
Evaluating Rhizobacterial Antagonists for Controlling Cercospora beticola and Promoting Growth in Beta vulgaris

 ,
,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Material
2.2. Isolation of Bacteria from Sugar Beet Rhizosphere Soil
2.3. Screening of Antagonistic Activity among Rhizosphere Bacteria
2.3.1. Dual-Culture Setup for Confrontation Testing
2.3.2. Antibiotic Activity through Bacterial Supernatant Analysis
2.4. Fourier Transform Infrared Spectroscopy (FTIR) Analysis
2.5. DNA Extraction and PCR Amplification of 16S rRNA Gene and Sanger Sequencing
2.6. PCR Amplifications of Lipopeptide-Encoding Genes
2.7. Biochemical and Plant Growth Promotion Tests
2.8. Effects of Bacterial Isolates on Sugar Beet Growth in Greenhouse Conditions
2.9. Effects of Four Bacterial Isolates on Sugar Beet Growth in the Field
2.10. Statistical Analysis
3. Results
3.1. Isolating and Identifying Bacteria from the Rhizosphere Soil of Sugar Beet
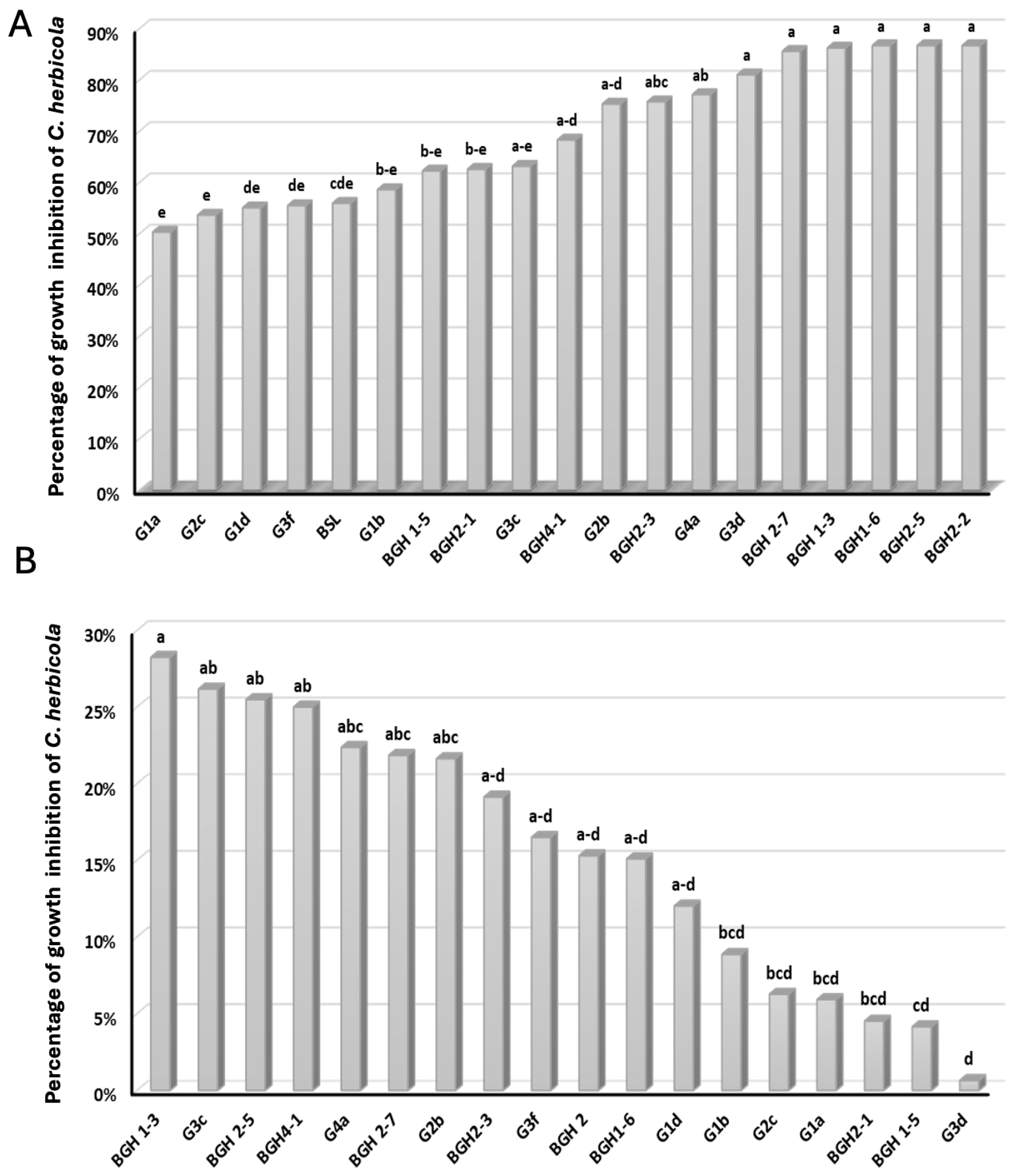
3.2. Antagonism against C. beticola in Confrontation Assay in PDA Medium and PDA Amended with Supernatant
3.3. PCR Detection of Lipopeptide Genes
3.4. Biochemical and Plant Growth Promotion Tests
3.5. Effects of Bacterial Isolates on Sugar Beet Growth in Controlled Conditions
3.6. Effects of Four Bacterial Isolates on Sugar Beet Growth in the Field
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tan, W.; Li, K.; Liu, D.; Xing, W. Cercospora leaf spot disease of sugar beet. Plant Signal. Behav. 2023, 18, 2214765. [Google Scholar] [CrossRef] [PubMed]
- Rangel, L.I.; Spanner, R.E.; Ebert, M.K.; Pethybridge, S.J.; Stukenbrock, E.H.; de Jonge, R.; Secor, G.A.; Bolton, M.D. Cercospora beticola: The intoxicating lifestyle of the leaf spot pathogen of sugar beet. Mol. Plant Pathol. 2020, 21, 1020–1041. [Google Scholar] [CrossRef] [PubMed]
- Skaracis, G.N.; Biancardi, E. Breeding for Cercospora resistance in sugarbeet. In Cercospora beticola Sacc. Biology, Agronomic Influence and Control Measures in Sugar Beet, 2nd ed.; Asher, M.J.C., Ed.; Advances in Sugar Beet Research; International Institute for Beet Research: Brussels, Belgium, 2000; pp. 177–195. [Google Scholar]
- Jacobsen, B.J.; Franc, G.D.; Harveson, R.M.; Hanson, L.E.; Hein, G.L. Foliar disease casused by fungi and Oomycetes. In Compendium of Beet Diseases and Pests: The Cercospora Leaf Spot, 2nd ed.; Harveson, R.M., Hanson, L.E., Hein, G.L., Eds.; American Phytopathological Society: St Paul, MN, USA, 2009; pp. 7–10. [Google Scholar]
- Rossi, V.; Meriggi, P.; Biancardi, E.; Rosso, F. Effect of Cercospora leaf spot on sugarbeet growth, yield and quality. In Cercospora beticola Sacc. Biology, Agronomic Influence and Control Measures in Sugar Beet; International Institute for Beet Research: Brussels, Belgium, 2000; pp. 49–76. [Google Scholar]
