
A Bioluminescence-Based Serum Bactericidal Assay to Detect Bactericidal Antibodies Against Neisseria meningitidis in Human Sera


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacteria
2.2. Serum Samples
2.3. Complement Source Validation Test
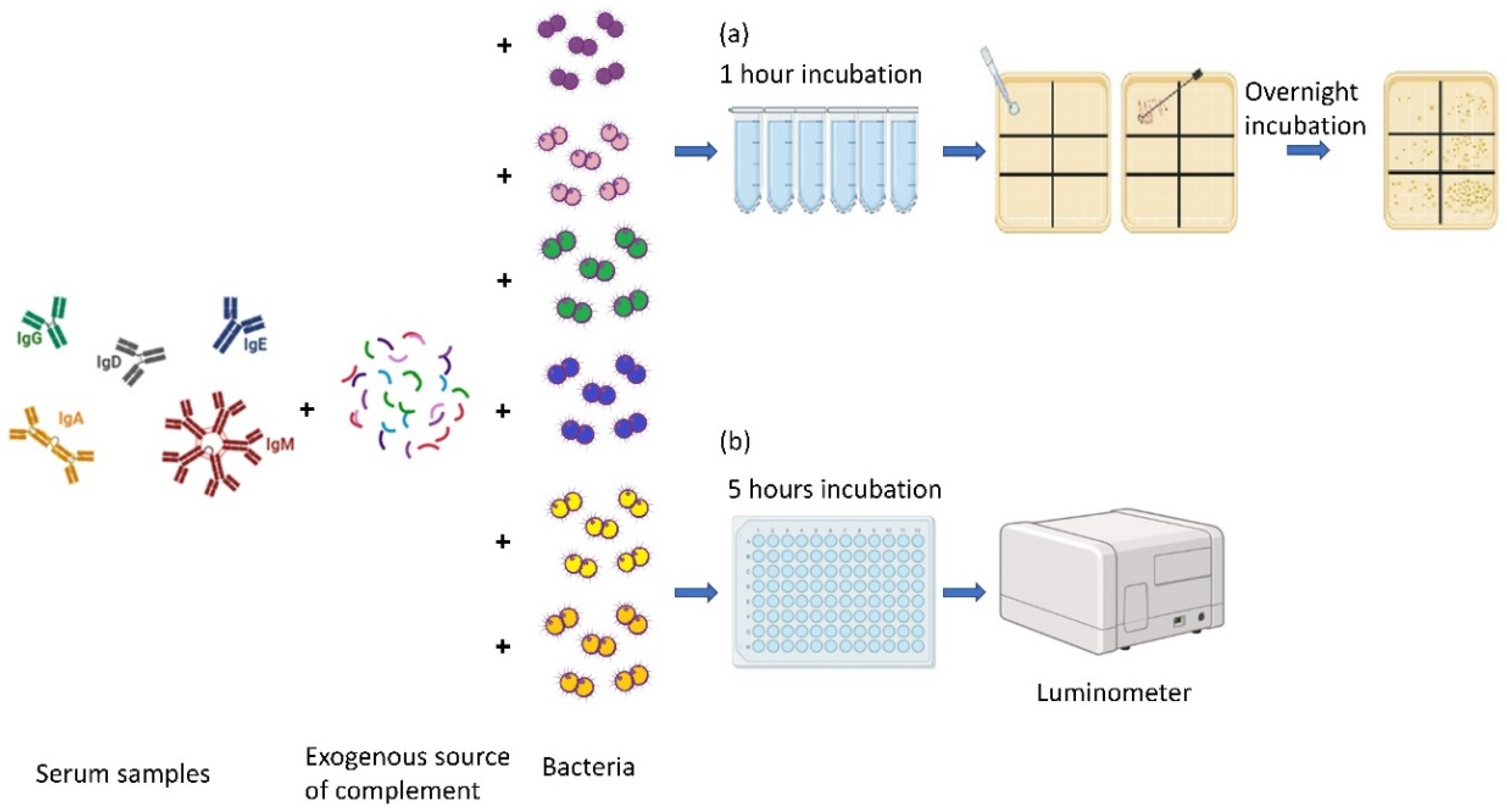
2.4. Conventional SBA
2.5. Bioluminescence SBA (BioLux-SBA)
2.6. Terminal Complement Complex Deposition Assay
2.7. Calculations
3. Results
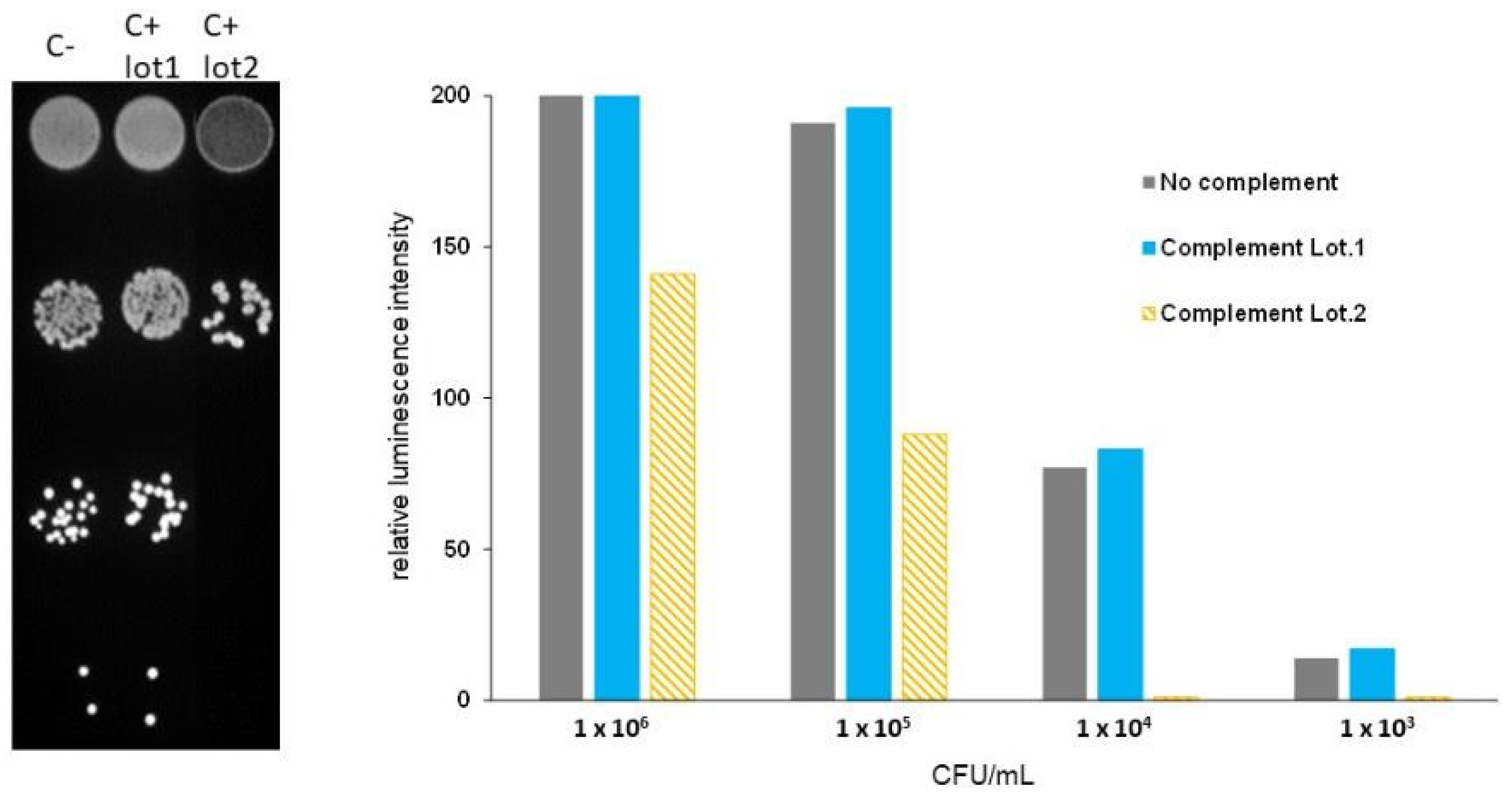
3.1. Complement Source Validation Assay
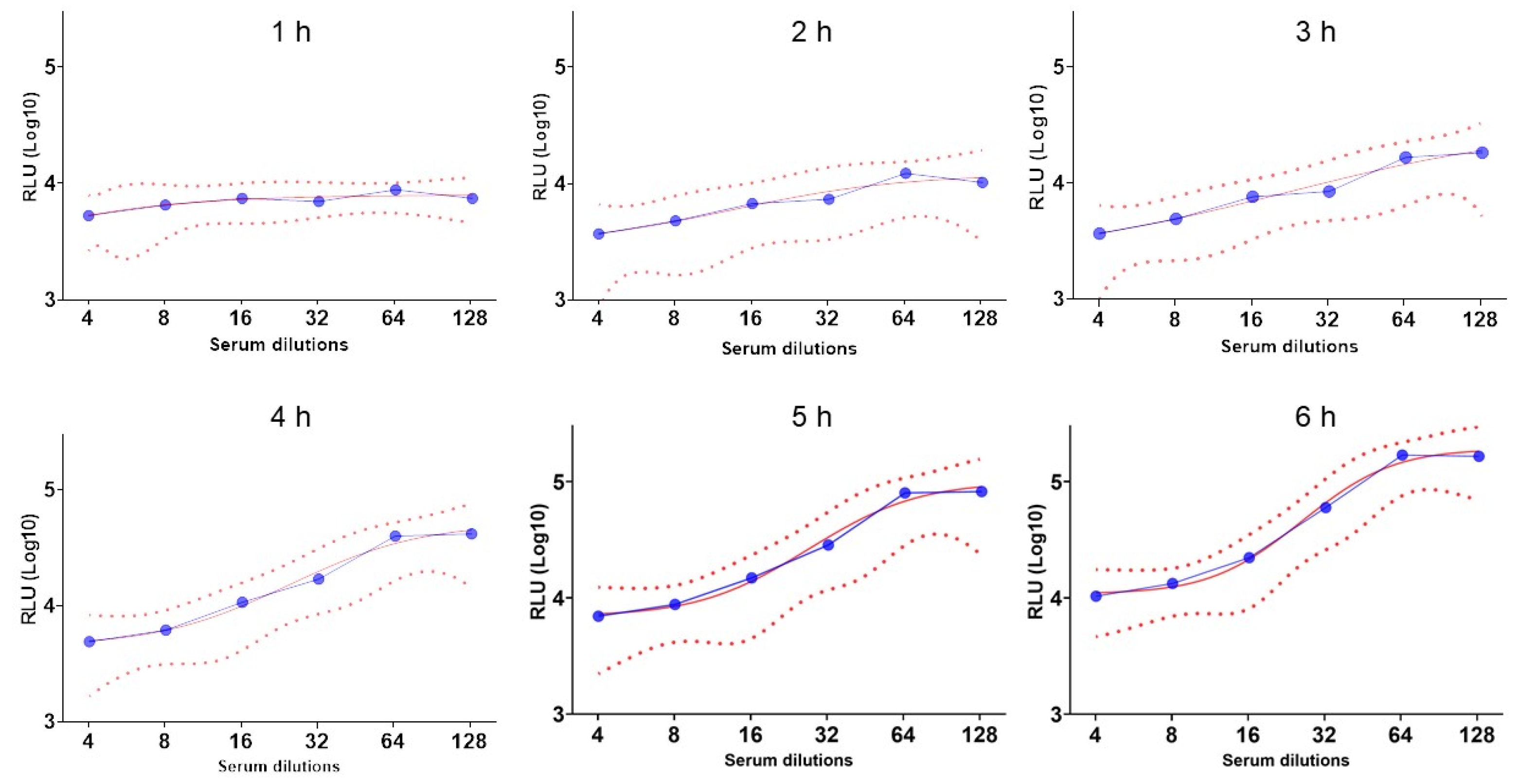
3.2. Set Up the Incubation Time During the BioLux-SBA
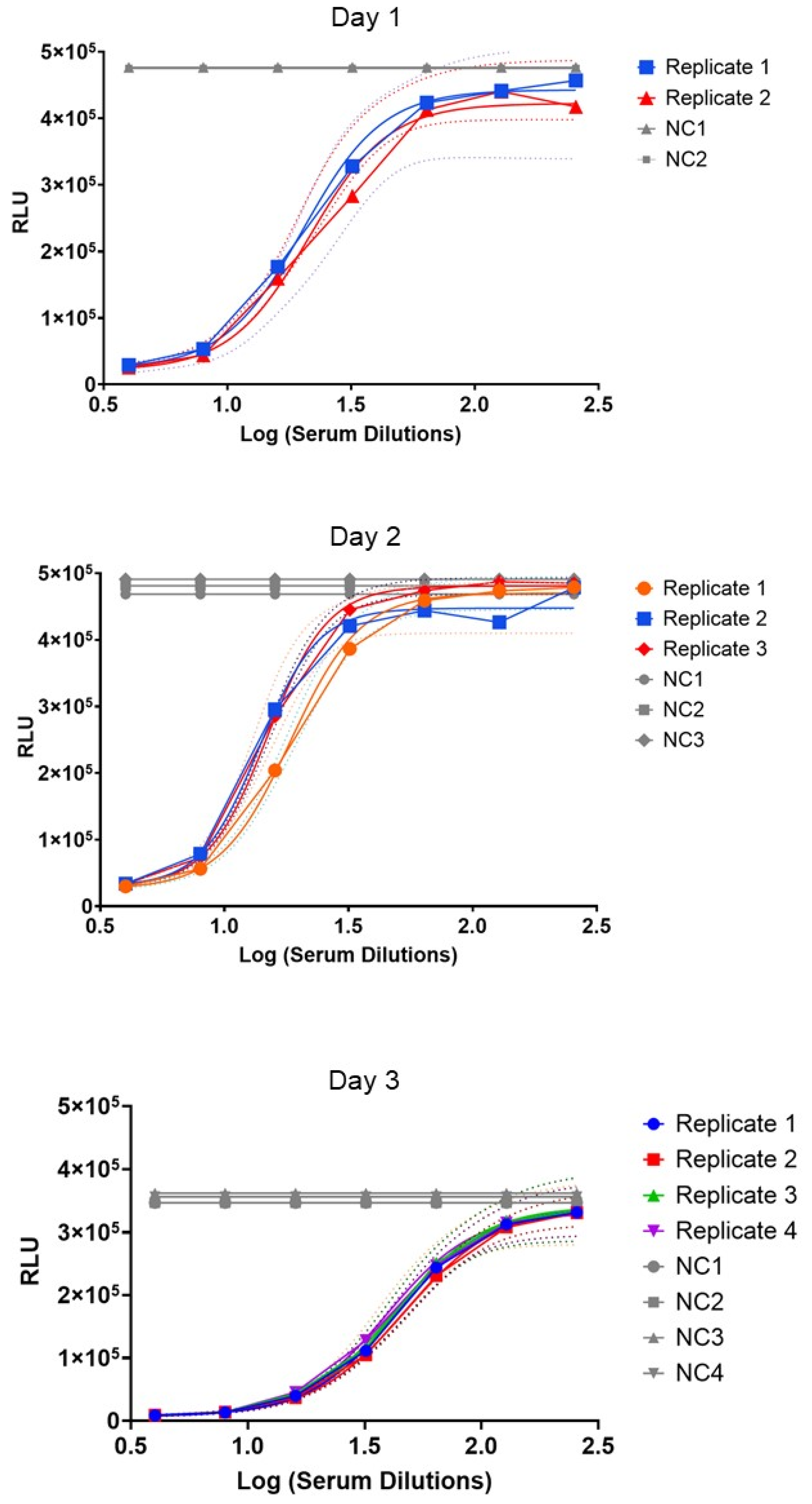
3.3. Repeatability and Intermediate Precision of the BioLux-SBA
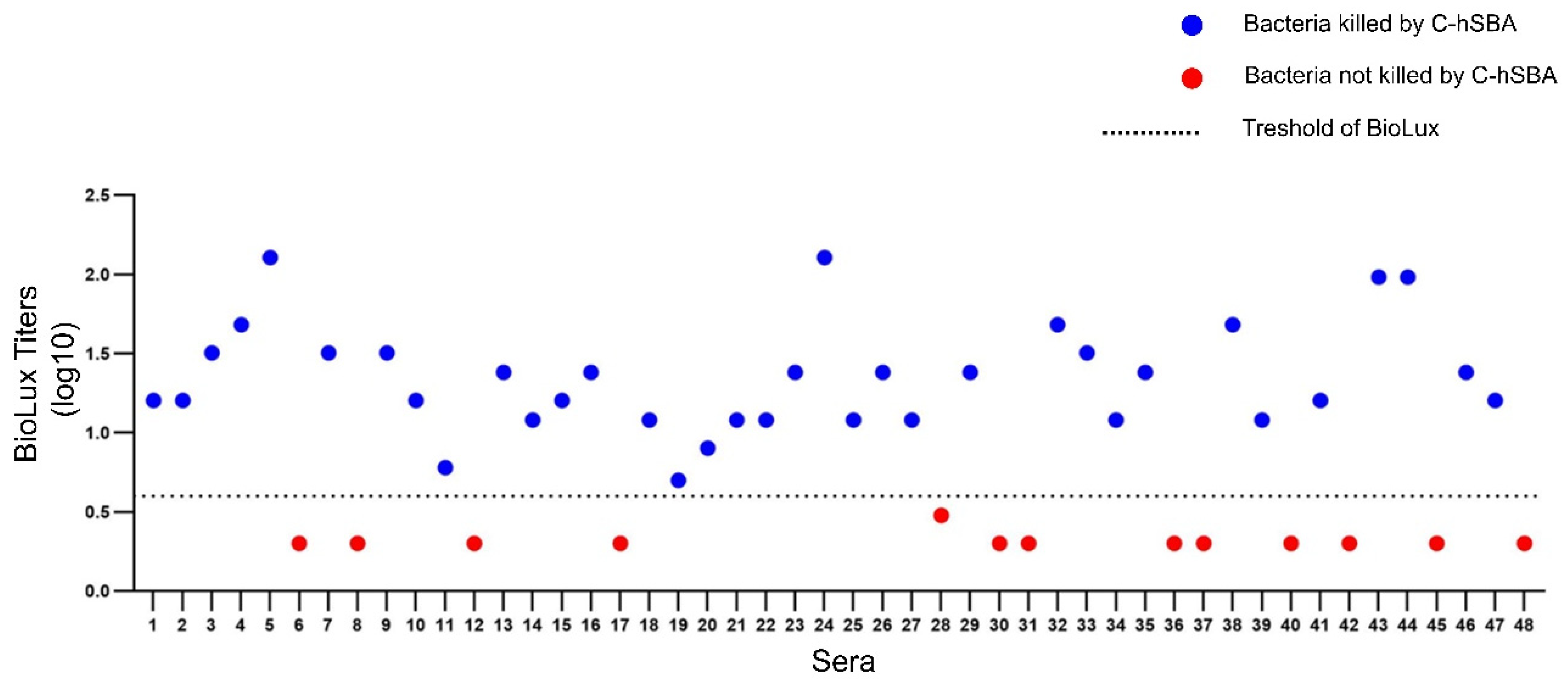
3.4. Correlation Between C-hSBA and BioLux-hSBA
3.5. Defining the Threshold of BioLux-hSBA
3.6. Terminal Complement Complex Deposition Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hollingshead, S.; Tang, C.M. An Overview of Neisseria meningitidis. In Neisseria meningitidis; Methods in Molecular Biology; Humana: New York, NY, USA, 2019; Volume 1969, pp. 1–16. [Google Scholar] [CrossRef]
