
Regulatory Mechanisms of Exogenous Acyl-Homoserine Lactones in the Aerobic Ammonia Oxidation Process Under Stress Conditions

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Reactor Operation and Experimental Design
2.2. Illumina MiSeq Sequencing and Sequence Processing
2.3. qPCR Assay of amoA Genes
- -
- AOA amoA genes: archaea-amoA F/R (Francis et al., 2005);
- -
- AOB amoA genes: amoA-1F/1R (Rotthauwe et al., 1997);
- -
- Comammox amoA genes: com amoA AF/SR (Shao and Wu, 2021).
3. Results and Discussion
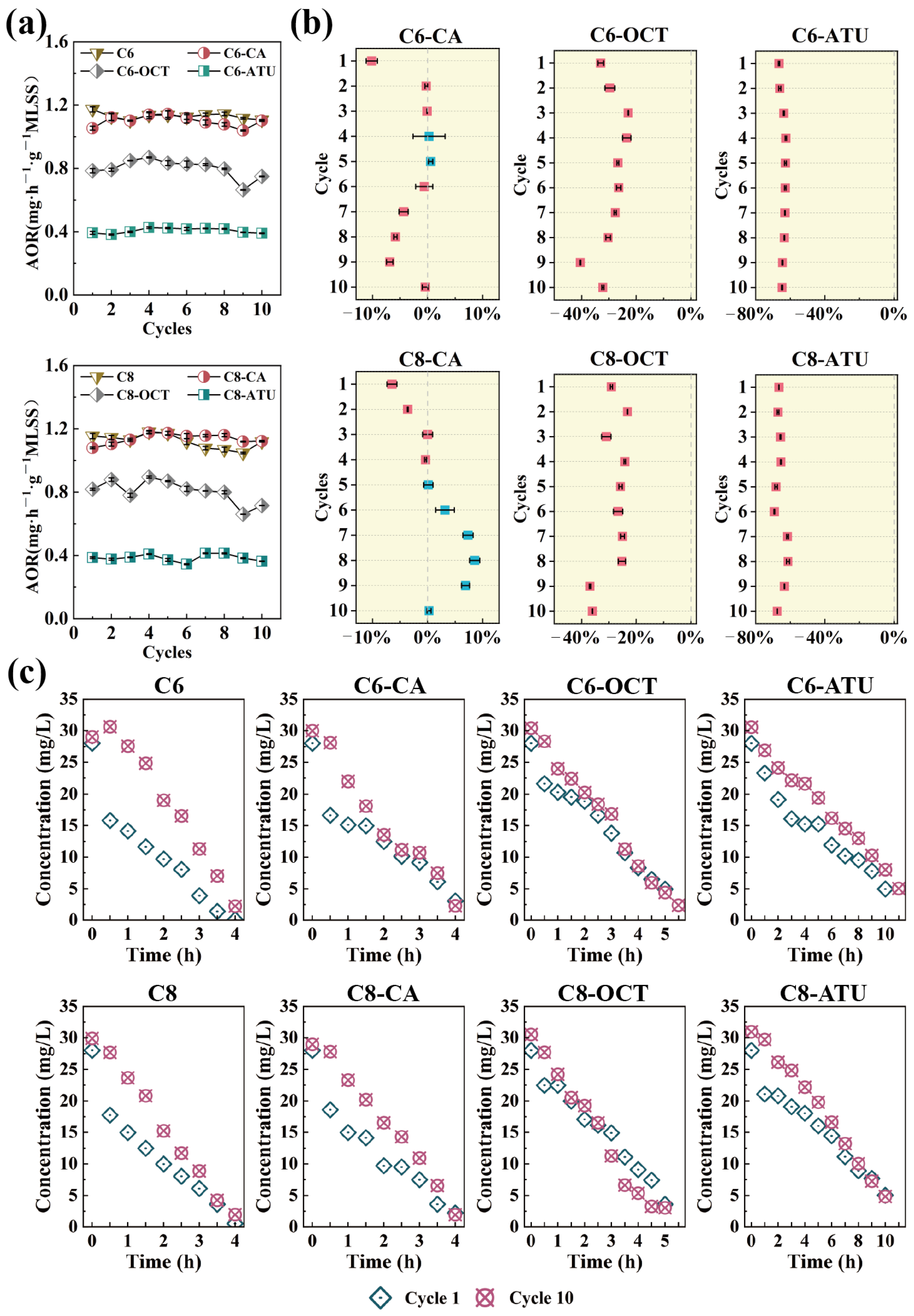
3.1. Effects of AHLs on Ammonia Oxidization
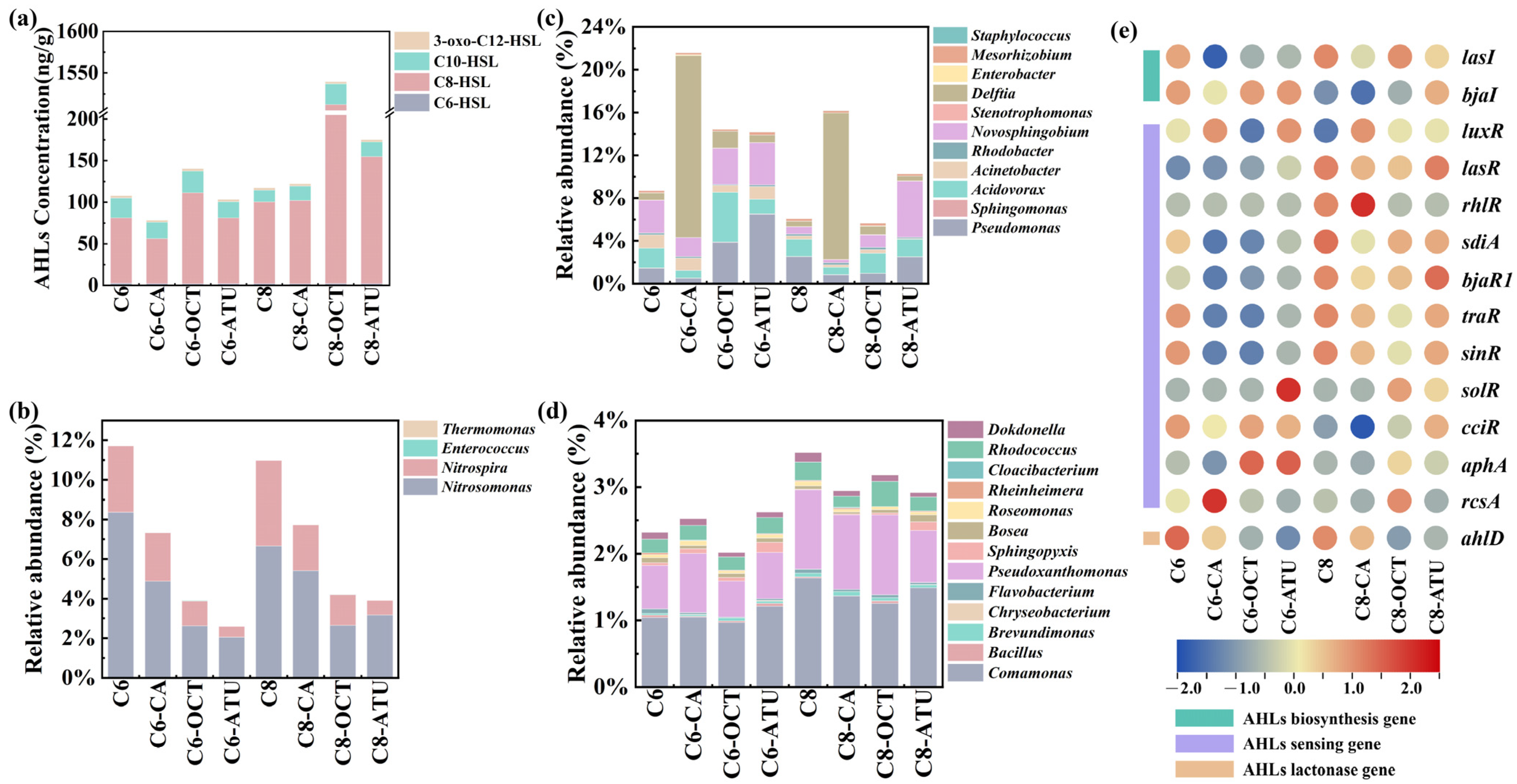
3.2. Functional Gene Abundance and AOM Diversity
3.3. Impact of AHL Addition on QS/QQ Bacteria and Genes
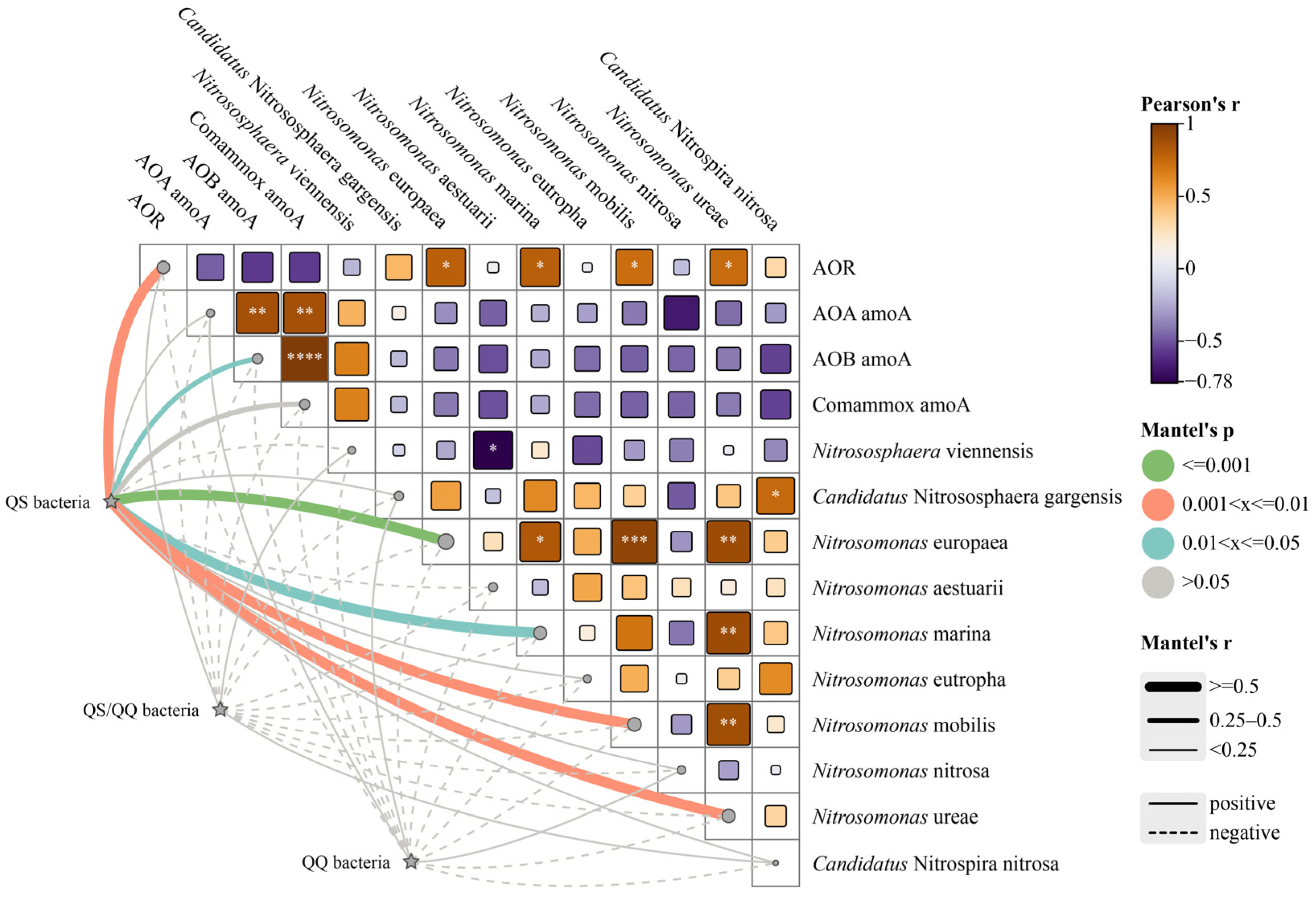
3.4. Potential Interactions Between AHL-Related Bacteria and the Ammonia Oxidation Process
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luo, Z.; Hu, S.; Chen, D.; Zhu, B. From Production to Consumption: A Coupled Human-Environmental Nitrogen Flow Analysis in China. Environ. Sci. Technol. 2018, 52, 2025–2035. [Google Scholar] [CrossRef] [PubMed]
- Wurtsbaugh, W.A.; Paerl, H.W.; Dodds, W.K. Nutrients, eutrophication and harmful algal blooms along the freshwater to marine continuum. Wiley Interdiscip. Rev. Water 2019, 6, e1373. [Google Scholar] [CrossRef]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.K.; Straka, L. New directions in biological nitrogen removal and recovery from wastewater. Curr. Opin. Biotechnol. 2019, 57, 50–55. [Google Scholar] [CrossRef]
- Martikainen, P.J. Heterotrophic nitrification–An eternal mystery in the nitrogen cycle. Soil Biol. Biochem. 2022, 168, 108611. [Google Scholar] [CrossRef]
- He, S.; Zhao, Z.; Tian, Z.; Xu, C.; Liu, Y.; He, D.; Zhang, Y.; Zheng, M. Comammox bacteria predominate among ammonia-oxidizing microorganisms in municipal but not in refinery wastewater treatment plants. J. Environ. Manag. 2022, 316, 115271. [Google Scholar] [CrossRef]
