
Itaconate: A Nexus Metabolite Fueling Leishmania Survival Through Lipid Metabolism Modulation

 ,
,  , and
, and
Abstract
1. Introduction
2. Material and Methods
2.1. Transcriptomic Data
2.2. Gene Expression Visualization of Acod1 and Il1b Gene Expression
2.3. Identification and Gene Set Enrichment Analysis of Genes Co-Expressed with Upregulated Acod1 Expression
2.4. Statistical Analysis
3. Results
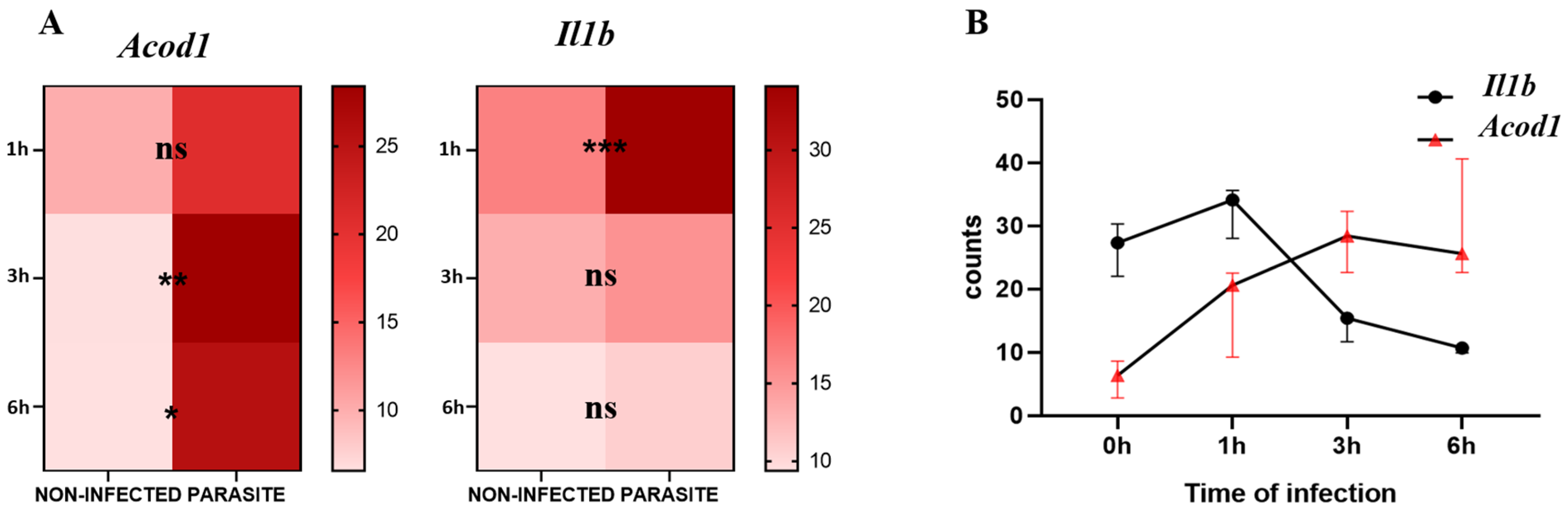
3.1. Kinetics of Il1b and Acod1 Gene Expression Levels in L. major-Infected Bone Marrow-Derived Macrophages (BMDMs)
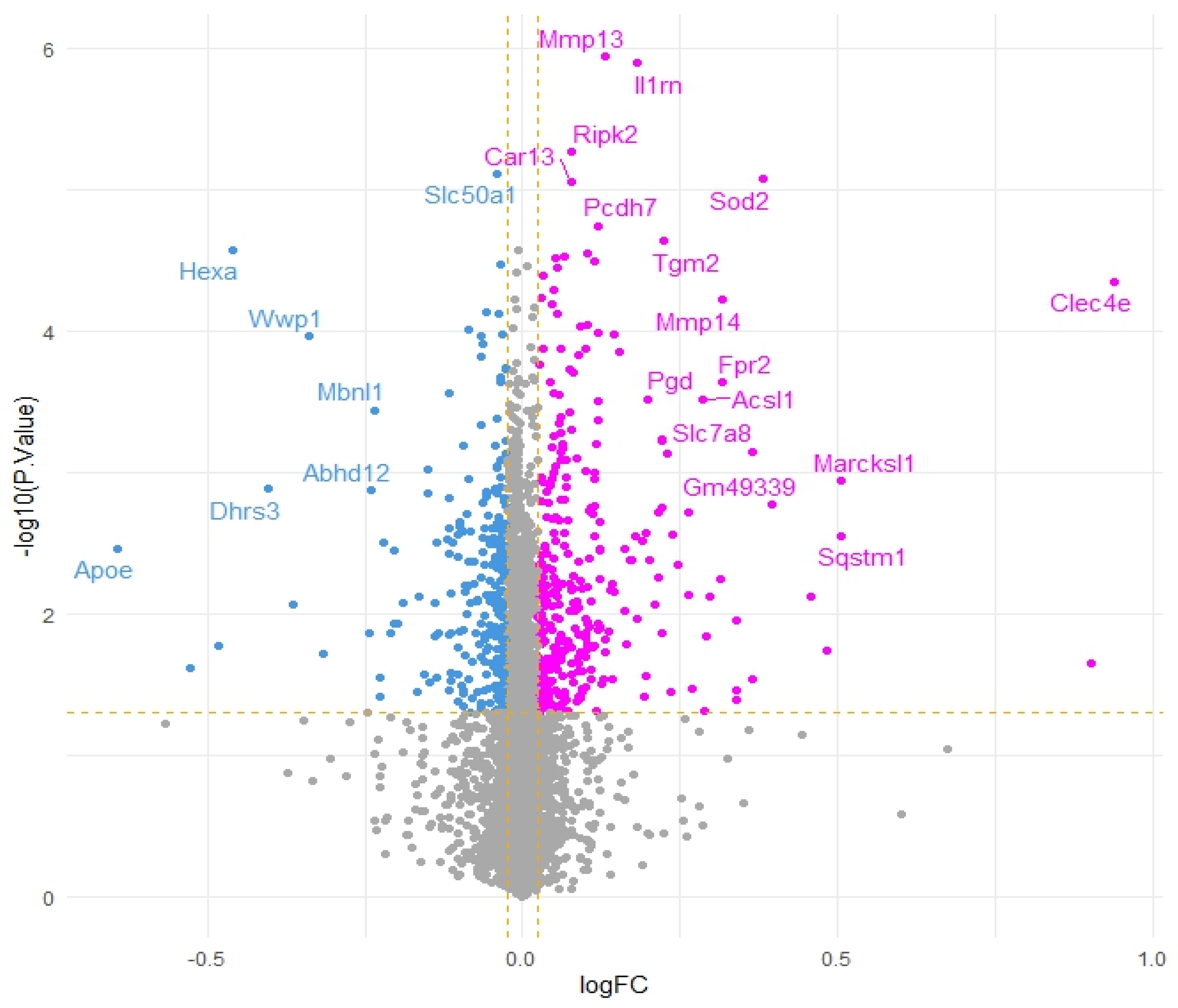
3.2. Transcriptomics
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kelly, B.; O’Neill, L.A. Metabolic Reprogramming in Macrophages and Dendritic Cells in Innate Immunity. Cell Res. 2015, 25, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, C.M.; Holowka, T.; Sun, J.; Blagih, J.; Amiel, E.; DeBerardinis, R.J.; Cross, J.R.; Jung, E.; Thompson, C.B.; Jones, R.G.; et al. Toll-like Receptor–Induced Changes in Glycolytic Metabolism Regulate Dendritic Cell Activation. Blood 2010, 115, 4742–4749. [Google Scholar] [CrossRef] [PubMed]
- Diskin, C.; Ryan, T.A.J.; O’Neill, L.A.J. Modification of Proteins by Metabolites in Immunity. Immunity 2021, 54, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.; O’Neill, L.A.J. Succinate: A Metabolic Signal in Inflammation. Trends Cell Biol. 2014, 24, 313–320. [Google Scholar] [CrossRef]
- Li, R.; Zhang, P.; Wang, Y.; Tao, K. Itaconate: A Metabolite Regulates Inflammation Response and Oxidative Stress. Oxidative Med. Cell. Longev. 2020, 2020, 5404780. [Google Scholar] [CrossRef]
- Wu, R.; Chen, F.; Wang, N.; Tang, D.; Kang, R. ACOD1 in Immunometabolism and Disease. Cell Mol. Immunol. 2020, 17, 822–833. [Google Scholar] [CrossRef] [PubMed]
