
Natural Phenolics Disrupt Microbial Communication by Inhibiting Quorum Sensing



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Chemicals
2.2.1. Isolation of Plant Chemicals
2.2.2. Purchased Chemicals
2.3. Bacterial Strains
2.4. Determination of Minimal Inhibitory Concentration (MIC)
2.5. Inhibition of AI-2-Dependent QS in V. harveyi MM30
2.6. Inhibition of AI-2 Independent QS in V. harveyi MM30
2.7. Inhibition of AI-2 Production in MRSA 7112
2.8. Statistical Analysis
3. Results and Discussion
3.1. Determination of MIC
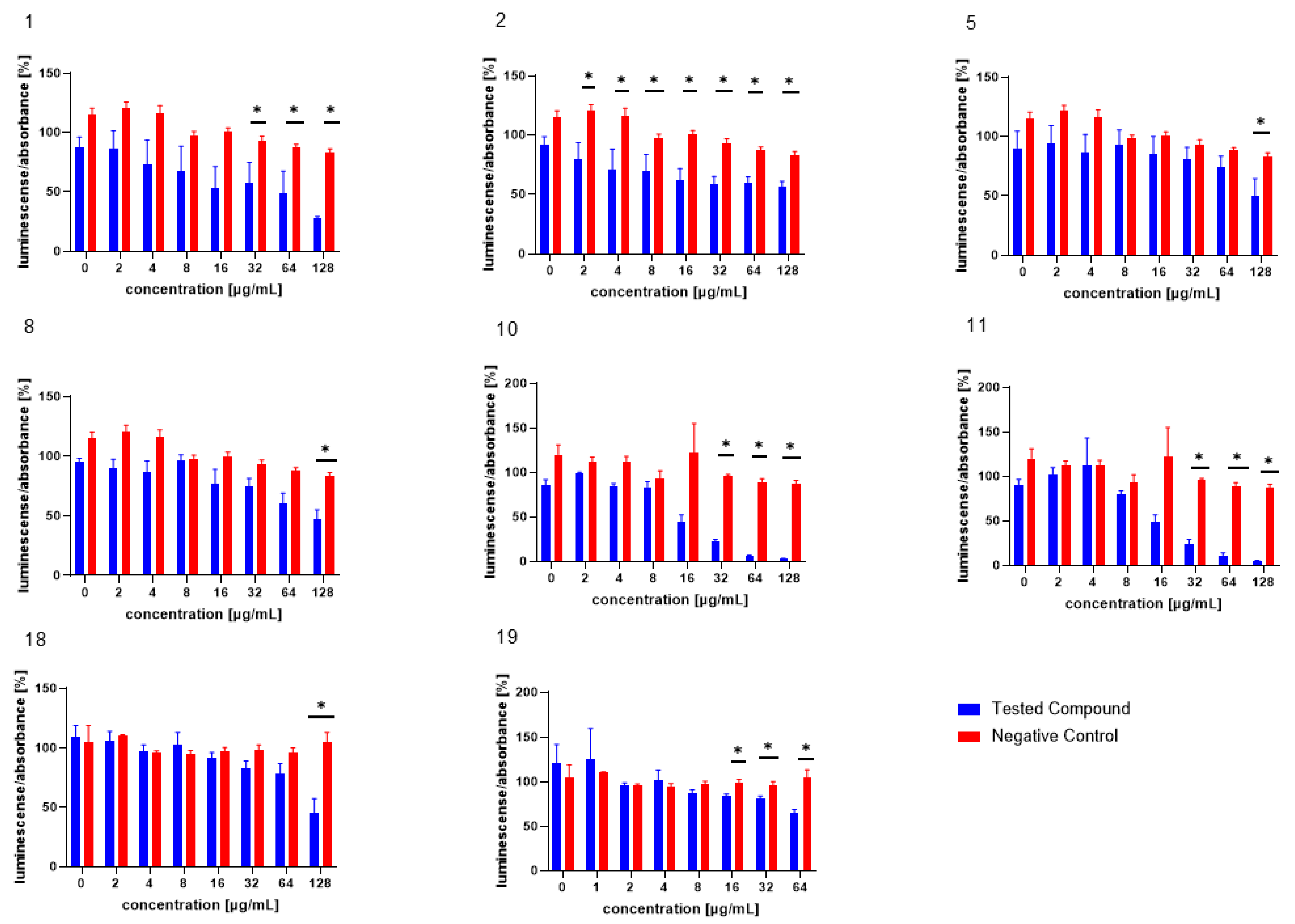
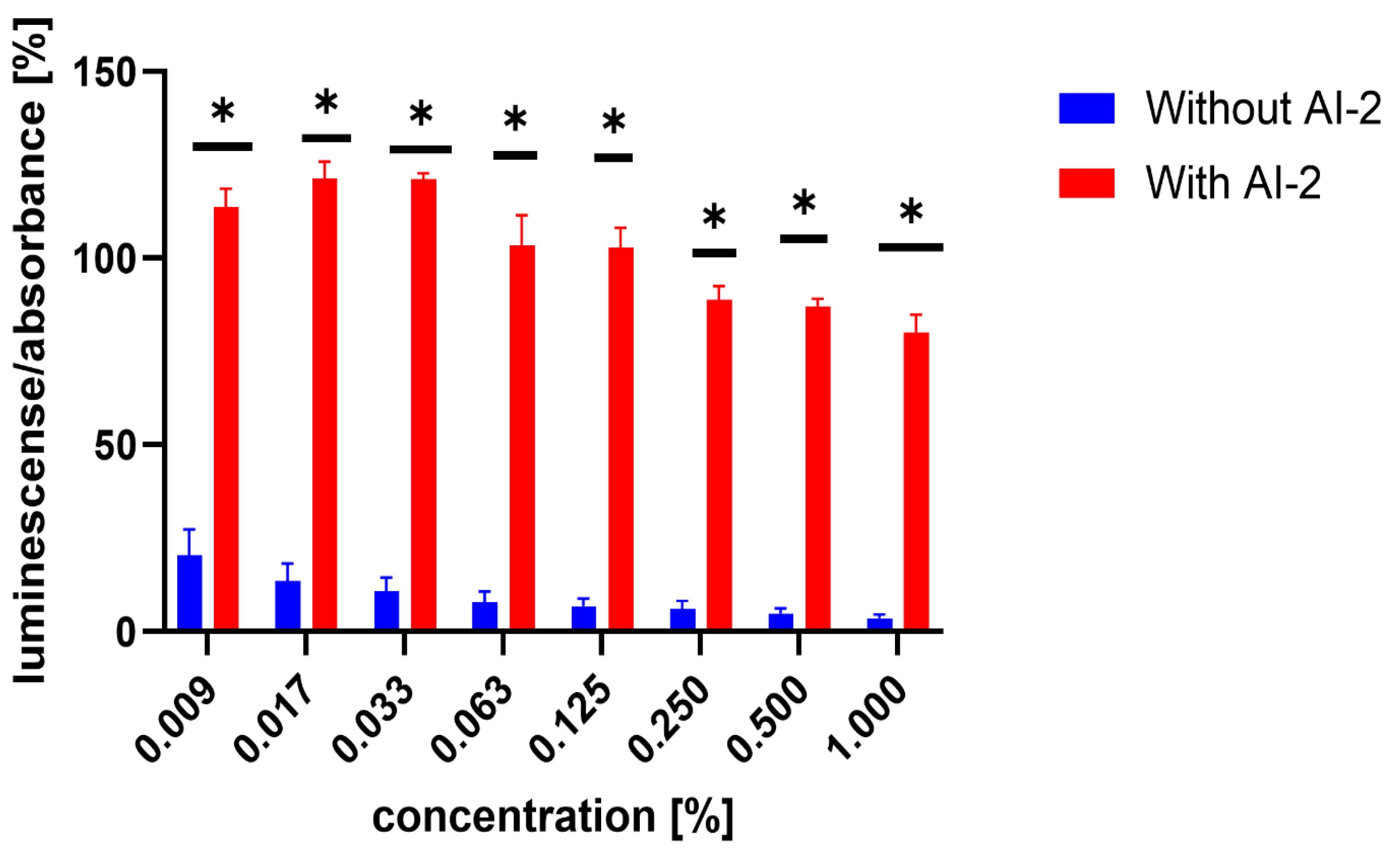
3.2. Inhibition of AI-2-Dependent QS in V. harveyi MM30
3.3. Inhibition of AI-2-Independent QS in V. harveyi MM30
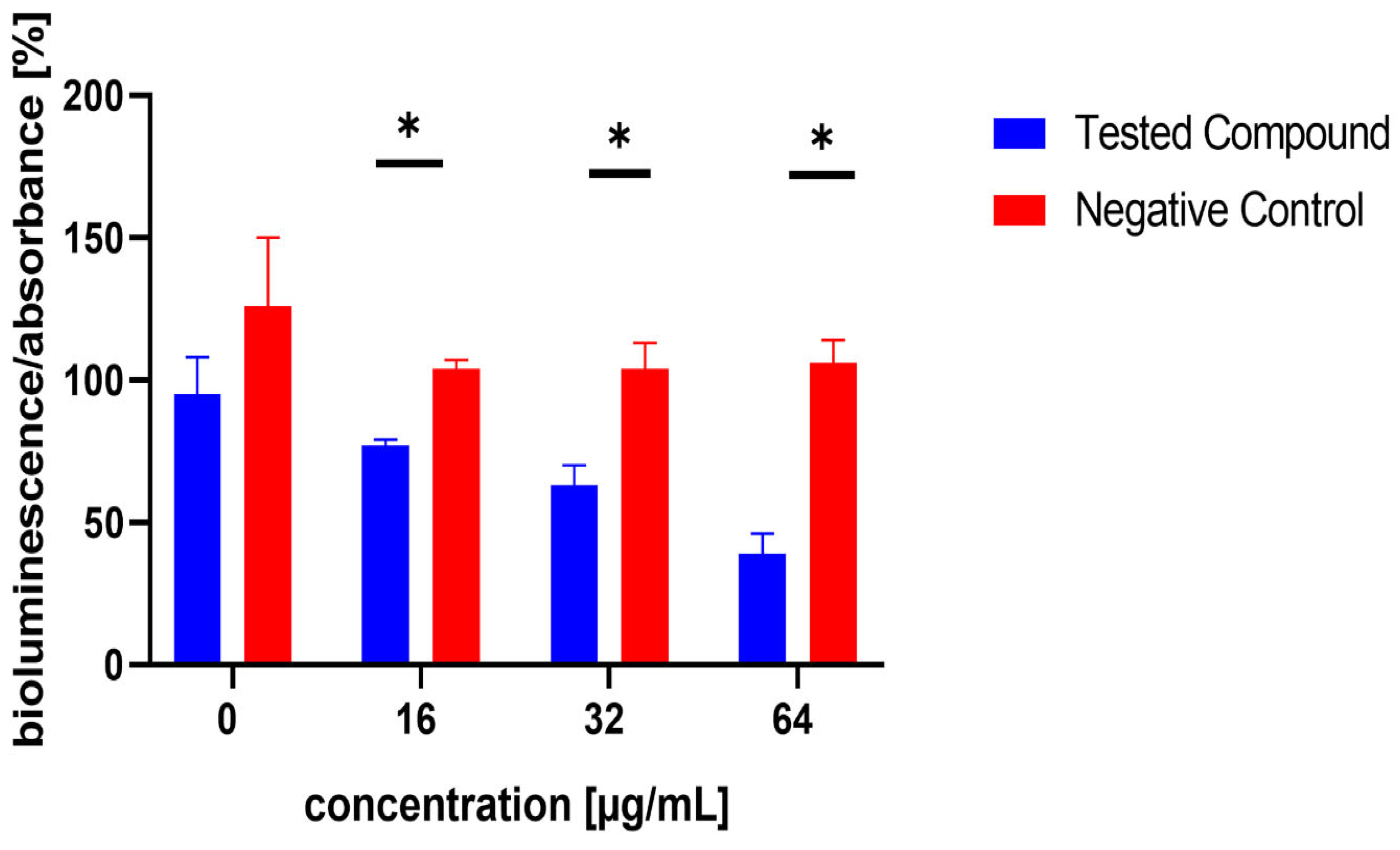
3.4. Inhibition of AI-2 Production in MRSA 7112
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
Abbreviations
| AB | Autoinducer-2 bioassay |
| AI-1 | Autoinducer-1 |
| AI-2 | Autoinducer-2 |
| CAI-1 | Cholera autoinducer-1 |
| CBDA | Cannabidiolic acid |
| CBG | Cannabigerol |
| CBGA | Cannabigerolic acid |
| DMSO | Dimethylsulfoxide |
| MH | Mueller–Hinton (medium) |
| MIC | Minimal inhibitory concentration |
| MRSA | Methicilin-resistant Staphylococcus aureus |
| NO | Nitric oxide |
| QS | Quorum sensing |
| QSI | Quorum-sensing inhibitor |
| THCA | Tetrahydrocannabinolic acid |
References
- Schluter, J.; Schoech, A.P.; Foster, K.R.; Mitri, S. The Evolution of Quorum Sensing as a Mechanism to Infer Kinship. PLoS Comput. Biol. 2016, 12, e1004848. [Google Scholar] [CrossRef] [PubMed]
