
The VapBC-4 Characterization Indicates It Is a Bona Fide Toxin-Antitoxin Module of Leptospira interrogans: Initial Evidence for a Role in Bacterial Adaptation

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. In Silico Studies
2.3. Genes and Plasmid Construction
2.4. Protein Synthesis and Purification
2.5. Production of VapB-4 Anti-Serum
Ethical Statement
2.6. E. coli Growth Kinetics
2.7. Pull-Down Assay
2.8. Affinity Dot Blot Assay
2.9. Ribonuclease Activity Assay
2.10. Transcription Studies by RT-qPCR
3. Results
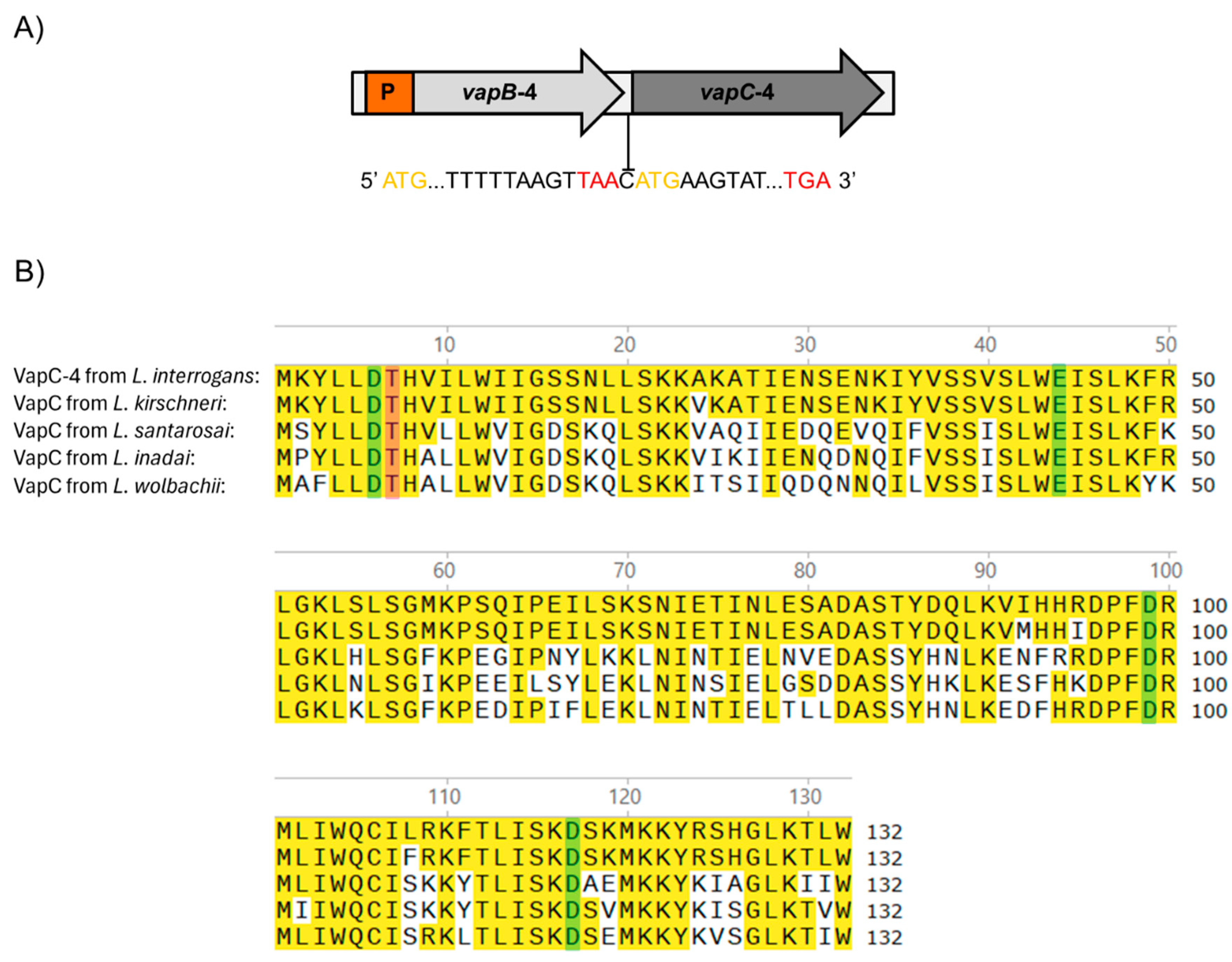
3.1. The VapBC-4 Module from L. interrogans
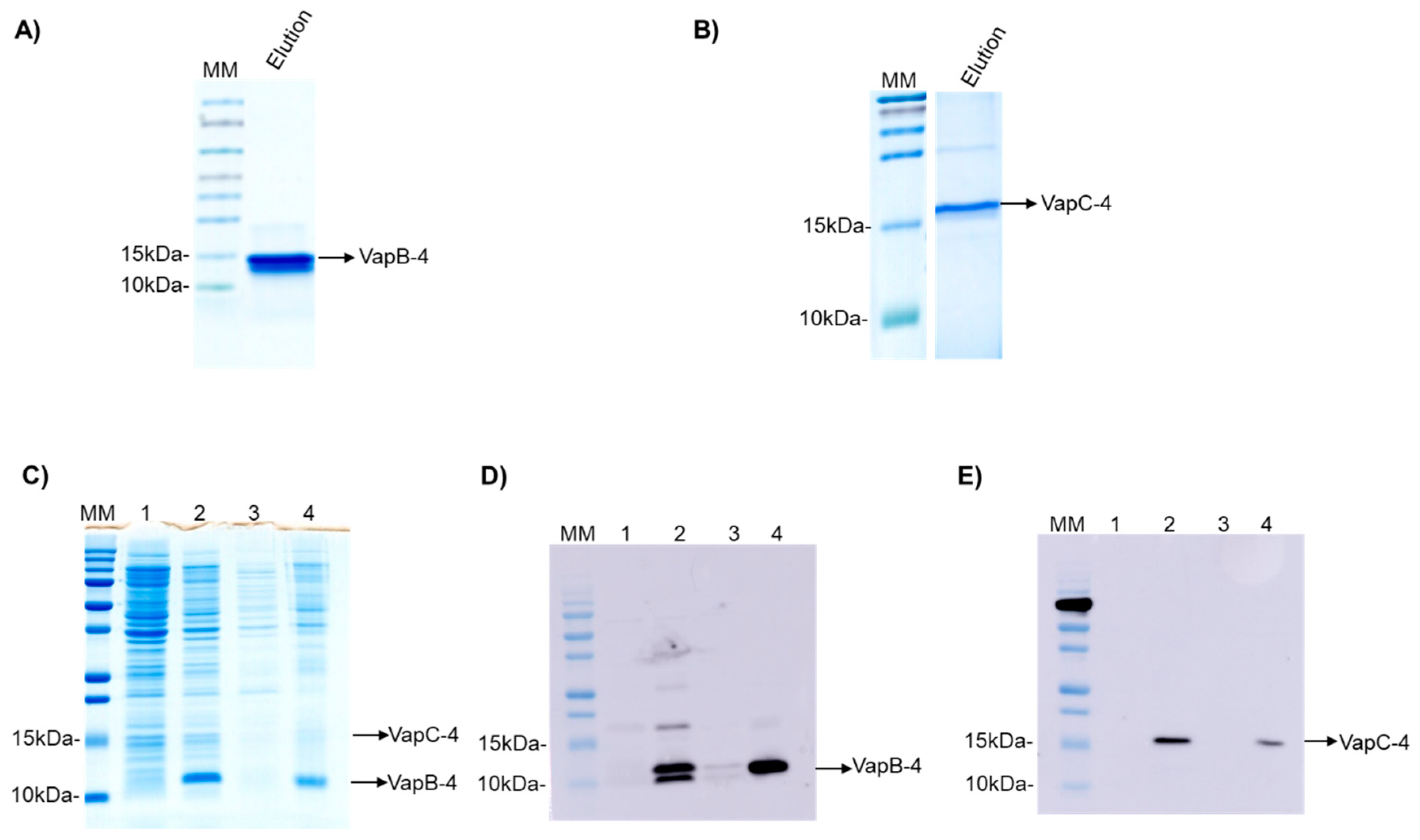
3.2. Production of Recombinant VapB-4 and VapC-4 Proteins Independently and in Tandem
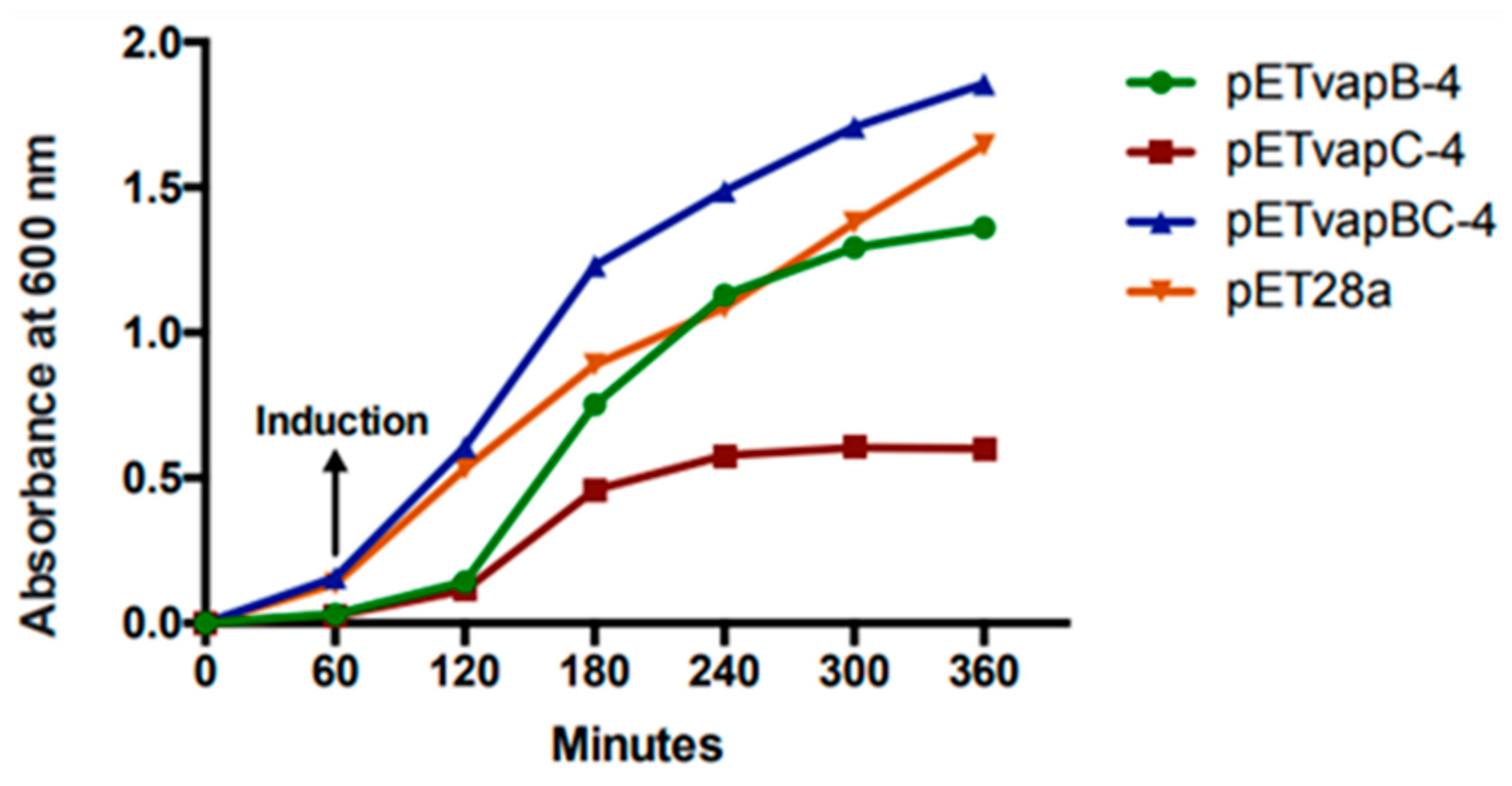
3.3. VapC-4 Toxin Inhibits E. coli Growth Rate
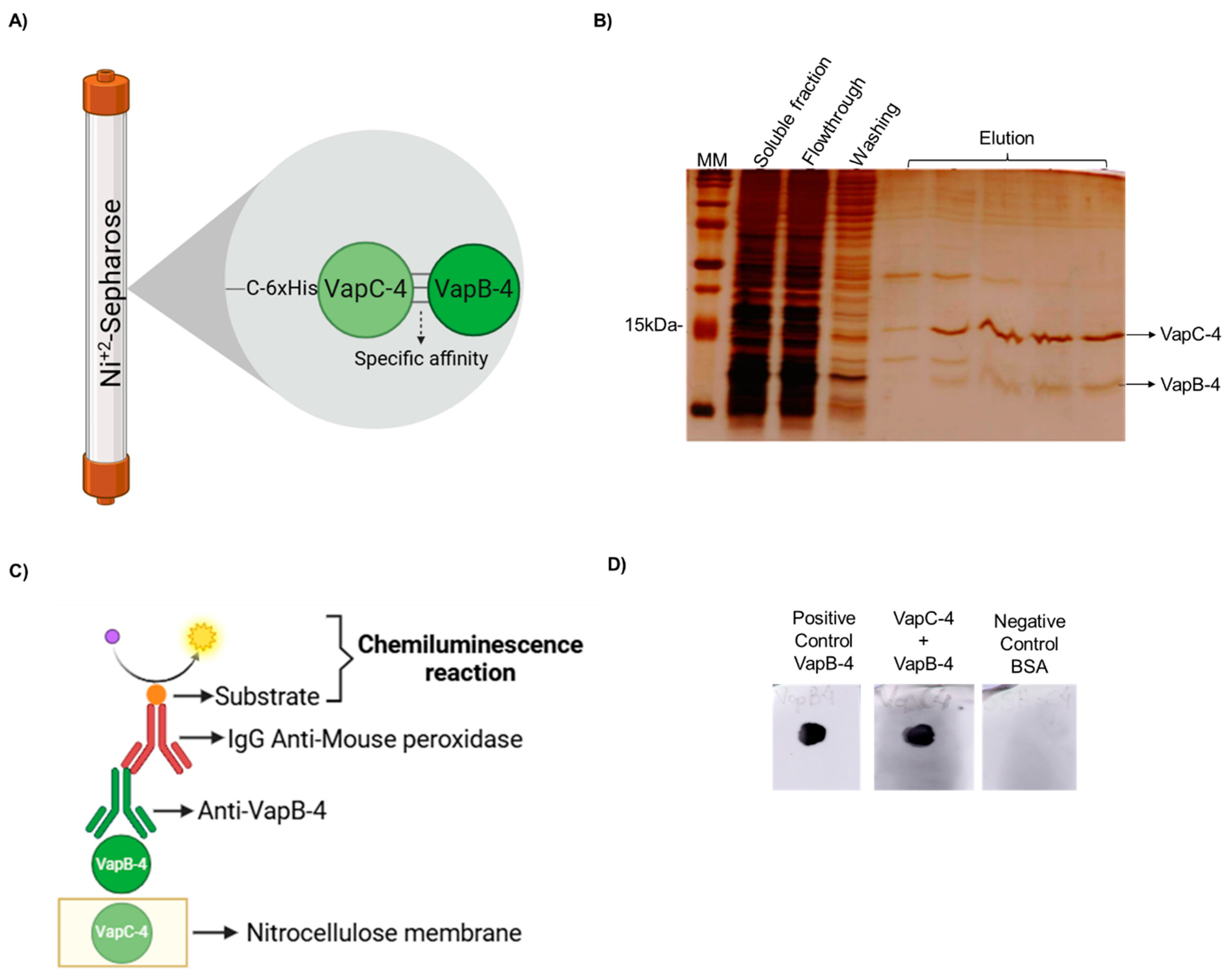
3.4. The Antitoxin VapB-4 Binds to the Toxin VapC-4 In Vivo and In Vitro
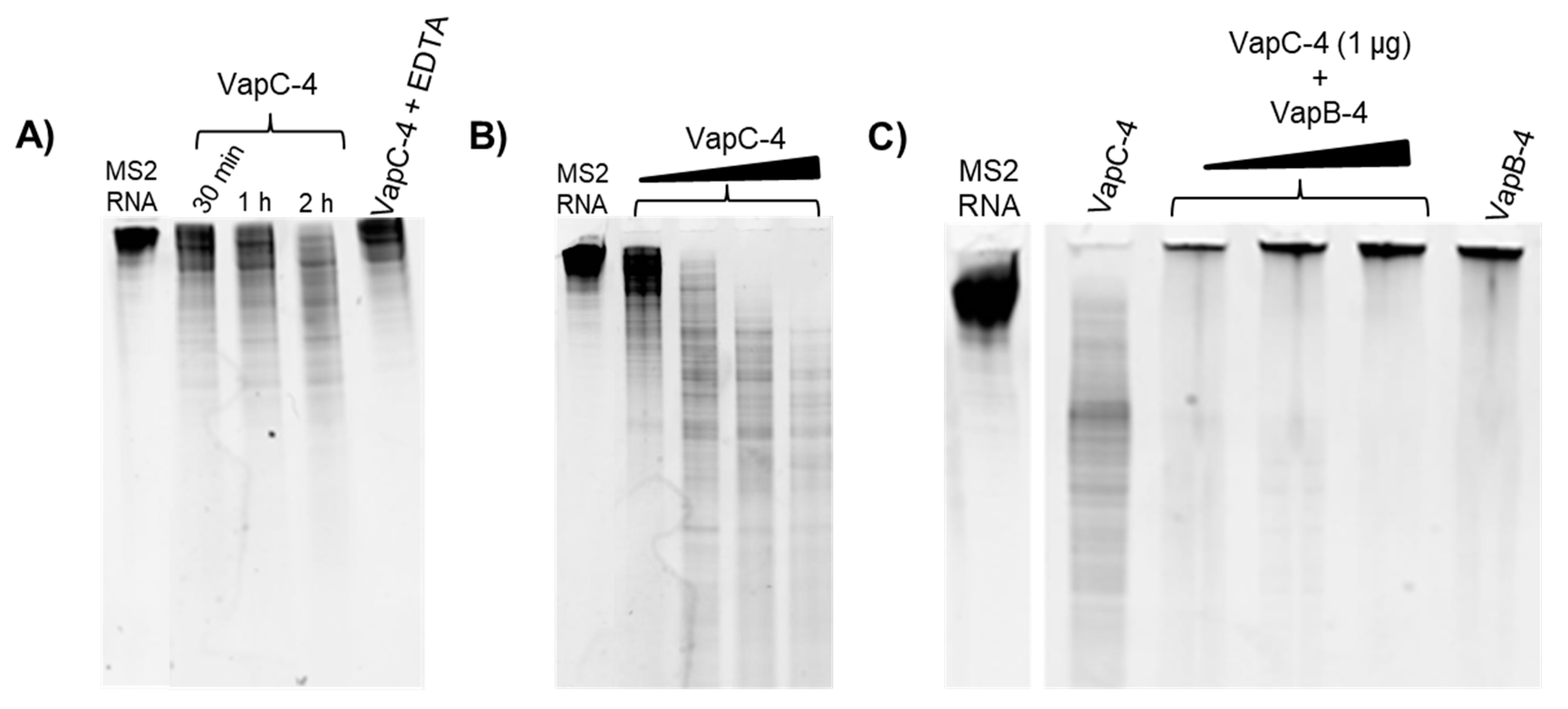
3.5. Determination of VapC-4 Ribonuclease Activity
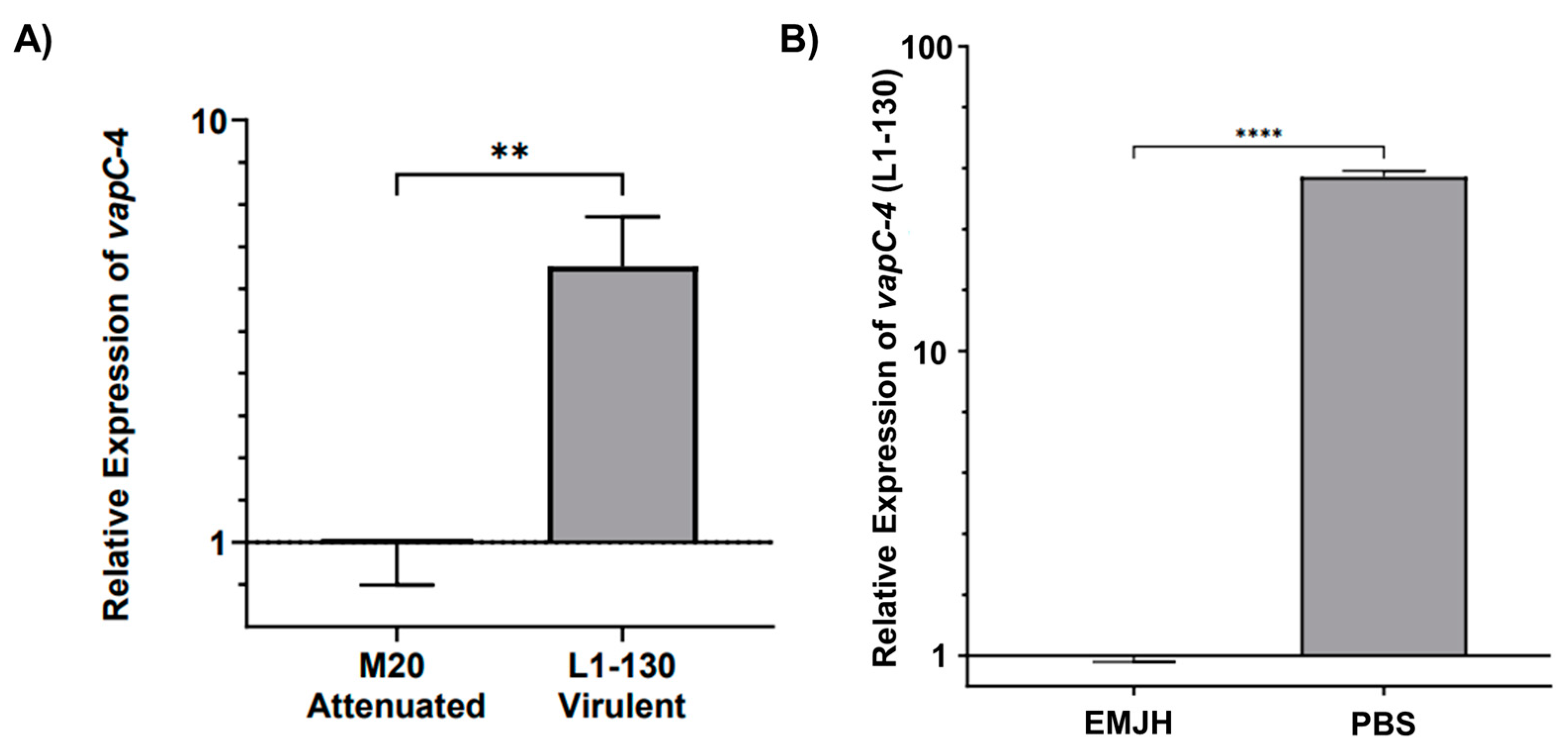
3.6. Analysis of vapC-4 Transcription
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chapartegui-González, I.; Khakhum, N.; Stockton, J.L.; Torres, A.G. Evaluating the Contribution of the Predicted Toxin-Antitoxin System HigBA to Persistence, Biofilm Formation, and Virulence in Burkholderia pseudomallei. Infect. Immun. 2022, 90, e0003522. [Google Scholar] [CrossRef] [PubMed]
- Jurėnas, D.; Fraikin, N.; Goormaghtigh, F.; Van Melderen, L. Biology and evolution of bacterial toxin-antitoxin systems. Nat. Rev. Microbiol. 2022, 20, 335–350. [Google Scholar] [CrossRef]
- Pandey, D.P.; Gerdes, K. Toxin-antitoxin loci are highly abundant in free-living but lost from host-associated prokaryotes. Nucleic Acids Res. 2005, 33, 966–976. [Google Scholar] [CrossRef]
- Page, R.; Peti, W. Toxin-antitoxin systems in bacterial growth arrest and persistence. Nat. Chem. Biol. 2016, 12, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, K.L.; Brodersen, D.E. Higher-Order Structure in Bacterial VapBC Toxin-Antitoxin Complexes. Subcell. Biochem. 2017, 83, 381–412. [Google Scholar] [CrossRef]
- Fozo, E.M.; Makarova, K.S.; Shabalina, S.A.; Yutin, N.; Koonin, E.V.; Storz, G. Abundance of type I toxin-antitoxin systems in bacteria: Searches for new candidates and discovery of novel families. Nucleic Acids Res. 2010, 38, 3743–3759. [Google Scholar] [CrossRef]