- Lartey, R.T.; Weiland, J.J.; Bucklin-Comiskey, S. A PCR protocol for rapid detection of Cercospora beticola in sugarbeet tissues. J. Sugar Beet Res. 2003, 40, 1–10. [Google Scholar] [CrossRef]
- Brown, J.K.M. Yield penalties of disease resistance in crops. Curr. Opin. Plant Biol. 2002, 5, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Gummert, A.; Ladewig, E.; Bürcky, K.; Märländer, B. Variety resistance to Cercospora leaf spot and fungicide application as tools of integrated pest management in sugar beet cultivation—A German case study. Crop Prot. 2015, 72, 182–194. [Google Scholar] [CrossRef]
- Vogel, J.; Kenter, C.; Holst, C.; Marlander, B. New Generation of Resistant Sugar Beet Varieties for Advanced Integrated Management of Cercospora Leaf Spot in Central Europe. Front. Plant Sci. 2018, 9, 222. [Google Scholar] [CrossRef] [PubMed]
- Panella, L.; Lewellen, R.T. Broadening the genetic base of sugar beet: Introgression from wild relatives. Euphytica 2007, 154, 383–400. [Google Scholar] [CrossRef]
- Davidson, R.M.; Hanson, L.E.; Franc, G.D.; Panella, L. Analysis of β-tubulin gene fragments from benzimidazole-sensitive and-tolerant Cercospora beticola. J. Phytopathol. 2006, 154, 321–328. [Google Scholar] [CrossRef]
- Secor, G.A.; Rivera, V.V.; Khan, M.F.R.; Gudmestad, N.C. Monitoring Fungicide Sensitivity of Cercospora beticola of Sugar Beet for Disease Management Decisions. Plant Dis. 2010, 94, 1272–1282. [Google Scholar] [CrossRef] [PubMed]
- Karaoglanidis, G.S.; Thanassoulopoulos, C.C. Cross-resistance patterns among sterol biosynthesis inhibiting fungicides (SBIs) in Cercospora beticola. Eur. J. Plant Pathol. 2003, 109, 929–934. [Google Scholar] [CrossRef]
- El Housni, Z.; Ezrari, S.; Tahiri, A.; Ouijja, A. Resistance of Cercospora beticola Sacc isolates to thiophanate methyl (benzimidazole), demethylation inhibitors and quinone outside inhibitors in Morocco. EPPO Bull. 2020, 50, 350–357. [Google Scholar] [CrossRef]
- Kirk, W.W.; Hanson, L.E.; Franc, G.D.; Stump, W.L.; Gachango, E.; Clark, G.; Stewart, J. First report of strobilurin resistance in Cercospora beticola in sugar beet (Beta vulgaris) in Michigan and Nebraska, USA. New Dis. Rep. 2012, 26, 3. [Google Scholar] [CrossRef]
- Hudec, K.; Mihók, M.; Roháčik, T.; Mišľan, Ľ. Sensitivity of Cercospora beticola to fungicides in Slovakia. Acta Fytotech. Zootech. 2020, 23, 147–154. [Google Scholar] [CrossRef]
- Trkulja, N.; Hristov, N. Morphological and genetic diversity of Cercospora beticola isolates. In Proceedings of the International Conference on BioScience: Biotechnology and Biodiversity-Step in the Future, the Fourth Joint UNS-PSU Conference, Novi Sad, Serbia, 18–20 June 2012; pp. 18–20. [Google Scholar]
