

Phenotypic and Molecular Characterization of Pyomelanin-Producing Acinetobacter baumannii ST2Pas;ST1816/ST195Oxf Causing the First European Nosocomial Outbreak
 and
and
Abstract
1. Introduction
2. Methods
2.1. Epidemiological Context
2.2. Phenotypic and Molecular Characterization of Strains
2.3. Software and Tools Used for Genomic Analysis
3. Results
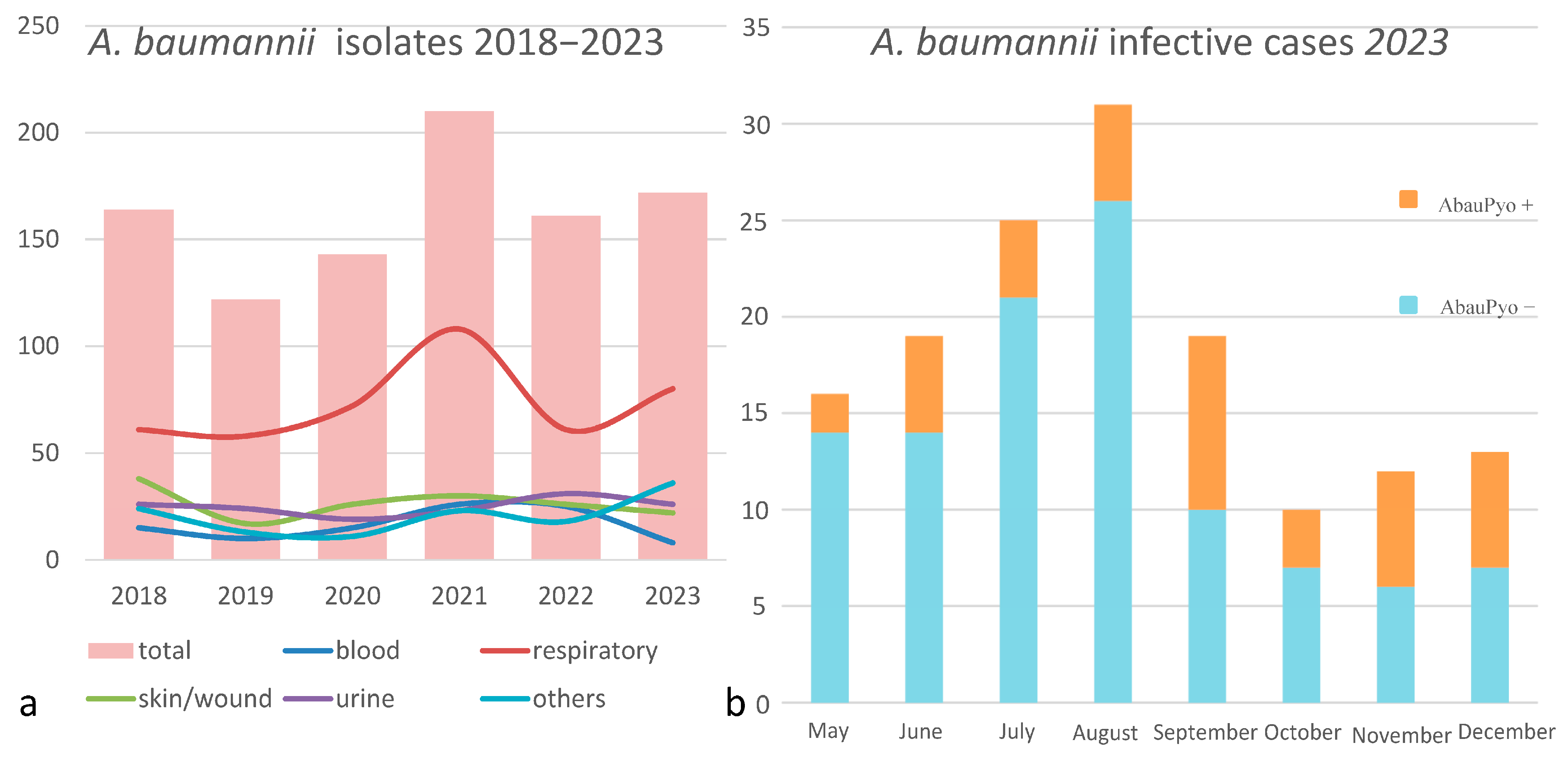
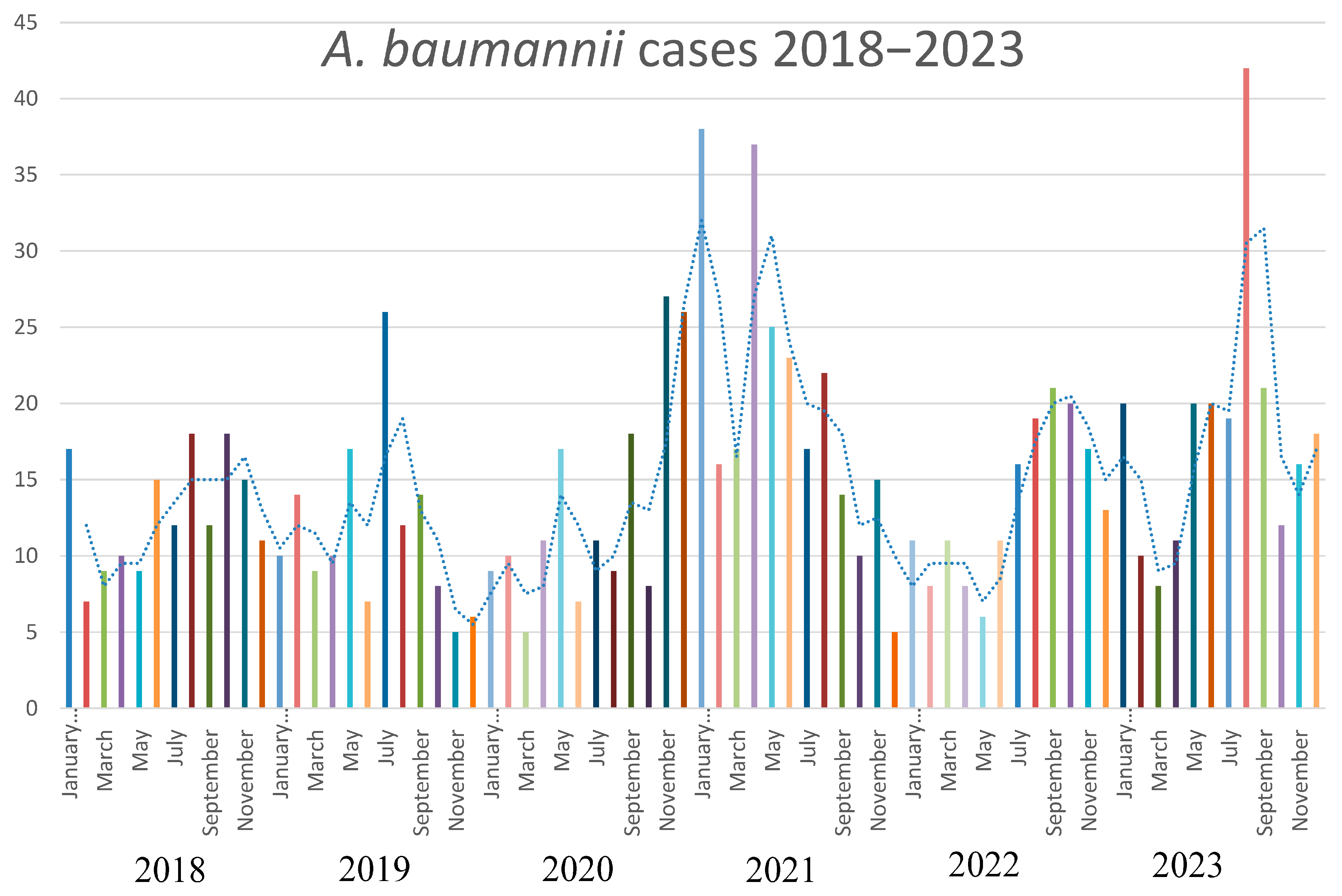
3.1. Phenotypic and Epidemiologic Analysis
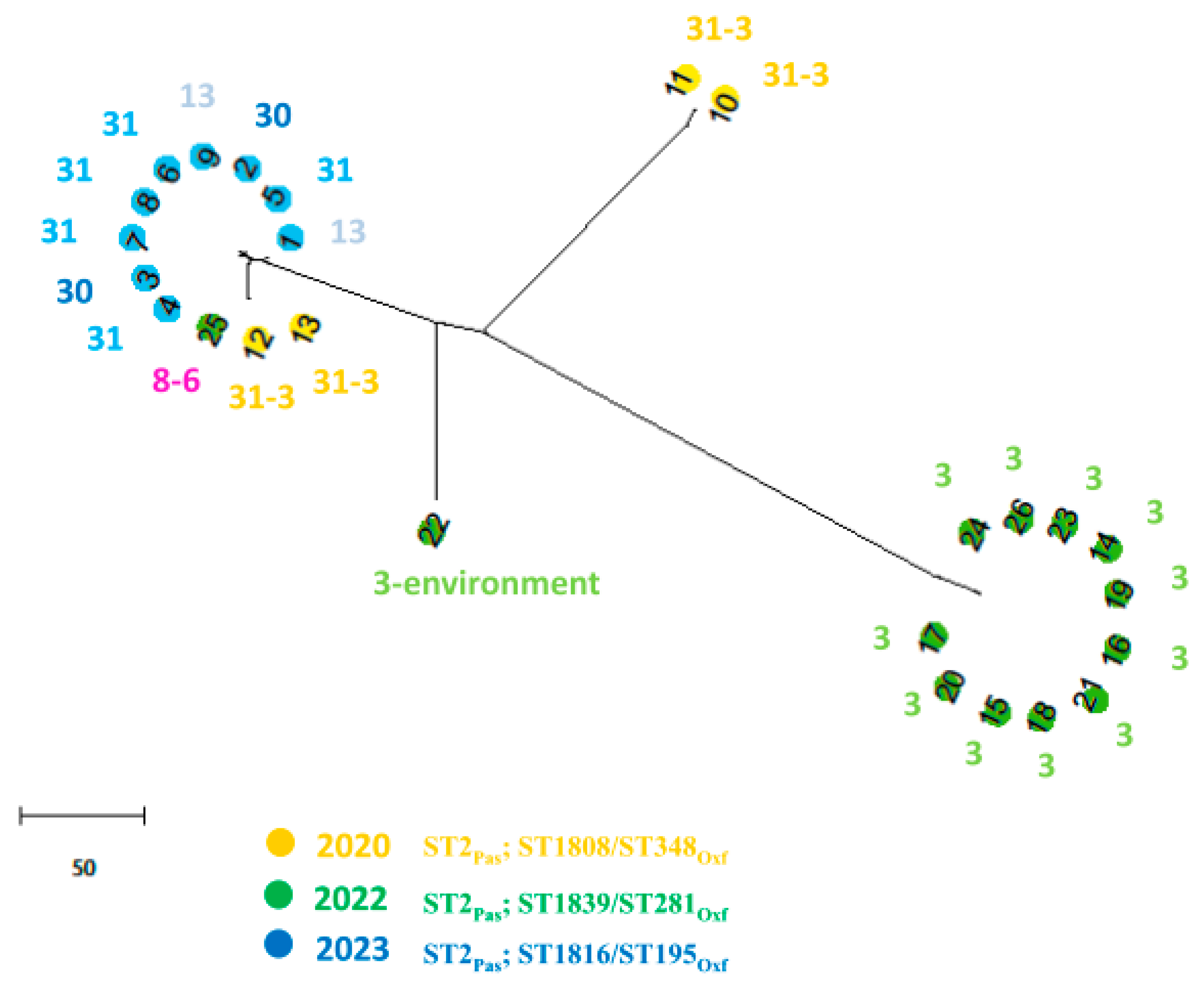
3.2. Molecular Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giammanco, A.; Calà, C.; Fasciana, T.; Dowzicky, M.J. Global Assessment of the Activity of Tigecycline against Multidrug-Resistant Gram-Negative Pathogens between 2004 and 2014 as Part of the Tigecycline Evaluation and Surveillance Trial. mSphere 2017, 2, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Rolain, J.M.; Diene, S.M.; Kempf, M.; Gimenez, G.; Robert, C.; Raoult, D. Real-time sequencing to decipher the molecular mechanism of resistance of a clinical pan-drug-resistant Acinetobacter baumannii isolate from Marseille, France. Antimicrob. Agents Chemother. 2013, 57, 592–596. [Google Scholar] [CrossRef]
- Agodi, A.; Barchitta, M.; Auxilia, F.; Brusaferro, S.; D’Errico, M.M.; Montagna, M.T.; Pasquarella, C.; Tardivo, S.; Arrigoni, C.; Fabiani, L.; et al. Epidemiology of intensive care unit-acquired sepsis in Italy: Results of the SPIN-UTI network. Ann. Ig. 2018, 30, 15–21. [Google Scholar]
- Zarrilli, R.; Bagattini, M.; Migliaccio, A.; Esposito, E.P.; Triassi, M. Molecular epidemiology of carbapenem-resistant Acinetobacter baumannii in Italy. Ann. Ig. 2021, 33, 401–409. [Google Scholar]
- Garnacho-Montero, J.; Ortiz-Leyba, C.; Fernández-Hinojosa, E.; Aldabó-Pallás, T.; Cayuela, A.; Marquez-Vácaro, J.A.; Garcia-Curiel, A.; Jiménez-Jiménez, F.J. Acinetobacter baumannii ventilator-associated pneumonia: Epidemiological and clinical findings. Intensive Care Med. 2005, 31, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Chopra, T.; Marchaim, D.; Awali, R.A.; Krishna, A.; Johnson, P.; Tansek, R.; Chaudary, K.; Lephart, P.; Slim, J.; Hothi, J.; et al. Epidemiology of bloodstream infections caused by Acinetobacter baumannii and impact of drug resistance to both carbapenems and ampicillin-sulbactam on clinical outcomes. Antimicrob. Agents Chemother. 2013, 57, 6270–6275. [Google Scholar] [CrossRef]
- World Health Statistics 2017: Monitoring Health for the SDGs, Sustainable Development Goals; World Health Organization: Geneva, Switzerland, 2017; ISBN 978-92-4-156548-6.