- Fozo, E.M.; Hemm, M.R.; Storz, G. Small toxic proteins and the antisense RNAs that repress them. Microbiol. Mol. Biol. Rev. 2008, 72, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Ramisetty, B.C.M.; Santhosh, R.S. Endoribonuclease type II toxin-antitoxin systems: Functional or selfish? Microbiology 2017, 163, 931–939. [Google Scholar] [CrossRef]
- Fineran, P.C.; Blower, T.R.; Foulds, I.J.; Humphreys, D.P.; Lilley, K.S.; Salmond, G.P. The phage abortive infection system, ToxIN, functions as a protein-RNA toxin-antitoxin pair. Proc. Natl. Acad. Sci. USA 2009, 106, 894–899. [Google Scholar] [CrossRef]
- Masuda, H.; Tan, Q.; Awano, N.; Yamaguchi, Y.; Inouye, M. A novel membrane-bound toxin for cell division, CptA (YgfX), inhibits polymerization of cytoskeleton proteins, FtsZ and MreB, in Escherichia coli. FEMS Microbiol. Lett. 2012, 328, 174–181. [Google Scholar] [CrossRef]
- Wang, X.; Lord, D.M.; Cheng, H.Y.; Osbourne, D.O.; Hong, S.H.; Sanchez-Torres, V.; Quiroga, C.; Zheng, K.; Herrmann, T.; Peti, W.; et al. A new type V toxin-antitoxin system where mRNA for toxin GhoT is cleaved by antitoxin GhoS. Nat. Chem. Biol. 2012, 8, 855–861. [Google Scholar] [CrossRef]
- Aakre, C.; Phung, T.; Huang, D.; Laub, M. A Bacterial Toxin Inhibits DNA Replication Elongation Through a Direct Interaction with the β Sliding Clamp. Mol. Cell. 2013, 52, 617–628. [Google Scholar] [CrossRef]
- Marimon, O.; Teixeira, J.M.; Cordeiro, T.N.; Soo, V.W.; Wood, T.L.; Mayzel, M.; Amata, I.; García, J.; Morera, A.; Gay, M.; et al. An oxygen-sensitive toxin-antitoxin system. Nat. Commun. 2016, 7, 13634. [Google Scholar] [CrossRef]
- Choi, J.S.; Kim, W.; Suk, S.; Park, H.; Bak, G.; Yoon, J.; Lee, Y. The small RNA, SdsR, acts as a novel type of toxin in Escherichia coli. RNA Biol. 2018, 15, 1319–1335. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, K.; Christensen, S.K.; Løbner-Olesen, A. Prokaryotic toxin-antitoxin stress response loci. Nat. Rev. Microbiol. 2005, 3, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.E.; Strugnell, R.A.; Rood, J.I. Molecular characterization of a genomic region associated with virulence in Dichelobacter nodosus. Infect. Immun. 1992, 60, 4586–4592. [Google Scholar] [CrossRef] [PubMed]
- Sevin, E.W.; Barloy-Hubler, F. RASTA-Bacteria: A web-based tool for identifying toxin-antitoxin loci in prokaryotes. Genome Biol. 2007, 8, R155. [Google Scholar] [CrossRef]