- Borrow, R.; Carlone, G.M.; Rosenstein, N.; Blake, M.; Feavers, I.; Martin, D.; Zollinger, W.; Robbins, J.; Aaberge, I.; Granoff, D.M.; et al. Neisseria meningitidis group B correlates of protection and assay standardization—International meeting report Emory University, Atlanta, Georgia, United States, 16–17 March 2005. Vaccine 2006, 24, 5093–5107. [Google Scholar] [CrossRef] [PubMed]
- Ladhani, S.N.; Campbell, H.; Andrews, N.; Parikh, S.R.; White, J.; Edelstein, M.; Clark, S.A.; Lucidarme, J.; Borrow, R.; Ramsay, M.E. First real world evidence of meningococcal group B vaccine, 4CMenB, protection against meningococcal group W disease; prospective enhanced national surveillance, England. Clin. Infect. Dis. 2021, 73, e1661–e1668. [Google Scholar] [CrossRef] [PubMed]
- Heist, G.D.; Solis-Cohen, S.; Solis-Cohen, M. A study of the virulence of meningococci for man and of human susceptibility to meningococcic infection. J. Immunol. 1922, 7, 1–33. [Google Scholar] [CrossRef]
- Goldschneider, I.; Gotschlich, E.C.; Artenstein, M.S. Human immunity to the meningococcus. I. The role of humoral antibodies. J. Exp. Med. 1969, 129, 1307–1326. [Google Scholar] [CrossRef]
