Sci. 2003, 82, 1378–1382. [Google Scholar] [CrossRef]
- Sharma, S.; Kulkarni, R.R.; Sharif, S.; Hassan, H.; Alizadeh, M.; Pratt, S.; Abdelaziz, K. In ovo feeding of probiotic lactobacilli differentially alters expression of genes involved in the development and immunological maturation of bursa of Fabricius in pre-hatched chicks. Poult. Sci. 2024, 103, 103237. [Google Scholar] [CrossRef]
- Alizadeh, M.; Bavananthasivam, J.; Shojadoost, B.; Astill, J.; Taha-Abdelaziz, K.; Alqazlan, N.; Boodhoo, N.; Shoja Doost, J.; Sharif, S. In ovo and oral administration of probiotic lactobacilli modulate cell- and antibody-mediated immune responses in newly hatched chicks. Front. Immunol. 2021, 12, 664387. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.P.; Wu, A.M.; Ding, X.M.; Lei, Y.; Bai, J.; Zhang, K.Y.; Chio, J.S. Effects of probiotic-supplemented diets on growth performance and intestinal immune characteristics of broiler chickens. Poult. Sci. 2013, 92, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Pender, C.M.; Kim, S.; Potter, T.D.; Ritzi, M.M.; Young, M.; Dalloul, R.A. In ovo supplementation of probiotics and its effects on performance and immune-related gene expression in broiler chicks. Poult. Sci. 2017, 96, 1052–1062. [Google Scholar] [CrossRef]
- Madej, J.P.; Skonieczna, J.; Siwek, M.; Kowalczyk, A.; Łukaszewicz, E.; Slawinska, A. Genotype-dependent development of cellular and humoral immunity in the spleen and cecal tonsils of chickens stimulated in ovo with bioactive compounds. Poult. Sci. 2020, 99, 4343–4350. [Google Scholar] [CrossRef]
- Lutful Kabir, S.M. The role of probiotics in the poultry industry. Int. J. Mol. Sci. 2009, 10, 3531–3546. [Google Scholar] [CrossRef] [PubMed]
- Robyn, J.; Rasschaert, G.; Hermans, D.; Pasmans, F.; Heyndrickx, M. In Vivo Broiler Experiments to Assess Anti-Campylobacter Jejuni Activity of a Live Enterococcus Faecalis Strain. Poult. Sci. 2013, 92, 265–271. [Google Scholar] [CrossRef]
- Ganan, M.; Martinez-Rodriguez, A.J.; Carrascosa, A.V.; Vesterlund, S.; Salminen, S.; Satokari, R. Interaction of Campylobacter Spp. and Human Probiotics in Chicken Intestinal Mucus. Zoonoses Public Health 2013, 60, 141–148. [Google Scholar] [CrossRef]
- Olsen, M.S.R.; Thøfner, I.; Sandvang, D.; Poulsen, L.L. Research Note: The Effect of a Probiotic E. Faecium 669 Mitigating Salmonella Enteritidis Colonization of Broiler Chickens by Improved Gut Integrity. Poult. Sci. 2022, 101, 102029. [Google Scholar] [CrossRef]
- Lyte, M.; Daniels, K.; Byrd, J.A.; Genovese, K.; Swaggerty, C.L.; Kogut, M.H. Use of a Microbial Endocrinology Designed Dopamine-Producing Probiotic to Control Gut Neurochemical Levels Associated with the Development of Gut Inflammation. Poult. Sci. 2025, 104, 105028. [Google Scholar] [CrossRef]
- Baffoni, L.; Gaggìa, F.; Garofolo, G.; Di Serafino, G.; Buglione, E.; Di Giannatale, E.; Di Gioia, D. Evidence of Campylobacter jejuni reduction in broilers with early synbiotic administration. Int. J. Food Microbiol. 2017, 251, 41–47. [Google Scholar] [CrossRef]
- Kobierecka, P.A.; Wyszyńska, A.K.; Aleksandrzak-Piekarczyk, T.; Kuczkowski, M.; Tuzimek, A.; Piotrowska, W.; Górecki, A.; Adamska, I.; Wieliczko, A.; Bardowski, J.; et al. In vitro characteristics of Lactobacillus spp. strains isolated from the chicken digestive tract and their role in the inhibition of campylobacter colonization. Microbiologyopen 2017, 6, e00512. [Google Scholar] [CrossRef]
- Smialek, M.; Burchardt, S.; Koncicki, A. The influence of probiotic supplementation in broiler chickens on population and carcass contamination with Campylobacter spp.—Field study. Res. Vet. Sci. 2018, 118, 312–316. [Google Scholar] [CrossRef]
- Tabashsum, Z.; Peng, M.; Alvarado-Martinez, Z.; Aditya, A.; Bhatti, J.; Romo, P.B.; Young, A.; Biswas, D. Competitive reduction of poultry-borne enteric bacterial pathogens in chicken gut with bioactive Lactobacillus casei. Sci. Rep. 2020, 10, 16259. [Google Scholar] [CrossRef]