- Xie, Y.; Wei, Y.; Shen, Y.; Li, X.; Zhou, H.; Tai, C.; Deng, Z.; Ou, H.Y. TADB 2.0: An updated database of bacterial type II toxin-antitoxin loci. Nucleic Acids Res. 2018, 46, D749–D753. [Google Scholar] [CrossRef]
- Chan, W.T.; Espinosa, M.; Yeo, C.C. Keeping the Wolves at Bay: Antitoxins of Prokaryotic Type II Toxin-Antitoxin Systems. Front. Mol. Biosci. 2016, 3, 9. [Google Scholar] [CrossRef]
- Gerdes, K.; Maisonneuve, E. Bacterial persistence and toxin-antitoxin loci. Annu. Rev. Microbiol. 2012, 66, 103–123. [Google Scholar] [CrossRef]
- Muthuramalingam, M.; White, J.C.; Bourne, C.R. Toxin-Antitoxin Modules Are Pliable Switches Activated by Multiple Protease Pathways. Toxins 2016, 8, 214. [Google Scholar] [CrossRef] [PubMed]
- Damiano, D.K.; Azevedo, B.O.P.; Fernandes, G.S.C.; Teixeira, A.F.; Gonçalves, V.M.; Nascimento, A.L.T.O.; Lopes, A.P.Y. The Toxin of VapBC-1 Toxin-Antitoxin Module from. Microorganisms 2024, 12, 1660. [Google Scholar] [CrossRef]
- Wen, Y.; Behiels, E.; Devreese, B. Toxin-Antitoxin systems: Their role in persistence, biofilm formation, and pathogenicity. Pathog. Dis. 2014, 70, 240–249. [Google Scholar] [CrossRef]
- Ogura, T.; Hiraga, S. Mini-F plasmid genes that couple host cell division to plasmid proliferation. Proc. Natl. Acad. Sci. USA 1983, 80, 4784–4788. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Bedzyk, L.A.; Thomas, S.M.; Ye, R.W.; Wood, T.K. Gene expression in Escherichia coli biofilms. Appl. Microbiol. Biotechnol. 2004, 64, 515–524. [Google Scholar] [CrossRef]
- Barth, V.C.; Chauhan, U.; Zeng, J.; Su, X.; Zheng, H.; Husson, R.N.; Woychik, N.A. VapC4 toxin engages small ORFs to initiate an integrated oxidative and copper stress response. Proc. Natl. Acad. Sci. USA 2021, 118, e2022136118. [Google Scholar] [CrossRef]
- Kim, Y.; Wang, X.; Ma, Q.; Zhang, X.S.; Wood, T.K. Toxin-antitoxin systems in Escherichia coli influence biofilm formation through YjgK (TabA) and fimbriae. J. Bacteriol. 2009, 191, 1258–1267. [Google Scholar] [CrossRef]
- Moreno-Del Álamo, M.; Marchisone, C.; Alonso, J.C. Antitoxin ε Reverses Toxin ζ-Facilitated Ampicillin Dormants. Toxins 2020, 12, 801. [Google Scholar] [CrossRef] [PubMed]
- Georgiades, K.; Raoult, D. Genomes of the most dangerous epidemic bacteria have a virulence repertoire characterized by fewer genes but more toxin-antitoxin modules. PLoS ONE 2011, 6, e17962. [Google Scholar] [CrossRef]