- Kolenbrander, P.E.; Andersen, R.N.; Blehert, D.S.; Egland, P.G.; Foster, J.S.; Palmer, R.J. Communication among Oral Bacteria. Microbiol. Mol. Biol. Rev. 2002, 66, 486–505. [Google Scholar] [CrossRef]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum Sensing in Bacteria: The LuxR-LuxI Family of Cell Density-Responsive Transcriptional Regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef]
- Defoirdt, T.; Sorgeloos, P. Monitoring of Vibrio harveyi Quorum Sensing Activity in Real Time during Infection of Brine Shrimp Larvae. ISME J. 2012, 6, 2314–2319. [Google Scholar] [CrossRef] [PubMed]
- Aharoni, R.; Bronstheyn, M.; Jabbour, A.; Zaks, B.; Srebnik, M.; Steinberg, D. Oxazaborolidine Derivatives Inducing Autoinducer-2 Signal Transduction in Vibrio harveyi. Bioorg. Med. Chem. 2008, 16, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.G.; Meighen, E.A. Purification and Structural Identification of an Autoinducer for the Luminescence System of Vibrio harveyi. J. Biol. Chem. 1989, 264, 21670–21676. [Google Scholar] [CrossRef]
- Kadirvel, M.; Fanimarvasti, F.; Forbes, S.; McBain, A.; Gardiner, J.M.; Brown, G.D.; Freeman, S. Inhibition of Quorum Sensing and Biofilm Formation in Vibrio harveyi by 4-Fluoro-DPD; a Novel Potent Inhibitor of AI-2 Signalling. Chem. Commun. 2014, 50, 5000–5002. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.A.; Pomianek, M.E.; Kraml, C.M.; Taylor, R.K.; Semmelhack, M.F.; Bassler, B.L. The Major Vibrio Cholerae Autoinducer and Its Role in Virulence Factor Production. Nature 2007, 450, 883–886. [Google Scholar] [CrossRef] [PubMed]
- Henares, B.; Xu, Y.; Boon, E. A Nitric Oxide-Responsive Quorum Sensing Circuit in Vibrio harveyi Regulates Flagella Production and Biofilm Formation. Int. J. Mol. Sci. 2013, 14, 16473–16484. [Google Scholar] [CrossRef]