- Lawson, C.E.; Lücker, S. Complete ammonia oxidation: An important control on nitrification in engineered ecosystems? Curr. Opin. Biotechnol. 2018, 50, 158–165. [Google Scholar] [CrossRef]
- Fan, X.Y.; Gao, J.; Pan, K.; Li, D.; Dai, H.; Li, X. Temporal heterogeneity and temperature response of active ammonia-oxidizing microorganisms in winter in full-scale wastewater treatment plants. Chem. Eng. J. 2019, 360, 1542–1552. [Google Scholar] [CrossRef]
- Zhao, J.; Bello, M.O.; Meng, Y.; Prosser, J.I.; Gubry-Rangin, C. Selective inhibition of ammonia oxidising archaea by simvastatin stimulates growth of ammonia oxidising bacteria. Soil Biol. Biochem. 2020, 141, 107673. [Google Scholar] [CrossRef]
- Johnston, J.; Du, Z.; Behrens, S. Ammonia-Oxidizing Bacteria Maintain Abundance but Lower amoA-Gene Expression during Cold Temperature Nitrification Failure in a Full-Scale Municipal Wastewater Treatment Plant. Microbiol. Spectr. 2023, 11, e02571-22. [Google Scholar] [CrossRef]
- Di, H.J.; Cameron, K.C.; Podolyan, A.; Robinson, A. Effect of soil moisture status and a nitrification inhibitor, dicyandiamide, on ammonia oxidizer and denitrifier growth and nitrous oxide emissions in a grassland soil. Soil Biol. Biochem. 2014, 73, 59–68. [Google Scholar] [CrossRef]
- Zhao, C.; Li, J.; Li, C.; Xue, B.; Wang, S.; Zhang, X.; Yang, X.; Shen, Z.; Bo, L.; Qiu, Z.; et al. Horizontal transfer of the multidrug resistance plasmid RP4 inhibits ammonia nitrogen removal dominated by ammonia-oxidizing bacteria. Water Res. 2022, 217, 118434. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Fujiwara, T.; Hidaka, T.; Nishimura, F.; Nakanishi, T.; Terada, A.; Hori, T. Evaluation of nitrous oxide emission during ammonia retention from simulated industrial wastewater by microaerobic activated sludge process. Water Res. 2023, 247, 120780. [Google Scholar] [CrossRef]