- Song, Y.; Tang, H.; Bao, R. Comparative analysis of five type II TA systems identified in. Front. Cell Infect. Microbiol. 2023, 13, 1127786. [Google Scholar] [CrossRef]
- Song, S.; Wood, T.K. A Primary Physiological Role of Toxin/Antitoxin Systems is Phage Inhibition. Front. Microbiol. 2020, 11, 1895. [Google Scholar] [CrossRef] [PubMed]
- Hayes, F.; Kędzierska, B. Regulating toxin-antitoxin expression: Controlled detonation of intracellular molecular timebombs. Toxins 2014, 6, 337–358. [Google Scholar] [CrossRef]
- Bloom-Ackermann, Z.; Steinberg, N.; Rosenberg, G.; Oppenheimer-Shaanan, Y.; Pollack, D.; Ely, S.; Storzi, N.; Levy, A.; Kolodkin-Gal, I. Toxin-Antitoxin systems eliminate defective cells and preserve symmetry in Bacillus subtilis biofilms. Environ. Microbiol. 2016, 18, 5032–5047. [Google Scholar] [CrossRef] [PubMed]
- Zadeh, R.G.; Kalani, B.S.; Ari, M.M.; Talebi, M.; Razavi, S.; Jazi, F.M. Isolation of persister cells within the biofilm and relative gene expression analysis of type II toxin/antitoxin system in Pseudomonas aeruginosa isolates in exponential and stationary phases. J. Glob. Antimicrob. Resist. 2022, 28, 30–37. [Google Scholar] [CrossRef]
- Coşkun, U.S.Ş.; Dagcioglu, Y. Evaluation of toxin-antitoxin genes, antibiotic resistance, and virulence genes in Pseudomonas aeruginosa isolates. Rev. Assoc. Med. Bras. (1992) 2023, 69, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.P.Y.; Azevedo, B.O.P.; Emídio, R.C.; Damiano, D.K.; Nascimento, A.L.T.O.; Barazzone, G.C. Analysis of Genetic VapC Profiles from the Toxin-Antitoxin Type II VapBC Modules among Pathogenic, Intermediate, and Non-Pathogenic. Microorganisms 2019, 7, 56. [Google Scholar] [CrossRef]
- Ren, S.X.; Fu, G.; Jiang, X.G.; Zeng, R.; Miao, Y.G.; Xu, H.; Zhang, Y.X.; Xiong, H.; Lu, G.; Lu, L.F.; et al. Unique physiological and pathogenic features of Leptospira interrogans revealed by whole-genome sequencing. Nature 2003, 422, 888–893. [Google Scholar] [CrossRef]
- Trindade, C.N.R.; Panzenhagen, P.H.N.; Junqueira, R.M.; Silva, D.C.V.; Conte-Junior, C.A.; Balassiano, I.T. Draft Genome Sequences of Leptospira interrogans Serovar Copenhageni Strains Isolated from Patients with Weil’s Disease in Brazil. Microbiol. Resour. Announc. 2020, 9, 10-1128. [Google Scholar] [CrossRef]
