
High Diversity and Prevalence of Rickettsial Agents in Rhipicephalus microplus Ticks from Livestock in Karst Landscapes of Southwest China

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction
2.3. PCR Assays and Sequencing
2.4. Phylogenetic Analysis
2.5. Statistical Analyses
3. Results
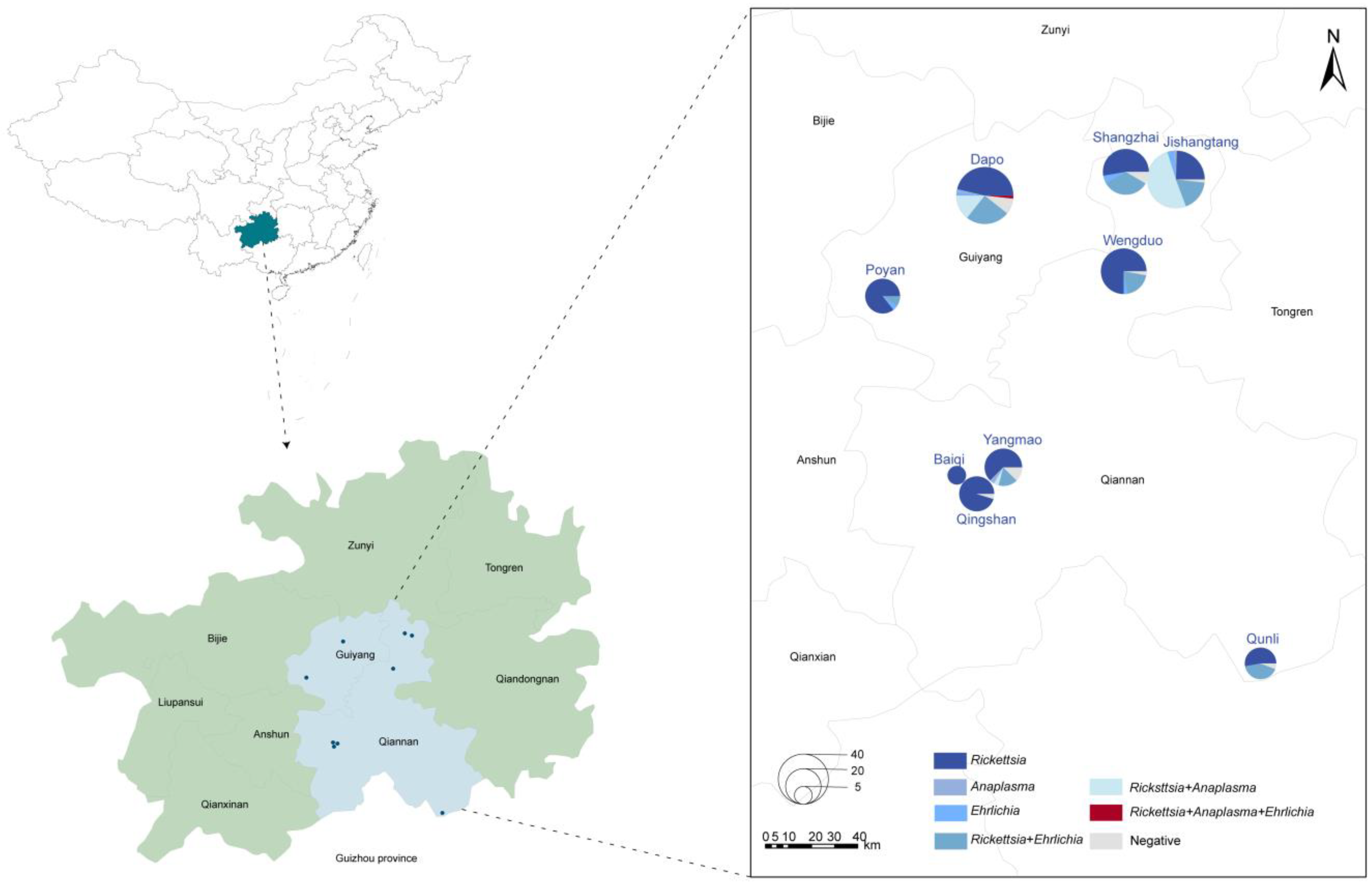
3.1. Tick Sampling
3.2. Phylogenic Analysis of Different Tick-Borne Pathogens
3.3. Prevalence of Different Tick-Borne Pathogens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dantas-Torres, F.; Chomel, B.B.; Otranto, D. Ticks and tick-borne diseases: A one health perspective. Trends Parasitol. 2012, 28, 437–446. [Google Scholar]
- Jia, N.; Wang, J.; Shi, W.; Du, L.; Sun, Y.; Zhan, W.; Jiang, J.F.; Wang, Q.; Zhang, B.; Ji, P.; et al. Large-scale comparative analyses of tick genomes elucidate their genetic diversity and vector capacities. Cell 2020, 182, 1328–1340.e13. [Google Scholar] [PubMed]
- Pace, E.J.; O’Reilly, M. Tickborne diseases: Diagnosis and management. Am. Fam. Physician 2020, 101, 530–540. [Google Scholar] [PubMed]
- Fang, L.Q.; Liu, K.; Li, X.L.; Liang, S.; Yang, Y.; Yao, H.W.; Sun, R.X.; Sun, Y.; Chen, W.J.; Zuo, S.Q.; et al. Emerging tick-borne infections in mainland China: An increasing public health threat. Lancet Infect. Dis. 2015, 15, 1467–1479. [Google Scholar] [PubMed]
- Johnson, N. Ticks: Biology, Ecology, and Diseases, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2023; pp. 25–40, 65–72. [Google Scholar]
- Desta, B. Review on the impact of ticks on livestock health and productivity. J. Biol. Agric. Healthc. 2016, 6, 1–7. [Google Scholar]
- Salje, J. Cells within cells: Rickettsiales and the obligate intracellular bacterial lifestyle. Nat. Rev. Microbiol. 2021, 19, 375–390. [Google Scholar]
- Giannotti, D.; Boscaro, V.; Husnik, F.; Vannini, C.; Keeling, P.J. The “other” Rickettsiales: An overview of the family “Candidatus Midichloriaceae”. Appl. Environ. Microbiol. 2022, 88, e0243221. [Google Scholar]
- Kang, Y.J.; Diao, X.N.; Zhao, G.Y.; Chen, M.H.; Xiong, Y.; Shi, M.; Fu, W.M.; Guo, Y.J.; Pan, B.; Chen, X.P.; et al. Extensive diversity of Rickettsiales bacteria in two species of ticks from China and the evolution of the Rickettsiales. BMC Evol. Biol. 2014, 14, 167. [Google Scholar]
- Li, H.; Zheng, Y.C.; Ma, L.; Jia, N.; Jiang, B.G.; Jiang, R.R.; Huo, Q.B.; Wang, Y.W.; Liu, H.B.; Chu, Y.L.; et al. Human infection with a novel tick-borne Anaplasma species in China: A surveillance study. Lancet Infect. Dis. 2015, 15, 663–670. [Google Scholar]