- Ahmed, M.F.A.; Mikhail, S.P.H.; Shaheen, S.I. Performance efficiency of some biocontrol agents on controlling Cercospora leaf spot disease of sugar beet plants under organic agriculture system. Eur. J. Plant Pathol. 2023, 167, 145–155. [Google Scholar] [CrossRef]
- Bonaterra, A.; Badosa, E.; Daranas, N.; Frances, J.; Rosello, G.; Montesinos, E. Bacteria as Biological Control Agents of Plant Diseases. Microorganisms 2022, 10, 1759. [Google Scholar] [CrossRef] [PubMed]
- Lahlali, R.; Aksissou, W.; Lyousfi, N.; Ezrari, S.; Blenzar, A.; Tahiri, A.; Ennahli, S.; Hrustić, J.; MacLean, D.; Amiri, S. Biocontrol activity and putative mechanism of Bacillus amyloliquefaciens (SF14 and SP10), Alcaligenes faecalis ACBC1, and Pantoea agglomerans ACBP1 against brown rot disease of fruit. Microb. Pathog. 2020, 139, 103914. [Google Scholar] [CrossRef]
- Ezrari, S.; Mhidra, O.; Radouane, N.; Tahiri, A.; Polizzi, G.; Lazraq, A.; Lahlali, R. Potential role of rhizobacteria isolated from citrus rhizosphere for biological control of citrus dry root rot. Plants 2021, 10, 872. [Google Scholar] [CrossRef] [PubMed]
- Radouane, N.; Adadi, H.; Ezrari, S.; Kenfaoui, J.; Belabess, Z.; Mokrini, F.; Barka, E.A.; Lahlali, R. Exploring the Bioprotective Potential of Halophilic Bacteria against Major Postharvest Fungal Pathogens of Citrus Fruit Penicillium digitatum and Penicillium italicum. Horticulturae 2023, 9, 922. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.L. Fungi vs. Fungi in Biocontrol: An Overview of Fungal Antagonists Applied against Fungal Plant Pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef]
- Dimkić, I.; Janakiev, T.; Petrović, M.; Degrassi, G.; Fira, D. Plant-associated Bacillus and Pseudomonas antimicrobial activities in plant disease suppression via biological control mechanisms-A review. Physiol. Mol. Plant Pathol. 2022, 117, 101754. [Google Scholar] [CrossRef]
- Arzanlou, M.; Mousavi, S.; Bakhshi, M.; Khakvar, R.; Bandehagh, A. Inhibitory effects of antagonistic bacteria inhabiting the rhizosphere of the sugarbeet plants, on Cercospora beticola Sacc., the causal agent of Cercospora leaf spot disease on sugarbeet. J. Plant Prot. Res. 2016, 56, 6–14. [Google Scholar] [CrossRef]
- Dervišević, M.; Đorđević, N.; Knežević, I.; Đorđević, S. Antagonistic activity of bacterial isolates against Cercospora beticola in laboratory conditions. In Proceedings of the Book of Abstracts: X International Symposium on Agricultural Sciences “AgroReS 2021”, Trebinje, Bosnia and Herzegovina, 27–29 May 2021; Poljoprivredni Fakultet: Banja Luka, Bosnia and Herzegovina, 2021; p. 47. [Google Scholar]
- Alotaibi, F.; St-Arnaud, M.; Hijri, M. In-Depth Characterization of Plant Growth Promotion Potentials of Selected Alkanes-Degrading Plant Growth-Promoting Bacterial Isolates. Front. Microbiol. 2022, 13, 863702. [Google Scholar] [CrossRef] [PubMed]