- Keller, L.; Surette, M.G. Communication in Bacteria: An Ecological and Evolutionary Perspective. Nat. Rev. Microbiol. 2006, 4, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Surette, M.G.; Miller, M.B.; Bassler, B.L. Quorum Sensing in Escherichia coli, Salmonella typhimurium, and Vibrio harveyi: A New Family of Genes Responsible for Autoinducer Production. Proc. Natl. Acad. Sci. USA 1999, 96, 1639–1644. [Google Scholar] [CrossRef] [PubMed]
- Aqawi, M.; Gallily, R.; Sionov, R.V.; Zaks, B.; Friedman, M.; Steinberg, D. Cannabigerol Prevents Quorum Sensing and Biofilm Formation of Vibrio harveyi. Front. Microbiol. 2020, 11, 858. [Google Scholar] [CrossRef]
- Kumar, L.; Chhibber, S.; Kumar, R.; Kumar, M.; Harjai, K. Zingerone Silences Quorum Sensing and Attenuates Virulence of Pseudomonas Aeruginosa. Fitoterapia 2015, 102, 84–95. [Google Scholar] [CrossRef]
- Martín-Rodríguez, A.J.; Ticona, J.C.; Jiménez, I.A.; Flores, N.; Fernández, J.J.; Bazzocchi, I.L. Flavonoids from Piper Delineatum Modulate Quorum-Sensing-Regulated Phenotypes in Vibrio harveyi. Phytochemistry 2015, 117, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Girennavar, B.; Cepeda, M.L.; Soni, K.A.; Vikram, A.; Jesudhasan, P.; Jayaprakasha, G.K.; Pillai, S.D.; Patil, B.S. Grapefruit Juice and Its Furocoumarins Inhibits Autoinducer Signaling and Biofilm Formation in Bacteria. Int. J. Food Microbiol. 2008, 125, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Vikram, A.; Jayaprakasha, G.K.; Jesudhasan, P.R.; Pillai, S.D.; Patil, B.S. Suppression of Bacterial Cell–Cell Signalling, Biofilm Formation and Type III Secretion System by Citrus Flavonoids. J. Appl. Microbiol. 2010, 109, 515–527. [Google Scholar] [CrossRef]
