Abstract
Escherichia coli (E. coli) represents a significant etiological agent of colibacillosis in poultry, resulting in considerable economic losses for the global poultry sector. The present study aimed to determine molecular characterization, antibiotic resistance, and biofilm formation of E. coli strains isolated from diseased broilers from four provinces of China. A total of 200 tissue samples were collected from the intestine, liver, crop, heart, and spleen and processed for microbiological examination. Molecular detection of E. coli strains, virulence genes, and serotypes was performed using polymerase chain reaction (PCR). Antibiotic susceptibility testing and biofilm formation were assessed using disk diffusion and 96-well microtiter plate assays. The study retrieved 68% (136/200) of E. coli strains from collected samples. Most of the E. coli strains were resistant to enrofloxacin (56%), followed by cefepime (54%), amoxicillin/clavulanate (52%), streptomycin (50%), ampicillin (48%), clindamycin (47%), kanamycin (41%), polymyxin B (37%), tetracycline (35%), sulfamethoxazole/trimethoprim (33%), ceftazidime (31%), meropenem (4.7%), and florfenicol (2.9%). Similarly, the E. coli strains tested positive for at least one virulence gene and specific serotypes. Among these, O145 was the most prevalent serotype, identified in 22 isolates (16.2%), followed by O8 (12.5%), O102 (11.8%), and O9 (11.0%). The tsh gene (10.2%) was the most prevalent virulence gene. This study found that 47.1% of E. coli strains were biofilm-producing, with 62.5% exhibiting weak biofilm production, 29.7% mild biofilm production, and 7.8% strong biofilm production. Similarly, 24.2% of the E. coli strains were avian pathogenic E. coli strains due to the presence of five or more virulence genes, specifically tsh, ompA, fimC, iss, fyuA, and astA, in a single strain by multiplex PCR. The present study recommends continuous surveillance and effective control measures to reduce the burden of avian pathogenic E. coli-related infections in poultry.
1. Introduction
Escherichia coli (E. coli) is a ubiquitous bacterium that typically predominates in the gut microflora of humans, animals, and birds [1]. Apart from commensal E. coli, various pathogenic strains cause intestinal infections [2]. E. coli in poultry leads to colibacillosis, which is manifested as perihepatitis, pericarditis, air-sacculitis, salpingitis, and peritonitis, potentially progressing to septicemia and death [3]. This is one of the leading causes of mortality and morbidity in poultry, affecting all stages of production and being economically devastating to the industry [4]. It is estimated that at least 30% of the commercial flocks in the United States are affected by colibacillosis at any point, resulting in multi-billion-dollar losses to the poultry industry annually [5,6]. E. coli can infect poultry through various routes. Oral and respiratory pathways are common routes that facilitate bacterial colonization of the gastrointestinal and respiratory tracts, allowing migration to internal organs and causing infection. Infected birds can transmit the bacteria to other birds by contaminating feed and water [3,4,7]. Previously, E. coli was regarded as a secondary pathogen that caused disease outbreaks with concurrent viral infections, improper management practices, or inadequate egg hygiene. However, recent research has identified its potential role as a primary pathogen that causes severe disease and high mortality in the absence of stressors [8,9,10].
The pathogenic attributes of E. coli are facilitated by multiple virulence factors, such as adhesins, invasins, protectins, iron acquisition mechanisms, toxins, and plasmids [11]. These factors enable attachment, invasion, colonization, replication, damage to the host cells, and evasion of the host immune response [12,13]. An array of virulence-associated genes encodes these virulence factors, including papC (pilus associated with pyelonephritis) and tsh (temperature-sensitive hemagglutinin), which are responsible for adhesion, and ibeA (invasion of the brain endothelium protein A), which governs the invasion of the host system. Other genes, such as iutA (aerobactin siderophore receptor) and iroN (salmochelin and catecholate siderophore receptor), facilitate iron acquisition from body fluids. The iss (increased serum survival), ompT, and ompA (outer membrane protease) protect the host immune response. Furthermore, astA (heat-stable enterotoxin), hlyE (putative avian hemolysin), iucD (aerobactin synthesis), vat (vacuolating autotransporter toxin), cvi/cva (structural genes of colicin V operon), fimC (type 1 fimbriae (D-mannose-specific adhesin), and fyuA (ferric yersinia uptake) enable E. coli to produce toxins that damage the host’s tissues [14,15]. Several studies have identified combinations of different virulence genes to predict the disease-causing potential of E. coli strains [16,17]. Virulence genes also significantly contribute in facilitating the biofilm formation by these bacterial species [18].
Biofilm formation can serve as a key survival strategy for bacteria, enabling them to survive within host cells while enhancing their antibiotic resistance. The biofilm is characterized by a polymeric matrix that adheres to the surfaces of bacterial cells [19] and provides numerous benefits to bacteria, such as structural integrity, enhanced adhesion through bacterial adhesins, and protection against immune responses [20]. Moreover, the high diversity of E. coli strains makes it challenging to ensure the accuracy of these predictors, and thus hinders the effective diagnosis, treatment, and prevention of E. coli infections in poultry [21]. The high diversity of E. coli strains is further evident in the number of serotypes established. Serotyping is vital for unraveling E. coli virulence mechanisms [22]. O (lipopolysaccharide) and H types (flagellar antigen) have been widely used to classify E. coli strains. Some of the O serogroups associated with E. coli strains in poultry are O1, O2, O8, O9, O18, O21 [23,24,25,26,27], O78, O102, O128, and O145 [28,29,30]. Serotyping, along with biofilm and virulence detection, is a better predictor of the virulence potential of E. coli [31].
E. coli has been reported to resist various antibiotics, such as tetracyclines, sulphonamides, and aminoglycosides, commonly used in the poultry industry to treat E. coli infections [3]. Antimicrobial resistance and virulence genes are often associated with plasmids that facilitate their transmission between bacteria, and thus need to be continuously monitored [32]. The present study investigated the molecular characterization, antibiotic resistance, and biofilm formation of E. coli strains isolated from tissue samples of diseased broiler birds from four provinces of China.
2. Materials and Methods
2.1. Sample Collection
Samples from four provinces recognized for broiler production were obtained to ensure comprehensive geographical representation and account for potential variations in E. coli strains. These samples may not accurately represent the entire population of affected broilers in these areas. They were chosen from farms with documented histories of colibacillosis outbreaks to enhance the chances of isolating pertinent strains. The samples were taken from broilers exhibiting specific clinical signs of colibacillosis, such as coughing, sneezing, nasal discharge, labored breathing, gasping, yellowish diarrhea, uncoordinated movements, and joint swelling. Characteristic post-mortem lesions, including fibrinous pericarditis, peritonitis, air sacculitis, and hepatomegaly, supported the suspected diagnosis. A licensed veterinarian performed a clinical evaluation to validate the suspicion of colibacillosis before sampling. Only cases detected with distinct clinical symptoms and post-mortem findings of colibacillosis were included in this study, and systemic infections were ruled out through clinical and post-mortem findings. A total of 200 tissue samples of liver (n = 20), spleen (n = 20), heart (n = 20), intestine (n = 120), and crop (n = 20) of colibacillosis-suspected broilers were received from October 2023 to October 2024 at Shanghai Veterinary Research Institute from commercial broiler farms in Liaoning (n = 50), Shandong (n = 50), Fujian (n = 50), and Xinjiang (n = 50) provinces of China. Each province received 5, 5, 5, 30, and 5 samples of liver, spleen, heart, intestine, and crop, respectively. Organ samples were collected from only those birds that had shown symptoms on post-mortem examination. All of the samples were labeled and processed immediately for bacteriological examination.
2.2. Bacterial Isolation and Primary Identification
A tissue sample of approximately 1 g was collected and homogenized in 1 mL of 0.9% sterile normal saline. A small volume of 100 μL was cultured in 3 mL of Luria-Bertani (LB) broth and incubated at 37 °C for 4 to 6 h to enrich the culture. After enrichment, a small inoculum of 100 μL was spread on MacConkey agar plates and incubated at 37 °C for 24 h in an aerobic incubator (Galaxy 48S, New Brunswick, and Eppendorf Company, Enfield, CT, USA). From each organ, 1 to 2 distinct pink colonies exhibiting morphological characteristics typical of E. coli were selected and streaked again on MacConkey agar plates until pure colonies were obtained. Following purification, a single purified colony was picked and streaked on Eosin Methylene Blue (EMB) agar for further phenotypic confirmation of E. coli. The metallic green sheen colonies on EMB agar were picked and cultured again in LB broth under the same incubation conditions to prepare a 20% glycerol stock and for DNA extraction.
2.3. Molecular Identification of E. coli
Bacterial DNA extraction was performed using the Universal DNA Extraction and Purification Kit (Tiangen, Beijing, China) following the manufacturer’s guidelines [33]. Polymerase chain reaction (PCR) was performed to detect E. coli using specific primers targeting the phoA gene (phoA-F 5′-GAAACAAAGCACTATTGCAC-3′, phoA-R 5′-GGCTTTTGTCACAGGGGTAA-3′) of E. coli from an earlier study [27]. Briefly, a 20 µL reaction mixture was prepared, which included primeMix 10 μL, phoA-F 1 μL, phoA-R 1 μL, DNA-free water 7 μL, and DNA template 1 μL. The reaction was carried out with an initial denaturation step at 94 °C for 5 min, followed by 30 cycles of denaturation at 94 °C for 30 s, annealing at 58 °C for 30 s, and extension at 72 °C for 30 s. The final amplification step lasted for 10 min at 72 °C. After amplification, the PCR products were run on a 1% agarose gel at 180 V and a 400-mA current for 25 min. The PCR-positive samples were sent for Sanger sequencing for final confirmation.
2.4. Molecular Identification of E. coli Serotypes and Virulence Genes
All PCR and sequencing confirmed that E. coli was examined for the presence of fifteen virulence genes, including iroN, iutA, iss, ompT, ompA, hlyE, astA, papC, tsh, ibeA, iucD, vat, cvi/cva, fimC, and fyuA (Eurofins Genomics LLC, Louisville, KY, USA). Conventional PCR was employed to detect virulence genes using specific gene primers at varying annealing temperatures (listed in Table 1). Briefly, a 20 µL reaction mixture was prepared, consisting of primeMix 10 μL, 1 μL of forward and reverse primers of each gene, 7 μL of DNA-free water, and 1 μL of DNA template. The reaction commenced with initial denaturation at 94 °C for 5 min, followed by 30 cycles of denaturation at 94 °C for 30 s, at the different annealing temperatures specified in Table 1 for 30 s, and amplification at 72 °C for 30 s. The final amplification step lasted for 10 min at 72 °C. All PCR products were analyzed through agarose gel electrophoresis using a 1% agarose gel (Bio-Rad Laboratories, Inc., Madrid, Spain) and visualized under UV light [34]. Additionally, these E. coli isolates were screened for the presence of O1, O2, O8, O9, O18, O21, O78, O102, O128, and O145 serotypes by PCR [25] that encode for different virulence genes such as iroN, iutA, iss, ompT, ompA, hlyE, astA, papC, tsh, ibeA, iucD, vat, cvi/cva, fimC, and fyuA. The primers for the E. coli serotypes and virulence genes were designed for serotyping and are shown in Table 1.

Table 1.
Primer sequences and annealing temperatures were used for the PCR analysis of virulence-associated genes.
2.5. Antibiotic Susceptibility of E. coli Isolates
All E. coli-positive isolates were analyzed for antibiotic susceptibility testing using the agar disc diffusion assay, consistent with the modified Kirby–Bauer disc diffusion technique. The multidisc dispenser (Abtek Biologicals Ltd., Liverpool, UK), containing various antibiotic discs, was aseptically placed on bacteria-inoculated Mueller–Hinton (MH) agar plates and incubated at 37 °C for 18–20 h. After incubation, the plates were examined for the zone of inhibition. The diameter of the inhibition zones produced by each antibiotic disk was measured using a calibrated Vernier caliper and interpreted according to the Clinical and Laboratory Standards Institute Standards. Antibiotic susceptibility testing was conducted using the following antibiotics from different classes: Lincosamides (Clindamycin, CLI-2 μg), Carbapenems (Meropenem, MEM-10 μg), Cephalosporins (Ceftazidime, CAZ-30 μg; Cefepime, FEP-30 μg), Amphiphenicols (Florfenicol, FFC-30 μg), Penicillin (Ampicillin, AMP-10 μg; Amoxicillin/Clavulanate, AMC 20 μg/10 μg), Aminoglycosides (Kanamycin, KAN-15 μg; Streptomycin, STR-10 μg), Tetracycline (Tetracycline, TCY-30 μg), Polypeptide (Polymyxin B, PB-30 μg), Co-trimoxazole (Sulfamethoxazole/Trimethoprim, SXT-25 μg), and Fluoroquinolone (Enrofloxacin, ENR-5 μg). The data were also analyzed for multidrug-resistant (MDR) strains. The resistance of E. coli isolates to ≥3 different antibiotic classes was referred to as MDR [38].
