
Multilocus Variable-Number Tandem-Repeat Analysis as an Investigation Tool in Cryptosporidium parvum Outbreaks in Finland and Sweden in 2022

 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Epidemiological Investigation
2.2. Statistical Analyses
2.3. Environmental and Traceback Investigation
2.4. Microbiological Investigation
2.5. Swedish Outbreak Investigation
3. Results
3.1. Clinical Microbiology Findings, Cryptosporidium Species Identification, and gp60 Subtyping
3.2. MLVA Typing of Patient Samples
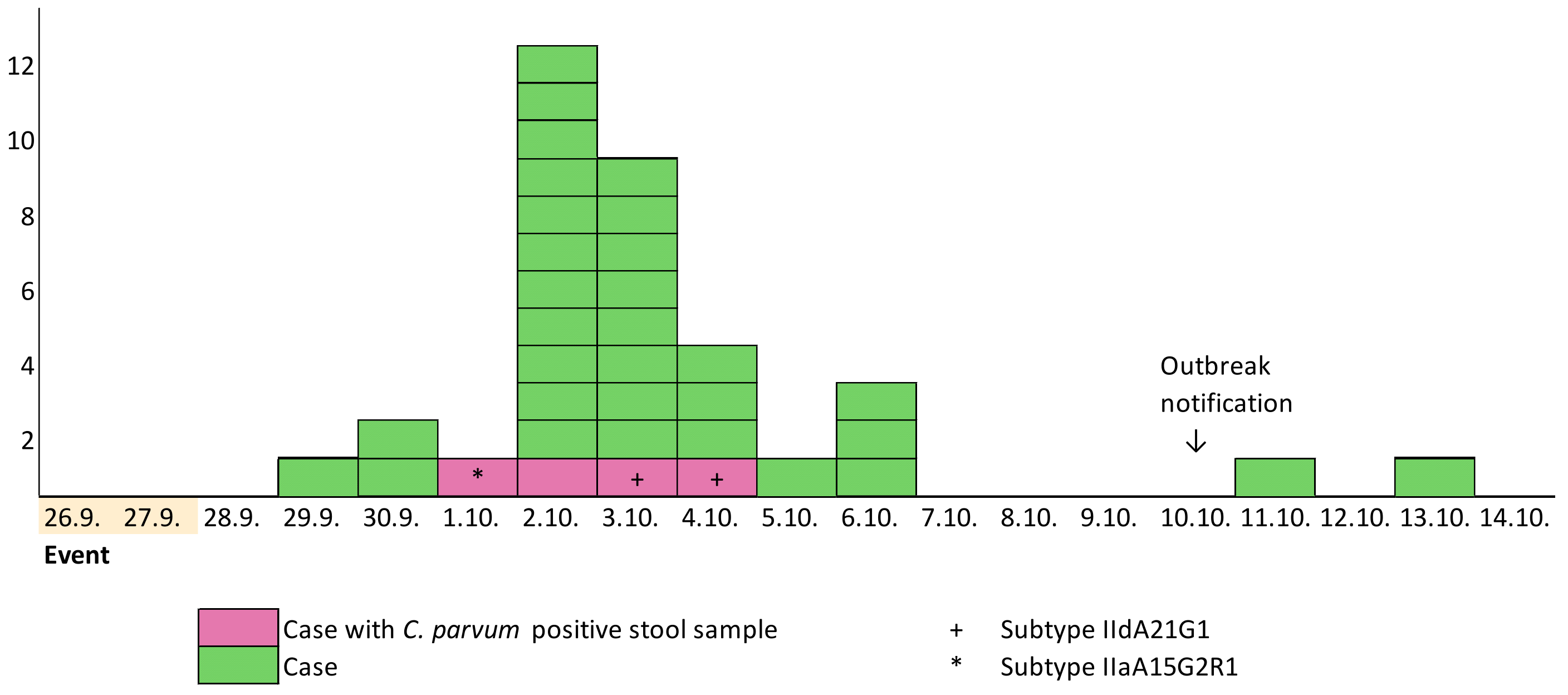
3.3. Descriptive Epidemiology
3.4. Analytical Epidemiology
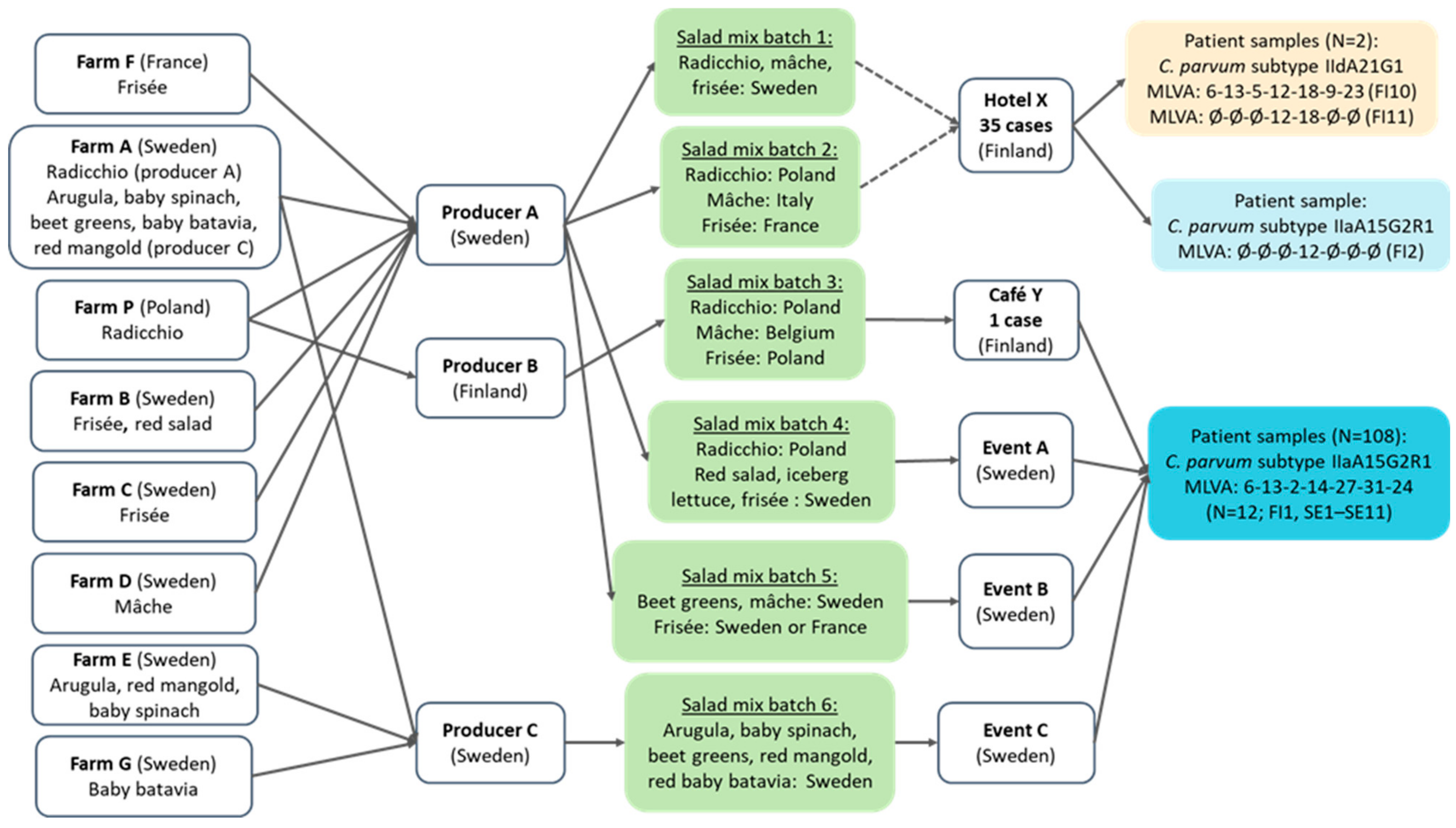
3.5. Environmental and Traceback Investigation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Checkley, W.; White, A.C., Jr.; Jaganath, D.; Arrowood, M.J.; Chalmers, R.M.; Chen, X.M.; Fayer, R.; Griffiths, J.K.; Guerrant, R.L.; Hedstrom, L.; et al. A review of the global burden, novel diagnostics, therapeutics, and vaccine targets for cryptosporidium. Lancet Infect. Dis. 2015, 15, 85–94. [Google Scholar] [CrossRef]
- Feng, Y.; Ryan, U.M.; Xiao, L. Genetic diversity and population structure of Cryptosporidium. Trends Parasitol. 2018, 34, 997–1011. [Google Scholar] [CrossRef]
- Cacciò, S.M.; Chalmers, R.M. Human cryptosporidiosis in Europe. Clin. Microbiol. Infect. 2016, 22, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, A.K.; Kumar, S.; Smith, W.A.; Sahu, P.S. Revisiting the global problem of cryptosporidiosis and recommendations. Trop. Parasitol. 2017, 7, 8–17. [Google Scholar] [PubMed]