- Ducousso-Detrez, A.; Lahrach, Z.; Fontaine, J.; Lounes-Hadj Sahraoui, A.; Hijri, M. Cultural techniques capture diverse phosphate-solubilizing bacteria in rock phosphate-enriched habitats. Front. Microbiol. 2024, 15, 1280848. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Aeron, A.; Khare, E.; Jha, C.K.; Meena, V.S.; Mohammed, S.; Aziz, A.; Islam, M.T.; Kim, K.; Meena, S.K.; Pattanayak, A.; et al. Revisiting the plant growth-promoting rhizobacteria: Lessons from the past and objectives for the future. Arch. Microbiol. 2020, 202, 665–676. [Google Scholar] [CrossRef] [PubMed]
- KaragÖZ, H.; Cakmakci, R.; Hosseinpour, A.; Kodaz, S. Alleviation of water stress and promotion of the growth of sugar beet (Beta vulgaris L.) plants by multi-traits rhizobacteria. Appl. Ecol. Environ. Res. 2018, 16, 6801–6813. [Google Scholar]
- Farhaoui, A.; Adadi, A.; Tahiri, A.; El Alami, N.; Khayi, S.; Mentag, R.; Ezrari, S.; Radouane, N.; Mokrini, F.; Belabess, Z.; et al. Biocontrol potential of plant growth-promoting rhizobacteria (PGPR) against Sclerotiorum rolfsii diseases on sugar beet (Beta vulgaris L.). Physiol. Mol. Plant Pathol. 2022, 119, 101829. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Llop, P.; Bonaterra, A.; Peñalver, J.; López, M.a.M. Development of a Highly Sensitive Nested-PCR Procedure Using a Single Closed Tube for Detection of Erwinia amylovora in Asymptomatic Plant Material. Appl. Environ. Microbiol. 2000, 66, 2071–2078. [Google Scholar] [CrossRef]
- Einloft, T.C.; Hartke, S.; de Oliveira, P.B.; Saraiva, P.S.; Dionello, R.G. Selection of rhizobacteria for biocontrol of Fusarium verticillioides on non-rhizospheric soil and maize seedlings roots. Eur. J. Plant Pathol. 2021, 160, 503–518. [Google Scholar] [CrossRef]
- Bailadores Bollona, J.P.; Delgado Paredes, G.E.; Wagner, M.L.; Rojas Idrogo, C. In vitro tissue culture, preliminar phytochemical analysis, and antibacterial activity of Psittacanthus linearis (Killip) JK Macbride (Loranthaceae). Rev. Colomb. Biotecnol. 2019, 21, 22–35. [Google Scholar]
- Wolf, P.F.J.; Verreet, J.A. An integrated pest management system in Germany for the control of fungal leaf diseases in sugar beet: The IPM sugar beet model. Plant Dis. 2002, 86, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Sehsah, M.D.; El-Kot, G.A.; El-Nogoumy, B.A.; Alorabi, M.; El-Shehawi, A.M.; Salama, N.H.; El-Tahan, A.M. Efficacy of Bacillus subtilis, Moringa oleifera seeds extract and potassium bicarbonate on Cercospora leaf spot on sugar beet. Saudi J. Biol. Sci. 2022, 29, 2219–2229. [Google Scholar] [CrossRef]
- Parthipan, P.; Preetham, E.; Machuca, L.L.; Rahman, P.K.S.M.; Murugan, K.; Rajasekar, A. Biosurfactant and degradative enzymes mediated crude oil degradation by bacterium Bacillus subtilis A1. Front. Microbiol. 2017, 8, 193. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.M.; Mody, K.; Mishra, A.; Jha, B. Physicochemical characterization of biosurfactant and its potential to remove oil from soil and cotton cloth. Carbohydr. Polym. 2012, 89, 1110–1116. [Google Scholar] [CrossRef]