- Yang, M.; Jia, Y.; Dong, Z.; Zhang, Y.; Xie, S.; Liu, Q.; Wang, Y. Rickettsia aeschlimannii infection in a woman from Xingjiang, northwestern China. Vector-Borne Zoonotic Dis. 2022, 22, 55–57. [Google Scholar]
- Duan, L.; Zhang, L.; Hou, X.; Bao, Z.; Zeng, Y.; He, L.; Liu, Z.; Zhou, H.; Hao, Q.; Dong, A. Surveillance of tick-borne bacterial infection in ticks and forestry populations in Inner Mongolia, China. Front. Public Health 2024, 12, 1302133. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Lian, Y.; Qin, X. Rocky desertification in southwest China: Impacts, causes, and restoration. Earth-Sci. Rev. 2014, 132, 1–12. [Google Scholar] [CrossRef]
- Chen, W.H.; Li, D.; Shu, H.L.; Liang, J.D.; Zhao, J.H.; Tian, W.Y.; Han, Y.F. Four new araneogenous species and a new genus in Hypocreales (Clavicipitaceae, Cordycipitaceae) from the karst region of China. MycoKeys 2025, 112, 335–359. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Meng, C.; Zhang, B.; Wang, X.; Tian, J.; Tang, G.; Wang, W.; Li, N.; Li, M.; Xu, X.; et al. Prevalence of spotted fever group Rickettsia and Candidatus Lariskella in multiple tick species from Guizhou Province, China. Biomolecules 2022, 12, 1701. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, K.; Zhao, S.; Yan, Y.; Wang, H.; Jing, J.; Jian, F.; Wang, R.; Zhang, L.; Ning, C. Detection and phylogenetic characterization of Anaplasma capra: An. emerging pathogen in sheep and goats in China. Front. Cell. Infect. Microbiol. 2018, 8, 283. [Google Scholar] [CrossRef]
- Guo, J.J.; Lin, X.D.; Chen, Y.M.; Hao, Z.Y.; Wang, Z.X.; Yu, Z.M.; Lu, M.; Li, K.; Qin, X.C.; Wang, W.; et al. Diversity and circulation of Jingmen tick virus in ticks and mammals. Virus Evol. 2020, 6, veaa051. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v6: Recent updates to the phylogenetic tree display and annotation tool. Nucleic Acids Res. 2024, 52, W78–W82. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. Mega11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Lagunova, E.K.; Liapunova, N.A.; Tuul, D.; Otgonsuren, G.; Nomin, D.; Erdenebat, N.; Abmed, D.; Danchinova, G.A.; Sato, K.; Kawabata, H.; et al. Co-infections with multiple pathogens in natural populations of Ixodes persulcatus ticks in Mongolia. Parasit. Vectors 2022, 15, 236. [Google Scholar] [CrossRef]
- Guo, W.P.; Wang, Y.H.; Lu, Q.; Xu, G.; Luo, Y.; Ni, X.; Zhou, E.M. Molecular detection of spotted fever group rickettsiae in hard ticks, northern China. Transbound. Emerg. Dis. 2019, 66, 1587–1596. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Guo, W.B.; Pan, Y.S.; Jiang, B.G.; Du, C.H.; Que, T.C.; Zhan, L.; Wu, J.H.; Yu, M.H.; Cui, X.M.; et al. Detection of novel spotted fever group Rickettsiae (Rickettsiales: Rickettsiaceae) in Ticks (Acari: Ixodidae) in southwestern China. J. Med. Entomol. 2021, 58, 1363–1369. [Google Scholar] [CrossRef]