- Sauder, L.A.; Ross, A.A.; Neufeld, J.D. Nitric oxide scavengers differentially inhibit ammonia oxidation in ammonia-oxidizing archaea and bacteria. Fems Microbiol. Lett. 2016, 363, fnw052. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chapman, S.J.; Nicol, G.W.; Yao, H. Nitrification and nitrifiers in acidic soils. Soil Biol. Biochem. 2018, 116, 290–301. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, S.; Ma, S.; Zheng, X.; Wang, Z.; Lu, C. Effects of commonly used nitrification inhibitors—Dicyandiamide (DCD), 3,4-dimethylpyrazole phosphate (DMPP), and nitrapyrin—On soil nitrogen dynamics and nitrifiers in three typical paddy soils. Geoderma 2020, 380, 114637. [Google Scholar] [CrossRef]
- Ginestet, P.; Audic, J.M.; Urbain, V.V.; Block, J.C. Estimation of Nitrifying Bacterial Activities by Measuring Oxygen Uptake in the Presence of the Metabolic Inhibitors Allylthiourea and Azide. Appl. Environ. Microbiol. 1998, 64, 2266–2268. [Google Scholar] [CrossRef]
- Taylor, A.E.; Vajrala, N.; Giguere, A.T.; Gitelman, A.I.; Bottomley, P.J. Use of Aliphatic n-Alkynes To Discriminate Soil Nitrification Activities of Ammonia-Oxidizing Thaumarchaea and Bacteria. Appl. Environ. Microbiol. 2013, 79, 6544–6551. [Google Scholar] [CrossRef]
- Guo, T.; Bai, S.H.; Omidvar, N.; Wang, Y.; Chen, F.; Zhang, M. Insight into the functional mechanisms of nitrogen-cycling inhibitors in decreasing yield-scaled ammonia volatilization and nitrous oxide emission: A global meta-analysis. Chemosphere 2023, 338, 139611. [Google Scholar] [CrossRef]
- Ma, X.; Liu, D.; Chu, X.; Huang, J.; Shu, Z.; Li, Y.; Jin, Y. Effects of exogenous signaling molecules on anaerobic sludge digestion: Quorum sensing and antibiotic resistance genes. Bioresour. Technol. 2024, 414, 131624. [Google Scholar] [CrossRef]