2.6. Biofilm Formation Assay of E. coli Isolates
Bacterial biofilms were measured in 96-well polyvinyl chloride (PVC) microplates using a crystal violet assay, as described previously [39]. Each strain’s overnight culture was diluted 1:100 in Mueller–Hinton Broth (MH Broth) (Thermo Scientific, Oxoid, UK), supplemented with 0.5% glucose (w/v), and inoculated into 96-well PVC microplates. Biofilms were allowed to develop at 37 °C for 24 h. The cells that adhered to the microtiter wells were stained with crystal violet (0.1%, w/v), subsequently solubilized with 95% ethanol, and the optical density (OD) was measured at 620 nm using a spectrophotometer. The E. coli DH5α (C) was a negative control strain. Biofilm formation capacity was assessed by OD evaluation: OD of the strain the same as or lower than C (negative control), weakly positive (C < strain ≤ 2C), mild positive (2C < strain ≤ 4C), and strongly positive (4C < strain) [40].
2.7. Pathogenicity Assay of E. coli Isolates for Avian Pathogenic Escherichia coli (APEC) Strains
E. coli-positive isolates were screened for key virulence genes (ompA, astA, iss, papC, tsh, ibeA, fimC, and fyuA) using a well-established multiplex PCR assay, and those harboring ≥5 genes were classified as avian pathogenic E. coli (APEC) strains, following the criteria of Johnson et al. [35] and Ewers et al. [41]. Primer sequences and gene definitions are shown in Table 1. Reactions were performed in a 25 μL volume containing 2.5 μL of 10x PCR buffer, 0.4 μL of 50 mm MgCl2, 1.25 μL of dNTP (10 μM) Pool, 2 U Taq DNA polymerase, 0.075 μL (200 μM) of each primer, and 2 μL of DNA sample. The reaction conditions were as follows: the annealing temperature was adjusted to 58 °C, followed by 94 °C for 5 min. The reaction consisted of 30 cycles of 94 °C for 30 s, 63 °C for 30 s, and 68 °C for 10 min, with a final extension step at 72 °C for 10 min.
2.8. Statistical Analysis
The data were saved and processed in Microsoft Excel to analyze percentage prevalence using the formula:
Descriptive statistics were used to analyze the antimicrobial susceptibility data. Moreover, a generalized linear model (GLM) with binomial distribution was used to assess differences in E. coli prevalence across different provinces and tissue types, accounting for unequal sample sizes. Additionally, post hoc pairwise comparisons with Bonferroni correction were applied to determine significant differences among groups. Analyses were performed using the stats model package in Python 3.12. A p-value < 0.05 was considered statistically significant and vice versa [17]. The graphs were made using GraphPad Prism version 8.2.1.
3. Results
3.1. Detection of E. coli isolates
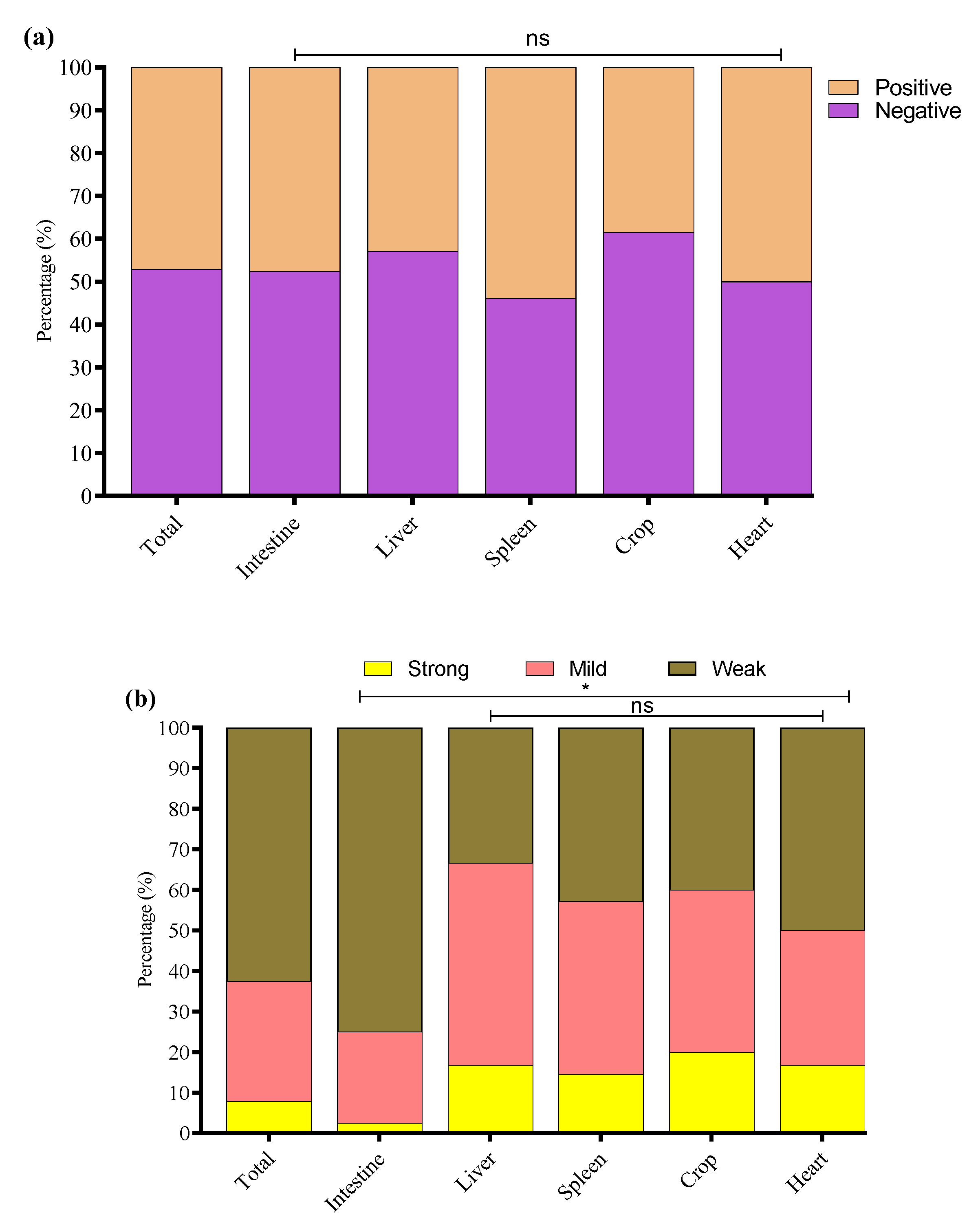
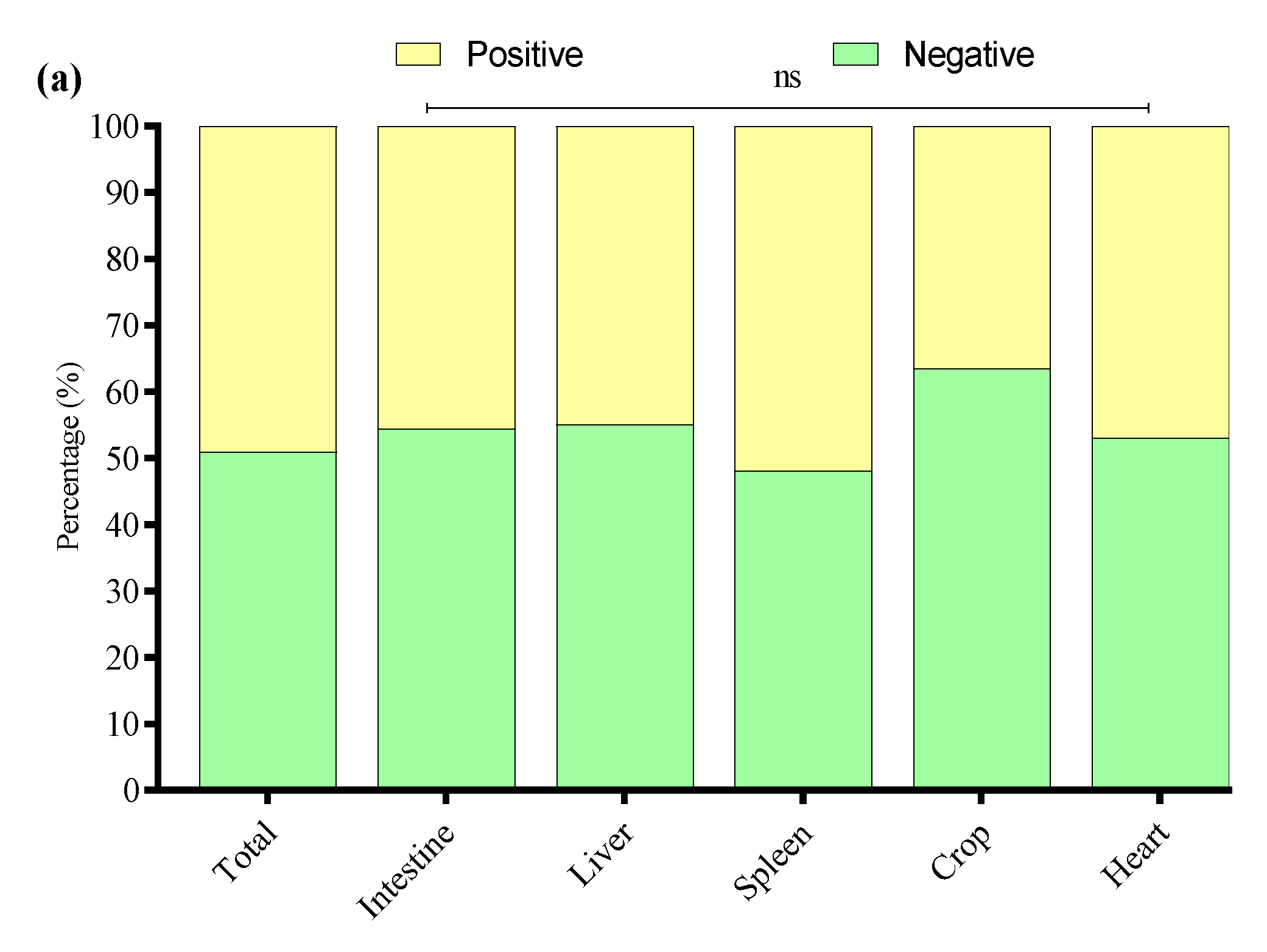
In this study, 200 tissue samples were collected from broiler birds suspected of having colibacillosis across four provinces of China. The study found that 68% (136/200) of the total samples tested positive for E. coli via PCR (Supplementary Figure S1). The GLM analysis showed no statistically significant differences in E. coli prevalence among the provinces. The prevalence of E. coli was highest in Xinjiang (80%, 40/50), followed by Fujian (68%, 34/50), Liaoning (64%, 32/50), and Shandong (60%, 30/50) (Figure 1a); the logistic regression model did not reveal statistically significant differences between provinces (p > 0.05 for all comparisons). These results suggest that E. coli prevalence is relatively consistent across geographically diverse poultry farms in China when accounting for sample size variability (Table 2). The GLM model estimates the log-odds of E. coli positivity relative to Fujian.
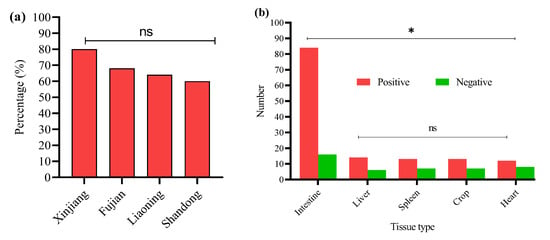
Figure 1.
(a) Percentage of positive E. coli from four Chinese provinces. (b) Distribution of E. coli from different sample types. * Indicates a significant difference; ns indicates a non-significant difference.
Table 2.
Generalized linear model (GLM) estimates for the effect of province on E. coli prevalence in broiler tissue samples.
E. coli was most frequently isolated from intestinal samples n = 84/120, with significantly lower detection in liver n = 14/20, spleen n = 13/20, crop n = 13/20, and heart n = 12/20 (Figure 1b). A GLM (binomial distribution) confirmed a significant effect of tissue type on E. coli prevalence (* p < 0.05), with intestinal isolates showing the highest likelihood of positivity. These findings indicate the intestine as the primary site of E. coli colonization in broilers, with potential systemic dissemination at later stages of infection. However, non-significant differences (ns p > 0.05) were noted among the liver, spleen, crop, and heart.