- Cotes-Perdomo, A.P.; Oviedo, A.; Castro, L.R. Molecular detection of pathogens in ticks associated with domestic animals from the Colombian Caribbean region. Exp. Appl. Acarol. 2020, 82, 137–150. [Google Scholar] [CrossRef]
- Lu, M.; Tian, J.H.; Yu, B.; Guo, W.P.; Holmes, E.C.; Zhang, Y.Z. Extensive diversity of Rickettsiales bacteria in ticks from Wuhan, China. Ticks Tick-Borne Dis. 2017, 8, 574–580. [Google Scholar] [CrossRef] [PubMed]
- de Souza Ramos, I.A.; Herrera, H.M.; Fernandes, S.J.; do Amaral, R.B.; Zanatto, D.C.S.; da Silva, T.M.V.; Horta, B.L.S.; Campos, J.B.V.; Alves, J.V.A.; de Macedo, G.C.; et al. Genetic diversity of Anaplasma marginale in beef cattle in the Brazilian Pantanal. Ticks Tick-Borne Dis. 2019, 10, 805–814. [Google Scholar] [CrossRef]
- Lu, M.; Tian, J.; Pan, X.; Qin, X.; Wang, W.; Chen, J.; Guo, W.; Li, K. Identification of Rickettsia spp., Anaplasma spp., and an Ehrlichia canis-like agent in Rhipicephalus microplus from southwest and south-central China. Ticks Tick-Borne Dis. 2022, 13, 101884. [Google Scholar] [CrossRef]
- Aguiar, D.M.; Araujo, J.P.J.; Nakazato, L.; Bard, E.; Cabezas-Cruz, A. Complete genome sequence of an Ehrlichia minasensis strain isolated from cattle. Microbiol. Resour. Announc. 2019, 8, e00161-19. [Google Scholar] [CrossRef]
- Intirach, J.; Lv, X.; Sutthanont, N.; Cai, B.; Champakaew, D.; Chen, T.; Han, Q.; Lv, Z. Molecular and next-generation sequencing analysis of tick-borne pathogens of Rhipicephalus ticks (Acari: Ixodidae) in cattle and dogs. Acta Trop. 2024, 252, 107138. [Google Scholar] [CrossRef]
- Ghafar, A.; Cabezas-Cruz, A.; Galon, C.; Obregon, D.; Gasser, R.B.; Moutailler, S.; Jabbar, A. Bovine ticks harbour a diverse array of microorganisms in Pakistan. Parasit. Vectors 2020, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Liao, J.; Chen, Q.; Ding, J.; Chang, H.; Lyu, Y.; Yu, L.; Wen, B.; Sun, Y.; Qin, T. Diversity of Rickettsiales bacteria in five species of ticks collected from Jinzhai County, Anhui Province, China in 2021–2022. Front. Microbiol. 2023, 14, 1141217. [Google Scholar] [CrossRef] [PubMed]