- Costa, F.; Hagan, J.E.; Calcagno, J.; Kane, M.; Torgerson, P.; Martinez-Silveira, M.S.; Stein, C.; Abela-Ridder, B.; Ko, A.I. Global Morbidity and Mortality of Leptospirosis: A Systematic Review. PLoS Negl. Trop. Dis. 2015, 9, e0003898. [Google Scholar] [CrossRef]
- Durski, K.N.; Jancloes, M.; Chowdhary, T.; Bertherat, E. A global, multi-disciplinary, multi-sectorial initiative to combat leptospirosis: Global Leptospirosis Environmental Action Network (GLEAN). Int. J. Environ. Res. Public Health 2014, 11, 6000–6008. [Google Scholar] [CrossRef]
- Haake, D.A.; Levett, P.N. Leptospirosis in humans. Curr. Top. Microbiol. Immunol. 2015, 387, 65–97. [Google Scholar] [CrossRef]
- Lehmann, J.S.; Matthias, M.A.; Vinetz, J.M.; Fouts, D.E. Leptospiral pathogenomics. Pathogens 2014, 3, 280–308. [Google Scholar] [CrossRef] [PubMed]
- Arcus, V.L.; McKenzie, J.L.; Robson, J.; Cook, G.M. The PIN-domain ribonucleases and the prokaryotic VapBC toxin-antitoxin array. Protein Eng. Des. Sel. 2011, 24, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Senissar, M.; Manav, M.C.; Brodersen, D.E. Structural conservation of the PIN domain active site across all domains of life. Protein Sci. 2017, 26, 1474–1492. [Google Scholar] [CrossRef]
- Chauhan, U.; Barth, V.C.; Woychik, N.A. tRNA. Antimicrob. Agents Chemother. 2022, 66, e0189621. [Google Scholar] [CrossRef]
- Takahashi, M.B.; Teixeira, A.F.; Nascimento, A.L.T.O. Host Cell Binding Mediated by. Int. J. Mol. Sci. 2022, 23, 15550. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, A.L.; Verjovski-Almeida, S.; Van Sluys, M.A.; Monteiro-Vitorello, C.B.; Camargo, L.E.; Digiampietri, L.A.; Harstkeerl, R.A.; Ho, P.L.; Marques, M.V.; Oliveira, M.C.; et al. Genome features of Leptospira interrogans serovar Copenhageni. Braz. J. Med. Biol. Res. 2004, 37, 459–477. [Google Scholar] [CrossRef]
- Sonnhammer, E.L.; Eddy, S.R.; Durbin, R. Pfam: A comprehensive database of protein domain families based on seed alignments. Proteins 1997, 28, 405–420. [Google Scholar] [CrossRef]
- Dienemann, C.; Bøggild, A.; Winther, K.S.; Gerdes, K.; Brodersen, D.E. Crystal structure of the VapBC toxin-antitoxin complex from Shigella flexneri reveals a hetero-octameric DNA-binding assembly. J. Mol. Biol. 2011, 414, 713–722. [Google Scholar] [CrossRef]