- Findlow, J.; Lucidarme, J.; Taha, M.K.; Burman, C.; Balmer, P. Correlates of protection for meningococcal surface protein vaccines: Lessons from the past. Expert Rev. Vaccines 2021, 21, 739–751. [Google Scholar] [CrossRef]
- Findlow, J.; Balmer, P.; Borrow, R. A review of complement sources used in serum bactericidal assays for evaluating immune responses to meningococcal ACWY conjugate vaccines. Hum. Vaccines Immunother. 2019, 15, 2491–2500. [Google Scholar] [CrossRef]
- Rambach, G.; Wurzner, R.; Speth, C. Complement: An efficient sword of innate immunity. Contrib. Microbiol. 2008, 15, 78–100. [Google Scholar]
- Heesterbeek, D.A.C.; Angelier, M.L.; Harrison, R.A.; Rooijakkers, S.H.M. Complement and Bacterial Infections: From Molecular Mechanisms to Therapeutic Applications. J. Innate Immun. 2018, 10, 455–464. [Google Scholar] [CrossRef]
- Rosain, J.; Hong, E.; Fieschi, C.; Martins, P.V.; El Sissy, C.; Deghmane, A.E.; Ouachee, M.; Thomas, C.; Launay, D.; de Pontual, L.; et al. Strains Responsible for Invasive Meningococcal Disease in Patients with Terminal Complement Pathway Deficiencies. J. Infect. Dis. 2017, 215, 1331–1338. [Google Scholar] [CrossRef]
- Zipfel, P.F.; Wurzner, R.; Skerka, C. Complement evasion of pathogens: Common strategies are shared by diverse organisms. Mol. Immunol. 2007, 44, 3850–3857. [Google Scholar] [CrossRef]
