
Induced Mutagenesis and Comparative Genomics of Raoultella sp. 64 for Enhanced Antimony Resistance and Biosorption


Abstract
1. Introduction
2. Materials and Methods
2.1. Screening of Sb-Resistant Bacteria Mutant
2.2. Whole Genome Sequencing, Assembly, and Annotation
2.3. Screening and Analysis of Differential Functional Genes
2.4. Pan-Genome Analyses of Raoultella spp.
2.5. Sb(III) Sorption of Sb-Resistant Bacteria R. ornithinolytica
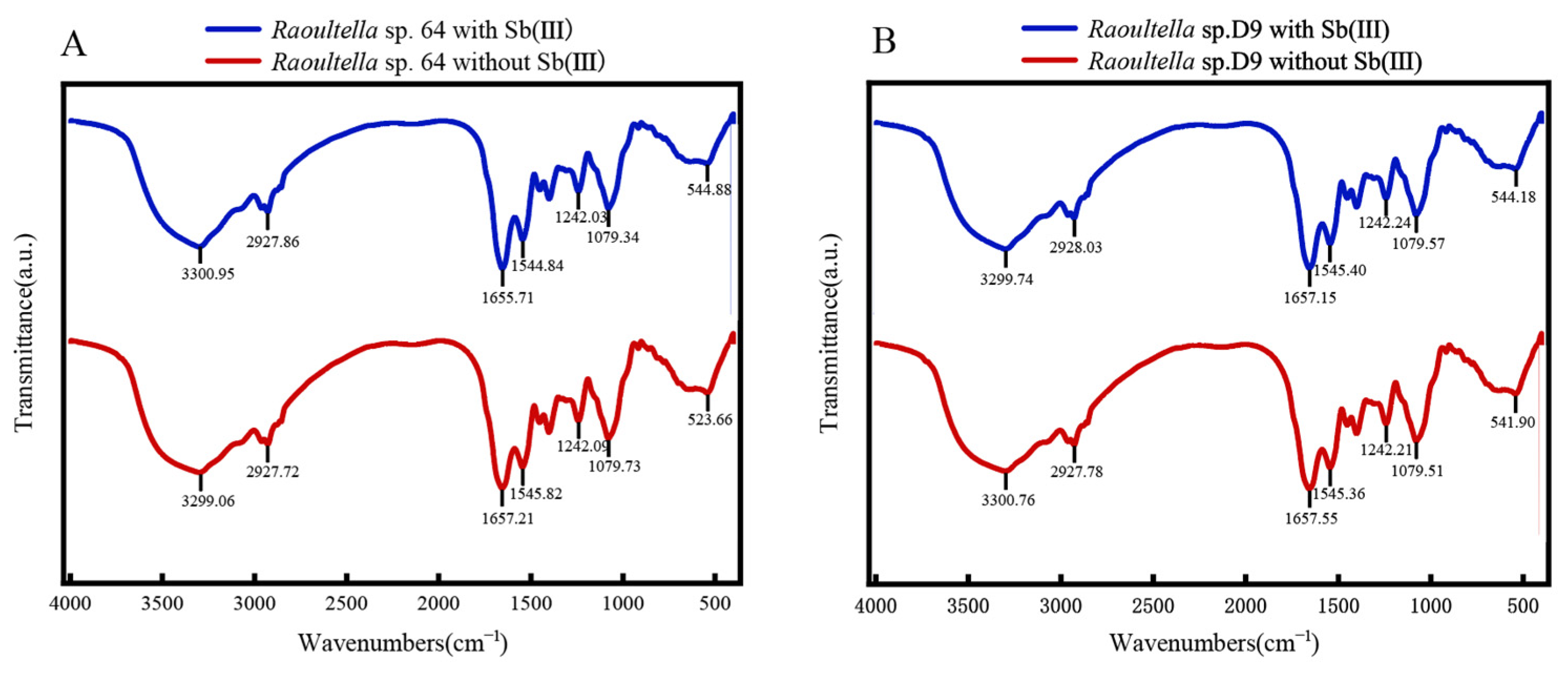
2.6. FTIR and SEM–EDX Analysis
2.7. Data and Statistical Analyses
3. Results
3.1. Screening of Sb(III) Tolerant Strains