- Xu, C.; Ni, L.; Du, C.; Shi, J.; Ma, Y.; Li, S.; Li, Y. Decoding Microcystis aeruginosa quorum sensing through AHL-mediated transcriptomic molecular regulation mechanisms. Sci. Total Environ. 2024, 926, 172101. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Chen, J.; Liu, X.; Gao, W.; Sun, L.; Wang, P.; Ren, Z.; Zhang, G.; Li, W. Roles and regulation of quorum sensing in anaerobic granular sludge: Research status, challenges, and perspectives. Bioresour. Technol. 2023, 387, 129644. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wang, X.; Chen, H.; Wang, M.; Wang, Y.; Chen, H.; Dai, H. Transcriptome analysis expands underlying mechanisms of quorum sensing mediating heterotrophic nitrification-aerobic denitrification process at low temperature. Bioresour. Technol. 2024, 414, 131581. [Google Scholar] [CrossRef] [PubMed]
- Maddela, N.R.; Sheng, B.; Yuan, S.; Zhou, Z.; Villamar-Torres, R.; Meng, F. Roles of quorum sensing in biological wastewater treatment: A critical review. Chemosphere 2019, 221, 616–629. [Google Scholar] [CrossRef]
- Tang, X.; Liu, S.; Zhang, Z.; Zhuang, G. Identification of the release and effects of AHLs in anammox culture for bacteria communication. Chem. Eng. J. 2015, 273, 184–191. [Google Scholar] [CrossRef]
- Sun, Y.; Guan, Y.; Zeng, D.; He, K.; Wu, G. Metagenomics-based interpretation of AHLs-mediated quorum sensing in Anammox biofilm reactors for low-strength wastewater treatment. Chem. Eng. J. 2018, 344, 42–52. [Google Scholar] [CrossRef]
- Feng, Z.; Sun, Y.; Li, T.; Meng, F.; Wu, G. Operational pattern affects nitritation, microbial community and quorum sensing in nitrifying wastewater treatment systems. Sci. Total Environ. 2019, 677, 456–465. [Google Scholar] [CrossRef]
- Gao, P.; Zhao, A.; Zhang, X.; Tang, P.; Li, D.; Liu, T.; Li, J.; Zhu, Y.; Wang, Z. Potential role of N-acyl homoserine lactone-mediated quorum sensing in the adaptation of anammox granular sludge system to salinity stress. Bioresour. Technol. 2025, 416, 131758. [Google Scholar] [CrossRef]
- Li, A.J.; Hou, B.L.; Li, M.X. Cell adhesion, ammonia removal and granulation of autotrophic nitrifying sludge facilitated by N-acyl-homoserine lactones. Bioresour. Technol. 2015, 196, 550–558. [Google Scholar] [CrossRef]