- Dasch, G.A.; Eremeeva, M.E.; Zambrano, M.L.; Premaratna, R.; Kularatne, S.A.M.; Jayanthe Rajapakse, R.P.V. Molecular characterization of Rickettsial agents in ticks (Acari: Ixodidae) from Sri Lanka. Am. J. Trop. Med. Hyg. 2022, 106, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Blazekova, V.; Stanko, M.; Sprong, H.; Kohl, R.; Zubrikova, D.; Vargova, L.; Bona, M.; Miklisova, D.; Vichova, B. Ixodiphagus hookeri (Hymenoptera: Encyrtidae) and tick-borne pathogens in ticks with sympatric occurrence (and different activities) in the Slovak Karst National Park (Slovakia), Central Europe. Pathogens 2024, 13, 385. [Google Scholar] [CrossRef]
- Bona, M.; Blaňárová, L.; Stanko, M.; Mošanský, L.; Čepčeková, E.; Víchová, B. Impact of climate factors on the seasonal activity of ticks and temporal dynamics of tick-borne pathogens in an area with a large tick species diversity in Slovakia, Central Europe. Biologia 2021, 77, 1619–1631. [Google Scholar] [CrossRef]
- Susnjar, J.; Cerar Kisek, T.; Strasek Smrdel, K.; Ruzic-Sabljic, E.; Adam, K.; Ivovic, V. Detection, identification and genotyping of Borrelia spp. in ticks of Coastal-Karst and Littoral-Inner Carniola regions in Slovenia. Folia Parasitol. 2023, 70, 7. [Google Scholar] [CrossRef]
- Strnad, M.; Honig, V.; Ruzek, D.; Grubhoffer, L.; Rego, R.O.M. Europe-wide meta-analysis of Borrelia burgdorferi Sensu Lato prevalence in questing Ixodes ricinus ticks. Appl. Environ. Microbiol. 2017, 83, e00609-17. [Google Scholar] [CrossRef]
- Shu, C.; Intirach, J.; Zhou, Y.; Gao, S.; Lv, X.; Jiao, H.; Hu, Y.; Lv, Z. Microbial community characteristics and pathogens detection in Rhipicephalus sanguineus and Haemaphysalis hystricis from Hainan Island, China. Front. Microbiol. 2024, 15, 1450219. [Google Scholar] [CrossRef]
- Lu, M.; Meng, C.; Gao, X.; Sun, Y.; Zhang, J.; Tang, G.; Li, Y.; Li, M.; Zhou, G.; Wang, W.; et al. Diversity of Rickettsiales in Rhipicephalus microplus ticks collected in domestic ruminants in Guizhou Province, China. Pathogens 2022, 11, 1108. [Google Scholar] [CrossRef]
- Gual-Gonzalez, L.; Torres, M.E.; Self, S.C.W.; Cantillo-Barraza, O.; Nolan, M.S. Spotted fever group Rickettsia spp. molecular and serological evidence among Colombian vectors and animal hosts: A historical review. Insects 2024, 15, 170. [Google Scholar] [CrossRef]
- Seo, J.Y.; Park, J.S.; Lee, H.I.; Ju, J.W. Molecular identification of spotted fever group Rickettsiae in ticks in the Republic of Korea. Pathogens 2024, 13, 575. [Google Scholar] [CrossRef]
- Byaruhanga, C.; Collins, N.E.; Knobel, D.L.; Khumalo, Z.T.H.; Chaisi, M.E.; Oosthuizen, M.C. Molecular detection and phylogenetic analysis of Anaplasma marginale and Anaplasma centrale amongst transhumant cattle in north-eastern Uganda. Ticks Tick-Borne Dis. 2018, 9, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Han, R.; Liu, Z.; Niu, Q.; Guan, G.; Liu, G.; Luo, J.; Yin, H. Insight into the genetic diversity of Anaplasma marginale in cattle from ten provinces of China. Parasit. Vectors 2017, 10, 565. [Google Scholar] [CrossRef]