- Hajfarajollah, H.; Mokhtarani, B.; Noghabi, K.A. Newly antibacterial and antiadhesive lipopeptide biosurfactant secreted by a probiotic strain, Propionibacterium freudenreichii. Appl. Biochem. Biotechnol. 2014, 174, 2725–2740. [Google Scholar] [CrossRef]
- Ramani, K.; Jain, S.C.; Mandal, A.B.; Sekaran, G. Microbial induced lipoprotein biosurfactant from slaughterhouse lipid waste and its application to the removal of metal ions from aqueous solution. Colloids Surf. B Biointerfaces 2012, 97, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Khmelevtsova, L.E.; Sazykin, I.S.; Azhogina, T.N.; Sazykina, M.A. Influence of Agricultural Practices on Bacterial Community of Cultivated Soils. Agriculture 2022, 12, 371. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, L.; Chen, Q.; Wen, X.; Liao, Y. Conservation tillage increases soil bacterial diversity in the dryland of northern China. Agron. Sustain. Dev. 2016, 36, 28. [Google Scholar] [CrossRef]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glockner, F.O.; Ludwig, W.; Schleifer, K.H.; Whitman, W.B.; Euzeby, J.; Amann, R.; Rossello-Mora, R. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef]
- Braun-Kiewnick, A.; Jacobsen, B.J.; Sands, D.C. Biological control of Pseudomonas syringae pv. syringae, the causal agent of basal kernel blight of barley, by antagonistic Pantoea agglomerans. Phytopathology 2000, 90, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Schuhegger, R.; Ihring, A.; Gantner, S.; Bahnweg, G.; Knappe, C.; Vogg, G.; Hutzler, P.; Schmid, M.; Van Breusegem, F.; Eberl, L.E.O. Induction of systemic resistance in tomato by N-acyl-L-homoserine lactone-producing rhizosphere bacteria. Plant Cell Environ. 2006, 29, 909–918. [Google Scholar] [CrossRef] [PubMed]
- Karimi, E.; Safaie, N.; Shams-Baksh, M.; Mahmoudi, B. Bacillus amyloliquefaciens SB14 from rhizosphere alleviates Rhizoctonia damping-off disease on sugar beet. Microbiol. Res. 2016, 192, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Khoa, N.Đ.; Giàu, N.Đ.N.; Tuấn, T.Q. Effects of Serratia nematodiphila CT-78 on rice bacterial leaf blight caused by Xanthomonas oryzae pv. oryzae. Biol. Control. 2016, 103, 1–10. [Google Scholar] [CrossRef]
- Sarhan, E.A.D. Induction of induced systemic resistance in fodder beet (Beta vulgaris L.) to Cercospora leaf spot caused by (Cercospora beticola Sacc.). Egypt. J. Phytopathol. 2018, 46, 39–59. [Google Scholar] [CrossRef]
- Pethybridge, S.J.; Vaghefi, N.; Kikkert, J.R. Management of Cercospora leaf spot in conventional and organic table beet production. Plant Dis. 2017, 101, 1642–1651. [Google Scholar] [CrossRef]