- Jamal Uddin, M.; Joe, Y.; Kim, S.-K.; Oh Jeong, S.; Ryter, S.W.; Pae, H.-O.; Chung, H.T. IRG1 Induced by Heme Oxygenase-1/Carbon Monoxide Inhibits LPS-Mediated Sepsis and pro-Inflammatory Cytokine Production. Cell Mol. Immunol. 2016, 13, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Kuo, P.-C.; Weng, W.-T.; Scofield, B.A.; Furnas, D.; Paraiso, H.C.; Yu, I.-C.; Yen, J.-H. Immunoresponsive Gene 1 Modulates the Severity of Brain Injury in Cerebral Ischaemia. Brain Commun. 2021, 3, fcab187. [Google Scholar] [CrossRef]
- Wu, R.; Liu, J.; Tang, D.; Kang, R. The Dual Role of ACOD1 in Inflammation. J. Immunol. 2023, 211, 518–526. [Google Scholar] [CrossRef]
- Cordes, T.; Wallace, M.; Michelucci, A.; Divakaruni, A.S.; Sapcariu, S.C.; Sousa, C.; Koseki, H.; Cabrales, P.; Murphy, A.N.; Hiller, K.; et al. Immunoresponsive Gene 1 and Itaconate Inhibit Succinate Dehydrogenase to Modulate Intracellular Succinate Levels. J. Biol. Chem. 2016, 291, 14274–14284. [Google Scholar] [CrossRef] [PubMed]
- Lampropoulou, V.; Sergushichev, A.; Bambouskova, M.; Nair, S.; Vincent, E.E.; Loginicheva, E.; Cervantes-Barragan, L.; Ma, X.; Huang, S.C.-C.; Griss, T.; et al. Itaconate Links Inhibition of Succinate Dehydrogenase with Macrophage Metabolic Remodeling and Regulation of Inflammation. Cell Metab. 2016, 24, 158–166. [Google Scholar] [CrossRef]
- Daniels, B.P.; Kofman, S.B.; Smith, J.R.; Norris, G.T.; Snyder, A.G.; Kolb, J.P.; Gao, X.; Locasale, J.W.; Martinez, J.; Gale, M.; et al. The Nucleotide Sensor ZBP1 and Kinase RIPK3 Induce the Enzyme IRG1 to Promote an Antiviral Metabolic State in Neurons. Immunity 2019, 50, 64–76.e4. [Google Scholar] [CrossRef]
- Liao, S.-T.; Han, C.; Xu, D.-Q.; Fu, X.-W.; Wang, J.-S.; Kong, L.-Y. 4-Octyl Itaconate Inhibits Aerobic Glycolysis by Targeting GAPDH to Exert Anti-Inflammatory Effects. Nat. Commun. 2019, 10, 5091. [Google Scholar] [CrossRef]
- Sakai, A.; Kusumoto, A.; Kiso, Y.; Furuya, E. Itaconate Reduces Visceral Fat by Inhibiting Fructose 2,6-Bisphosphate Synthesis in Rat Liver. Nutrition 2004, 20, 997–1002. [Google Scholar] [CrossRef]