- Al-Hosary, A.; Raileanu, C.; Tauchmann, O.; Fischer, S.; Nijhof, A.M.; Silaghi, C. Epidemiology and genotyping of Anaplasma marginale and co-infection with piroplasms and other Anaplasmataceae in cattle and buffaloes from Egypt. Parasit. Vectors 2020, 13, 495. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Pan, K.; Teng, Z.; Wang, H.; Zhang, X.; Ren, H.; Yi, L.; He, J.; Cai, K.; Qin, T. Molecular detection reveals diverse tick-borne bacterial and protozoan pathogens in two tick species from Yingshan County of Hubei Province, China in 2021–2022. Front Microbiol 2023, 14, 1298037. [Google Scholar] [CrossRef] [PubMed]
- Cruz, A.C.; Zweygarth, E.; Ribeiro, M.F.; da Silveira, J.A.; de la Fuente, J.; Grubhoffer, L.; Valdes, J.J.; Passos, L.M. New species of Ehrlichia isolated from Rhipicephalus (Boophilus) microplus shows an ortholog of the E. canis major immunogenic glycoprotein gp36 with a new sequence of tandem repeats. Parasit. Vectors 2012, 5, 291. [Google Scholar] [CrossRef]
- Cabezas-Cruz, A.; Zweygarth, E.; Broniszweska, M.; Passos, L.M.; Ribeiro, M.F.; Manrique, M.; Tobes, R.; de la Fuente, J. Complete genome sequence of Ehrlichia mineirensis, a novel organism closely related to Ehrlichia canis with a new host association. Genome Announc. 2015, 3, e01450-14. [Google Scholar] [CrossRef]
- Sgroi, G.; D’Alessio, N.; Veneziano, V.; Rofrano, G.; Fusco, G.; Carbonara, M.; Dantas-Torres, F.; Otranto, D.; Iatta, R. Ehrlichia canis in human and tick, Italy, 2023. Emerg. Infect. Dis. 2024, 30, 2651–2654. [Google Scholar] [CrossRef]
- Shi, W.; Zhang, Y. The distribution, protection and utilization of karst caves in Hainan province, China. In Proceedings of the 16th International Congress of Speleology, Guilin, China, 21–28 July 2013. [Google Scholar]
- Galvão, P.; Halihan, T.; Hirata, R. Evaluating karst geotechnical risk in the urbanized area of Sete Lagoas, Minas Gerais, Brazil. Hydrogeol. J. 2015, 23, 1499–1513. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Sites | No. of Tested | Pathogens (%, 95% CI) | ||||
|---|---|---|---|---|---|---|
| Candidatus Rickettsia Jingxinensis | Anaplasma marginale | Candidatus Ehrlichia Carsus | Ehrlichia minasensis | Ehrlichia canis | ||
| Wengduo, Guiding | 36 | 34 (94.4, 81.9–98.5) | 0 | 7 (19.4, 9.8–35.0) | 1 (2.8, 0.5–14.2) | 0 |
| Jishangtang, Weng’an | 36 | 31 (86.1, 71.3–93.9) | 0 | 13 (36.1, 22.5–52.4) | 0 | 0 |