- Yaseen, Y.; Gancel, F.; Béchet, M.; Drider, D.; Jacques, P. Study of the correlation between fengycin promoter expression and its production by Bacillus subtilis under different culture conditions and the impact on surfactin production. Arch. Microbiol. 2017, 199, 1371–1382. [Google Scholar] [CrossRef]
- Mostertz, J.; Scharf, C.; Hecker, M.; Homuth, G. Transcriptome and proteome analysis of Bacillus subtilis gene expression in response to superoxide and peroxide stress. Microbiology 2004, 150, 497–512. [Google Scholar] [CrossRef] [PubMed]
- Nakano, S.; Küster-Schöck, E.; Grossman, A.D.; Zuber, P. Spx-dependent global transcriptional control is induced by thiol-specific oxidative stress in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2003, 100, 13603–13608. [Google Scholar] [CrossRef]
- El Housni, Z.; Tahiri, A.; Ezrari, S.; Radouane, N.; Ouijja, A. Occurrence of Cercospora beticola Sacc populations resistant to benzimidazole, demethylation-inhibiting, and quinone outside inhibitors fungicides in Morocco. Eur. J. Plant Pathol. 2023, 165, 73–83. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Saad, A.M.; Soliman, S.M.; Salem, H.M.; Ahmed, A.I.; Mahmood, M.; El-Tahan, A.M.; Ebrahim, A.A.M.; Abd El-Mageed, T.A.; Negm, S.H.; et al. Plant growth-promoting microorganisms as biocontrol agents of plant diseases: Mechanisms, challenges and future perspectives. Front. Plant Sci. 2022, 13, 923880. [Google Scholar] [CrossRef]
- ÇInar, V.M.; Aydın, Ü. The effects of some biofertilizers on yield, chlorophyll index and sugar content in sugar beet (Beta vulgaris var. saccharifera L.). Ege Üniv. Ziraat Fakültesi Derg. 2021, 58, 163–170. [Google Scholar] [CrossRef]
- Collins, D.P.; Jacobsen, B.J. Optimizing a Bacillus subtilis isolate for biological control of sugar beet Cercospora leaf spot. Biol. Control. 2003, 26, 153–161. [Google Scholar] [CrossRef]
- Gowtham, H.G.; Murali, M.; Singh, S.B.; Lakshmeesha, T.R.; Narasimha Murthy, K.; Amruthesh, K.N.; Niranjana, S.R. Plant growth promoting rhizobacteria Bacillus amyloliquefaciens improves plant growth and induces resistance in chilli against anthracnose disease. Biol. Control 2018, 126, 209–217. [Google Scholar] [CrossRef]
- Gao, H.; Lu, C.; Wang, H.; Wang, L.; Yang, Y.; Jiang, T.; Li, S.; Xu, D.; Wu, L. Production exopolysaccharide from Kosakonia cowanii LT-1 through solid-state fermentation and its application as a plant growth promoter. Int. J. Biol. Macromol. 2020, 150, 955–964. [Google Scholar] [CrossRef]
- Shoaib, A.; Ali, H.; Javaid, A.; Awan, Z.A. Contending charcoal rot disease of mungbean by employing biocontrol Ochrobactrum ciceri and zinc. Physiol. Mol. Biol. Plants 2020, 26, 1385–1397. [Google Scholar] [CrossRef]