- Ibrahim, S.; Al-Saryi, N.; Al-Kadmy, I.M.S.; Aziz, S.N. Multidrug-resistant Acinetobacter baumannii as an emerging concern in hospitals. Mol. Biol. Rep. 2021, 48, 6987–6998. [Google Scholar] [CrossRef]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Microbiol. 2018, 16, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options. Front. Cell Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef]
- Yabuuchi, E.; Ohyama, A. Characterization of “pyomelanin”-producing strains of Pseudomonas aeruginosa. Int. J. Syst. Bacteriol. 1972, 22, 53–64. [Google Scholar] [CrossRef]
- Fonseca, É.; Freitas, F.; Caldart, R.; Morgado, S.; Vicente, A.C. Pyomelanin biosynthetic pathway in pigment-producer strains from the pandemic Acinetobacter baumannii IC-5. Mem. Inst. Oswaldo Cruz. 2020, 115, e200371. [Google Scholar] [CrossRef]
- Zhao, F.; Liu, H.; Yao, Y.; Zhang, L.; Zhou, Z.; Leptihn, S.; Yu, Y.; Hua, X.; Fu, Y. Description of a Rare Pyomelanin-Producing Carbapenem-Resistant Acinetobacter baumannii Strain Coharboring Chromosomal OXA-23 and NDM-1. Microbiol. Spectr. 2022, 10, e0214422. [Google Scholar] [CrossRef] [PubMed]
- Coelho-Souza, T.; Martins, N.; Maia, F.; Frases, S.; Bonelli, R.R.; Riley, L.W.; Moreira, B.M. Pyomelanin production: A rare phenotype in Acinetobacter baumannii. J. Med. Microbiol. 2014, 63, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Chatfield, C.H.; Cianciotto, N.P. The secreted pyomelanin pigment of Legionella pneumophila confers ferric reductase activity. Infect. Immun. 2007, 75, 4062–4070. [Google Scholar] [CrossRef]
- Valeru, S.P.; Rompikuntal, P.K.; Ishikawa, T.; Vaitkevicius, K.; Sjöling, A.; Dolganov, N.; Zhu, J.; Schoolnik, G.; Wai, S.N. Role of melanin pigment in expression of Vibrio cholerae virulence factors. Infect. Immun. 2009, 77, 935–942. [Google Scholar] [CrossRef]
- Arias-Barrau, E.; Olivera, E.R.; Luengo, J.M.; Fernández, C.; Galán, B.; García, J.L.; Díaz, E.; Miñambres, B. The homogentisate pathway a central catabolic pathway involved in the degradation of L-phenylalanine, L-tyrosine, and 3-hydroxyphenylacetate in Pseudomonas putida. J. Bacteriol. 2004, 186, 5062–5077. [Google Scholar] [CrossRef]
- Ranjan, V.K.; Saha, T.; Mukherjee, S.; Chakraborty, R. Draft genome sequence of a novel bacterium, Pseudomonas sp strain MR 02, capable of pyomelanin production, isolated from the Mahananda River at Siliguri, West Bengal, India. Genome Announc. 2018, 6, e01443-17. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.C.; Newman, D.K. A putative ABC transporter, hatABCDE, is among molecular determinants of pyomelanin production in Pseudomonas aeruginosa. J. Bacteriol. 2010, 192, 5962–5971. [Google Scholar] [CrossRef]
- Loi, J.; Yi, T.S.; Pinsa, A.; Mulyana, S.; Singaretnam, L.G.; Riwanto, M.; Chang, J. Pyomelanin production from a marine isolate of Acinetobacter spp. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 2250–2259. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; UGENE team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Johansson, M.H.K.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Machado, M.P.; Silva, D.N.; Rossi, M.; Moran-Gilad, J.; Santos, S.; Ramirez, M.; Carriço, J.A. chewBBACA: A complete suite for gene-by-gene schema creation and strain identification. Microb. Genom. 2018, 4, e000166. [Google Scholar] [CrossRef]