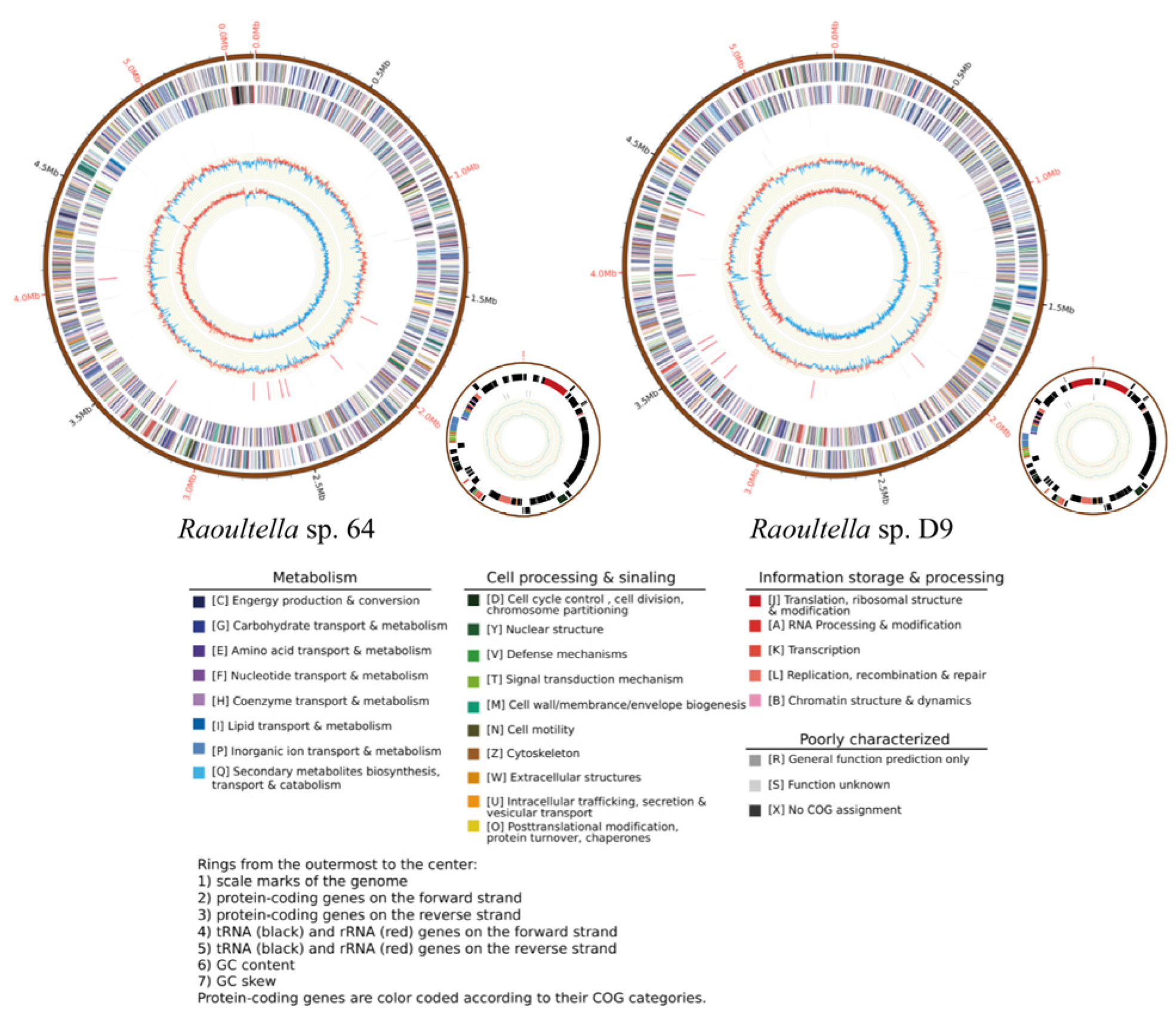
3.2. Genomic Analysis of 64
3.3. Comparative Genomic of Raoultella
3.4. Sorption of Sb(III) by R. ornithinolytica 64 and D9
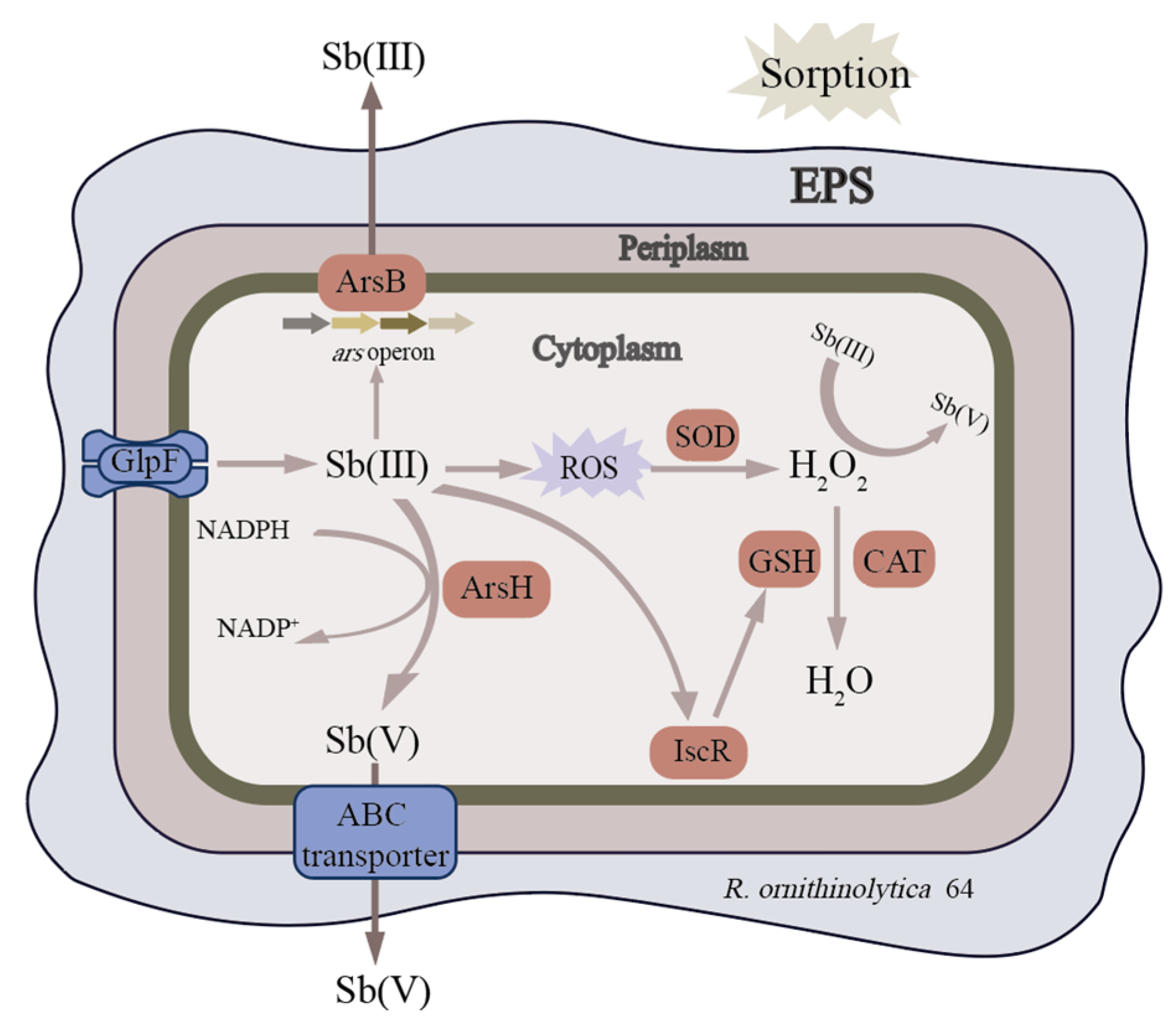
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Courtin-Nomade, A.; Rakotoarisoa, O.; Bril, H.; Grybos, M.; Forestier, L.; Foucher, F.; Kunz, M. Weathering of Sb-rich mining and smelting residues: Insight in solid speciation and soil bacteria toxicity. Geochemistry 2012, 72, 29–39. [Google Scholar] [CrossRef]
- Callahan, M.A. United States Environmental Protection Agency Office of Water Planning and Standards, and United States Environmental Protection Agency Office of Water Planning and Standards Monitoring and Data Support Division. In Water-Related Environmental Fate of 129 Priority Pollutants; Office of Water Planning and Standards, Office of Water and Waste Management, U.S. Environmental Protection Agency: Washington, DC, USA, 1980. [Google Scholar]
- European Council, Council Directive 76/464/EEC of 4 May 1976 on pollution caused by certain dangerous substances discharged into the aquatic environment of the Community, 1976, OJ L 129, 23–29.