- Chen, F.; Elgaher, W.a.M.; Winterhoff, M.; Büssow, K.; Waqas, F.H.; Graner, E.; Pires-Afonso, Y.; Casares Perez, L.; de la Vega, L.; Sahini, N.; et al. Citraconate Inhibits ACOD1 (IRG1) Catalysis, Reduces Interferon Responses and Oxidative Stress, and Modulates Inflammation and Cell Metabolism. Nat. Metab. 2022, 4, 534–546. [Google Scholar] [CrossRef]
- Sohail, A.; Iqbal, A.A.; Sahini, N.; Chen, F.; Tantawy, M.; Waqas, S.F.H.; Winterhoff, M.; Ebensen, T.; Schultz, K.; Geffers, R.; et al. Itaconate and Derivatives Reduce Interferon Responses and Inflammation in Influenza A Virus Infection. PLoS Pathog. 2022, 18, e1010219. [Google Scholar] [CrossRef]
- He, Y.; Hara, H.; Núñez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Swain, A.; Bambouskova, M.; Kim, H.; Andhey, P.S.; Duncan, D.; Auclair, K.; Chubukov, V.; Simons, D.M.; Roddy, T.P.; Stewart, K.M.; et al. Comparative Evaluation of Itaconate and Its Derivatives Reveals Divergent Inflammasome and Type I Interferon Regulation in Macrophages. Nat. Metab. 2020, 2, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.L.; Ryan, D.G.; Prag, H.A.; Dikovskaya, D.; Menon, D.; Zaslona, Z.; Jedrychowski, M.P.; Costa, A.S.H.; Higgins, M.; Hams, E.; et al. Itaconate Is an Anti-Inflammatory Metabolite That Activates Nrf2 via Alkylation of KEAP1. Nature 2018, 556, 113–117. [Google Scholar] [CrossRef]
- Singh, O.P.; Sundar, S. Developments in Diagnosis of Visceral Leishmaniasis in the Elimination Era. J. Parasitol. Res. 2015, 2015, 239469. [Google Scholar] [CrossRef]
- Assouab, A.; Kihel, A.; Rouahi, M.; Larribau, M.; Karim, Z.; Akarid, K. Cutaneous Leishmaniasis and Iron Metabolism: Current Insights and Challenges. Front. Immunol. 2024, 15, 1488590. [Google Scholar] [CrossRef]
- Almeida, F.S.; Vanderley, S.E.R.; Comberlang, F.C.; de Andrade, A.G.; Cavalcante-Silva, L.H.A.; Silva, E.d.S.; Palmeira, P.H.d.S.; do Amaral, I.P.G.; Keesen, T.S.L. Leishmaniasis: Immune Cells Crosstalk in Macrophage Polarization. Trop. Med. Infect. Dis. 2023, 8, 276. [Google Scholar] [CrossRef]
- Ferreira, C.; Estaquier, J.; Silvestre, R. Immune-Metabolic Interactions between Leishmania and Macrophage Host. Curr. Opin. Microbiol. 2021, 63, 231–237. [Google Scholar] [CrossRef]