- Neveling, D.P.; Dicks, L.M.T. Probiotics: An antibiotic replacement strategy for healthy broilers and productive rearing. Probiotics Antimicrob. Proteins 2021, 13, 1–11. [Google Scholar] [CrossRef]
- Kpodo, K.R.; Proszkowiec-Weglarz, M. Physiological Effects of in Ovo Delivery of Bioactive Substances in Broiler Chickens. Front. Vet. Sci. 2023, 10, 1124007. [Google Scholar] [CrossRef] [PubMed]
- Muyyarikkandy, M.S.; Schlesinger, M.; Ren, Y.; Gao, M.; Liefeld, A.; Reed, S.; Amalaradjou, M.A. In ovo probiotic supplementation promotes muscle growth and development in broiler embryos. Poult. Sci. 2023, 102, 102744. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterium Species | Primer Sequence (5′→3′) | Product Size (bp) | Reference |
|---|---|---|---|
| Bacillus subtilis | F:AAGTCGAGCGGACAGATGG R:CCAGTTTCCAATGACCCTCCCC | 600 | [27] |
| Enterococcus faecium | F:TTGAGGCAGACCAGATTGACG R:TATGACAGCGACTCCGATTCC | 658 | [26] |
| Lactobacillus fermentum | F:GACCAGCGCACCAAGTGATA R:AGCGTAGCGTTCGTGGTAAT | 129 | [28] |
| Species | Primer Sequence (5′→3′) | Product Size (bp) | Gene | Reference |
|---|---|---|---|---|
| Campylobacter spp. | F:AAGTCGAGCGGACAGATGG R:CCAGTTTCCAATGACCCTCCCC | 439 | 16S rRNA | [32] |
| Campylobacter jejuni | F:ACTTCTTTATTGCTTGCTGC R:GCCACAACAAGTAAAGAAGC | 323 | hipO | [33] |
| Campylobacter coli | F:GTAAAACCAAAGCTTATCGTG R:TCCAGCAATGTGTGCAATG | 126 | glyA | [33] |
| D7 | D21 | D35 | |
|---|---|---|---|
| Cortex/medulla diameter ratio in thymic lobules | |||
| C | 0.95 ± 0.23 | 0.87 ± 0.32 | 0.89 ± 0.26 |
| BAC | 0.98 ± 0.26 | 0.88 ± 0.23 | 0.87 ± 0.26 |
| LAC | 0.96 ± 0.25 | 0.88 ± 0.22 | 0.88 ± 0.30 |
| ENT | 0.97 ± 0.28 | 0.92 ± 0.34 | 0.84 ± 0.24 |
| Cortex/medulla area ratio in bursal follicles | |||
| C | 0.50 ± 0.13 | 0.53 ± 0.13 | 0.57 ± 0.19 |
| BAC | 0.53 ± 0.24 | 0.54 ± 0.19 | 0.55 ± 0.17 |
| LAC | 0.46 ± 0.17 | 0.56 ± 0.13 | 0.56 ± 0.19 |
| ENT | 0.40 ± 0.11 * | 0.53 ± 0.11 | 0.62 ± 0.21 |
| Germinal centers area in spleen (percentage of the field of view) | |||
| C | (not visible) | 2.63 ± 1.77 | 3.49 ± 1.35 |
| BAC | — | 2.19 ± 1.24 | 2.51 ± 1.43 |
| LAC | — | 2.34 ± 1.36 | 2.95 ± 1.94 |
| ENT | — | 2.25 ± 1.67 | 3.23 ± 1.63 |
| Feed Conversion Ratio (FCR) | Mortality in the Flock (%) | Average Slaughter Weight (kg) | Fattening Days | European Broiler Index (EBI) | |
|---|---|---|---|---|---|
| I repetition | |||||
| C | 1.58 | 3.72 | 2.52 | 38 | 404.09 |
| BAC | 1.59 | 4.14 | 2.60 | 38 | 412.50 |
| LAC | 1.58 | 4.71 | 2.54 | 38 | 403.12 |
| ENT | 1.60 | 4.75 | 2.50 | 38 | 391.65 |
| II repetition | |||||
| C | 1.66 | 2.80 | 3.28 | 50 | 384.11 |
| BAC | 1.71 | 2.90 | 3.14 | 50 | 356.60 |
| LAC | 1.64 | 2.60 | 3.27 | 50 | 388.41 |
| ENT | 1.66 | 4.20 | 3.23 | 50 | 372.81 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madej, J.P.; Woźniak-Biel, A.; Gaweł, A.; Bobrek, K. Boosting Broiler Health and Productivity: The Impact of in ovo Probiotics and Early Posthatch Feeding with Bacillus subtilis, Lactobacillus fermentum, and Enterococcus faecium. Microorganisms 2025, 13, 1219. https://doi.org/10.3390/microorganisms13061219
Madej JP, Woźniak-Biel A, Gaweł A, Bobrek K. Boosting Broiler Health and Productivity: The Impact of in ovo Probiotics and Early Posthatch Feeding with Bacillus subtilis, Lactobacillus fermentum, and Enterococcus faecium. Microorganisms. 2025; 13(6):1219. https://doi.org/10.3390/microorganisms13061219
Chicago/Turabian StyleMadej, Jan P., Anna Woźniak-Biel, Andrzej Gaweł, and Kamila Bobrek. 2025. "Boosting Broiler Health and Productivity: The Impact of in ovo Probiotics and Early Posthatch Feeding with Bacillus subtilis, Lactobacillus fermentum, and Enterococcus faecium" Microorganisms 13, no. 6: 1219. https://doi.org/10.3390/microorganisms13061219
APA StyleMadej, J. P., Woźniak-Biel, A., Gaweł, A., & Bobrek, K. (2025). Boosting Broiler Health and Productivity: The Impact of in ovo Probiotics and Early Posthatch Feeding with Bacillus subtilis, Lactobacillus fermentum, and Enterococcus faecium. Microorganisms, 13(6), 1219. https://doi.org/10.3390/microorganisms13061219