- Oppenheim, D.S.; Yanofsky, C. Translational coupling during expression of the tryptophan operon of Escherichia coli. Genetics 1980, 95, 785–795. [Google Scholar] [CrossRef]
- Cataudella, I.; Trusina, A.; Sneppen, K.; Gerdes, K.; Mitarai, N. Conditional cooperativity in toxin-antitoxin regulation prevents random toxin activation and promotes fast translational recovery. Nucleic Acids Res. 2012, 40, 6424–6434. [Google Scholar] [CrossRef]
- Cooper, C.R.; Daugherty, A.J.; Tachdjian, S.; Blum, P.H.; Kelly, R.M. Role of vapBC toxin-antitoxin loci in the thermal stress response of Sulfolobus solfataricus. Biochem. Soc. Trans. 2009, 37, 123–126. [Google Scholar] [CrossRef]
- Nascimento, A.L.; Ko, A.I.; Martins, E.A.; Monteiro-Vitorello, C.B.; Ho, P.L.; Haake, D.A.; Verjovski-Almeida, S.; Hartskeerl, R.A.; Marques, M.V.; Oliveira, M.C.; et al. Comparative genomics of two Leptospira interrogans serovars reveals novel insights into physiology and pathogenesis. J. Bacteriol. 2004, 186, 2164–2172. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.P.; Lopes, L.M.; Fraga, T.R.; Chura-Chambi, R.M.; Sanson, A.L.; Cheng, E.; Nakajima, E.; Morganti, L.; Martins, E.A. VapC from the leptospiral VapBC toxin-antitoxin module displays ribonuclease activity on the initiator tRNA. PLoS ONE 2014, 9, e101678. [Google Scholar] [CrossRef]
- Ricaldi, J.N.; Fouts, D.E.; Selengut, J.D.; Harkins, D.M.; Patra, K.P.; Moreno, A.; Lehmann, J.S.; Purushe, J.; Sanka, R.; Torres, M.; et al. Whole genome analysis of Leptospira licerasiae provides insight into leptospiral evolution and pathogenicity. PLoS Negl. Trop. Dis. 2012, 6, e1853. [Google Scholar] [CrossRef] [PubMed]
- Ramisetty, B.C.; Santhosh, R.S. Horizontal gene transfer of chromosomal Type II toxin-antitoxin systems of Escherichia coli. FEMS Microbiol. Lett. 2016, 363, fnv238. [Google Scholar] [CrossRef]
- Engelberg-Kulka, H.; Glaser, G. Addiction modules and programmed cell death and antideath in bacterial cultures. Annu. Rev. Microbiol. 1999, 53, 43–70. [Google Scholar] [CrossRef] [PubMed]
- Tchamedeu Kameni, A.P.; Couture-Tosi, E.; Saint-Girons, I.; Picardeau, M. Inactivation of the spirochete recA gene results in a mutant with low viability and irregular nucleoid morphology. J. Bacteriol. 2002, 184, 452–458. [Google Scholar] [CrossRef]
- Bhowmick, A.; Recalde, A.; Bhattacharyya, C.; Banerjee, A.; Das, J.; Rodriguez-Cruz, U.E.; Albers, S.V.; Ghosh, A. Role of VapBC4 toxin-antitoxin system of. mBio 2024, 15, e0275324. [Google Scholar] [CrossRef]