- Goldman-Pinkovich, A.; Kannan, S.; Nitzan-Koren, R.; Puri, M.; Pawar, H.; Bar-Avraham, Y.; McDonald, J.; Sur, A.; Zhang, W.-W.; Matlashewski, G.; et al. Sensing Host Arginine Is Essential for Leishmania Parasites’ Intracellular Development. mBio 2020, 11, e02023-20. [Google Scholar] [CrossRef]
- Darif, D.; Nait Irahal, I.; Hammi, I.; Kihel, A.; Kachmar, M.R.; Riyad, M.; Hmimid, F.; Akarid, K. Capparis Spinosa Inhibits Leishmania Major Growth through Nitric Oxide Production in Vitro and Arginase Inhibition in Silico. Exp. Parasitol. 2023, 245, 108452. [Google Scholar] [CrossRef]
- Akarid, K.; Arnoult, D.; Micic-Polianski, J.; Sif, J.; Estaquier, J.; Ameisen, J.C. Leishmania Major. -Mediated Prevention of Programmed Cell Death Induction in Infected Macrophages Is Associated with the Repression of Mitochondrial Release of Cytochrome c. J. Leukoc. Biol. 2004, 76, 95–103. [Google Scholar] [CrossRef]
- Assouab, A.; El Filaly, H.; Akarid, K. Inhibiting Human and Leishmania Arginases Using Cannabis Sativa as a Potential Therapy for Cutaneous Leishmaniasis: A Molecular Docking Study. TropicalMed 2022, 7, 400. [Google Scholar] [CrossRef]
- Rodrigues, V.; André, S.; Maksouri, H.; Mouttaki, T.; Chiheb, S.; Riyad, M.; Akarid, K.; Estaquier, J. Transcriptional Analysis of Human Skin Lesions Identifies Tryptophan-2,3-Deoxygenase as a Restriction Factor for Cutaneous Leishmania. Front. Cell Infect. Microbiol. 2019, 9, 338. [Google Scholar] [CrossRef]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for Functional Genomics Data Sets--Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef]
- Rabhi, I.; Rabhi, S.; Ben-Othman, R.; Rasche, A.; Consortium, S.; Daskalaki, A.; Trentin, B.; Piquemal, D.; Regnault, B.; Descoteaux, A.; et al. Transcriptomic Signature of Leishmania Infected Mice Macrophages: A Metabolic Point of View. PLoS Neglected Trop. Dis. 2012, 6, e1763. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) Hallmark Gene Set Collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Wickham, H. Manipulating Data. In ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009; pp. 157–175. ISBN 978-0-387-98141-3. [Google Scholar]