- Knijn, A.; Michelacci., V.; Orsini, M.; Morabito, S. Advanced Research Infrastructure for Experimentation in genomicS (ARIES): A lustrum of Galaxy experience. bioRxiv 2020. [Google Scholar] [CrossRef]
- Giordano, C.; Barnini, S. Glycine restores the sensitivity to antibiotics in multidrug-resistant bacteria. Microbiol. Spectr. 2024, 12, e0016424. [Google Scholar] [CrossRef] [PubMed]
- Falcone, M.; Tiseo, G.; Carbonara, S.; Marino, A.; Di Caprio, G.; Carretta, A.; Mularoni, A.; Mariani, M.F.; Maraolo, A.E.; Scotto, R.; et al. Mortality attributable to bloodstream infections caused by different carbapenem-resistant Gram negative bacilli: Results from a nationwide study in Italy (ALARICO Network). Clin. Infect. Dis. 2023, 76, 2059–2069. [Google Scholar] [CrossRef]
- Zarrilli, R.; Pournaras, S.; Giannouli, M.; Tsakris, A. Global evolution of multidrug-resistant Acinetobacter baumannii clonal lineages. Int. J. Antimicrob. Agents 2013, 41, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Tiseo, G.; Giordano, C.; Leonildi, A.; Riccardi, N.; Galfo, V.; Limongi, F.; Nicastro, M.; Barnini, S.; Falcone, M. Salvage therapy with sulbactam/durlobactam against cefiderocol-resistant Acinetobacter baumannii in a critically ill burn patient: Clinical challenges and molecular characterization. JAC Antimicrob. Resist. 2023, 5, dlad078. [Google Scholar] [CrossRef]
- Falcone, M.; Tiseo, G.; Nicastro, M.; Leonildi, A.; Vecchione, A.; Casella, C.; Forfori, F.; Malacarne, P.; Guarracino, F.; Barnini, S.; et al. Cefiderocol as Rescue Therapy for Acinetobacter baumannii and Other Carbapenem-resistant Gram-negative Infections in Intensive Care Unit Patients. Clin. Infect. Dis. 2021, 72, 2021–2024. [Google Scholar] [CrossRef]
- Roca, I.; Espinal, P.; Vila-Farrés, X.; Vila, J. The Acinetobacter baumannii oxymoron: Commensal hospital dweller turned pan-drug-resistant menace. Front. Microbiol. 2012, 3, 148. [Google Scholar] [CrossRef] [PubMed]
- Nosanchuk, J.D.; Casadevall, A. The contribution of melanin to microbial pathogenesis. Cell Microbiol. 2003, 5, 203–223. [Google Scholar] [CrossRef] [PubMed]
- Imperi, F.; Antunes, L.C.; Blom, J.; Villa, L.; Iacono, M.; Visca, P.; Carattoli, A. The genomics of Acinetobacter baumannii: Insights into genome plasticity, antimicrobial resistance and pathogenicity. IUBMB Life 2011, 63, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Hèritier, C.; Poirel, L.; Nordmann, P. Cephalosporinase overexpression resulting from insertion of ISAba1 in Acinetobacter baumannii. Clin. Microbiol. Infect. 2006, 12, 123–130. [Google Scholar] [CrossRef]
- Mussi, M.A.; Limansky, A.S.; Viale, A.M. Acquisition of resistance to carbapenems in multidrug-resistant clinical strains of Acinetobacter baumannii: Natural insertional inactivation of a gene encoding a member of a novel family of b-barrel outer membrane proteins. Antimicrob. Agents Chemother. 2005, 49, 1432–1440. [Google Scholar] [CrossRef] [PubMed]
- Hornsey, M.; Phee, L.; Wareham, D.W. A novel variant, NDM-5, of the New Delhi metallo-b-lactamase in a multidrug-resistant Escherichia coli ST648 isolate recovered from a patient in the United Kingdom. Antimicrob. Agents Chemother. 2011, 55, 5952–5954. [Google Scholar] [CrossRef]