- Parai, D.; Banerjee, M.; Dey, P.; Chakraborty, A.; Islam, E.; Mukherjee, S.K. Effect of Reserpine on Pseudomonas aeruginosa Quorum Sensing Mediated Virulence Factors and Biofilm Formation. Biofouling 2018, 34, 320–334. [Google Scholar] [CrossRef] [PubMed]
- Brackman, G.; Defoirdt, T.; Miyamoto, C.; Bossier, P.; Van Calenbergh, S.; Nelis, H.; Coenye, T. Cinnamaldehyde and Cinnamaldehyde Derivatives Reduce Virulence in Vibrio Spp. by Decreasing the DNA-Binding Activity of the Quorum Sensing Response Regulator LuxR. BMC Microbiol. 2008, 8, 149. [Google Scholar] [CrossRef] [PubMed]
- Paczkowski, J.E.; Mukherjee, S.; McCready, A.R.; Cong, J.-P.; Aquino, C.J.; Kim, H.; Henke, B.R.; Smith, C.D.; Bassler, B.L. Flavonoids Suppress Pseudomonas Aeruginosa Virulence through Allosteric Inhibition of Quorum-Sensing Receptors. J. Biol. Chem. 2017, 292, 4064–4076. [Google Scholar] [CrossRef] [PubMed]
- Soni, D.; Smoum, R.; Breuer, A.; Mechoulam, R.; Steinberg, D. Effect of the Synthetic Cannabinoid HU-210 on Quorum Sensing and on the Production of Quorum Sensing-Mediated Virulence Factors by Vibrio harveyi. BMC Microbiol. 2015, 15, 159. [Google Scholar] [CrossRef] [PubMed]
- Friedman, L.; Smoum, R.; Feldman, M.; Mechoulam, R.; Steinberg, D. Does the Endocannabinoid Anandamide Affect Bacterial Quorum Sensing, Vitality, and Motility? Cannabis. Cannabinoid. Res. 2019, 4, 102–109. [Google Scholar] [CrossRef]