- El Sissy, C.; Rosain, J.; Vieira-Martins, P.; Bordereau, P.; Gruber, A.; Devriese, M.; de Pontual, L.; Taha, M.K.; Fieschi, C.; Picard, C.; et al. Clinical and Genetic Spectrum of a Large Cohort with Total and Sub-total Complement Deficiencies. Front. Immunol. 2019, 10, 1936. [Google Scholar] [CrossRef]
- Aruta, M.G.; Carducci, M.; Micoli, F.; Necchi, F.; Rossi, O. Increasing the High Throughput of a Luminescence-Based Serum Bactericidal Assay (L-SBA). BioTech 2021, 10, 19. [Google Scholar] [CrossRef] [PubMed]
- Kleinschmidt, A.; Vadivelu, K.; Serino, L.; Neidig, N.; de Wergifosse, B. Endogenous complement human serum bactericidal assay (enc-hSBA) for vaccine effectiveness assessments against meningococcal serogroup B. npj Vaccines 2021, 6, 29. [Google Scholar] [CrossRef]
- Matthias, K.A.; Reveille, A.; Dhara, K.; Lyle, C.S.; Natuk, R.J.; Bonk, B.; Bash, M.C. Development and validation of a standardized human complement serum bactericidal activity assay to measure functional antibody responses to Neisseria gonorrhoeae. Vaccine 2025, 43, 126508. [Google Scholar] [CrossRef]