- Lopes, B.S.; Amyes, S.G.B. Role of ISAba1 and ISAba125 in governing the expression of blaADC in clinically relevant Acinetobacter baumannii strains resistant to cephalosporins. J. Med. Microbiol. 2012, 61, 1103–1108. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| HYDROGEN PEROXIDE % | SALINE | ||||
|---|---|---|---|---|---|
| 0.75% | 0.37% | 0.16% | 0.08% | CTRL | |
| MP | 34 | 24 | 10 | 0 | 0 |
| MNP | 40 | 32 | 30 | 24 | 0 |
| FP | 36 | 25 | 10 | 0 | 0 |
| FNP | 36 | 32 | 30 | 24 | 0 |
| Strain | Isolation Date (Source) | Breakpoints | Genome Size (base) | Sequence Type | Resistome | Mobilome | Virulence | Accession Number | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amikacin (mg/L) | Meropenem (mg/L) | Colistin (mg/L) | Cefiderocol (mm) | MLST | Aminoglycoside Resistance | B-Lactam Resistance | MGEs | Colicin V | RecA | Pili Type | Porin | Pyomelanin | ||||
| 1 | 28/05/23 (sputum) | >16 | 64 | ≤0.5 | 21 mm | 3,987,231 | ST2Pas; ST1816/ST195Oxf | aph(6)-Id, aph(3′)-Ia, aph(3″)-Ib, armA | OXA -23, OXA-66, TEM-1D, and ADC-25 | ISAba24, ISAba26, IS17, is26, ISAba125 | ✓ | ✓ | I/II/IV | OprB, Aquaporin Z | ✓ | SAMN41030183 |
| 2 | 14/06/23 (urine) | >16 | 32 | ≤1 | 22 mm | 3,999,467 | ST2Pas; ST1816/ST195Oxf | aph(6)-Id, aph(3′)-Ia, aph(3″)-Ib, armA | OXA -23, OXA-66, TEM-1D, and ADC-25 | ISAba24, ISVsa3, ISAba26, IS17, ISAba25 | ✓ | ✓ | I/II/IV | OprB, Aquaporin Z | ✓ | SAMN41030184 |
| 3 | 20/06/23 (CVC swab) | >16 | 32 | ≤1 | 22 mm | 3,995,867 | ST2Pas; ST1816/ST195Oxf | aph(6)-Id, aph(3′)-Ia, aph(3″)-Ib, armA | OXA -23, OXA-66, TEM-1D, and ADC-25 | ISAba24, ISVsa3, ISAba26, IS17, IS26, ISAba125 | ✓ | ✓ | I/II/IV | OprB, Aquaporin Z | ✓ | SAMN41030185 |
| 4 | 23/06/23 (BAS) | >16 | 64 | 1 | 22 mm | 3,998,140 | ST2Pas; ST1816/ST195Oxf | aph(6)-Id, aph(3′)-Ia, aph(3″)-Ib, armA | OXA -23, OXA-66, TEM-1D, and ADC-25 | ISAba24, ISVsa3, ISAba26, IS17, ISAba125 | ✓ | ✓ | I/II/IV | OprB, Aquaporin Z | ✓ | SAMN41030186 |
| 5 | 27/06/23 (BAS) | >16 | 64 | ≤0.5 | 21 mm | 3,990,297 | ST2Pas; ST1816/ST195Oxf | aph(6)-Id, aph(3′)-Ia, aph(3″)-Ib, armA | OXA -23, OXA-66, TEM-1D, and ADC-25 | ISAba24, ISVsa3, ISAba26, IS17, IS26, ISAba25 | ✓ | ✓ | I/II/IV | OprB, Aquaporin Z | ✓ | SAMN41030187 |
| 6 | 27/06/23 (BAL) | >16 | >64 | ≤0.5 | 22 mm | 3,988,632 | ST2Pas; ST1816/ST195Oxf | aph(6)-Id, aph(3′)-Ia, aph(3″)-Ib, armA | OXA -23, OXA-66, TEM-1D, and ADC-25 | ISAba24, ISVsa3, ISAba26, IS17, IS26, ISAba125 | ✓ | ✓ | I/II/IV | OprB, Aquaporin Z | ✓ | SAMN41030188 |
| 7 | 12/07/23 (BAL) | >16 | >64 | ≤0.5 | 22 mm | 3,989,774 | ST2Pas; ST1816/ST195Oxf | aph(6)-Id, aph(3′)-Ia, aph(3″)-Ib, armA | OXA -23, OXA-66, TEM-1D, and ADC-25 | ISAba24, ISVsa3, ISAba26, IS17, ISAba125 | ✓ | ✓ | I/II/IV | OprB, Aquaporin Z | ✓ | SAMN41030189 |
| 8 | 13/07/23 (BAS) | >16 | 64 | ≤0.5 | 22 mm | 3,988,236 | ST2Pas; ST1816/ST195Oxpiraginef | aph(6)-Id, aph(3′)-Ia, aph(3″)-Ib, armA | OXA -23, OXA-66, TEM-1D, and ADC-25 | ISAba24, ISVsa3, ISAba26, IS17, ISAba125 | ✓ | ✓ | I/II/IV | OprB, Aquaporin Z | ✓ | SAMN41030190 |