- Cline, M.S.; Smoot, M.; Cerami, E.; Kuchinsky, A.; Landys, N.; Workman, C.; Christmas, R.; Avila-Campilo, I.; Creech, M.; Gross, B.; et al. Integration of Biological Networks and Gene Expression Data Using Cytoscape. Nat. Protoc. 2007, 2, 2366–2382. [Google Scholar] [CrossRef]
- Wang, X.; Song, H.; Liang, J.; Jia, Y.; Zhang, Y. Abnormal Expression of HADH, an Enzyme of Fatty Acid Oxidation, Affects Tumor Development and Prognosis (Review). Mol. Med. Rep. 2022, 26, 355. [Google Scholar] [CrossRef]
- Go, G.-W.; Mani, A. Low-Density Lipoprotein Receptor (LDLR) Family Orchestrates Cholesterol Homeostasis. Yale J. Biol. Med. 2012, 85, 19–28. [Google Scholar]
- Lin, S.-Y.; Lin, S.-C. SRC Promotes Lipogenesis: Implications for Obesity and Breast Cancer. Mol. Cell. Oncol. 2021, 8, 1866975. [Google Scholar] [CrossRef]
- Lopez-Castejon, G.; Brough, D. Understanding the Mechanism of IL-1β Secretion. Cytokine Growth Factor. Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef]
- Kihel, A.; Hammi, I.; Darif, D.; Lemrani, M.; Riyad, M.; Guessous, F.; Akarid, K. The Different Faces of the NLRP3 Inflammasome in Cutaneous Leishmaniasis: A Review. Cytokine 2021, 147, 155248. [Google Scholar] [CrossRef]
- Lima-Junior, D.S.; Costa, D.L.; Carregaro, V.; Cunha, L.D.; Silva, A.L.N.; Mineo, T.W.P.; Gutierrez, F.R.S.; Bellio, M.; Bortoluci, K.R.; Flavell, R.A.; et al. Inflammasome-Derived IL-1β Production Induces Nitric Oxide–Mediated Resistance to Leishmania. Nat. Med. 2013, 19, 909–915. [Google Scholar] [CrossRef]
- Palacios, G.; Vega-García, E.; Valladares, B.; Pérez, J.A.; Dorta-Guerra, R.; Carmelo, E. Gene Expression Profiling of Classically Activated Macrophages in Leishmania Infantum Infection: Response to Metabolic Pre-Stimulus with Itaconic Acid. TropicalMed 2023, 8, 264. [Google Scholar] [CrossRef]