3.2. Detection of O-Serotypes in E. coli-Positive Samples
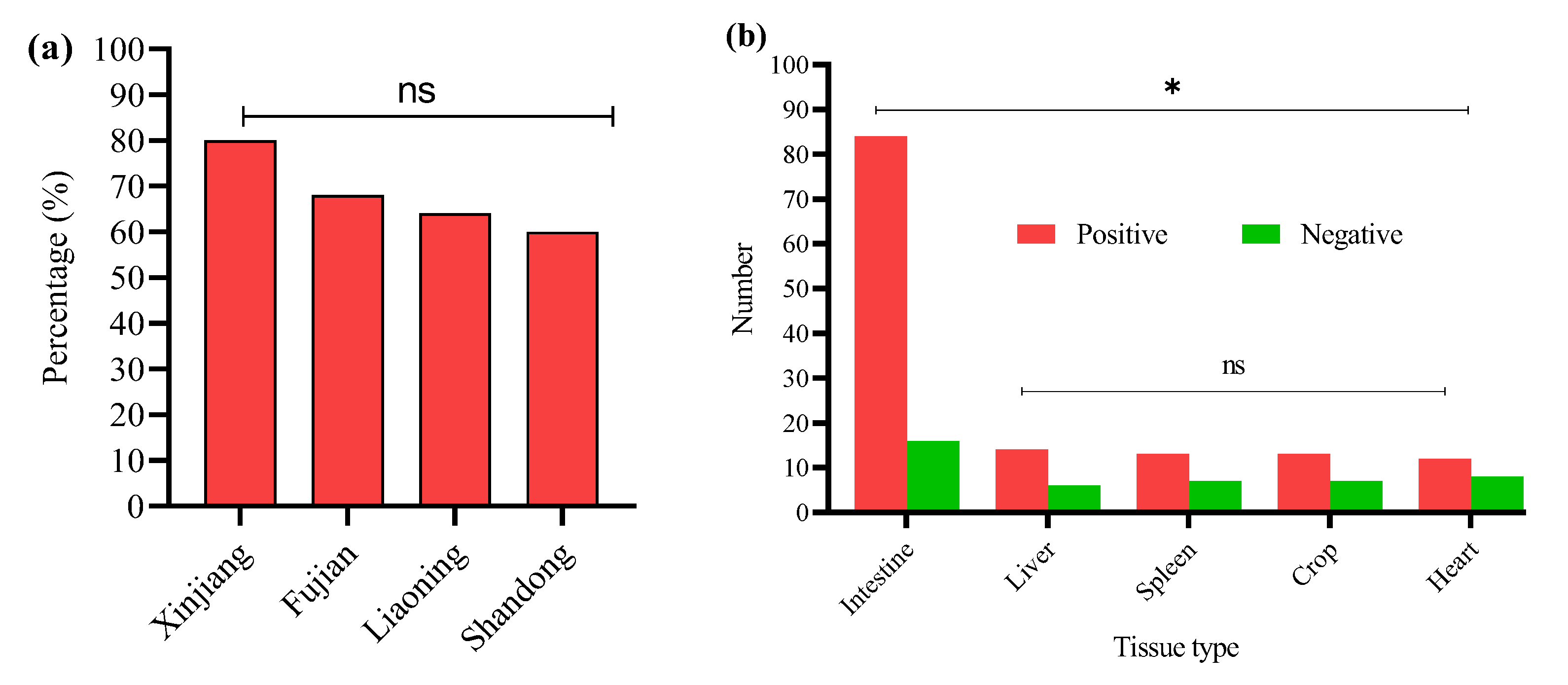
The prevalence of E. coli serotypes was detected by PCR (Supplementary Figure S2). Using a PCR-based method, we determined the O group of 136 E. coli isolates. The serotype prevalence showed that O145 was the most prevalent serotype, identified in 22 isolates (16.2%), followed by O8 (12.5%), O102 (11.8%), and O9 (11.0%). The O145 serotype was consistently found across all four regions, with the highest occurrence in Shandong (17.6%), and similar levels in Liaoning (15.6%), Fujian (16.7%), and Xinjiang (15%). Other frequently detected serotypes included rfbO18 (10.3%), rfbO2 (8.8%), rfbO78 (8.1%), and rfbO1 (7.4%). Some serotypes, like O128 and O21, were found in fewer isolates and were unevenly distributed across regions (Figure 2).
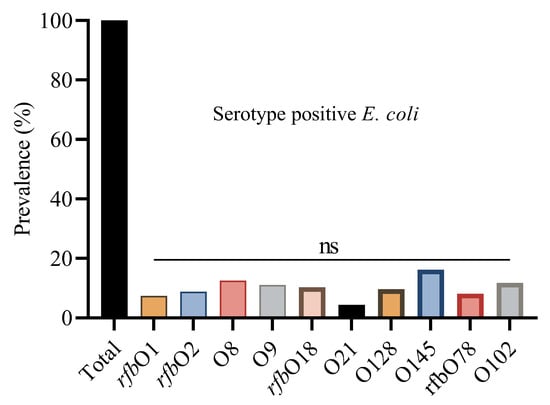
Figure 2.
Overall percentage prevalence of E. coli serotypes. ns indicates a non-significant difference.
Statistical analysis showed non-significant differences (ns) in serotype distribution among the provinces. This suggests that these serotypes are geographically distributed and may reflect common circulating E. coli strains in Chinese broiler populations (Table 3).
Table 3.
Prevalence of E. coli serotypes in different regions.
3.3. Detection Rate of Virulence Genes in Positive O-Serotype Samples
This study analyzed fifteen virulence genes, i.e., iroN, iutA, iss, ompT, ompA, hlyE, astA, papC, tsh, ibeA, iucD, vat, cvi/cva, fimC, and fyuA in 136 positive serotype E. coli strains using PCR (Supplementary Figure S2). Overall, the positive serotype strains were found positive for at least one of the virulence genes, with tsh (10.2%) being the most prevalent, followed by iss (8.8%), ompA, papC, and fyuA (8% each), astA, ibeA, and fimC (7.3% each), iutA, vat, and hlyE (5.8% each), iucD (5.1%), cvi/cva and iroN (4.4%), and ompT (2.9%) (Table 4).
Table 4.
Prevalence of virulence genes in E. coli strains isolated from four regions of China.