- Dupont, D.; Arnout, S.; Jones, P.T.; Binnemans, K. Antimony Recovery from End-of-Life Products and Industrial Process Residues: A Critical Review. J. Sustain. Metall. 2016, 2, 79–103. [Google Scholar] [CrossRef]
- Briki, M.; Yi, Z.; Yang, G.; Shao, M.; Ding, H.; Ji, H. Distribution and health risk assessment to heavy metals near smelting and mining areas of Hezhang, China. Environ. Monit. Assess. 2017, 189, 1–19. [Google Scholar] [CrossRef]
- Filella, M.; Belzile, N.; Chen, Y. Antimony in the environment: A review focused on natural waters: I. Occurrence. Earth-Sci. Rev. 2002, 57, 125–176. [Google Scholar] [CrossRef]
- Nakamaru, Y.M.; Altansuvd, J. Speciation and bioavailability of selenium and antimony in non-flooded and wetland soils: A review. Chemosphere 2014, 111, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gao, Y.; Ning, X.; Li, Z. Research progress and hotspots on microbial remediation of heavy metal-contaminated soil: A systematic review and future perspectives. Environ. Sci. Pollut. Res. 2023, 30, 118192–118212. [Google Scholar] [CrossRef]
- Chen, X.; Yu, T.; Zeng, X. Functional features of a novel Sb(III)- and As(III)-oxidizing bacterium: Implications for the interactions between bacterial Sb(III) and As(III) oxidation pathways. Chemosphere 2024, 352, 141385. [Google Scholar] [CrossRef]
- Li, J. The Mechanisms of Enzymatic and H2O2-Mediated Non-Enzymatic Antimonite Oxidation in Agrobacterium tumefaciens GW4; Huazhong Agricultural University: Wuhan, China, 2017. [Google Scholar]
- Achour, A.R.; Bauda, P.; Billard, P. Diversity of arsenite transporter genes from arsenic-resistant soil bacteria. Res. Microbiol. 2007, 158, 128–137. [Google Scholar] [CrossRef]
- Leuz, A.-K.; Johnson, C.A. Oxidation of Sb(III) to Sb(V) by O2 and H2O2 in aqueous solutions. Geochim. Et Cosmochim. Acta 2005, 69, 1165–1172. [Google Scholar] [CrossRef]
- Li, J.; Yang, B.; Shi, M.; Yuan, K.; Guo, W.; Wang, Q.; Wang, G. Abiotic and biotic factors responsible for antimonite oxidation in Agrobacterium tumefaciens GW4. Sci. Rep. 2017, 7, 43225. [Google Scholar] [CrossRef]
- Zhou, L.; Li, A.; Ma, F.; Yang, J.; Pi, S.; Tang, A. Sb(V) Reduced to Sb(III) and More Easily Adsorbed in the Form of Sb(OH)3 by Microbial Extracellular Polymeric Substances and Core-Shell Magnetic Nanocomposites. ACS Sustain. Chem. Eng. 2019, 7, 10075–10083. [Google Scholar] [CrossRef]
- Lee, H.; Popodi, E.; Tang, H.; Foster, P.L. Rate and molecular spectrum of spontaneous mutations in the bacterium Escherichia coli as determined by whole-genome sequencing. Proc. Natl. Acad. Sci. USA 2012, 109, E2774–E2783. [Google Scholar] [CrossRef] [PubMed]
- Cronan, J.E., Jr.; Ray, T.K.; Vagelos, P.R. Selection and Characterization of an E. coli Mutant Defective in Membrane Lipid Biosynthesis. Proc. Natl. Acad. Sci. USA 1970, 65, 737–744. [Google Scholar] [CrossRef]