| Shangzhai, Weng’an | 57 | 53 (93.0, 83.3–97.2) | 30 (52.6, 39.9–65.0) | 4 (7.0, 2.8–16.7) | 8 (14.0, 7.3–25.3) | 0 |
| Baiqi, Huishui | 6 | 6 (100, 61.0–100) | 0 | 0 | 0 | 0 |
| Qingshan, Huishui | 21 | 20 (95.2, 77.3–99.2) | 0 | 0 | 0 | 0 |
| Yangmao, Huishui | 24 | 20 (83.3, 64.2–93.3) | 2 (8.3, 2.3–25.9) | 2 (8.3, 2.3–25.9) | 2 (8.3, 2.3–25.9) | 0 |
| Qunli, Libo | 17 | 16 (94.1, 73.0–99.0) | 0 | 3 (17.7, 6.2–41.0) | 0 | 4 (23.5, 9.6–47.3) |
| Dapo, Xifeng | 56 | 49 (87.5, 76.4–93.8) | 10 (17.9, 10.0–29.8) | 15 (26.8, 17.0–39.6) | 1 (1.8, 0.3–9.5) | 0 |
| Poyan, Qingzheng | 21 | 20 (95.2, 77.3–99.2) | 0 | 2(9.5, 2.7–28.9) | 1 (4.8, 0.9–22.7) | 0 |
| Total | 274 | 249 (90.9, 86.9–93.7) | 42 (15.3, 11.5–20.1) | 46 (16.8, 12.8–21.7) | 13 (4.7, 2.5–7.5) | 4 (1.5, 0.6–3.7) |
| Pathogen Co-Infections | No. of Positive | Positive Rate (95% CI) |
|---|---|---|
| Ca. R. jingxinensis + A. marginale | 38 | 13.9% (10.3–18.5) |
| Ca. R. jingxinensis + E. canis | 4 | 1.5% (0.6–3.7) |
| Ca. R. jingxinensis + E. minasensis | 12 | 4.4% (2.5–7.5) |
| Ca. R. jingxinensis + Ca. E. carsus | 39 | 14.2% (10.6–18.9) |
| Ca. R. jingxinensis + A. marginale + Ca. E. carsus | 1 | 0.4% (0.1–2.0) |
| Total | 94 | 34.3% (28.9–40.1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.-T.; Wang, Y.-F.; Zhang, M.-Z.; Zhu, D.-Y.; Sun, Y.; Gong, C.-W.; Zhan, L.; Cui, X.-M.; Cao, W.-C. High Diversity and Prevalence of Rickettsial Agents in Rhipicephalus microplus Ticks from Livestock in Karst Landscapes of Southwest China. Microorganisms 2025, 13, 765. https://doi.org/10.3390/microorganisms13040765
Liu Y-T, Wang Y-F, Zhang M-Z, Zhu D-Y, Sun Y, Gong C-W, Zhan L, Cui X-M, Cao W-C. High Diversity and Prevalence of Rickettsial Agents in Rhipicephalus microplus Ticks from Livestock in Karst Landscapes of Southwest China. Microorganisms. 2025; 13(4):765. https://doi.org/10.3390/microorganisms13040765
Chicago/Turabian StyleLiu, Ya-Ting, Yi-Fei Wang, Ming-Zhu Zhang, Dai-Yun Zhu, Yi Sun, Cai-Wei Gong, Lin Zhan, Xiao-Ming Cui, and Wu-Chun Cao. 2025. "High Diversity and Prevalence of Rickettsial Agents in Rhipicephalus microplus Ticks from Livestock in Karst Landscapes of Southwest China" Microorganisms 13, no. 4: 765. https://doi.org/10.3390/microorganisms13040765
APA StyleLiu, Y.-T., Wang, Y.-F., Zhang, M.-Z., Zhu, D.-Y., Sun, Y., Gong, C.-W., Zhan, L., Cui, X.-M., & Cao, W.-C. (2025). High Diversity and Prevalence of Rickettsial Agents in Rhipicephalus microplus Ticks from Livestock in Karst Landscapes of Southwest China. Microorganisms, 13(4), 765. https://doi.org/10.3390/microorganisms13040765