- Martinelli, D.; Grossmann, G.; Séquin, U.; Brandl, H.; Bachofen, R. Effects of Natural and Chemically Synthesized Furanones on Quorum Sensing in Chromobacterium Violaceum. BMC Microbiol. 2004, 4, 25. [Google Scholar] [CrossRef]
- Laganenka, L.; Sander, T.; Lagonenko, A.; Chen, Y.; Link, H.; Sourjik, V. Quorum Sensing and Metabolic State of the Host Control Lysogeny-Lysis Switch of Bacteriophage T1. mBio 2019, 10, e01884-19. [Google Scholar] [CrossRef]
- Norizan, S.; Yin, W.-F.; Chan, K.-G. Caffeine as a Potential Quorum Sensing Inhibitor. Sensors 2013, 13, 5117–5129. [Google Scholar] [CrossRef] [PubMed]
- Castillo, S.; Heredia, N.; García, S. 2(5H)-Furanone, Epigallocatechin Gallate, and a Citric-Based Disinfectant Disturb Quorum-Sensing Activity and Reduce Motility and Biofilm Formation of Campylobacter Jejuni. Folia Microbiol. 2015, 60, 89–95. [Google Scholar] [CrossRef]
- Xu, W.; Zhang, X.; Wang, L.; Zeng, W.; Sun, Y.; Zhou, C.; Zhou, T.; Shen, M. Effect of Chlorogenic Acid on the Quorum-sensing System of Clinically Isolated Multidrug-resistant Pseudomonas aeruginosa. J. Appl. Microbiol. 2022, 132, 1008–1017. [Google Scholar] [CrossRef]
- Čulenová, M.; Sychrová, A.; Hassan, S.T.S.; Berchová-Bímová, K.; Svobodová, P.; Helclová, A.; Michnová, H.; Hošek, J.; Vasilev, H.; Suchý, P.; et al. Multiple In Vitro Biological Effects of Phenolic Compounds from Morus Alba Root Bark. J. Ethnopharmacol. 2020, 248, 112296. [Google Scholar] [CrossRef]
- Ramić, D.; Klančnik, A.; Možina, S.S.; Dogsa, I. Elucidation of the AI-2 Communication System in the Food-Borne Pathogen Campylobacter Jejuni by Whole-Cell-Based Biosensor Quantification. Biosens. Bioelectron. 2022, 212, 114439. [Google Scholar] [CrossRef] [PubMed]