- De Clippeleir, H.; Defoirdt, T.; Vanhaecke, L.; Vlaeminck, S.E.; Carballa, M.; Verstraete, W.; Boon, N. Long-chain acylhomoserine lactones increase the anoxic ammonium oxidation rate in an OLAND biofilm. Appl. Microbiol. Biotechnol. 2011, 90, 1511–1519. [Google Scholar] [CrossRef]
- Wang, N.; Gao, J.; Liu, Y.; Wang, Q.; Zhuang, G. Realizing the role of N-acyl-homoserine lactone-mediated quorum sensing in nitrification and denitrification: A review. Chemosphere 2021, 274, 129970. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; He, J.; Yu, H.; Liu, J.; Zhang, J. A strategy to speed up formation and strengthen activity of biofilms at low temperature. RSC Adv. 2017, 37, 22788–22796. [Google Scholar] [CrossRef]
- Zhou, Q.; Xie, X.; Feng, F.; Huang, S.; Sun, Y. Impact of acyl-homoserine lactones on the response of nitrogen cycling in sediment to florfenicol stress. Sci. Total Environ. 2021, 785, 147294. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Q.; Dong, D.; Hu, H.; Ren, H. AHLs-mediated quorum sensing threshold and its response towards initial adhesion of wastewater biofilms. Water Res. 2021, 194, 116925. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, L.; Sun, P.; He, J.; Li, J. Effects of AHLs inhibitors and exogenous AHLs on the stability and activity of Anammox granules at low temperatures. Water Environ. Res. 2021, 93, 1576–1588. [Google Scholar] [CrossRef]
- Li, C.; Li, W.; Li, H.; Hou, M.; Wu, X.; Zhuang, J.; Liu, Y. The effect of quorum sensing on performance of salt-tolerance aerobic granular sludge: Linking extracellular polymeric substances and microbial community. Biodegradation 2019, 30, 447–456. [Google Scholar] [CrossRef]
- Gao, J.; Duan, Y.; Liu, Y.; Zhuang, X.; Liu, Y.; Bai, Z.; Ma, W.; Zhuang, G. Long- and short-chain AHLs affect AOA and AOB microbial community composition and ammonia oxidation rate in activated sludge. J. Environ. Sci. 2019, 78, 53–62. [Google Scholar] [CrossRef]