In Xinjiang, the prevalent virulence genes were astA, iss, and ibeA, each at 10%. Similarly, in Shandong, the prevalent genes were tsh and fyuA, each at 11.7%. In Liaoning, the prevalent gene was tsh (12.5%), while in Fujian, the prevalent genes identified were papC, tsh, ibeA, iucD, and cvi/cva, each at 10%.
3.4. Antibiotic Susceptibility Profile
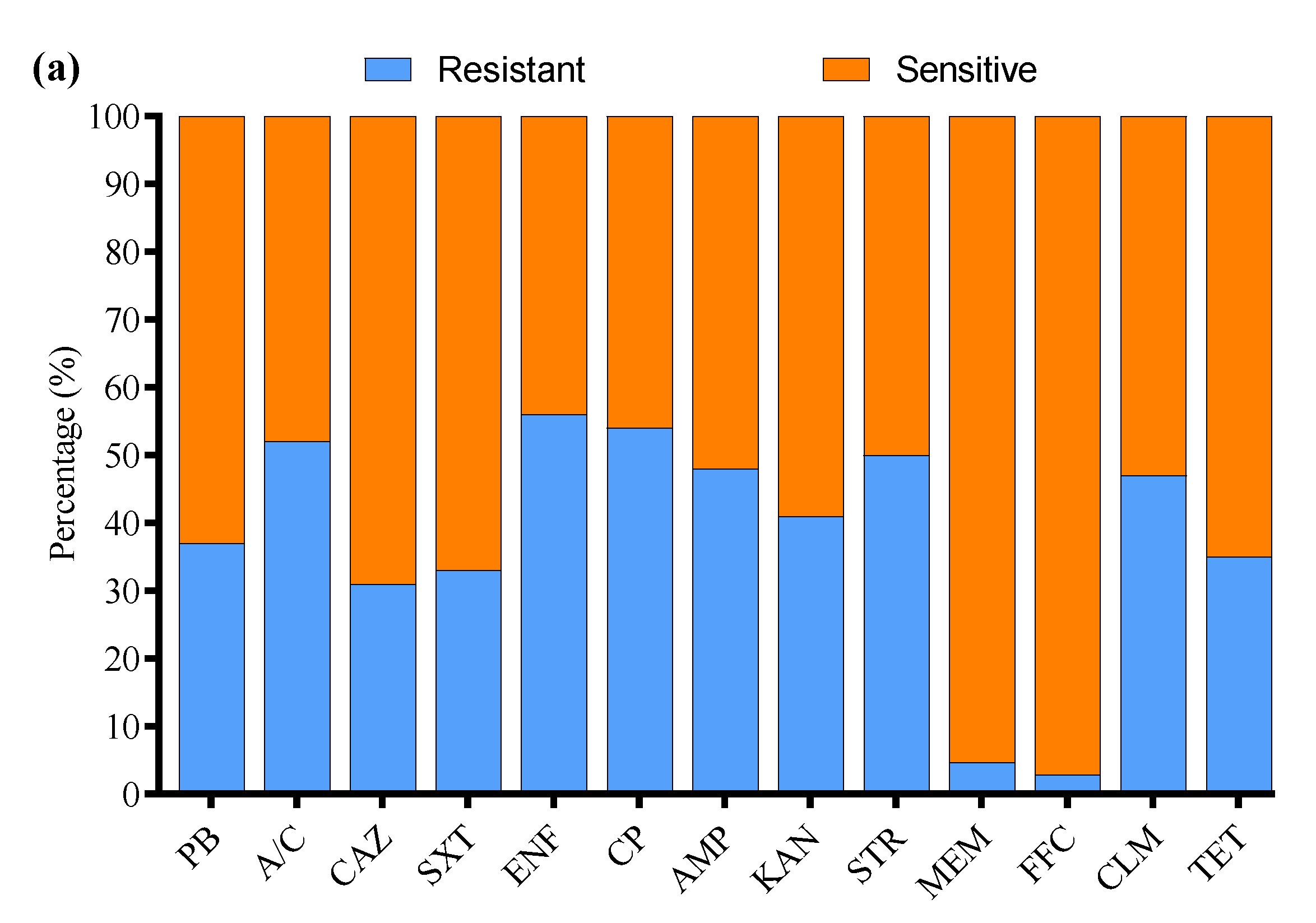
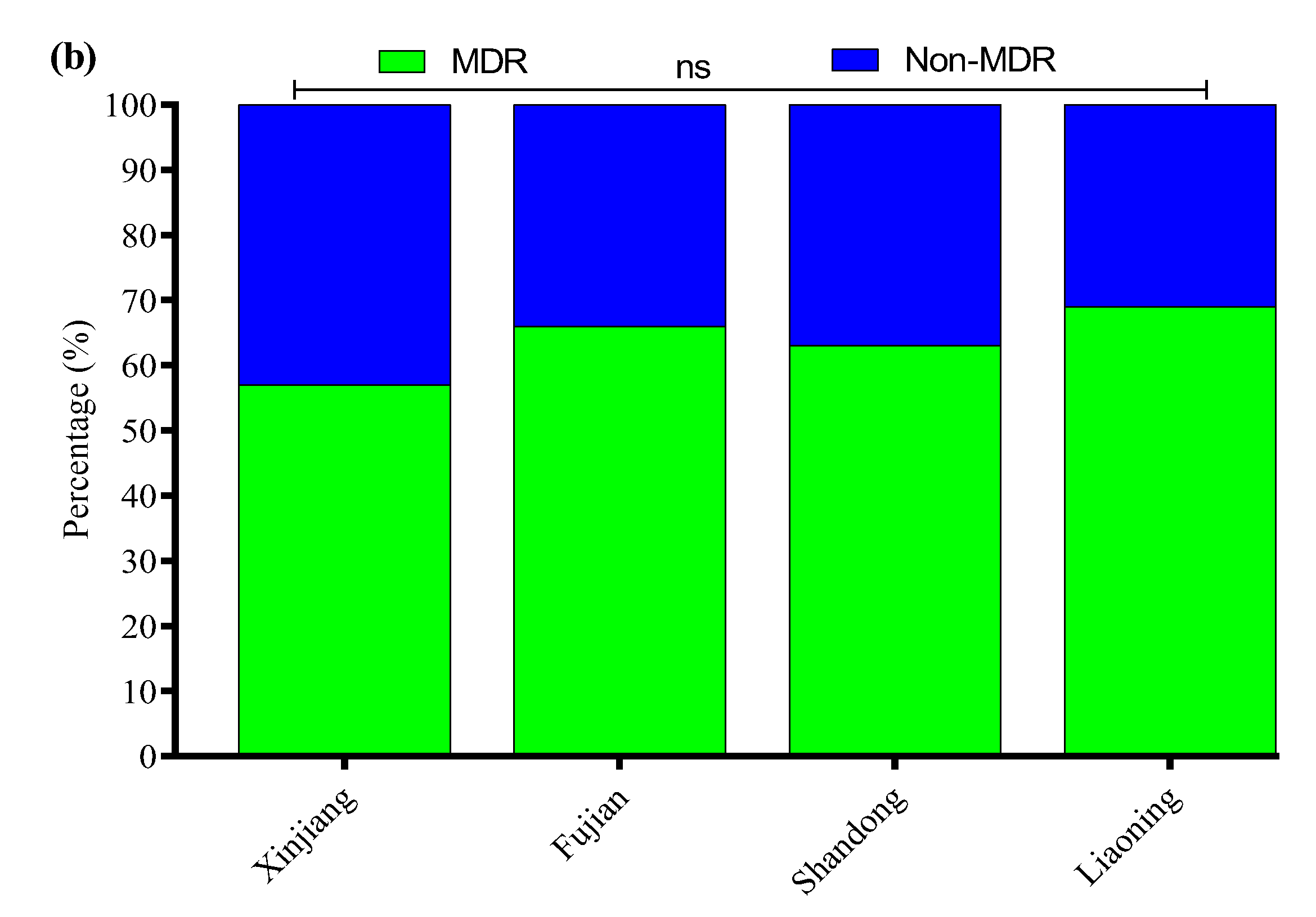
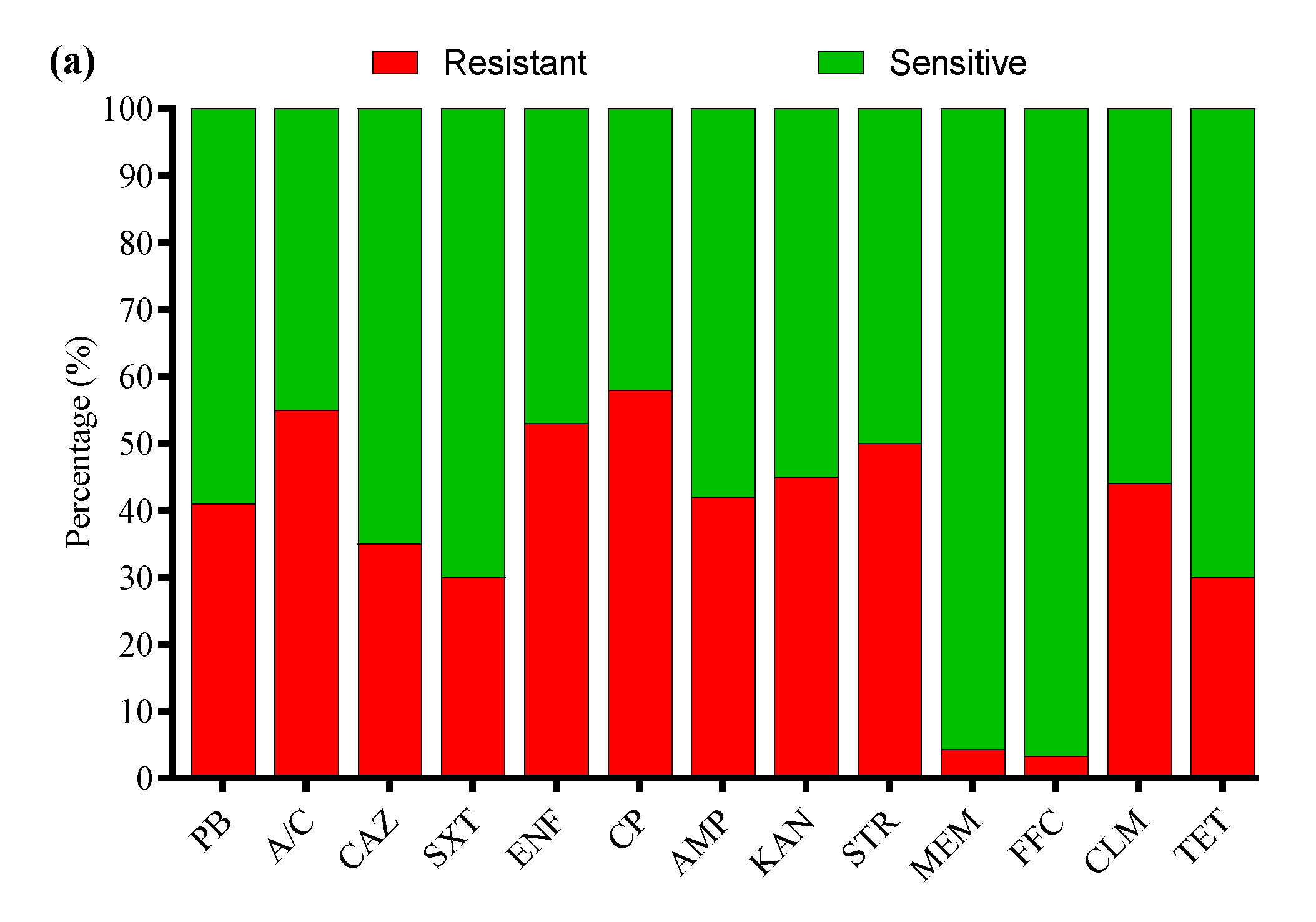
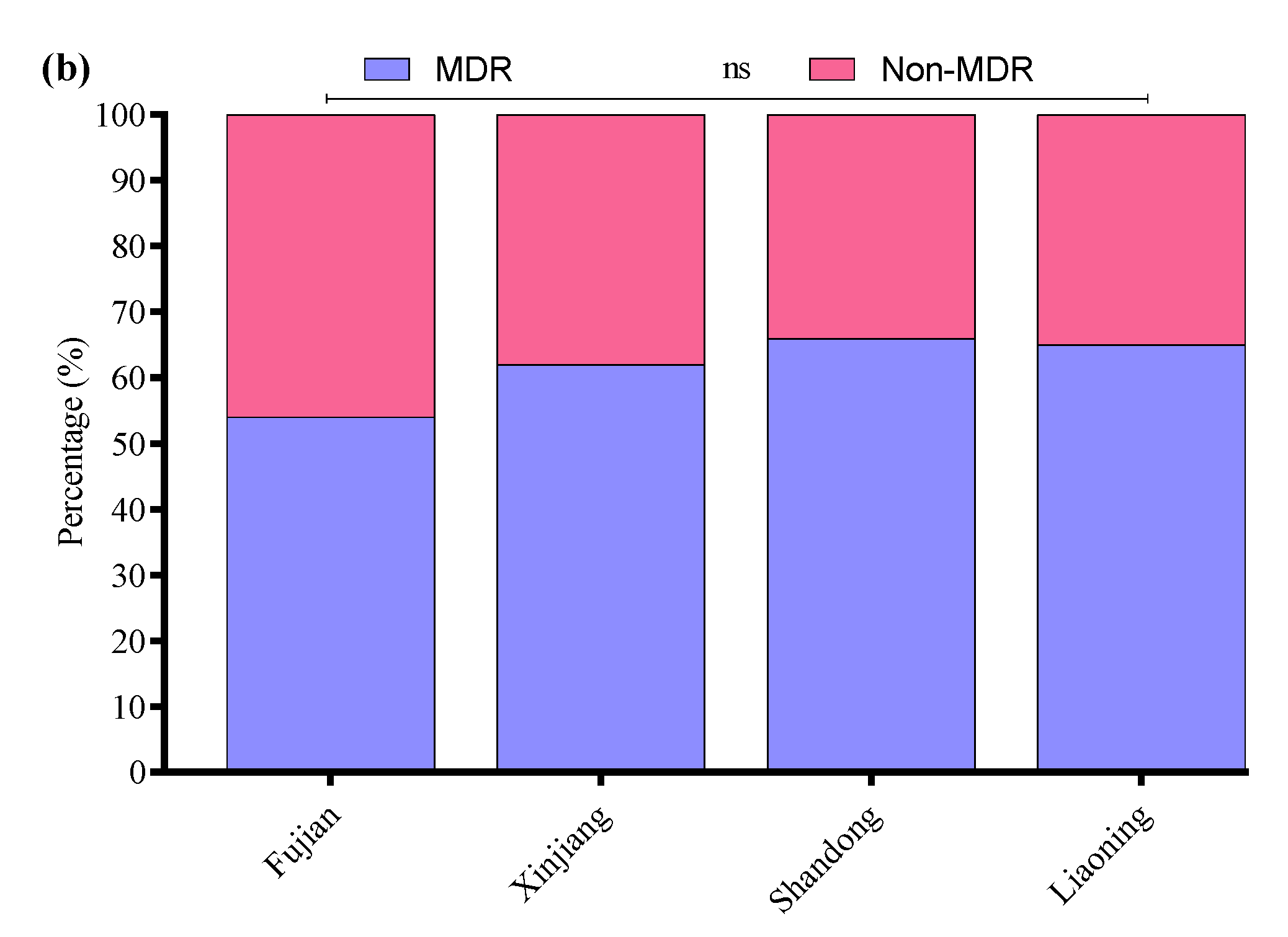
All 136 positive E. coli strains for O-serotypes and virulence genes were subjected to antibiotic susceptibility testing using the Kirby–Bauer disk diffusion assay (Supplementary Figure S3). Most of the E. coli strains were resistant to enrofloxacin (56%), followed by cefepime (54%), amoxicillin/clavulanate (52%), streptomycin (50%), ampicillin (48%), clindamycin (47%), kanamycin (41%), polymyxin B (37%), tetracycline (35%), sulfamethoxazole/trimethoprim (33%), ceftazidime (31%), meropenem (4.7%), and florfenicol (2.9%) (Figure 3a). Moreover, the MDR analysis showed a higher prevalence of MDR E. coli isolates in Liaoning (69%), compared to Fujian (66%), Shandong (63%), and Xinjiang (57%) (Figure 3b).
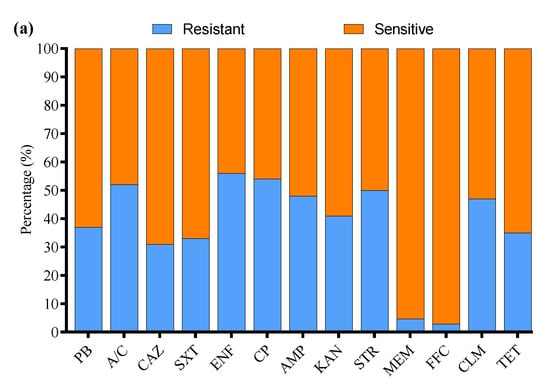
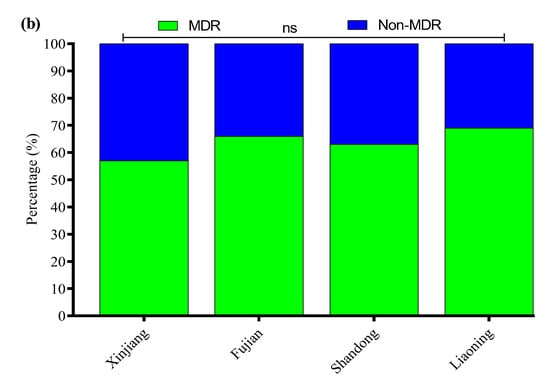
Figure 3.
(a) Antibiotic susceptibility profile of E. coli-positive isolates. (b) Percentage prevalence of MDR E. coli isolates in different provinces of China. ns indicates a non-significant difference.
3.5. Biofilm Formation Ability
The biofilm-forming abilities of 136 positive O-serotypes and virulence genes of E. coli strains were evaluated using a crystal violet 96-well microtiter plate assay (Figure 4).

Figure 4.
Biofilm formation of E. coli isolates: (a) Liaoning spleen E. coli (LSEC 7) strong biofilm, (b) Liaoning liver E. coli (LLEC 7) strong biofilm, (c) Xinjiang intestine E. coli (XIEC 15) strong biofilm, (d) Liaoning heart E. coli (LHEC 10) mild biofilm, (e) Shandong spleen E. coli (SSEC 5) mild biofilm, (f) Shandong heart E. coli (SHEC 7) weak biofilm, (g) Liaoning liver E. coli (LLEC 3) weak biofilm, (h) Xinjiang intestine E. coli (XIEC 9) weak biofilm, and (i) Xinjiang intestine E. coli (XIEC 5) Negative/No biofilm formation.