- Costa-da-Silva, A.C.; Nascimento, D.D.O.; Ferreira, J.R.M.; Guimarães-Pinto, K.; Freire-de-Lima, L.; Morrot, A.; Decote-Ricardo, D.; Filardy, A.A.; Freire-de-Lima, C.G. Immune Responses in Leishmaniasis: An Overview. TropicalMed 2022, 7, 54. [Google Scholar] [CrossRef]
- Tomiotto-Pellissier, F.; Bortoleti, B.T.D.S.; Assolini, J.P.; Gonçalves, M.D.; Carloto, A.C.M.; Miranda-Sapla, M.M.; Conchon-Costa, I.; Bordignon, J.; Pavanelli, W.R. Macrophage Polarization in Leishmaniasis: Broadening Horizons. Front. Immunol. 2018, 9, 2529. [Google Scholar] [CrossRef] [PubMed]
- Maksouri, H.; Dang, P.M.-C.; Rodrigues, V.; Estaquier, J.; Riyad, M.; Akarid, K. Moroccan Strains of Leishmania Major and Leishmania Tropica Differentially Impact on Nitric Oxide Production by Macrophages. Parasit. Vectors 2017, 10, 506. [Google Scholar] [CrossRef]
- Li, Z.; Zheng, W.; Kong, W.; Zeng, T. Itaconate: A Potent Macrophage Immunomodulator. Inflammation 2023, 46, 1177–1191. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, Y.; Deng, Q.; Hu, Y.; Dong, J.; Wang, W.; Wang, Y.; Li, C. Macrophage Polarization, Metabolic Reprogramming, and Inflammatory Effects in Ischemic Heart Disease. Front. Immunol. 2022, 13, 934040. [Google Scholar] [CrossRef]
- Mainali, R.; Buechler, N.; Otero, C.; Edwards, L.; Key, C.-C.; Furdui, C.; Quinn, M.A. Itaconate Stabilizes CPT1a to Enhance Lipid Utilization during Inflammation. eLife 2024, 12, RP92420. [Google Scholar] [CrossRef]
- Popa, F.I.; Perlini, S.; Teofoli, F.; Degani, D.; Funghini, S.; La Marca, G.; Rinaldo, P.; Vincenzi, M.; Antoniazzi, F.; Boner, A.; et al. 3-Hydroxyacyl-Coenzyme A Dehydrogenase Deficiency: Identification of a New Mutation Causing Hyperinsulinemic Hypoketotic Hypoglycemia, Altered Organic Acids and Acylcarnitines Concentrations. In JIMD Reports—Case and Research Reports, 2011/2; SSIEM, Ed.; JIMD Reports; Springer: Berlin/Heidelberg, Germany, 2011; Volume 2, pp. 71–77. ISBN 978-3-642-24757-6. [Google Scholar]