- Tzeng, Y.L.; Giuntini, S.; Berman, Z.; Sannigrahi, S.; Granoff, D.M.; Stephens, D.S. Neisseria meningitidis Urethritis Outbreak Isolates Express a Novel Factor H Binding Protein Variant That Is a Potential Target of Group B-Directed Meningococcal (MenB) Vaccines. Infect. Immun. 2020, 88, e00462-20. [Google Scholar] [CrossRef] [PubMed]
- Alexander, F.; Brunt, E.; Humphries, H.; Cavell, B.; Leung, S.; Allen, L.; Halkerston, R.; Lesne, E.; Penn, E.; Thomas, S.; et al. Generation of a Universal Human Complement Source by Large-Scale Depletion of IgG and IgM from Pooled Human Plasma. Methods Mol. Biol. 2022, 2414, 341–362. [Google Scholar] [CrossRef] [PubMed]
- Caron, F.; du Chatelet, I.P.; Leroy, J.P.; Ruckly, C.; Blanchard, M.; Bohic, N.; Massy, N.; Morer, I.; Floret, D.; Delbos, V.; et al. From tailor-made to ready-to-wear meningococcal B vaccines: Longitudinal study of a clonal meningococcal B outbreak. Lancet Infect. Dis. 2011, 11, 455–463. [Google Scholar] [CrossRef]
- Tettelin, H.; Saunders, N.J.; Heidelberg, J.; Jeffries, A.C.; Nelson, K.E.; Eisen, J.A.; Ketchum, K.A.; Hood, D.W.; Peden, J.F.; Dodson, R.J.; et al. Complete genome sequence of Neisseria meningitidis serogroup B strain MC58. Science 2000, 287, 1809–1815. [Google Scholar] [CrossRef]