The study noted that 64/136 (47.1%) E. coli strains were biofilm producers while 72/136 (52.9%) were non-biofilm producers. Among biofilm producers, 47.6% (40/84) were E. coli of intestinal origin, 42.9% (6/14) of liver origin, 53.9% (7/13) of spleen origin, 38.5% (5/13) of crop origin, and 50% (6/12) of heart origin (Figure 5a). Among the biofilm-producing E. coli, 62.5% (40/64) were weak, 29.7% (19/64) were mild, and 7.8% (5/64) were strong biofilm-producing E. coli (Figure 5b). To investigate the potential link between adhesion-related virulence genes and biofilm formation, we conducted a correlation analysis between the presence of papC and tsh genes and the intensity of biofilm formation, categorized as none, weak, mild, or strong. Our findings revealed a weak negative correlation between papC and biofilm formation score (r = −0.048), and a low positive correlation for tsh (r = 0.155). These results indicate that while tsh may have a minor association with increased biofilm production, neither gene alone strongly predicts biofilm formation in the tested strains. This suggests that biofilm formation in E. coli may be governed by multiple factors beyond papC and tsh, requiring further investigation into regulatory pathways and additional genetic contributors. The percentage distribution of weak, mild, and strong biofilm-producing E. coli from different tissues is presented in Figure 5b.
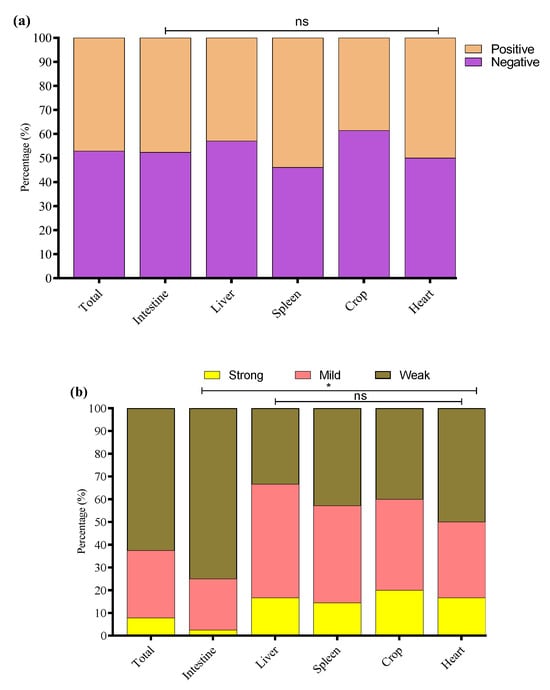
Figure 5.
(a) Overall, biofilm-forming (positive) and non-biofilm-forming (negative) E. coli strains were isolated from different tissues. (b) Percentage of weak, mild, and strong biofilm-producing E. coli among biofilm-positive E. coli in various tissue samples. * Indicates a significant difference; ns indicates a non-significant difference.
3.6. Detection of APEC Strains in E. coli Isolates
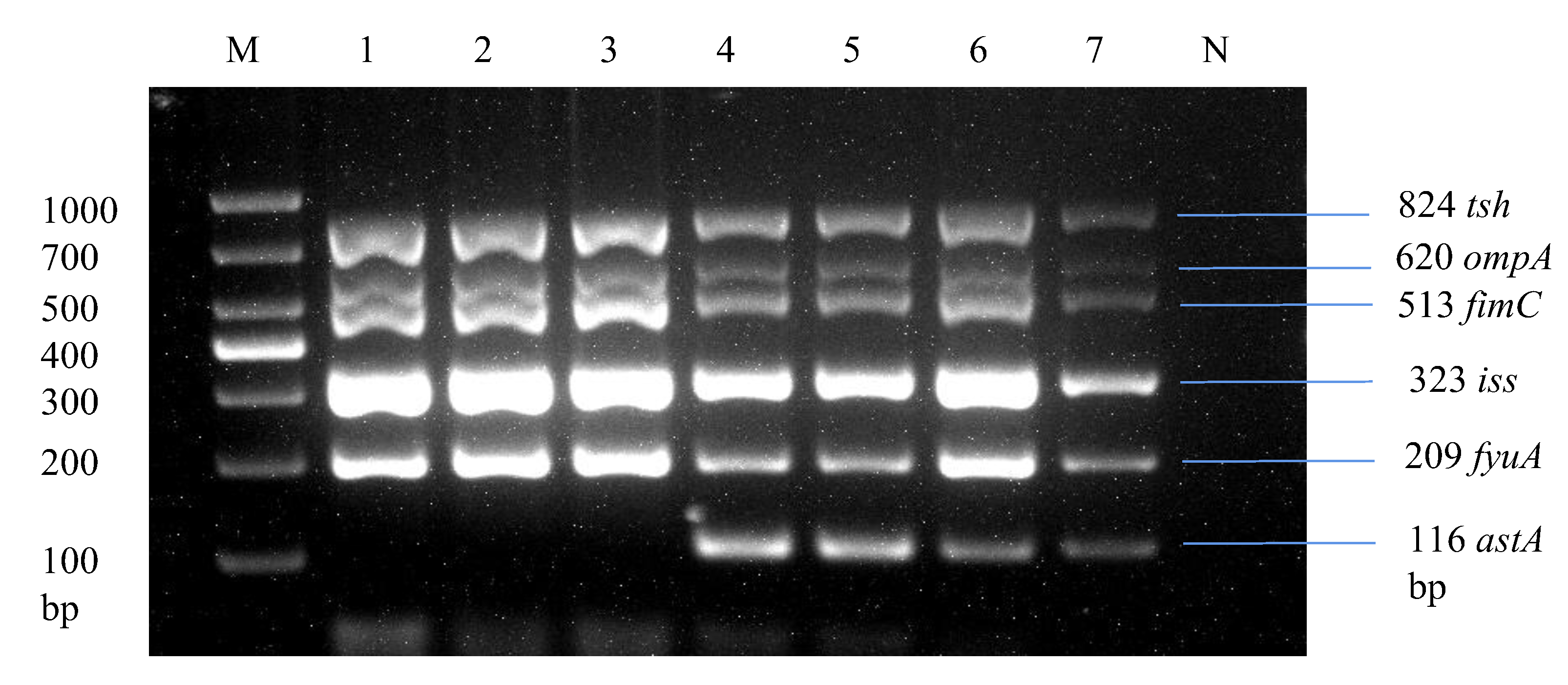
Through multiplex PCR, we analyzed strains by screening the most prevalent virulence genes across E. coli isolates, as shown in Table 3, including ompA, astA, iss, papC, tsh, ibeA, fimC, and fyuA. Overall, 24.2% (33/136) of the E. coli strains were found to be positive for 5 or 6 virulence genes, specifically tsh, ompA, fimC, iss, fyuA, and astA, in a single strain, as illustrated in Figure 6. These strains were classified as avian pathogenic E. coli strains due to the presence of five or more virulence genes of the total eight virulence genes in a single strain, which is the basic criterion for determining a pathogenic strain as per the standards of Johnson et al. [35] and Ewers et al. [41].
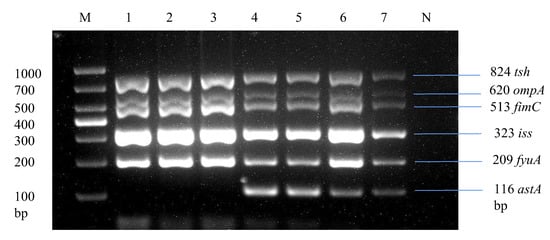
Figure 6.
Development of a multiplex PCR for detecting APEC strains. Lane 1: Liaoning spleen APEC strain. Lane 2: Liaoning liver APEC strain. Lane 3: Xinjiang intestine APEC strain. Lane 1–3: Products of multiplex PCR containing tsh, ompA, fimC, iss, and fyuA virulence genes. Lane 4: Liaoning heart APEC strain. Lane 5: Shandong spleen APEC strain. Lane 6: Fujian intestine APEC strain. Lane 7: Fujian liver APEC strain. Lane 4–7: Products of multiplex PCR containing tsh, ompA, fimC, iss, fyuA, and astA virulence genes.
3.7. Antibiotic Susceptibility Profile of APEC Strains
All 33 APEC strains were subjected to AST using the Kirby–Bauer disk diffusion assay. Most of the APEC strains were resistant to cefepime (58%), followed by amoxicillin/clavulanate (55%), enrofloxacin (53%), streptomycin (50%), kanamycin (45%), clindamycin (44%), ampicillin (42%), polymyxin B (41%), ceftazidime (35%), tetracycline and sulfamethoxazole/trimethoprim (30% each), meropenem (4.3%), and florfenicol (3.3%) (Figure 7a). Moreover, the MDR analysis showed a higher prevalence of MDR APEC strains from Shandong (66%) compared to Liaoning (65%), Xinjiang (62%), and Fujian (54%) (Figure 7b).
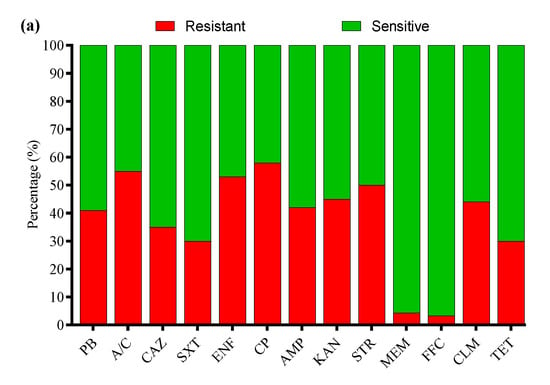
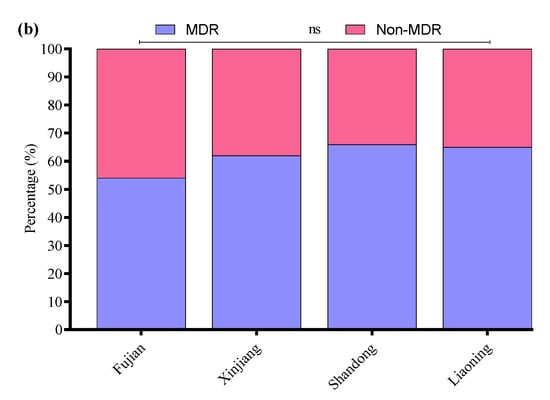
Figure 7.
(a) Antibiotic susceptibility profile of APEC strains. (b) Percentage prevalence of MDR APEC strains in different provinces of China. ns indicates a non-significant difference.
3.8. Biofilm Formation of APEC Strains
The biofilm-forming abilities of 33 APEC strains were evaluated through a crystal violet 96-well microtiter plate assay. The study noted that 49.1% of APEC strains were associated with biofilm production, while 50.9% were not. Among biofilm producers, 45.6% were of intestinal origin, 44.9% of liver origin, 51.9% of spleen origin, 36.5% of crop origin, and 47% of heart origin (Figure 8a). Among the biofilm-producing APEC strains, 62% were weak, 30% were mild, and 8% were strong biofilm producers. The percentage distribution of weak, mild, and strong biofilm-producing APEC strains across different tissues is presented in Figure 8b.
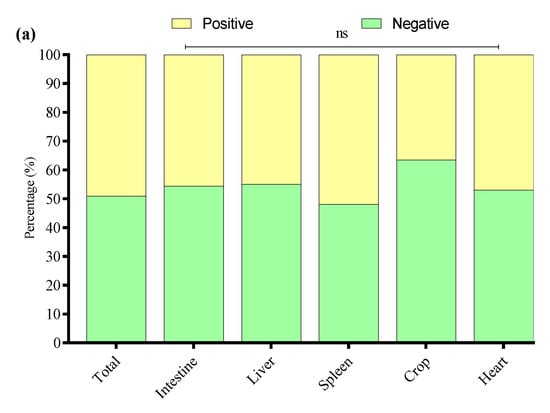
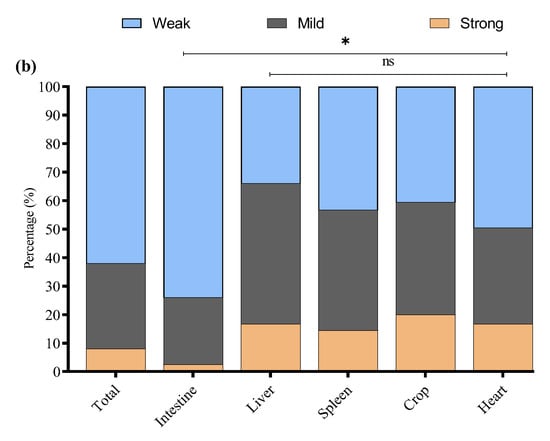
Figure 8.