- Hong, N.T.X.; Baruah, K.; Vanrompay, D.; Bossier, P. Characterization of Phenotype Variations of Luminescent and Non-luminescent Variants of Vibrio harveyi Wild Type and Quorum Sensing Mutants. J. Fish Dis. 2016, 39, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Mok, K.C. Vibrio harveyi Quorum Sensing: A Coincidence Detector for Two Autoinducers Controls Gene Expression. EMBO J. 2003, 22, 870–881. [Google Scholar] [CrossRef] [PubMed]
- Cluzel, M.-E.; Zanella-Cléon, I.; Cozzone, A.J.; Fütterer, K.; Duclos, B.; Molle, V. The Staphylococcus Aureus Autoinducer-2 Synthase LuxS Is Regulated by Ser/Thr Phosphorylation. J. Bacteriol. 2010, 192, 6295–6301. [Google Scholar] [CrossRef] [PubMed]
- Appendino, G.; Giana, A.; Gibbons, S.; Maffei, M.; Gnavi, G.; Grassi, G.; Sterner, O. A Polar Cannabinoid from Cannabis sativa Var. Carma. Nat. Prod. Commun. 2008, 3, 1934578X0800301. [Google Scholar] [CrossRef]
- Appendino, G.; Gibbons, S.; Giana, A.; Pagani, A.; Grassi, G.; Stavri, M.; Smith, E.; Rahman, M.M. Antibacterial Cannabinoids from Cannabis sativa: A Structure−Activity Study. J. Nat. Prod. 2008, 71, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- DeKeersmaecker, S.C.J.; Vanderleyden, J. Constraints on Detection of Autoinducer-2 (AI-2) Signalling Molecules Using Vibrio harveyi as a Reporter. Microbiology 2003, 149, 1953–1956. [Google Scholar] [CrossRef]
- Filer, C.N. Acidic Cannabinoid Decarboxylation. Cannabis. Cannabinoid. Res. 2022, 7, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Naik, R.; Harmalkar, D.S.; Xu, X.; Jang, K.; Lee, K. Bioactive Benzofuran Derivatives: Moracins A–Z in Medicinal