- Nyabako, B.A.; Fang, H.; Cui, F.; Liu, K.; Tao, T.; Zan, X.; Sun, W. Enhanced Acid Tolerance in Lactobacillus acidophilus by Atmospheric and Room Temperature Plasma (ARTP) Coupled with Adaptive Laboratory Evolution (ALE). Appl. Biochem. Biotechnol. 2020, 191, 1499–1514. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Zhang, X.; Dou, Y.; Bao, X. Whole genome sequencing and comparative genomics analysis of a new pyrroloquinoline quinone-producing methylotroph. Microbiol. China 2020, 47, 4182–4195. [Google Scholar] [CrossRef]
- Eichinger, R.; Garny, H.; Šácha, P.; Danker, J.; Dietmüller, S.; Oberländer-Hayn, S. Effects of missing gravity waves on stratospheric dynamics; part 1: Climatology. Clim. Dyn. 2020, 54, 3165–3183. [Google Scholar] [CrossRef]
- Li, J.; Wang, Q.; Zhang, S.; Qin, D.; Wang, G. Phylogenetic and genome analyses of antimony-oxidizing bacteria isolated from antimony mined soil. Int. Biodeterior. Biodegrad. 2013, 76, 76–80. [Google Scholar] [CrossRef]
- Delcher, A.L.; Bratke, K.A.; Powers, E.C.; Salzberg, S.L. Identifying bacterial genes and endosymbiont DNA with Glimmer. Bioinformatics 2007, 23, 673–679. [Google Scholar] [CrossRef]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A Program for Improved Detection of Transfer RNA Genes in Genomic Sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Stærfeldt, H.-H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Kalvari, I.; Argasinska, J.; Quinones-Olvera, N.; Nawrocki, E.P.; Rivas, E.; Eddy, S.R.; Bateman, A.; Finn, R.D.; Petrov, A.I. Rfam 13.0: Shifting to a genome-centric resource for non-coding RNA families. Nucleic Acids Res. 2018, 46, D335–D342. [Google Scholar] [CrossRef] [PubMed]
- Huerta-Cepas, J.; Forslund, K.; Coelho, L.P.; Szklarczyk, D.; Jensen, L.J.; von Mering, C.; Bork, P. Fast Genome-Wide Functional Annotation through Orthology Assignment by eggNOG-Mapper. Mol. Biol. Evol. 2017, 34, 2115–2122. [Google Scholar] [CrossRef] [PubMed]
- Pal, C.; Bengtsson-Palme, J.; Rensing, C.; Kristiansson, E.; Larsson, D.G.J. BacMet: Antibacterial biocide and metal resistance genes database. Nucleic Acids Res. 2014, 42, D737–D743. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, L.; Xing, F.; Kudrna, D.A. Extensive sequence divergence between the reference genomes of two elite indica rice varieties Zhenshan 97 and Minghui 63. Proc. Natl. Acad. Sci. USA 2016, 113, E5163–E5171. [Google Scholar] [CrossRef]
- Fitch, W.M. Distinguishing homologous from analogous proteins. Syst. Biol. 1970, 19, 99–113. [Google Scholar] [CrossRef]
- Zhang, D.; Pan, X.; Zhao, L.Y.; Mu, G.-J. Biosorption of antimony (Sb) by the Cyanobacterium Synechocystis sp. Pol. J. Environ. Stud. 2011, 20, 1353–1358. [Google Scholar]
- Loni, P.C.; Wu, M.; Wang, W.; Wang, H.; Ma, L.; Liu, C.; Song, Y.; H Tuovinen, O. Mechanism of microbial dissolution and oxidation of antimony in stibnite under ambient conditions. J. Hazard. Mater. 2020, 385, 121561. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Li, L.-L.; Zhao, S.; Chen, Y.; Yu, A.-F. Bioleaching of metals from spent fluid catalytic cracking catalyst using adapted Acidithiobacillus caldus. Environ. Sci. Pollut. Res. 2023, 30, 125689–125701. [Google Scholar] [CrossRef]
- Rong, Q.; Ling, C.; Lu, D.; Zhang, C.; Zhao, H.; Zhong, K.; Nong, X.; Qin, X. Sb(III) resistance mechanism and oxidation characteristics of Klebsiella aerogenes X. Chemosphere 2022, 293, 133453. [Google Scholar] [CrossRef]