(a) Overall biofilm forming (positive) and non-biofilm forming (negative) by APEC strains isolated from different tissues. (b) Percentage of weak, mild, and strong biofilm-producing among biofilm-positive APEC strains in various tissue samples. * Indicates a significant difference; ns indicates a non-significant difference.
4. Discussion
E. coli is recognized as a prevalent etiological agent of Gram-negative infections [42]. Extraintestinal pathogenic E. coli (ExPEC) is classified as a facultative pathogen and encompasses several subtypes, including uropathogenic E. coli (UPEC), neonatal meningitis E. coli (NMEC), sepsis-associated E. coli (SEPEC), and avian pathogenic E. coli (APEC). Many studies reported the characteristics of E. coli isolates from layers worldwide [43,44]. However, there is limited information about the E. coli isolates from broiler birds. Recent studies have highlighted the importance of broilers as reservoirs for E. coli infection through vertical transmission to chicks and subsequent horizontal transmission between chicks [45,46]. Moreover, the evolving genetic diversity of E. coli strains requires continuous monitoring among all poultry species [47]. Strains of ExPEC are characterized by the presence of distinct virulence factors (VFs), which include adhesins, toxins, and iron acquisition mechanisms [11,48]. Numerous studies have explored the correlation between antimicrobial resistance and the presence of specific VFs with the capacity of ExPEC strains to form biofilms, which are implicated in urinary tract infections, bloodstream infections, and other extraintestinal conditions.
In this study, we characterize E. coli isolates from broilers with colibacillosis in Fujian, Liaoning, Shandong, and Xinjiang provinces, and also provide information on their genotypic-virulence properties. The prevalence of virulence genes tested in the present study is 68%. The genes encoding for temperature-sensitive hemagglutinin (tsh), outer membrane protease (ompA), type 1 fimbriae (D-mannose-specific adhesin) (fimC), increased serum survival (iss), ferric yersinia uptake (fyuA), and heat-stable enterotoxin (astA) were found to exhibit the highest prevalence among the isolates. The tsh and iss genes are associated with the ColV plasmid [49] and have been identified as genes more predominantly associated with highly avian pathogenic E. coli predictors. A similarly high prevalence of these genes was observed in E. coli isolated from broilers and broiler breeders with colibacillosis from different geographical locations, such as Canada [50], Brazil [51], Egypt [52], Korea [32], and the United States [10].
Similarly, the occurrence of virulence genes analyzed, papC, hlyE, ibeA, and tsh, was also less than the minimal predictors and was similar to that reported in E. coli from broilers in Nepal [36]. The gene encoding the mechanism for adhesion, tsh, was the lowest among the isolates and was comparable to that observed in E. coli isolated from cellulitis lesions in turkeys from Iowa, USA [33]. The current results indicate the virulence-defining nature of the minimal-predictor genes in E. coli isolates from broilers with colibacillosis. E. coli may be classified by somatic (O), capsular (K), and flagellar (H) antigens [53]. Pathogenicity is linked to distinct O-antigen serotypes, i.e., O1, O2, O8, O9, O18, O21, O78, O102, O128, and O145, which have been linked to avian pathogenic E. coli outbreaks, accounting for over 50% of reported cases [37]. Previous epidemiological research found that O1, O2, O18, and O78 accounted for more than 85% of E. coli in Eastern Chinese farms. The O-antigen is an essential element of the lipopolysaccharide (LPS) layer found in the outer membrane of E. coli. Similarly, a previous study reports serogroups from APEC strains, with O78 (16%) and O2 (10%) [54], which were slightly more prevalent compared to our results. Furthermore, another study reported for the first time that O145 may be emerging as a predominant serogroup of APEC in China, with a prevalence of (37.9%), which was higher than that of the other traditional APEC serogroups (O1 (4.7%), O2 (4.7%), O9 (7.1%), O21 (2.3%), O78 (16.7%)). The possible reason for its prevalence and oversight is the failure of vaccines that target the other major serogroups [26]. Similarly, a study reported O8 (37.6%) was the most prevalent serotype of APEC isolated from Wenchang chicken embryos, followed by O9 (16.9%), O102 (6.9%), O128 (3.8%), O21 (3%), and O78 (2.3%) [25]. Serotyping E. coli bacteria in isolated or diseased tissues is critical for disease diagnosis [55], epidemiology, and vaccine development [37].
The complex mechanism of biofilm formation encompasses several genes and regulatory networks. The initial stages of biofilm development, particularly within the first 12 h, are significantly influenced by adhesion genes such as papC and tsh, crucial for transcription and motility [56]. Previous research has examined the relationship between biofilm production and phylogenetic classifications. Notably, phylogroups B2 and D are frequently associated with enhanced biofilm formation, resistance to multiple drugs, a high capacity for iron uptake, and the presence of toxin-related genes [57].
In a previous epidemiological study, a substantial proportion of E. coli isolates derived from urine samples exhibited resistance to combinations of β-lactam antibiotics, β-lactamase inhibitors, quinolones, and cephalosporins, while showing low resistance levels to fosfomycin (2.7%), imipenem (3.2%), and meropenem (3.2%) [58]. The slight difference in our results is due to the frequent use of these antibiotics in poultry farming for therapeutic and prophylactic purposes. In these regions, antibiotics are often used without strict regulations, leading to overuse or misuse, which promotes the development of resistant strains. Furthermore, another study indicated that E. coli isolates demonstrated considerable antibiotic resistance, including quinolones, cephalosporins, aminoglycosides, carbapenems, and penicillin [59]. In Iran, research on uropathogenic E. coli strains revealed a predominant resistance to cefepime (100%) and cephalothin (74%), although these strains remained sensitive to imipenem (100%), vancomycin (100%), and doxycycline (100%) [60]. Additionally, a study conducted in Spain in 2022 collected 376 extraintestinal pathogenic E. coli strains, which exhibited high resistance rates to ciprofloxacin (48.7%), trimethoprim-sulfamethoxazole (47.9%), and ampicillin (38%) [18].
The capacity of isolates to form biofilms was also examined, as the biofilm-forming characteristics of Gram-negative bacteria play a crucial role in their virulence. Our findings reveal that 64 strains exhibited biofilm formation. A 2021 study assessed the biofilm formation capabilities of E. coli strains, reporting that 99% of the isolates demonstrated this ability [61]. Shah et al. identified that 50% of E. coli isolates derived from urine samples form biofilm [62]. Detho et al. also found that over half of the E. coli isolates demonstrate biofilms in vitro [63]. Another study with 126 E. coli isolates established that 80.2% could produce biofilms, with 42.1%, 16.7%, and 21.4% of biofilm producers categorized as weak, medium, and strong, respectively [64]. Additionally, a study explored the correlation between specific virulence factors and the biofilm-forming ability of extraintestinal pathogenic E. coli, concluding that 84.3% of the isolates were capable of biofilm formation. This elevated percentage was suggested to be associated with the virulence genes present in the strains [18]. Although papC and tsh are adhesion-related virulence genes believed to contribute to the early stages of biofilm formation, our findings indicate only a weak association between these genes and biofilm intensity. Specifically, the tsh gene exhibited a weak positive correlation with biofilm score (r = 0.155), while papC showed a weak negative correlation (r = −0.048). Previous studies have reported mixed findings regarding this association. Fattahi et al. demonstrated that papC-positive E. coli strains had a high capacity for biofilm production in urinary tract isolates [65]. Similarly, Laconi et al. found that ESBL/pAmpC-producing E. coli strains from broiler chickens frequently harbored virulence factors associated with enhanced biofilm formation [66]. Biofilms have the potential to develop on the surfaces of water systems, feeders, and drinking apparatus, serving as a continual source of bacterial contamination that proves challenging to eradicate through standard cleaning and disinfection techniques [67]. The ongoing presence of these pathogens can result in chronic infections among poultry and the transmission of zoonotic diseases [68]. The bacteria embedded within biofilms exhibit heightened resistance to antibiotics and disinfectants, complicating control efforts [64]. To address these challenges, farm management practices must incorporate regular cleaning, application of agents that disrupt biofilms, and ongoing monitoring of water and feed systems.
5. Conclusions
The present study reports a high prevalence of E. coli (68%) in colibacillosis-suspected tissue samples from commercial broiler farms in four Chinese provinces. The E. coli strains exhibited significant antibiotic resistance to most antibiotics. Furthermore, the molecular characterization of E. coli strains revealed that 52.9% carried at least one serotype and virulence gene. Similarly, 24.2% of the E. coli strains were classified as avian pathogenic E. coli due to the presence of five or more virulence genes, specifically tsh, ompA, fimC, iss, fyuA, and astA, in a single strain. Moreover, about half of the E. coli strains were identified as biofilm producers. Biofilm formation in E. coli may facilitate the development of more effective therapeutic strategies for managing infections. Biosecurity protocols play a crucial role in the management and prevention of colibacillosis. Implementing vaccination strategies aimed at particular E. coli serogroups, including O1, O2, O8, O9, O18, O21, O78, O102, O128, and O145, can markedly decrease the frequency of infections. Furthermore, alleviating stress through improved environmental conditions and upholding rigorous hygiene and sanitation standards in water supply systems and feeding apparatus are vital for reducing infection risks. When these biosecurity measures are followed, they can significantly enhance the control of colibacillosis in agricultural settings. However, this study has some limitations, particularly regarding the need for a more in-depth examination of the antibiotic resistance mechanisms exhibited by the strains. Further investigation into other genes that may influence biofilm formation is necessary. These findings can also contribute to essential insights into the virulence mechanisms of E. coli and support the formulation of more effective control strategies for this vital poultry pathogen.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms13051017/s1, Figure S1: Molecular identification of E. coli; Figure S2: Molecular detection rate of E. coli virulence genes and serotypes; Figure S3: Antibiotic susceptibility profile of E. coli strains.
Author Contributions
S.N.: Investigation, Methodology, Writing—original draft; M.S., C.H. and Y.B.: Formal analysis, Writing—original draft, Writing—review and editing; Z.W., W.J., W.F., X.W. and L.N.: Investigation, Methodology; Z.C., H.Y. and X.H.: Supervision, Conceptualization, Funding acquisition, Writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research project was financially supported by the National Natural Science Foundation of China (Grant Nos. U22A20518; 32072829; 32302883), Natural Science Foundation of Shanghai (Grant No. 22ZR1475800), and the Natural Science Foundation of Fujian Province of China (Grant No. 2021J01132429).
Institutional Review Board Statement
The experimental procedures and protocols used in this study were approved by the Shanghai Veterinary Research Institute’s Ethical Committee (Approval number: SV-20231015-Go2, 10 February 2023).
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Materials. Further inquiries can be directed to the corresponding authors.