- Hu, H.; He, J.; Liu, J.; Yu, H.; Zhang, J. Biofilm activity and sludge characteristics affected by exogenous N-acyl homoserine lactones in biofilm reactors. Bioresour. Technol. 2016, 211, 339–347. [Google Scholar] [CrossRef]
- Yan, X.; Zheng, S.; Huo, Z.; Shi, B.; Huang, J.; Yang, J.; Ma, J.; Han, Y.; Wang, Y.; Cheng, K. Effects of exogenous N-acyl-homoserine lactones on nutrient removal, sludge properties and microbial community structures during activated sludge process. Chemosphere 2020, 255, 126945. [Google Scholar] [CrossRef]
- Zeng, X.; Hu, H. Potential roles of acyl homoserine lactones (AHLs) in nitrifying bacteria survival under certain adverse circumstances. Sci. Rep. 2023, 13, 705. [Google Scholar] [CrossRef]
- Burton, E.O.; Read, H.W.; Pellitteri, M.C.; Hickey, W.J. Identification of acyl-homoserine lactone signal molecules produced by Nitrosomonas europaea strain Schmidt. Appl. Environ. Microbiol. 2005, 71, 4906–4909. [Google Scholar] [CrossRef] [PubMed]
- Yeon, K.M.; Cheong, W.S.; Oh, H.S.; Lee, W.N.; Hwang, B.K.; Lee, C.H.; Beyenal, H.; Lewandowski, Z. Quorum Sensing: A New Biofouling Control Paradigm in a Membrane Bioreactor for Advanced Wastewater Treatment. Environ. Sci. Technol. 2009, 43, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Decho, A.W.; Norman, R.S.; Visscher, P.T. Quorum sensing in natural environments: Emerging views from microbial mats. Trends Microbiol. 2010, 18, 73–80. [Google Scholar] [CrossRef]
- Tan, C.H.; Koh, K.S.; Xie, C.; Zhang, J.; Kjelleberg, S. Community quorum sensing signalling and quenching: Microbial granular biofilm assembly. Npj Biofilms Microbiomes 2015, 1, 15006. [Google Scholar] [CrossRef]
- Ishizaki, S.; Sugiyama, R.; Okabe, S. Membrane fouling induced by AHL-mediated soluble microbial product (SMP) formation by fouling-causing bacteria co-cultured with fouling-enhancing bacteria. Sci. Rep. 2017, 7, 8612. [Google Scholar] [CrossRef]