- Hunter, P.R.; Hughes, S.; Woodhouse, S.; Raj, N.; Syed, Q.; Chalmers, R.M.; Verlander, N.Q.; Goodacre, J. Health sequelae of human cryptosporidiosis in immunocompetent patients. Clin. Infect. Dis. 2004, 39, 504–510. [Google Scholar] [CrossRef]
- Kosek, M.; Alcantara, C.; Lima, A.A.; Guerrant, R.L. Cryptosporidiosis: An update. Lancet Infect. Dis. 2001, 1, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Hijjawi, N.; Xiao, L. Foodborne cryptosporidiosis. Int. J. Parasitol. 2018, 48, 1–12. [Google Scholar] [CrossRef]
- Bujila, I.; Troell, K.; Ögren, J.; Hansen, A.; Killander, G.; Agudelo, L.; Lebbad, M.; Beser, J. Cryptosporidium species and subtypes identified in human domestic cases through the national microbiological surveillance programme in Sweden from 2018 to 2022. BMC Infect. Dis. 2024, 24, 146. [Google Scholar] [CrossRef]
- Åberg, R.; Sjöman, M.; Hemminki, K.; Pirnes, A.; Räsänen, S.; Kalanti, A.; Pohjanvirta, T.; Caccio, S.M.; Pihlajasaari, A.; Toikkanen, S.; et al. Cryptosporidium parvum caused a large outbreak linked to frisée salad in Finland, 2012. Zoonoses Public Health 2015, 62, 618–624. [Google Scholar] [CrossRef]
- Ryan, U.M.; Feng, Y.; Fayer, R.; Xiao, L. Taxonomy and molecular epidemiology of Cryptosporidium and Giardia—A 50 year perspective (1971–2021). Int. J. Parasitol. 2021, 51, 1099–1119. [Google Scholar] [CrossRef]
- Strong, W.B.; Gut, J.; Nelson, R.G. Cloning and sequence analysis of a highly polymorphic Cryptosporidium parvum gene encoding a 60-kilodalton glycoprotein and characterization of its 15- and 45-kilodalton zoite surface antigen products. Infect. Immun. 2000, 68, 4117–4134. [Google Scholar] [CrossRef]