Acknowledgments
The authors appreciate the support of the National Natural Science Foundation of China and the Shanghai Natural Science Foundation of China for funding this research study.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| PB | Polymyxin B |
| A/C | Amoxicillin/Clavulanate |
| CAZ | Ceftazidime |
| SXT | Sulfamethoxazole/Trimethoprim |
| ENF | Enrofloxacin |
| CP | Cefepime |
| AMP | Ampicillin |
| KAN | Kanamycin |
| STR | Streptomycin |
| MEM | Meropenem |
| FFC | Florfenicol |
| CLM | Clindamycin |
| TET | Tetracycline |
References
- Sannes, M.R.; Kuskowski, M.A.; Owens, K.; Gajewski, A.; Johnson, J.R. Virulence factor profiles and phylogenetic background of Escherichia coli isolates from veterans with bacteremia and uninfected control subjects. J. Infect. Dis. 2004, 190, 2121–2128. [Google Scholar] [CrossRef] [PubMed]
- Kathayat, D.; Lokesh, D.; Ranjit, S.; Rajashekara, G. Avian Pathogenic Escherichia coli (APEC): An Overview of Virulence and Pathogenesis Factors, Zoonotic Potential, and Control Strategies. Pathogens 2021, 10, 467. [Google Scholar] [CrossRef] [PubMed]
- Fancher, C.A.; Zhang, L.; Kiess, A.S.; Adhikari, P.A.; Dinh, T.T.N.; Sukumaran, A.T. Avian Pathogenic Escherichia coli and Clostridium perfringens: Challenges in No Antibiotics Ever Broiler Production and Potential Solutions. Microorganisms 2020, 8, 1533. [Google Scholar] [CrossRef] [PubMed]
- Ghunaim, H.; Abu-Madi, M.A.; Kariyawasam, S. Advances in vaccination against avian pathogenic Escherichia coli respiratory disease: Potentials and limitations. Vet. Microbiol. 2014, 172, 13–22. [Google Scholar] [CrossRef]
- Mellata, M. Human and avian extraintestinal pathogenic Escherichia coli: Infections, zoonotic risks, and antibiotic resistance trends. Foodborne Pathog. Dis. 2013, 10, 916–932. [Google Scholar] [CrossRef]
- Johnson, T.J.; Wannemuehler, Y.; Kariyawasam, S.; Johnson, J.R.; Logue, C.M.; Nolan, L.K. Prevalence of avian-pathogenic Escherichia coli strain O1 genomic islands among extraintestinal and commensal E. coli isolates. J. Bacteriol. 2012, 194, 2846–2853. [Google Scholar] [CrossRef]
- Shoaib, M.; Tang, M.; Awan, F.; Aqib, A.I.; Hao, R.; Ahmad, S.; Wang, S.; Shang, R.; Pu, W. Genomic Characterization of Extended-Spectrum β-Lactamase (ESBL) Producing E. coli Harboring blaOXA−1-catB3-arr-3 Genes Isolated From Dairy Farm Environment in China. Transbound. Emerg. Dis. 2024, 17, 3526395. [Google Scholar] [CrossRef]
- Collingwood, C.; Kemmett, K.; Williams, N.; Wigley, P. Is the Concept of Avian Pathogenic Escherichia coli as a Single Pathotype Fundamentally Flawed? Front. Vet. Sci. 2014, 1, 5. [Google Scholar] [CrossRef]
- Agunos, A.; Carson, C.; Léger, D. Antimicrobial therapy of selected diseases in turkeys, laying hens, and minor poultry species in Canada. Can. Vet. J. 2013, 54, 1041–1052. [Google Scholar]
- Newman, D.M.; Barbieri, N.L.; de Oliveira, A.L.; Willis, D.; Nolan, L.K.; Logue, C.M. Characterizing avian pathogenic Escherichia coli (APEC) from colibacillosis cases, 2018. PeerJ 2021, 9, e11025. [Google Scholar] [CrossRef]
- Nawaz, S.; Wang, Z.; Zhang, Y.; Jia, Y.; Jiang, W.; Chen, Z.; Yin, H.; Huang, C.; Han, X. Avian pathogenic Escherichia coli (APEC): Current insights and future challenges. Poult. Sci. 2024, 103, 104359. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; An, C.; Bao, Y.; Zhao, X.; Jernigan, R.L.; Lithio, A.; Nettleton, D.; Li, L.; Wurtele, E.S.; Nolan, L.K.; et al. ArcA Controls Metabolism, Chemotaxis, and Motility Contributing to the Pathogenicity of Avian Pathogenic Escherichia coli. Infect. Immun. 2015, 83, 3545–3554. [Google Scholar] [CrossRef] [PubMed]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, N.L.; Vande Vorde, J.A.; Baker, A.R.; Horn, F.; Li, G.; Logue, C.M.; Nolan, L.K. FNR Regulates the Expression of Important Virulence Factors Contributing to the Pathogenicity of Avian Pathogenic Escherichia coli. Front. Cell Infect. Microbiol. 2017, 7, 265. [Google Scholar] [CrossRef]
- Meng, Q.; Meng, Q.; Wang, S.; Han, X.; Han, Y.; Ding, C.; Dai, J.; Yu, S. Multiplex PCR assay for detection of virulence genes in avian pathogenic Escherichia coli. Wei Sheng Wu Xue Bao 2014, 54, 696–702. [Google Scholar]
- Shoaib, M.; Tang, M.; Aqib, A.I.; Zhang, X.; Wu, Z.; Wen, Y.; Hou, X.; Xu, J.; Hao, R.; Wang, S.; et al. Dairy farm waste: A potential reservoir of diverse antibiotic resistance and virulence genes in aminoglycoside- and beta-lactam-resistant Escherichia coli in Gansu Province, China. Environ. Res. 2024, 263, 120190. [Google Scholar] [CrossRef]
- Shoaib, M.; He, Z.; Geng, X.; Tang, M.; Hao, R.; Wang, S.; Shang, R.; Wang, X.; Zhang, H.; Pu, W. The emergence of multi-drug resistant and virulence gene carrying Escherichia coli strains in the dairy environment: A rising threat to the environment, animal, and public health. Front. Microbiol. 2023, 14, 1197579. [Google Scholar] [CrossRef]
- Ballén, V.; Gabasa, Y.; Ratia, C.; Sánchez, M.; Soto, S. Correlation Between Antimicrobial Resistance, Virulence Determinants and Biofilm Formation Ability Among Extraintestinal Pathogenic Escherichia coli Strains Isolated in Catalonia, Spain. Front. Microbiol. 2022, 12, 803862. [Google Scholar] [CrossRef]
- Rijavec, M.; Müller-Premru, M.; Zakotnik, B.; Žgur-Bertok, D. Virulence factors and biofilm production among Escherichia coli strains causing bacteraemia of urinary tract origin. J. Med. Microbiol. 2008, 57, 1329–1334. [Google Scholar] [CrossRef]
- Limoli, D.H.; Jones, C.J.; Wozniak, D.J. Bacterial Extracellular Polysaccharides in Biofilm Formation and Function. Microbiol. Spectr. 2015, 3, 223–247. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, W.; Zhu, Q.; Shoaib, M.; Chengye, W.; Zhu, Z.; Wei, X.; Bai, Y.; Zhang, J. The prevalent dynamic and genetic characterization of mcr-1 encoding multidrug resistant Escherichia coli strains recovered from poultry in Hebei, China. J. Glob. Antimicrob. Resist. 2024, 38, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.E.; Blanco, M.; Mora, A.; Jansen, W.H.; García, V.; Vázquez, M.L.; Blanco, J. Serotypes of Escherichia coli isolated from septicaemic chickens in Galicia (northwest Spain). Vet. Microbiol. 1998, 61, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Nolan, L.K. Characterizing the APEC pathotype. Vet. Res. 2005, 36, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Knöbl, T.; Moreno, A.M.; Paixão, R.; Gomes, T.A.; Vieira, M.A.; da Silva Leite, D.; Blanco, J.E.; Ferreira, A.J. Prevalence of avian pathogenic Escherichia coli (APEC) clone harboring sfa gene in Brazil. Sci. World J. 2012, 2012, 437342. [Google Scholar] [CrossRef]
- Tian, R.; Xie, F.; Wang, X.; Dai, L.; Wang, J.; Liu, Y.; Zhao, C.; Li, Q.; Zhang, W. Epidemiological investigation and drug resistance analysis of avian pathogenic Escherichia coli (APEC) of Wenchang chickens in Hainan, China. Avian Pathol. 2025. online ahead of print. [Google Scholar] [CrossRef]
- Wang, Z.; Zheng, X.; Guo, G.; Hu, Z.; Miao, J.; Dong, Y.; Xu, Z.; Zhou, Q.; Wei, X.; Han, X.; et al. O145 may be emerging as a predominant serogroup of Avian pathogenic Escherichia coli (APEC) in China. Vet. Microbiol. 2022, 266, 109358. [Google Scholar] [CrossRef]
- Wang, S.; Meng, Q.; Dai, J.; Han, X.; Han, Y.; Ding, C.; Liu, H.; Yu, S. Development of an allele-specific PCR assay for simultaneous sero-typing of avian pathogenic Escherichia coli predominant O1, O2, O18 and O78 strains. PLoS ONE 2014, 9, e96904. [Google Scholar] [CrossRef]
- Ronco, T.; Stegger, M.; Olsen, R.H.; Sekse, C.; Nordstoga, A.B.; Pohjanvirta, T.; Lilje, B.; Lyhs, U.; Andersen, P.S.; Pedersen, K. Spread of avian pathogenic Escherichia coli ST117 O78:H4 in Nordic broiler production. BMC Genom. 2017, 18, 13. [Google Scholar] [CrossRef]
- Doyle, M.P.; Schoeni, J.L. Isolation of Escherichia coli O157:H7 from retail fresh meats and poultry. Appl. Environ. Microbiol. 1987, 53, 2394–2396. [Google Scholar] [CrossRef]
- Cortés, P.; Blanc, V.; Mora, A.; Dahbi, G.; Blanco, J.E.; Blanco, M.; López, C.; Andreu, A.; Navarro, F.; Alonso, M.P.; et al. Isolation and characterization of potentially pathogenic antimicrobial-resistant Escherichia coli strains from chicken and pig farms in Spain. Appl. Environ. Microbiol. 2010, 76, 2799–2805. [Google Scholar] [CrossRef]
- Ishii, N.; Nakahigashi, K.; Baba, T.; Robert, M.; Soga, T.; Kanai, A.; Hirasawa, T.; Naba, M.; Hirai, K.; Hoque, A.; et al. Multiple high-throughput analyses monitor the response of E. coli to perturbations. Science 2007, 316, 593–597. [Google Scholar] [CrossRef]
- Kim, Y.B.; Yoon, M.Y.; Ha, J.S.; Seo, K.W.; Noh, E.B.; Son, S.H.; Lee, Y.J. Molecular characterization of avian pathogenic Escherichia coli from broiler chickens with colibacillosis. Poult. Sci. 2020, 99, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, A.L.; Newman, D.M.; Sato, Y.; Noel, A.; Rauk, B.; Nolan, L.K.; Barbieri, N.L.; Logue, C.M. Characterization of Avian Pathogenic Escherichia coli (APEC) Associated With Turkey Cellulitis in Iowa. Front. Vet. Sci. 2020, 7, 380. [Google Scholar] [CrossRef] [PubMed]
- Ievy, S.; Islam, M.S.; Sobur, M.A.; Talukder, M.; Rahman, M.B.; Khan, M.F.R.; Rahman, M.T. Molecular Detection of Avian Pathogenic Escherichia coli (APEC) for the First Time in Layer Farms in Bangladesh and Their Antibiotic Resistance Patterns. Microorganisms 2020, 8, 1021. [Google Scholar] [CrossRef]