- Khoo, S.K.; Loll, B.; Chan, W.T.; Shoeman, R.L.; Ngoo, L.; Yeo, C.C.; Meinhart, A. Molecular and structural characterization of the PezAT chromosomal toxin-antitoxin system of the human pathogen Streptococcus pneumoniae. J. Biol. Chem. 2007, 282, 19606–19618. [Google Scholar] [CrossRef]
- Ning, D.; Jiang, Y.; Liu, Z.; Xu, Q. Characterization of a chromosomal type II toxin-antitoxin system mazEaFa in the Cyanobacterium Anabaena sp. PCC 7120. PLoS ONE 2013, 8, e56035. [Google Scholar] [CrossRef] [PubMed]
- Ning, D.; Liu, S.; Xu, W.; Zhuang, Q.; Wen, C.; Tang, X. Transcriptional and proteolytic regulation of the toxin-antitoxin locus vapBC10 (ssr2962/slr1767) on the chromosome of Synechocystis sp. PCC 6803. PLoS ONE 2013, 8, e80716. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Guo, Y.; Zeng, Z.; Liu, X.; Shi, F.; Wang, X. Identification and characterization of a HEPN-MNT family type II toxin-antitoxin in Shewanella oneidensis. Microb. Biotechnol. 2015, 8, 961–973. [Google Scholar] [CrossRef]
- Zheng, C.; Zhao, X.; Zeng, T.; Cao, M.; Xu, J.; Shi, G.; Li, J.; Chen, H.; Bei, W. Identification of four type II toxin-antitoxin systems in Actinobacillus pleuropneumoniae. FEMS Microbiol. Lett. 2017, 364, fnx126. [Google Scholar] [CrossRef] [PubMed]
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, Targets, and Triggers: An Overview of Toxin-Antitoxin Biology. Mol. Cell 2018, 70, 768–784. [Google Scholar] [CrossRef]
- Winther, K.S.; Gerdes, K. Enteric virulence associated protein VapC inhibits translation by cleavage of initiator tRNA. Proc. Natl. Acad. Sci. USA 2011, 108, 7403–7407. [Google Scholar] [CrossRef]
- Starosta, A.L.; Lassak, J.; Jung, K.; Wilson, D.N. The bacterial translation stress response. FEMS Microbiol. Rev. 2014, 38, 1172–1201. [Google Scholar] [CrossRef]
- Christensen, S.K.; Mikkelsen, M.; Pedersen, K.; Gerdes, K. RelE, a global inhibitor of translation, is activated during nutritional stress. Proc. Natl. Acad. Sci. USA 2001, 98, 14328–14333. [Google Scholar] [CrossRef]
- Silva, J.C.A.; Marques-Neto, L.M.; Carvalho, E.; Del Carpio, A.M.G.; Henrique, C.; Leite, L.C.C.; Mitsunari, T.; Elias, W.P.; Munhoz, D.D.; Piazza, R.M.F. Chromosomal Type II Toxin-Antitoxin Systems May Enhance Bacterial Fitness of a Hybrid Pathogenic. Toxins 2024, 16, 469. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′ → 3′) | Restriction Site | Construction |