- Samaras, A.; Roumeliotis, E.; Ntasiou, P.; Karaoglanidis, G. Bacillus subtilis MBI600 promotes growth of tomato plants and induces systemic resistance contributing to the control of soilborne pathogens. Plants 2021, 10, 1113. [Google Scholar] [CrossRef]
- Muthu Narayanan, M.; Ahmad, N.; Shivanand, P.; Metali, F. The role of endophytes in combating fungal-and bacterial-induced stress in plants. Molecules 2022, 27, 6549. [Google Scholar] [CrossRef]
- Li, J.; Ovakim, D.H.; Charles, T.C.; Glick, B.R. An ACC deaminase minus mutant of Enterobacter cloacae UW4No longer promotes root elongation. Curr. Microbiol. 2000, 41, 101–105. [Google Scholar] [CrossRef]
- Grover, M.; Bodhankar, S.; Sharma, A.; Sharma, P.; Singh, J.; Nain, L. PGPR mediated alterations in root traits: Way toward sustainable crop production. Front. Sustain. Food Syst. 2021, 4, 618230. [Google Scholar] [CrossRef]
- Desbrosses, G.; Contesto, C.; Varoquaux, F.; Galland, M.; Touraine, B. PGPR-Arabidopsis interactions is a useful system to study signaling pathways involved in plant developmental control. Plant Signal. Behav. 2009, 4, 319–321. [Google Scholar] [CrossRef]
- Meena, M.; Swapnil, P.; Divyanshu, K.; Kumar, S.; Harish; Tripathi, Y.N.; Zehra, A.; Marwal, A.; Upadhyay, R.S. PGPR-mediated induction of systemic resistance and physiochemical alterations in plants against the pathogens: Current perspectives. J. Basic Microbiol. 2020, 60, 828–861. [Google Scholar] [CrossRef]
- Ramarathnam, R.; Bo, S.; Chen, Y.; Fernando, W.G.D.; Xuewen, G.; De Kievit, T. Molecular and biochemical detection of fengycin-and bacillomycin D-producing Bacillus spp., antagonistic to fungal pathogens of canola and wheat. Can. J. Microbiol. 2007, 53, 901–911. [Google Scholar] [CrossRef]
- Dimkić, I.; Živković, S.; Berić, T.; Ivanović, Ž.; Gavrilović, V.; Stanković, S.; Fira, D. Characterization and evaluation of two Bacillus strains, SS-12.6 and SS-13.1, as potential agents for the control of phytopathogenic bacteria and fungi. Biological Control 2013, 65, 312–321. [Google Scholar] [CrossRef]
- Hsieh, F.C.; Li, M.C.; Lin, T.C.; Kao, S.S. Rapid detection and characterization of surfactin-producing Bacillus subtilis and closely related species based on PCR. Curr. Microbiol. 2004, 49, 186–191. [Google Scholar] [CrossRef]
- Syed-Ab-Rahman, S.F.; Carvalhais, L.C.; Chua, E.; Xiao, Y.; Wass, T.J.; Schenk, P.M. Identification of soil bacterial isolates suppressing different phytophthora spp. And promoting plant growth. Front. Plant Sci. 2018, 871. [Google Scholar] [CrossRef]
- Etesami, H.; Alikhani, H.A.; Mirseyed Hosseini, H. Evaluation of halotolerant endophytic bacteria isolated from the halophyte suaeda for biological control of fungal rice pathogens. 2019. Arch. Für Phytopathol. Und Pflanzenschutz 2019, 52. [Google Scholar]