| 9 | 13/07/23 (BAL) | >16 | 64 | 0.5 | 20 mm | 3,990,333 | ST2Pas; ST1816/ST195Oxf | aph(6)-Id, aph(3′)-Ia, aph(3″)-Ib, armA | OXA -23, OXA-66, TEM-1D, and ADC-25 | ISAba24, ISVsa3, ISAba26, IS17, IS26, ISAba25 | ✓ | ✓ | I/II/IV | OprB, Aquaporin Z | ✓ | SAMN41030191 |
| 10 | 24/04/20 (blood) | >16 | >64 | 1 | 6 mm | 3,846,537 | ST2Pas; ST1808/ST348Oxf | aph(6)-Id, aph(3″)-Ib, armA | OXA -72, OXA-66, TEM-1D, and ADC-25 | ISVsa3, ISAba26, ISAba13, ISEc28, ISEc29, ISAba1, IS17, IS26, ISAba36 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030730 |
| 11 | 24/04/20 (blood) | >16 | >64 | 1 | 20 mm | 3,790,446 | ST2Pas; ST1808/ST348Oxf | aph(6)-Id, aph(3″)-Ib, armA | OXA -72, OXA-66, TEM-1D, and ADC-25 | ISVsa3, ISAba26, ISAba13, ISEc29, IS17, IS26, ISAba36 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030731 |
| 12 | 24/04/20 (skin) | >16 | 64 | ≤0.5 | 6 mm | 3,999,068 | ST2Pas; ST1808/ST348Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, | OXA -23, OXA-66, TEM-1D, and ADC-25 | ISAba24, ISVsa3, ISAba125, ISAba26, IS17, IS26 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030732 |
| 13 | 24/04/20 (skin) | >16 | 64 | ≤0.5 | 20 mm | 4,032,758 | ST2Pas; ST1808/ST348Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, | OXA -23, OXA-66, TEM-1D, and ADC-25 | ISAba24, ISVsa3, ISAba125, ISAba26, IS17, ISAba36, IS26 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030733 |
| 14 | 03/11/22 (rectal swab) | >16 | 32 | ≤0.5 | <17 mm | 3,975,455 | ST2Pas; ST1839/ST281Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, aadA1, aac(6′)-Ib-cr | OXA -225, OXA-66, ADC-25 | ISAba24, ISVsa3, ISAba125 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030734 |
| 15 | 28/10/22 (skin) | >16 | 32 | ≤0.5 | <17 mm | 3,976,418 | ST2Pas; ST1839/ST281Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, aadA1, mazzettiaac(6′)-Ib-bonuccellicr | OXA -26, OXA-66, ADC-25 | ISAba24, ISVsa3, ISAba125, IS26 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030735 |
| 16 | 07/11/22 (blood) | >16 | 32 | ≤0.5 | <17 mm | 3,974,827 | ST2Pas; ST1839/ST281Oxf | aph(3′)-Ia, petracciniaph(3″)-Ib, ardi garbomA, aadA1, aac(6′)-Ib-cr | OXA -225, OXA-66, OXA-23, ADC-25 | ISAba24, ISVsa3, ISAba125 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030736 |
| 17 | 27/10/22 (skin) | >16 | 16 | ≤0.5 | 12 mm | 3,971,629 | ST2Pas; ST1839/ST281Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, aadA1, aac(6′)-Ib-cr | OXA-66, OXA-23, ADC-25 | ISAba24, ISVsa3, ISAba125, IS26 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030737 |
| 18 | 01/11/22 (CVC) | >16 | 32 | ≤0.5 | 11 mm | 3,972,425 | ST2Pas; ST1839/ST281Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, aadA1, aac(6′)-Ib-cr | OXA-66, OXA-23, ADC-25 | ISAba24, ISVsa3, ISAba125 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SUB12614636 |
| 19 | 07/11/22 (blood) | >16 | 16 | ≤0.5 | <17 mm | 3,974,920 | ST2Pas; ST1839/ST281Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, aadA1, aac(6′)-Ib-cr | OXA-66, OXA-23, ADC-25 | ISAba24, ISVsa3, ISAba125, IS26 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030738 |
| 20 | 10/10/2022 (skin) | >16 | 16 | ≤1 | <17 mm | 3,971,101 | ST2Pas; ST1839/ST281Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, aadA1, aac(6′)-Ib-cr | OXA -225, OXA-66, ADC-25 | ISAba24, ISVsa3, ISAba125, IS26 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030739 |