- Morris, A.; Robinson, G.; Swain, M.T.; Chalmers, R.M. Direct sequencing of Cryptosporidium in stool samples for public health. Front. Public. Health 2019, 7, 360. [Google Scholar] [CrossRef] [PubMed]
- Chalmers, R.M.; Robinson, G.; Elwin, K.; Elson, R. Analysis of the Cryptosporidium spp. and gp60 subtypes linked to human outbreaks of cryptosporidiosis in England and Wales, 2009 to 2017. Parasit. Vectors 2019, 12, 95. [Google Scholar] [CrossRef]
- Robinson, G.; Pérez-Cordón, G.; Hamilton, C.; Katzer, F.; Connelly, L.; Alexander, C.L.; Chalmers, R.M. Validation of a multilocus genotyping scheme for subtyping Cryptosporidium parvum for epidemiological purposes. Food Waterborne Parasitol. 2022, 27, e00151. [Google Scholar] [CrossRef] [PubMed]
- Häkkänen, T.; Rimhanen-Finne, R.; Antikainen, J.; Ruotsalainen, E.; Vainio, A. Molecular characteristics of Cryptosporidium spp. in human cases in five Finnish hospital districts during 2021: First findings of Cryptosporidium mortiferum (Cryptosporidium chipmunk genotype I) in Finland. Int. J. Parasitol. 2024, 54, 225–231. [Google Scholar] [CrossRef]
- Cryptosporidium Reference Unit, Public Health Wales. Assay Transfer Document: Cp MLVA V8 February 2024. C. parvum Multiple Locus Variable-Number Tandem Repeat Analysis. Available online: https://phw.nhs.wales/services-and-teams/cryptosporidium-reference-unit/mlva-assay-protocol/ (accessed on 19 February 2024).