- Guiddir, T.; Deghmane, A.E.; Giorgini, D.; Taha, M.K. Lipocalin 2 in cerebrospinal fluid as a marker of acute bacterial meningitis. BMC Infect. Dis. 2014, 14, 276. [Google Scholar] [CrossRef]
- Kellogg, D.S., Jr.; Peacock, W.L., Jr.; Deacon, W.E.; Brown, L.; Pirkle, D.I. Neisseria gonorrhoeae. I. Virulence Genetically Linked to Clonal Variation. J. Bacteriol. 1963, 85, 1274–1279. [Google Scholar] [CrossRef]
- Levy, M.; Aouiti Trabelsi, M.; Taha, M.K. Evidence for Multi-Organ Infection During Experimental Meningococcal Sepsis due to ST-11 Isolates in Human Transferrin-Transgenic Mice. Microorganisms 2020, 8, 1456. [Google Scholar] [CrossRef]
- Sevestre, J.; Hong, E.; Delbos, V.; Terrade, A.; Mallet, E.; Deghmane, A.E.; Lemee, L.; Taha, M.K.; Caron, F. Durability of immunogenicity and strain coverage of MenBvac, a meningococcal vaccine based on outer membrane vesicles: Lessons of the Normandy campaign. Vaccine 2017, 35, 4029–4033. [Google Scholar] [CrossRef]
- Borrow, R.; Carlone, G.M. Serogroup B and C serum bactericidal assays. Methods Mol. Med. 2001, 66, 289–304. [Google Scholar] [CrossRef]