- Deng, R.; Chen, Y.; Deng, X.; Huang, Z.; Zhou, S.; Ren, B.; Jin, G.; Hursthouse, A. A Critical Review of Resistance and Oxidation Mechanisms of Sb-Oxidizing Bacteria for the Bioremediation of Sb(III) Pollution. Front. Microbiol. 2021, 12, 738596. [Google Scholar] [CrossRef]
- Xu, C.; Zhou, T.; Kuroda, M.; Rosen, B.P. Metalloid resistance mechanisms in prokaryotes. J. Biochem. 1998, 123, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Rosen, B.P.; Borbolla, M.G. A plasmid-encoded arsenite pump produces arsenite resistance in Escherichia coli. Biochem. Biophys. Res. Commun. 1984, 124, 760–765. [Google Scholar] [CrossRef]
- Liu, H.; Zhuang, W.; Zhang, S.; Rensing, C.; Huang, J.; Li, J.; Wang, G. Global Regulator IscR Positively Contributes to Antimonite Resistance and Oxidation in Comamonas testosteroni S44. Front. Mol. Biosci. 2015, 2, 70. [Google Scholar] [CrossRef]
- Wang, D.; Zhu, F.; Wang, Q.; Rensing, C.; Yu, P.; Gong, J.; Wang, G. Disrupting ROS-protection mechanism allows hydrogen peroxide to accumulate and oxidize Sb(III) to Sb(V) in Pseudomonas stutzeri TS44. BMC Microbiol. 2016, 16, 279. [Google Scholar] [CrossRef] [PubMed]
- Páez-Espino, A.D.; Nikel, P.I.; Chavarría, M.; de Lorenzo, V. ArsH protects Pseudomonas putida from oxidative damage caused by exposure to arsenic. Environ. Microbiol. 2020, 22, 2230–2242. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Wang, W.; Amanze, C.; Anaman, R.; Fosua, B.A.; Zeng, W. Antimony oxidation and whole genome sequencing of Phytobacter sp. X4 isolated from contaminated soil near a flotation site. J. Hazard. Mater. 2023, 445, 130462. [Google Scholar] [CrossRef]
- Cross, B.; Garcia, A.; Faustoferri, R.; Quivey, R.G. PlsX deletion impacts fatty acid synthesis and acid adaptation in Streptococcus mutans. Microbiology 2016, 162, 662–671. [Google Scholar] [CrossRef]
- Towijit, U.; Songruk, N.; Lindblad, P.; Incharoensakdi, A.; Jantaro, S. Co-overexpression of native phospholipid-biosynthetic genes plsX and plsC enhances lipid production in Synechocystis sp. PCC 6803. Sci. Rep. 2018, 8, 13510. [Google Scholar] [CrossRef]
- Lemire, J.A.; Harrison, J.J.; Turner, R.J. Antimicrobial activity of metals: Mechanisms, molecular targets and applications. Nat. Rev. Microbiol. 2013, 11, 371–384. [Google Scholar] [CrossRef]
- Thakur, M.; Praveen, S.; Divte, P.R.; Mitra, R.; Kumar, M.; Gupta, C.K.; Kalidindi, U.; Bansal, R.; Roy, S.; Anand, A.; et al. Metal tolerance in plants: Molecular and physicochemical interface determines the “not so heavy effect” of heavy metals. Chemosphere 2022, 287, 131957. [Google Scholar] [CrossRef]
- Yang, J.; Calero, C.; Bonomi, M.; Martí, J. Specific Ion Binding at Phospholipid Membrane Surfaces. J. Chem. Theory Comput. 2015, 11, 4495–4499. [Google Scholar] [CrossRef] [PubMed]
- Kiran, I.; Akar, T.; Tunali, S. Biosorption of Pb(II) and Cu(II) from aqueous solutions by pretreated biomass of Neurospora crassa. Process Biochem. 2005, 40, 3550–3558. [Google Scholar] [CrossRef]