- Garcia-R, J.C.; Hayman, D.T.S. A review and analysis of cryptosporidiosis outbreaks in New Zealand. Parasitology 2023, 150, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Chalmers, R.M.; Cacciò, S. Towards a consensus on genotyping schemes for surveillance and outbreak investigations of Cryptosporidium, Berlin, June 2016. Euro Surveill 2016, 21, 30338. [Google Scholar] [CrossRef]
- Risby, H.; Robinson, G.; Chandra, N.; King, G.; Vivancos, R.; Smith, R.; Thomas, D.; Fox, A.; McCarthy, N.; Chalmers, R.M. Application of a new multi-locus variable number tandem repeat analysis (MLVA) scheme for the seasonal investigation of Cryptosporidium parvum cases in Wales and the northwest of England, spring 2022. Curr. Res. Parasitol. Vector Borne Dis. 2023, 4, 100151. [Google Scholar] [CrossRef]
- Gopfert, A.; Chalmers, R.M.; Whittingham, S.; Wilson, L.; van Hove, M.; Ferraro, C.F.; Robinson, G.; Young, N.; Nozad, B. An outbreak of Cryptosporidium parvum linked to pasteurised milk from a vending machine in England: A descriptive study, March 2021. Epidemiol. Infect. 2022, 150, e185. [Google Scholar] [CrossRef]
- Kubina, S.; Costa, D.; Cazeaux, C.; Villena, I.; Favennec, L.; Razakandrainibe, R.; La Carbona, S. Persistence and survival of Cryptosporidium parvum oocysts on lamb’s lettuce leaves during plant growth and in washing conditions of minimally-processed salads. Int. J. Food Microbiol. 2023, 388, 110085. [Google Scholar] [CrossRef]
- Utaaker, K.S.; Skjerve, E.; Robertson, L.J. Keeping it cool: Survival of Giardia cysts and Cryptosporidium oocysts on lettuce leaves. Int. J. Food Microbiol. 2017, 255, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Suominen, K.A.; Björkstrand, M.; Ollgren, J.; Autio, T.J.; Rimhanen-Finne, R. Cryptosporidiosis in Finland is predominantly of domestic origin: Investigation of increased reporting, 1995–2020. Infect. Dis. 2023, 55, 116–124. [Google Scholar] [CrossRef]
- Enbom, T.; Suominen, K.; Laitinen, S.; Ollgren, J.; Autio, T.; Rimhanen-Finne, R. Cryptosporidium parvum: An emerging occupational zoonosis in Finland. Acta Vet. Scand. 2023, 65, 25. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Huang, J.; Qin, H.; Wang, L.; Li, J.; Zhang, L. Cryptosporidium parvum and gp60 genotype prevalence in dairy calves worldwide: A systematic review and meta-analysis. Acta Trop. 2023, 240, 106843. [Google Scholar] [CrossRef] [PubMed]
- Communicable Diseases Act (21.12.2016/1227). Available online: https://www.finlex.fi/fi/laki/ajantasa/2016/20161227 (accessed on 21 March 2024).

{kind=link}
{kind=link}
| Cryptosporidium Species (N) | Gp60 Subtype (n/N) |
|---|---|
| C. parvum (21) | IIaA15G2R1 (10/21) 1 |
| IIdA21G1 (2/21) 2 | |
| IIaA13G2R1 (2/21) | |
| IIaA15G1R1 (1/21) | |
| IIaA16G1R1 (1/21) | |
| IIaA19G2R1 (1/21) | |
| subtype family IIa (1/21) | |
| NT (3/21) 3 | |
| C. mortiferum (13) | XIVaA20G2T1 (7/13) |
| NT (6/13) 3 | |
| C. hominis (2) | IdA16 (1/2) |
| IeA11G3T3 (1/2) | |
| C. ditrichi (1) | NT 3 |
| Sample | Gp60 Subtype | MLVA Profile | Epidemiological Link to Producer A’s Salad Mix | Origin of Sample |
|---|---|---|---|---|