Chemistry. Eur. J. Med. Chem. 2015, 90, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Škovranová, G.; Čulenová, M.; Treml, J.; Dzurická, L.; Marova, I.; Sychrová, A. Prenylated Phenolics from Morus Alba against MRSA Infections as a Strategy for Wound Healing. Front. Pharmacol. 2022, 13, 1068371. [Google Scholar] [CrossRef]
- Rasmussen, T.B.; Manefield, M.; Andersen, J.B.; Eberl, L.; Anthoni, U.; Christophersen, C.; Steinberg, P.; Kjelleberg, S.; Givskov, M. How Delisea Pulchra Furanones Affect Quorum Sensing and Swarming Motility in Serratia Liquefaciens MG1. Microbiology 2000, 146, 3237–3244. [Google Scholar] [CrossRef]
- Ahmad, A.; Viljoen, A.M.; Chenia, H.Y. The Impact of Plant Volatiles on Bacterial Quorum Sensing. Lett. Appl. Microbiol. 2015, 60, 8–19. [Google Scholar] [CrossRef]
- Yue, J.; Yang, H.; Liu, S.; Song, F.; Guo, J.; Huang, C. Influence of Naringenin on the Biofilm Formation of Streptococcus Mutans. J. Dent. 2018, 76, 24–31. [Google Scholar] [CrossRef] [PubMed]
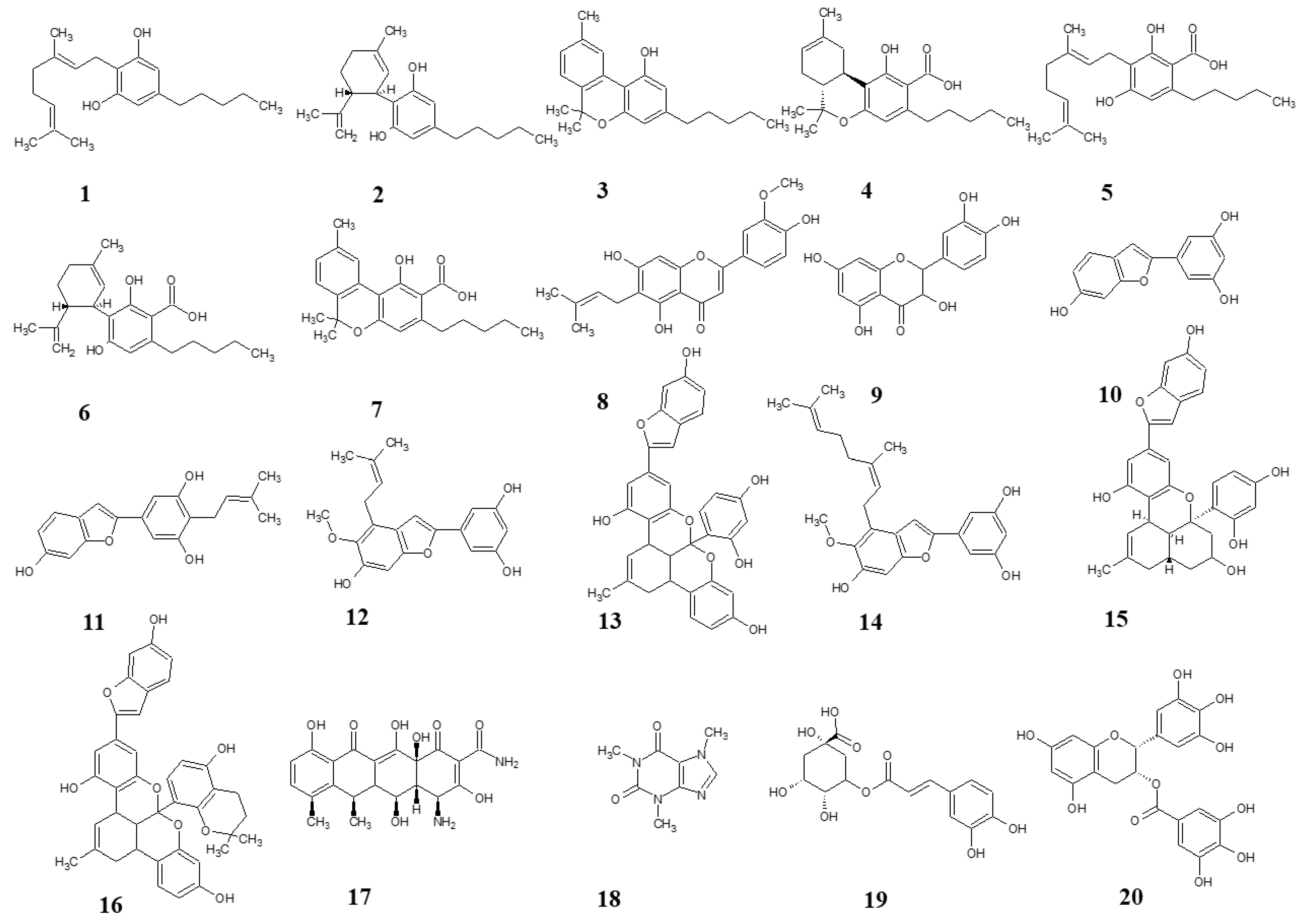




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class of Compounds | Number | Name | MIC MRSA | MIC V.h. |
|---|---|---|---|---|
| Neutral cannabinoid | 1 | Cannabigerol | 64 µg/mL | >128 µg/mL |
| 2 | Cannabidiol | 32 µg/mL | >128 µg/mL | |
| 3 | Cannabinol | 64 µg/mL | >128 µg/mL | |
| Cannabinoid acid | 4 | Tetrahydrocannabinolic acid | 128 µg/mL | >128 µg/mL |
| 5 | Cannabigerolic acid | 128 µg/mL | >128 µg/mL | |
| 6 | Cannabidiolic acid | 64 µg/mL | >128 µg/mL | |
| 7 | Cannabinolic acid | 128 µg/mL | >128 µg/mL | |
| Prenylated flavonoid | 8 | Cannflavin B | 64 µg/mL | >128 µg/mL |