- Chandra, P.; He, L.; Zimmerman, M.; Yang, G.; Köster, S.; Ouimet, M.; Wang, H.; Moore, K.J.; Dartois, V.; Schilling, J.D.; et al. Inhibition of Fatty Acid Oxidation Promotes Macrophage Control of Mycobacterium Tuberculosis. mBio 2020, 11, e01139-20. [Google Scholar] [CrossRef]
- Wheeler, D.L.; Iida, M.; Dunn, E.F. The Role of Src in Solid Tumors. Oncologist 2009, 14, 667–678. [Google Scholar] [CrossRef]
- Schilke, R.M.; Blackburn, C.M.R.; Rao, S.; Krzywanski, D.M.; Finck, B.N.; Woolard, M.D. Macrophage-Associated Lipin-1 Promotes β-Oxidation in Response to Proresolving Stimuli. Immunohorizons 2020, 4, 659–669. [Google Scholar] [CrossRef]
- Chandran, S.; Schilke, R.M.; Blackburn, C.M.R.; Yurochko, A.; Mirza, R.; Scott, R.S.; Finck, B.N.; Woolard, M.D. Lipin-1 Contributes to IL-4 Mediated Macrophage Polarization. Front. Immunol. 2020, 11, 787. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Wang, H.; Han, C.; Cao, X. Src Promotes Anti-Inflammatory (M2) Macrophage Generation via the IL-4/STAT6 Pathway. Cytokine 2018, 111, 209–215. [Google Scholar] [CrossRef]
- Fortéa, J.O.Y.; De La Llave, E.; Regnault, B.; Coppée, J.-Y.; Milon, G.; Lang, T.; Prina, E. Transcriptional Signatures of BALB/c Mouse Macrophages Housing Multiplying Leishmania Amazonensis Amastigotes. BMC Genom. 2009, 10, 119. [Google Scholar] [CrossRef] [PubMed]
- Semini, G.; Paape, D.; Paterou, A.; Schroeder, J.; Barrios-Llerena, M.; Aebischer, T. Changes to Cholesterol Trafficking in Macrophages by Leishmania Parasites Infection. Microbiologyopen 2017, 6, e00469. [Google Scholar] [CrossRef]
- Andrade-Neto, V.V.; Manso, P.P.D.A.; Pereira, M.G.; De Cicco, N.N.T.; Atella, G.C.; Pelajo-Machado, M.; Menna-Barreto, R.F.S.; Torres-Santos, E.C. Host Cholesterol Influences the Activity of Sterol Biosynthesis Inhibitors in Leishmania Amazonensis. Mem. Inst. Oswaldo Cruz 2022, 117, e220407. [Google Scholar] [CrossRef]
- Parihar, S.P.; Hartley, M.-A.; Hurdayal, R.; Guler, R.; Brombacher, F. Topical Simvastatin as Host-Directed Therapy against Severity of Cutaneous Leishmaniasis in Mice. Sci. Rep. 2016, 6, 33458. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