| FI10 | IIdA21G1 | 6-13-5-12-18-9-23 | Yes | Finland (outbreak) |
| FI11 | IIdA21G1 | Ø-Ø-Ø-12-18-Ø-Ø 1 | Yes | Finland (outbreak) |
| FI2 | IIaA15G2R1 | Ø-Ø-Ø-12-Ø-Ø-Ø 1 | Yes | Finland (outbreak) |
| SE1–SE11 | IIaA15G2R1 | 6-13-2-14-27-31-24 | Yes | Sweden (outbreak) |
| SE12 | IIaA15G2R1 | Ø-13-2-14-27-31-24 1 | Yes | Sweden (outbreak) |
| SE13 | IIaA15G2R1 | Ø-Ø-2-14-27-31-Ø 1 | No | Sweden (outbreak) |
| SE14 | IIaA15G2R1 | 6-Ø-2-14-27-Ø-24 1 | Yes | Sweden (outbreak) |
| FI1 | IIaA15G2R1 | 6-13-2-14-27-31-24 | No | Finland |
| FI7 | IIaA15G2R1 | 6-13-2-14-27-31-24 | NA 2 | Finland |
| FI4–FI6, FI8 | IIaA15G2R1 | 4-14-5-7-27-31-15 | NA 2 | Finland |
| FI9 | IIaA15G2R1 | 4-14-5-7-27-32/35-15 (mixed) | NA 2 | Finland |
| FI3 | subtype family IIa | Ø-Ø-Ø-7-Ø-Ø-Ø 1 | NA 2 | Finland |
| Exposure (Meal) | Food Eaten | Food Not Eaten | Relative Risk | p-Value | % of Cases (N = 35) Exposed | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Total | Cases | AR (%) | Total | Cases | AR (%) | RR | 95% CI | |||
| Chicken artichoke salad (Dinner on 26 September) | 47 | 29 | 62 | 11 | 1 | 9.1 | 6.8 | 1.0–45 | 0.002 | 83 |
| Green salad 1 (Dinner on 26 September) | 56 | 34 | 61 | 8 | 0 | 0 | 6.0 1 | 0.9–39 | 0.005 | 97 |
| Arugula potatoes (Dinner on 26 September) | 43 | 26 | 60 | 12 | 2 | 17 | 3.6 | 1.0–13 | 0.010 | 74 |
| Bratwurst (Breakfast on 27 September) | 6 | 6 | 100 | 52 | 27 | 52 | 1.9 | 1.5–2.5 | 0.032 | 17 |
| Fruit (Breakfast on 26 September) | 9 | 8 | 89 | 55 | 27 | 49 | 1.8 | 1.3–2.6 | 0.033 | 23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suominen, K.; Vainio, A.; Hokkanen, P.; Åberg, R.; Isosomppi, S.; Särelä, E.; Kitowska, W.; Gonzalez-Perez, A.C.; Ollgren, J.; Bujila, I.; et al. Multilocus Variable-Number Tandem-Repeat Analysis as an Investigation Tool in Cryptosporidium parvum Outbreaks in Finland and Sweden in 2022. Microorganisms 2025, 13, 821. https://doi.org/10.3390/microorganisms13040821
Suominen K, Vainio A, Hokkanen P, Åberg R, Isosomppi S, Särelä E, Kitowska W, Gonzalez-Perez AC, Ollgren J, Bujila I, et al. Multilocus Variable-Number Tandem-Repeat Analysis as an Investigation Tool in Cryptosporidium parvum Outbreaks in Finland and Sweden in 2022. Microorganisms. 2025; 13(4):821. https://doi.org/10.3390/microorganisms13040821
Chicago/Turabian StyleSuominen, Kristiina, Anni Vainio, Pirkko Hokkanen, Riikka Åberg, Sanna Isosomppi, Eeva Särelä, Wioleta Kitowska, Ana Cristina Gonzalez-Perez, Jukka Ollgren, Ioana Bujila, and et al. 2025. "Multilocus Variable-Number Tandem-Repeat Analysis as an Investigation Tool in Cryptosporidium parvum Outbreaks in Finland and Sweden in 2022" Microorganisms 13, no. 4: 821. https://doi.org/10.3390/microorganisms13040821
APA StyleSuominen, K., Vainio, A., Hokkanen, P., Åberg, R., Isosomppi, S., Särelä, E., Kitowska, W., Gonzalez-Perez, A. C., Ollgren, J., Bujila, I., Troell, K., Hansen, A., Lindblad, M., & Rimhanen-Finne, R. (2025). Multilocus Variable-Number Tandem-Repeat Analysis as an Investigation Tool in Cryptosporidium parvum Outbreaks in Finland and Sweden in 2022. Microorganisms, 13(4), 821. https://doi.org/10.3390/microorganisms13040821