| 21 | 06/10/2022 (blood) | >16 | 16 | ≤1 | <17 mm | 4,121,556 | ST2Pas; ST1839/ST281Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, aadA1, aac(6′)-Ib-cr | OXA -225, OXA-66, ADC-25, blaCMY-33, blaOXA-10 | IS26, ISEc9, ISAba24, Tn6196, ISVsa3, IS5075, ISAba125 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030740 |
| 22 | Environmental swab | >16 | 32 | ≤0.5 | 20 mm | 3,942,955 | ST2Pas; ST1839/ST281Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, aadA1, aac(6′)-Ib-cr | OXA -23, OXA-66, ADC-25 | Tn6207, ISEc29, ISAba26,IS26 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030741 |
| 23 | 22/08/22 (BAL) | >16 | 16 | ≤0.5 | 18 mm | 3,975,018 | ST2Pas; ST1839/ST281Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, aadA1, aac(6′)-Ib-cr | OXA -225, OXA-66, ADC-25 | ISAba24, ISVsa3, ISAba125, IS26 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030742 |
| 24 | 17/09/22 (blood) | >16 | 16 | ≤0.5 | 9 mm | 3,977,422 | ST2Pas; ST1839/ST281Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, aadA1, aac(6′)-Ib-cr | OXA -225, OXA-66, ADC-25 | ISAba24, ISVsa3, ISAba125 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030743 |
| 25 | 14/06/22 (wound) | >16 | 32 | ≤0.5 | 9 mm | 3,935,147 | ST2Pas; ST1839/ST281Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, aadA1, aac(6′)-Ib-cr | OXA -23, OXA-66, ADC-25, TEM-1D | ISVsa3, ISEc29, ISAba125, ISAba26, IS17, IS26 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030744 |
| 26 | 15/09/22 (blood) | >16 | 16 | ≤0.5 | 14 mm | 3,972,320 | ST2Pas; ST1839/ST281Oxf | aph(3′)-Ia, aph(3″)-Ib, armA, aadA1, aac(6′)-Ib-cr | OXA -225, OXA-66, | ISAba24, ISVsa3, ISAba125 | ✓ | ✓ | I/IV | OmpA, OprD, OprB, Aquaporin Z | - | SAMN41030745 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leonildi, A.; Rosellini, A.; Gemignani, G.; Tiseo, G.; Falcone, M.; Giordano, C.; Barnini, S. Phenotypic and Molecular Characterization of Pyomelanin-Producing Acinetobacter baumannii ST2Pas;ST1816/ST195Oxf Causing the First European Nosocomial Outbreak. Microorganisms 2025, 13, 493. https://doi.org/10.3390/microorganisms13030493
Leonildi A, Rosellini A, Gemignani G, Tiseo G, Falcone M, Giordano C, Barnini S. Phenotypic and Molecular Characterization of Pyomelanin-Producing Acinetobacter baumannii ST2Pas;ST1816/ST195Oxf Causing the First European Nosocomial Outbreak. Microorganisms. 2025; 13(3):493. https://doi.org/10.3390/microorganisms13030493
Chicago/Turabian StyleLeonildi, Alessandro, Alfredo Rosellini, Giulia Gemignani, Giusy Tiseo, Marco Falcone, Cesira Giordano, and Simona Barnini. 2025. "Phenotypic and Molecular Characterization of Pyomelanin-Producing Acinetobacter baumannii ST2Pas;ST1816/ST195Oxf Causing the First European Nosocomial Outbreak" Microorganisms 13, no. 3: 493. https://doi.org/10.3390/microorganisms13030493
APA StyleLeonildi, A., Rosellini, A., Gemignani, G., Tiseo, G., Falcone, M., Giordano, C., & Barnini, S. (2025). Phenotypic and Molecular Characterization of Pyomelanin-Producing Acinetobacter baumannii ST2Pas;ST1816/ST195Oxf Causing the First European Nosocomial Outbreak. Microorganisms, 13(3), 493. https://doi.org/10.3390/microorganisms13030493