- Johnson, T.J.; Wannemuehler, Y.; Doetkott, C.; Johnson, S.J.; Rosenberger, S.C.; Nolan, L.K. Identification of minimal predictors of avian pathogenic Escherichia coli virulence for use as a rapid diagnostic tool. J. Clin. Microbiol. 2008, 46, 3987–3996. [Google Scholar] [CrossRef]
- Subedi, M.; Luitel, H.; Devkota, B.; Bhattarai, R.K.; Phuyal, S.; Panthi, P.; Shrestha, A.; Chaudhary, D.K. Antibiotic resistance pattern and virulence genes content in avian pathogenic Escherichia coli (APEC) from broiler chickens in Chitwan, Nepal. BMC Vet. Res. 2018, 14, 113. [Google Scholar] [CrossRef]
- Ewers, C.; Janssen, T.; Kiessling, S.; Philipp, H.C.; Wieler, L.H. Molecular epidemiology of avian pathogenic Escherichia coli (APEC) isolated from colisepticemia in poultry. Vet. Microbiol. 2004, 104, 91–101. [Google Scholar] [CrossRef]
- WHO. Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report 2023; WHO: Geneva, Switzerland, 2023. [Google Scholar]
- Hu, J.; Lv, X.; Niu, X.; Yu, F.; Zuo, J.; Bao, Y.; Yin, H.; Huang, C.; Nawaz, S.; Zhou, W.; et al. Effect of nutritional and environmental conditions on biofilm formation of avian pathogenic Escherichia coli. J. Appl. Microbiol. 2022, 132, 4236–4251. [Google Scholar] [CrossRef]
- Mumin, Y.M.; Yüksel, G.; Özad Düzgün, A. Investigation of virulence factor genes and biofilm formation of antibiotic resistant clinical E.coli isolates. Microb. Pathog. 2025, 199, 107257. [Google Scholar] [CrossRef]
- Ewers, C.; Janssen, T.; Kiessling, S.; Philipp, H.C.; Wieler, L.H. Rapid detection of virulence-associated genes in avian pathogenic Escherichia coli by multiplex polymerase chain reaction. Avian Dis. 2005, 49, 269–273. [Google Scholar] [CrossRef]
- Akar, Z.; Akay, S.; Ejder, N.; Özad Düzgün, A. Determination of the Cytotoxicity and Antibiofilm Potential Effect of Equisetum arvense Silver Nanoparticles. Appl. Biochem. Biotechnol. 2024, 196, 909–922. [Google Scholar] [CrossRef] [PubMed]
- McPeake, S.J.; Smyth, J.A.; Ball, H.J. Characterisation of avian pathogenic Escherichia coli (APEC) associated with colisepticaemia compared to faecal isolates from healthy birds. Vet. Microbiol. 2005, 110, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Solà-Ginés, M.; Cameron-Veas, K.; Badiola, I.; Dolz, R.; Majó, N.; Dahbi, G.; Viso, S.; Mora, A.; Blanco, J.; Piedra-Carrasco, N.; et al. Diversity of Multi-Drug Resistant Avian Pathogenic Escherichia coli (APEC) Causing Outbreaks of Colibacillosis in Broilers during 2012 in Spain. PLoS ONE 2015, 10, e0143191. [Google Scholar] [CrossRef]
- Naundrup Thøfner, I.C.; Poulsen, L.L.; Bisgaard, M.; Christensen, H.; Olsen, R.H.; Christensen, J.P. Longitudinal Study on Causes of Mortality in Danish Broiler Breeders. Avian Dis. 2019, 63, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Giovanardi, D.; Campagnari, E.; Ruffoni, L.S.; Pesente, P.; Ortali, G.; Furlattini, V. Avian pathogenic Escherichia coli transmission from broiler breeders to their progeny in an integrated poultry production chain. Avian Pathol. 2005, 34, 313–318. [Google Scholar] [CrossRef]
- Sattar, S.; Bailie, M.; Yaqoob, A.; Khanum, S.; Fatima, K.; Altaf, A.; Ahmed, I.; Shah, S.T.A.; Munawar, J.; Zehra, Q.A.; et al. Characterization of two novel lytic bacteriophages having lysis potential against MDR avian pathogenic Escherichia coli strains of zoonotic potential. Sci. Rep. 2023, 13, 10043. [Google Scholar] [CrossRef]
- Naves, P.; del Prado, G.; Huelves, L.; Gracia, M.; Ruiz, V.; Blanco, J.; Dahbi, G.; Blanco, M.; del Carmen Ponte, M.; Soriano, F. Correlation between virulence factors and in vitro biofilm formation by Escherichia coli strains. Microb. Pathog. 2008, 45, 86–91. [Google Scholar] [CrossRef]
- Johnson, T.J.; Johnson, S.J.; Nolan, L.K. Complete DNA sequence of a ColBM plasmid from avian pathogenic Escherichia coli suggests that it evolved from closely related ColV virulence plasmids. J. Bacteriol. 2006, 188, 5975–5983. [Google Scholar] [CrossRef]
- Varga, C.; Brash, M.L.; Slavic, D.; Boerlin, P.; Ouckama, R.; Weis, A.; Petrik, M.; Philippe, C.; Barham, M.; Guerin, M.T. Evaluating Virulence-Associated Genes and Antimicrobial Resistance of Avian Pathogenic Escherichia coli Isolates from Broiler and Broiler Breeder Chickens in Ontario, Canada. Avian Dis. 2018, 62, 291–299. [Google Scholar] [CrossRef]
- De Carli, S.; Ikuta, N.; Lehmann, F.K.; da Silveira, V.P.; de Melo Predebon, G.; Fonseca, A.S.; Lunge, V.R. Virulence gene content in Escherichia coli isolates from poultry flocks with clinical signs of colibacillosis in Brazil. Poult. Sci. 2015, 94, 2635–2640. [Google Scholar] [CrossRef]
- Li, Y.; Chen, L.; Wu, X.; Huo, S. Molecular characterization of multidrug-resistant avian pathogenic Escherichia coli isolated from septicemic broilers. Poult. Sci. 2015, 94, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Ewers, C.; Li, G.; Wilking, H.; Kiessling, S.; Alt, K.; Antáo, E.M.; Laturnus, C.; Diehl, I.; Glodde, S.; Homeier, T.; et al. Avian pathogenic, uropathogenic, and newborn meningitis-causing Escherichia coli: How closely related are they? Int. J. Med. Microbiol. 2007, 297, 163–176. [Google Scholar] [CrossRef]
- Jamali, H.; Akrami, F.; Bouakkaz, S.; Dozois, C.M. Prevalence of specific serogroups, antibiotic resistance and virulence factors of avian pathogenic Escherichia coli (APEC) isolated from clinical cases: A systematic review and meta-analysis. Microb. Pathog. 2024, 194, 106843. [Google Scholar] [CrossRef]
- Samuel, G.; Reeves, P. Biosynthesis of O-antigens: Genes and pathways involved in nucleotide sugar precursor synthesis and O-antigen assembly. Carbohydr. Res. 2003, 338, 2503–2519. [Google Scholar] [CrossRef]
- Holden, E.R.; Yasir, M.; Turner, A.K.; Wain, J.; Charles, I.G.; Webber, M.A. Massively parallel transposon mutagenesis identifies temporally essential genes for biofilm formation in Escherichia coli. Microb. Genom. 2021, 7, 000673. [Google Scholar] [CrossRef]
- Javed, S.; Mirani, Z.A.; Pirzada, Z.A. Phylogenetic Group B2 Expressed Significant Biofilm Formation among Drug Resistant Uropathogenic Escherichia coli. Libyan J. Med. 2021, 16, 1845444. [Google Scholar] [CrossRef]
- Düzgün, A.; Okumuş, F.; Saral, A.; Çiçek, A.; Cinemre, S. Determination of antibiotic resistance genes and virulence factors in Escherichia coli isolated from Turkish patients with urinary tract infection. Rev. Soc. Bras. Med. Trop. 2019, 52, e20180499. [Google Scholar] [CrossRef]
- Özad Düzgün, A.; Yüksel, G. Detection of virulence factor genes, antibiotic resistance genes and biofilm formation in clinical Gram-negative bacteria and first report from Türkiye of K.oxytoca carrying both blaOXA-23 and blaOXA-51 genes. Biologia 2023, 78, 2245–2251. [Google Scholar] [CrossRef]
- Raeispour, M.; Ranjbar, R. Antibiotic resistance, virulence factors and genotyping of Uropathogenic Escherichia coli strains. Antimicrob. Resist. Infect. Control 2018, 7, 118. [Google Scholar] [CrossRef]
- Naziri, Z.; Kilegolan, J.A.; Moezzi, M.S.; Derakhshandeh, A. Biofilm formation by uropathogenic Escherichia coli: A complicating factor for treatment and recurrence of urinary tract infections. J. Hosp. Infect. 2021, 117, 9–16. [Google Scholar] [CrossRef]
- Shah, T.A.; Preethishree, P.; Ashwini; Pai, V. Bacterial Profile of Urinary Tract Infections: Evaluation of Biofilm Formation and Antibiotic Resistance Pattern of Uropathogenic Escherichia coli. J. Pure Appl. Microbiol. 2020, 14, 2577–2584. [Google Scholar] [CrossRef]
- Detho, H.; Bano, S.; Tunio, S.A.; Abbasi, S.M.; Ahmed, M. Biofilm formation and antibiotic resistance in uropathogenic Escherichia coli: The quest for effective treatment of urinary tract infections. Pure Appl. Biol. (PAB) 2023, 13, 38–46. [Google Scholar] [CrossRef]
- Rastegar, E.; Malekzadegan, Y.; Khashei, R.; Hadi, N. Quinolone resistance and biofilm formation capability of uropathogenic Escherichia coli isolates from an Iranian inpatients’ population. Mol. Biol. Rep. 2023, 50, 8073–8079. [Google Scholar] [CrossRef] [PubMed]
- Fattahi, S.; Kafil, H.S.; Nahai, M.R.; Asgharzadeh, M.; Nori, R.; Aghazadeh, M. Relationship of biofilm formation and different virulence genes in uropathogenic Escherichia coli isolates from Northwest Iran. GMS Hyg. Infect. Control 2015, 10, Doc11. [Google Scholar] [CrossRef] [PubMed]
- Laconi, A.; Tolosi, R.; Apostolakos, I.; Piccirillo, A. Biofilm Formation Ability of ESBL/pAmpC-Producing Escherichia coli Isolated from the Broiler Production Pyramid. Antibiotics 2023, 12, 155. [Google Scholar] [CrossRef]
- Trusz, A.; Gorlach, J.; Gazda, D.; Piekarska, K. Biofilm formation in the drinking water distribution system, on selected pipe materials in flow reactors–preliminary investigations. Ecohydrol. Hydrobiol. 2024, 24, 535–542. [Google Scholar] [CrossRef]
- Araújo, D.; Silva, A.R.; Fernandes, R.; Serra, P.; Barros, M.M.; Campos, A.M.; Oliveira, R.; Silva, S.; Almeida, C.; Castro, J. Emerging Approaches for Mitigating Biofilm-Formation-Associated Infections in Farm, Wild, and Companion Animals. Pathogens 2024, 13, 320. [Google Scholar] [CrossRef]

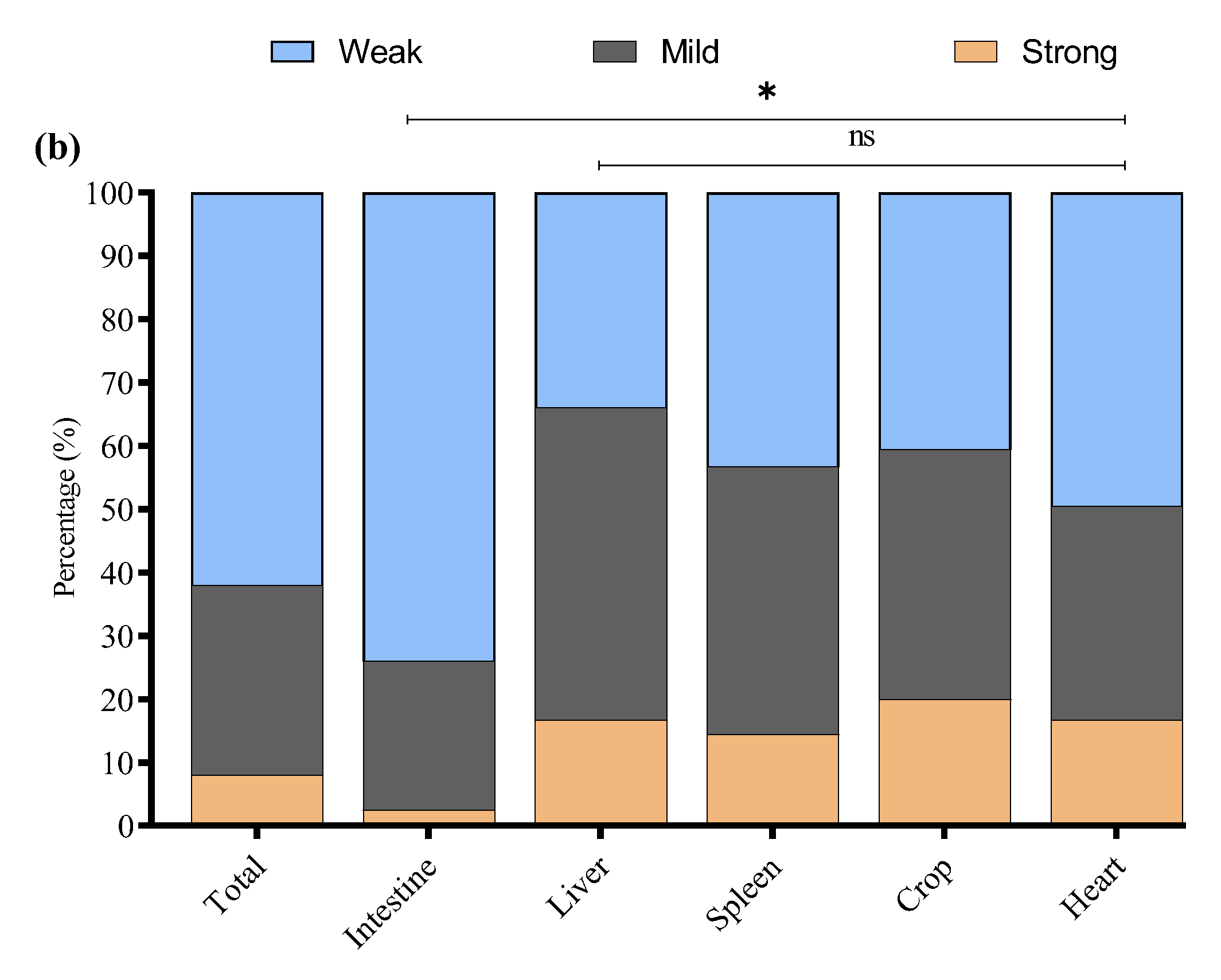
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).