- Borrow, R.; Aaberge, I.S.; Santos, G.F.; Eudey, T.L.; Oster, P.; Glennie, A.; Findlow, J.; Hoiby, E.A.; Rosenqvist, E.; Balmer, P.; et al. Interlaboratory standardization of the measurement of serum bactericidal activity by using human complement against meningococcal serogroup b, strain 44/76-SL, before and after vaccination with the Norwegian MenBvac outer membrane vesicle vaccine. Clin. Diagn. Lab. Immunol. 2005, 12, 970–976. [Google Scholar] [CrossRef]
- Borrow, R.; Balmer, P.; Miller, E. Meningococcal surrogates of protection—Serum bactericidal antibody activity. Vaccine 2005, 23, 2222–2227. [Google Scholar] [CrossRef]
- Mak, P.A.; Santos, G.F.; Masterman, K.A.; Janes, J.; Wacknov, B.; Vienken, K.; Giuliani, M.; Herman, A.E.; Cooke, M.; Mbow, M.L.; et al. Development of an automated, high-throughput bactericidal assay that measures cellular respiration as a survival readout for Neisseria meningitidis. Clin. Vaccine Immunol. 2011, 18, 1252–1260. [Google Scholar] [CrossRef]
- Romero-Steiner, S.; Spear, W.; Brown, N.; Holder, P.; Hennessy, T.; Gomez De Leon, P.; Carlone, G.M. Measurement of serum bactericidal activity specific for Haemophilus influenzae type b by using a chromogenic and fluorescent metabolic indicator. Clin. Diagn. Lab. Immunol. 2004, 11, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Lewis, L.A.; Ram, S. Meningococcal disease and the complement system. Virulence 2014, 5, 98–126. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fantoni, G.; Deghmane, A.-E.; Caron, F.; Taha, M.-K. A Bioluminescence-Based Serum Bactericidal Assay to Detect Bactericidal Antibodies Against Neisseria meningitidis in Human Sera. Microorganisms 2025, 13, 595. https://doi.org/10.3390/microorganisms13030595
Fantoni G, Deghmane A-E, Caron F, Taha M-K. A Bioluminescence-Based Serum Bactericidal Assay to Detect Bactericidal Antibodies Against Neisseria meningitidis in Human Sera. Microorganisms. 2025; 13(3):595. https://doi.org/10.3390/microorganisms13030595
Chicago/Turabian StyleFantoni, Giulia, Ala-Eddine Deghmane, François Caron, and Muhamed-Kheir Taha. 2025. "A Bioluminescence-Based Serum Bactericidal Assay to Detect Bactericidal Antibodies Against Neisseria meningitidis in Human Sera" Microorganisms 13, no. 3: 595. https://doi.org/10.3390/microorganisms13030595
APA StyleFantoni, G., Deghmane, A.-E., Caron, F., & Taha, M.-K. (2025). A Bioluminescence-Based Serum Bactericidal Assay to Detect Bactericidal Antibodies Against Neisseria meningitidis in Human Sera. Microorganisms, 13(3), 595. https://doi.org/10.3390/microorganisms13030595