| Non-prenylated flavonoid | 9 | Quercetin | >128 µg/mL | >128 µg/mL |
| Non-prenylated arylbenzofuran | 10 | Moracin M | >128 µg/mL | >128 µg/mL |
| Prenylated arylbenzofuran | 11 | Moracin C | 128 µg/mL | >128 µg/mL |
| 12 | Moracin T | >128 µg/mL | >128 µg/mL | |
| 13 | Albanol B | 16 µg/mL | >128 µg/mL | |
| 14 | Mulberrofuran Y | 128 µg/mL | >128 µg/mL | |
| 15 | Mulberrofuran G | >128 µg/mL | >128 µg/mL | |
| 16 | Mulberrofuran K | >128 µg/mL | >128 µg/mL | |
| Positive control | 17 | Doxycycline | 4 µg/mL | 8 µg/mL |
| 18 | Caffeine | N.D. * | >128 µg/mL | |
| 19 | Chlorogenic acid | N.D. | 128 µg/mL | |
| 20 | Epigallocatechine gallate | N.D. | 16 µg/mL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Helcman, M.; Šmejkal, K.; Čulenová, M.; Béres, T.; Treml, J. Natural Phenolics Disrupt Microbial Communication by Inhibiting Quorum Sensing. Microorganisms 2025, 13, 287. https://doi.org/10.3390/microorganisms13020287
Helcman M, Šmejkal K, Čulenová M, Béres T, Treml J. Natural Phenolics Disrupt Microbial Communication by Inhibiting Quorum Sensing. Microorganisms. 2025; 13(2):287. https://doi.org/10.3390/microorganisms13020287
Chicago/Turabian StyleHelcman, Martin, Karel Šmejkal, Marie Čulenová, Tibor Béres, and Jakub Treml. 2025. "Natural Phenolics Disrupt Microbial Communication by Inhibiting Quorum Sensing" Microorganisms 13, no. 2: 287. https://doi.org/10.3390/microorganisms13020287
APA StyleHelcman, M., Šmejkal, K., Čulenová, M., Béres, T., & Treml, J. (2025). Natural Phenolics Disrupt Microbial Communication by Inhibiting Quorum Sensing. Microorganisms, 13(2), 287. https://doi.org/10.3390/microorganisms13020287