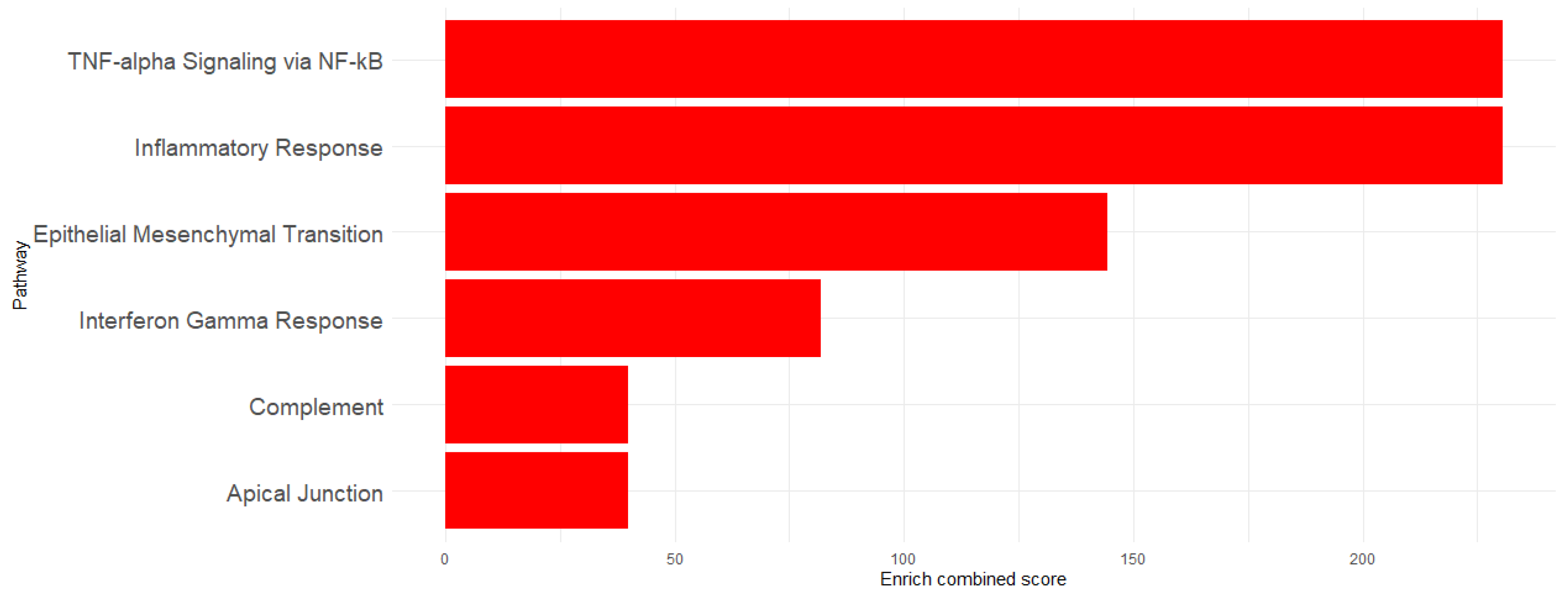
| Pathway | Overlap | p-Value | Adjusted p-Value | odd_R | Combined Score | Genes |
|---|---|---|---|---|---|---|
| TNF-alpha Signaling via NF-kB | 6/200 | 2.68 × 10−6 | 4.16 × 10−5 | 17.97999 | 230.6618 | Dram1/Ripk2/Traf1 /Cd69/Sod2/Ldlr |
| Inflammatory Response | 6/200 | 2.68 × 10−6 | 4.16 × 10−5 | 17.97999 | 230.6618 | Mmp14/Ripk2/Pcdh7/ Itga5/Cd69/Ldlr |
| Epithelial Mesenchymal Transition | 5/200 | 4.71 × 10−5 | 4.87 × 10−4 | 14.47985 | 144.2694 | Mmp14/Itgb5/Tpm4/ Itga5/Tgm2 |
| Interferon Gamma Response | 4/200 | 6.69 × 10−4 | 0.005185 | 11.20408 | 81.89774 | Ripk2/St8sia4/ Cd69/Sod2 |
| Apical Junction | 3/200 | 0.007415 | 0.038311 | 8.134038 | 39.89133 | Src/Traf1/Hadh |
| Complement | 3/200 | 0.007415 | 0.038311 | 8.134038 | 39.89133 | Mmp14/Mmp13/Src |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kihel, A.; El Filaly, H.; Darif, D.; Assouab, A.; Riyad, M.; Nait Irahal, I.; Akarid, K. Itaconate: A Nexus Metabolite Fueling Leishmania Survival Through Lipid Metabolism Modulation. Microorganisms 2025, 13, 531. https://doi.org/10.3390/microorganisms13030531
Kihel A, El Filaly H, Darif D, Assouab A, Riyad M, Nait Irahal I, Akarid K. Itaconate: A Nexus Metabolite Fueling Leishmania Survival Through Lipid Metabolism Modulation. Microorganisms. 2025; 13(3):531. https://doi.org/10.3390/microorganisms13030531
Chicago/Turabian StyleKihel, Ayyoub, Hajar El Filaly, Dounia Darif, Aicha Assouab, Myriam Riyad, Imane Nait Irahal, and Khadija Akarid. 2025. "Itaconate: A Nexus Metabolite Fueling Leishmania Survival Through Lipid Metabolism Modulation" Microorganisms 13, no. 3: 531. https://doi.org/10.3390/microorganisms13030531
APA StyleKihel, A., El Filaly, H., Darif, D., Assouab, A., Riyad, M., Nait Irahal, I., & Akarid, K. (2025). Itaconate: A Nexus Metabolite Fueling Leishmania Survival Through Lipid Metabolism Modulation. Microorganisms, 13(3), 531. https://doi.org/10.3390/microorganisms13030531