- Li, D.; Xu, X.; Yu, H.; Han, X. Characterization of Pb2+ biosorption by psychrotrophic strain Pseudomonas sp. I3 isolated from permafrost soil of Mohe wetland in Northeast China. J. Environ. Manag. 2017, 196, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Shafqat, F.; Bhatt, H.N.; Hanif, M.A.; Zubair, A. Kinetic and equilibrium studies of Cr (III) and Cr (VI) sorption from aqueous solution using Rosa Gruss an Teplitz (red rose) waste biomass. J. Chil. Chem. Soc. 2008, 53, 1667–1672. [Google Scholar] [CrossRef]
- Lu, W.; Shi, J.; Wang, C.; Chang, J. Biosorption of lead, copper and cadmium by an indigenous isolate Enterobacter sp. J1 possessing high heavy-metal resistance. J. Hazard. Mater. 2006, 134, 80–86. [Google Scholar] [CrossRef]
- Li, K.; Ramakrishna, W. Effect of multiple metal resistant bacteria from contaminated lake sediments on metal accumulation and plant growth. J. Hazard. Mater. 2011, 189, 531–539. [Google Scholar] [CrossRef]
- Chakravarty, R.; Banerjee, P.C. Mechanism of cadmium binding on the cell wall of an acidophilic bacterium. Bioresour. Technol. 2012, 108, 176–183. [Google Scholar] [CrossRef]
- Gu, J.; Sunahara, G.; Duran, R.; Yao, J.; Cui, Y.; Tang, C.; Li, H.; Mihucz, V.G. Sb(III)-resistance mechanisms of a novel bacterium from non-ferrous metal tailings. Ecotoxicol. Environ. Saf. 2019, 186, 109773. [Google Scholar] [CrossRef]
- Sanders, O.I.; Rensing, C.; Kuroda, M.; Mitra, B.; Rosen, B.P. Antimonite is accumulated by the glycerol facilitator glpf in Escherichia coli. J. Bacteriol. 1997, 179, 3365–3367. [Google Scholar] [CrossRef]
- Inam, M.A.; Khan, R.; Park, D.R.; Khan, S.; Uddin, A.; Yeom, I.T. Complexation of Antimony with Natural Organic Matter: Performance Evaluation during Coagulation-Flocculation Process. Int. J. Environ. Res. Public Health 2019, 16, 1092. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | R. ornithinolytica 64 | R. ornithinolytica D9 |
|---|---|---|
| Size (bp) | 5,351,241 | 5,351,236 |
| Total genes | 4975 | 4974 |
| Average length (bp) | 947.81 | 948.13 |
| GC% | 55.91 | 55.91 |
| tRNA | 89 | 88 |
| rRNA | 25 | 25 |
| Gene island | 7 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, T.; Cao, S.; Li, X.; Wang, C.; Peng, X. Induced Mutagenesis and Comparative Genomics of Raoultella sp. 64 for Enhanced Antimony Resistance and Biosorption. Microorganisms 2025, 13, 880. https://doi.org/10.3390/microorganisms13040880
Huang T, Cao S, Li X, Wang C, Peng X. Induced Mutagenesis and Comparative Genomics of Raoultella sp. 64 for Enhanced Antimony Resistance and Biosorption. Microorganisms. 2025; 13(4):880. https://doi.org/10.3390/microorganisms13040880
Chicago/Turabian StyleHuang, Tianhua, Shiran Cao, Xiaohan Li, Chuhan Wang, and Xiawei Peng. 2025. "Induced Mutagenesis and Comparative Genomics of Raoultella sp. 64 for Enhanced Antimony Resistance and Biosorption" Microorganisms 13, no. 4: 880. https://doi.org/10.3390/microorganisms13040880
APA StyleHuang, T., Cao, S., Li, X., Wang, C., & Peng, X. (2025). Induced Mutagenesis and Comparative Genomics of Raoultella sp. 64 for Enhanced Antimony Resistance and Biosorption. Microorganisms, 13(4), 880. https://doi.org/10.3390/microorganisms13040880