- Li, Y.; Cao, J.; Li, B.; Li, W.; Fang, F.; Tong, Z.; Yu, H. Outcompeting Presence of Acyl-Homoserine-Lactone (AHL)- Quenching Bacteria over AHL-Producing Bacteria in Aerobic Granules. Environ. Sci. Technol. Lett. 2020, 3, 36–40. [Google Scholar] [CrossRef]
- Mellbye, B.L.; Spieck, E.; Bottomley, P.J.; Sayavedrasoto, L.A. Acyl-Homoserine Lactone Production in Nitrifying Bacteria of the Genera Nitrosospira, Nitrobacter, and Nitrospira Identified via a Survey of Putative Quorum-Sensing Genes. Appl. Environ. Microbiol. 2017, 83, e01540-17. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibitor and Dose | Blank Control | Caffeic Acid, 100 μm | 1-Octyne, 0.03% v/v | ATU, 100 μm |
|---|---|---|---|---|
| C6-HSL, 50 μM (GROUP 1) | C6 | C6-CA | C6-OCT | C6-ATU |
| C8-HSL, 50 μM (GROUP 2) | C8 | C8-CA | C8-OCT | C8-ATU |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, C.; Pan, K.; Wei, Y.; Zhou, X.; Su, Q.; Bi, X.; Ng, H. Regulatory Mechanisms of Exogenous Acyl-Homoserine Lactones in the Aerobic Ammonia Oxidation Process Under Stress Conditions. Microorganisms 2025, 13, 663. https://doi.org/10.3390/microorganisms13030663
Qiu C, Pan K, Wei Y, Zhou X, Su Q, Bi X, Ng H. Regulatory Mechanisms of Exogenous Acyl-Homoserine Lactones in the Aerobic Ammonia Oxidation Process Under Stress Conditions. Microorganisms. 2025; 13(3):663. https://doi.org/10.3390/microorganisms13030663
Chicago/Turabian StyleQiu, Chen, Kailing Pan, Yuxuan Wei, Xiaolin Zhou, Qingxian Su, Xuejun Bi, and Howyong Ng. 2025. "Regulatory Mechanisms of Exogenous Acyl-Homoserine Lactones in the Aerobic Ammonia Oxidation Process Under Stress Conditions" Microorganisms 13, no. 3: 663. https://doi.org/10.3390/microorganisms13030663
APA StyleQiu, C., Pan, K., Wei, Y., Zhou, X., Su, Q., Bi, X., & Ng, H. (2025). Regulatory Mechanisms of Exogenous Acyl-Homoserine Lactones in the Aerobic Ammonia Oxidation Process Under Stress Conditions. Microorganisms, 13(3), 663. https://doi.org/10.3390/microorganisms13030663