|---|---|---|---|
| F-vapB4-Nhis 1 | gagccatgggcCACCACCACCACCACCACATGAAATCGTATCCGGTTGG | Nco I | pET-vapB-4 |
| R-vapB4-Nhis 1 | gagcctcgagTGTTAACTTAAAAACTCTTCTTCGG | Xho I | |
| F-vapC4-Nhis 1 | gagccatgggcCACCACCACCACCACCACATGAAGTATTTGCTTGATAC | Nco I | pET-vapC-4 |
| R-vapC4-Nhis 1 | gagcctcgagttaTCACCAAAGTGTTTTTAATCCATG | Xho I | |
| F-vapBC4-Chis 1 | gagccatgggcATGAAATCGTATCCGGTTGG | Nco I | pET-vapBC-4 |
| R-vapBC4-Chis 1 | gagcctcgagCCAAAGTGTTTTTAATCCATGTGATC | Xho I | |
| F-qPCR-vapB-4 2 | CCGGTTGGCGAGCTTAAATC | - | - |
| R-qPCR-vapB-4 2 | TTACCTTTCCATCCAATAATCCT | - | - |
| F-qPCR-vapC-4 2 | ACGTTAGCTCAGTTTCTCTTTGG | - | - |
| R-qPCR-vapC-4 2 | ATCGTATGTGCTTGCATCGG | - | - |
| F-qPCR-vapBC-4 2 | GCCGATTGCCATGATTATTCCT | - | - |
| R-qPCR-vapBC-4 2 | AGATGAACCGATTATCCAAAGAA | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azevedo, B.O.P.; Damiano, D.K.; Teixeira, A.F.; Nascimento, A.L.T.O.; Fernandes, L.G.V.; Lopes, A.P.Y. The VapBC-4 Characterization Indicates It Is a Bona Fide Toxin-Antitoxin Module of Leptospira interrogans: Initial Evidence for a Role in Bacterial Adaptation. Microorganisms 2025, 13, 879. https://doi.org/10.3390/microorganisms13040879
Azevedo BOP, Damiano DK, Teixeira AF, Nascimento ALTO, Fernandes LGV, Lopes APY. The VapBC-4 Characterization Indicates It Is a Bona Fide Toxin-Antitoxin Module of Leptospira interrogans: Initial Evidence for a Role in Bacterial Adaptation. Microorganisms. 2025; 13(4):879. https://doi.org/10.3390/microorganisms13040879
Chicago/Turabian StyleAzevedo, Bruna Oliveira Pigatto, Deborah Kohn Damiano, Aline Florencio Teixeira, Ana Lucia Tabet Oller Nascimento, Luis Guilherme Virgilio Fernandes, and Alexandre Paulo Yague Lopes. 2025. "The VapBC-4 Characterization Indicates It Is a Bona Fide Toxin-Antitoxin Module of Leptospira interrogans: Initial Evidence for a Role in Bacterial Adaptation" Microorganisms 13, no. 4: 879. https://doi.org/10.3390/microorganisms13040879
APA StyleAzevedo, B. O. P., Damiano, D. K., Teixeira, A. F., Nascimento, A. L. T. O., Fernandes, L. G. V., & Lopes, A. P. Y. (2025). The VapBC-4 Characterization Indicates It Is a Bona Fide Toxin-Antitoxin Module of Leptospira interrogans: Initial Evidence for a Role in Bacterial Adaptation. Microorganisms, 13(4), 879. https://doi.org/10.3390/microorganisms13040879