- Dinesh, R.; Anandaraj, M.; Kumar, A.; Bini, Y.K.; Subila, K.P.; Aravind, R. Isolation, characterization, and evaluation of multi-trait plant growth promoting rhizobacteria for their growth promoting and disease suppressing effects on ginger. Microbiol. Res. 2015, 173, 34–43. [Google Scholar] [CrossRef]
- Jamilah, I.T.; Meryandini, A.; Rusmana, I.; Suwanto, A.; Mubarik, N.R. Activity of proteolytic and amylolytic enzymes from Bacillus spp. isolated from shrimp ponds. Microbiol. Indones. 2009, 3, 4. [Google Scholar] [CrossRef]
- Pertiwiningrum, A.; Anggraini, F.D.; Fitrianto, N.A. Isolation and identification of bacterial protease enzyme of leather waste. J. Indones. Trop. Anim. Agric. 2017, 42, 1. [Google Scholar] [CrossRef]
- Hsu, S.C.; Lockwood, J.L. Powdered Chitin Agar as a Selective Medium for Enumeration of Actinomycetes in Water and Soil1. Applied Microbiology 1975, 29, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.A.; Forzani, M.V.; Soares, R.d.S.; Sibov, S.T.; Vieira, J.D.G. Isolation and selection of plant growth-promoting bacteria associated with sugarcane. Pesqui. Agropecuária Trop. 2016, 46, 149–158. [Google Scholar] [CrossRef]
- Modi, K.; Patel, P.; Parmar, K. Isolation, screening and characterization of PGPR from rhizosphere of rice. Int. J. Pure Appl. Biosci. 2017, 5, 264–270. [Google Scholar]
- Yuttavanichakul, W.; Lawongsa, P.; Wongkaew, S.; Teaumroong, N.; Boonkerd, N.; Nomura, N.; Tittabutr, P. Improvement of peanut rhizobial inoculant by incorporation of plant growth promoting rhizobacteria (PGPR) as biocontrol against the seed borne fungus, Aspergillus niger. Biol. Control. 2012, 63, 87–97. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Strain ID | Accession Number | Length (bp) | Coverage (%) | Identity (%) | Closest Taxon (Accession Number) |
|---|---|---|---|---|---|
| BGH2-7 | MT256074 | 1501 | 99 | 95.56 | Bacillus vallismortis (FJ386541) |
| BGH4-1 | MW002558 | 559 | 100 | 96.08 | B. halotolerans (MT271912) |
| G1B | MT256075 | 1477 | 99 | 97.35 | B. subtilis (KP876486) |
| G2B | MT256077 | 1495 | 99 | 96.45 | B. halotolerans (MF417800) |
| G3C | MT256076 | 1483 | 99 | 96.46 | B. amyloliquefaciens (PP125657) |
| G1D | MW002221 | 1019 | 99 | 97.84 | B. subtilis (KY818937) |
| G3F | MT254817 | 1524 | 99 | 94.72 | B. subtilis (OM978656) |
| G3D | MT256072 | 1500 | 98 | 95.29 | B. subtilis (KY652939) |
| BGH2-3 | MW086541 | 536 | 99 | 95.86 | Enterobacter sp. (JX103562) |
| BGH1-5 | MW092092 | 1000 | 100 | 93.47 | Kosakonia cowanii MG871199 |
| BGH2-5 * | MT254758 | 1513 | 98 | 94.16 | Pantoea agglomerans MZ647535 |
| BGH1-6 * | MT254751 | 576 | 100 | 100 | P. agglomerans (OQ202156) |
| BGH2-1 | MT254818 | 903 | 98 | 91.12 | P. conspicua (MW568057) |
| G4A | MW092005 | 555 | 99 | 99.29 | Pantoea sp. (JN853255) |
| G1A | MW079530 | 539 | 100 | 99.63 | Pseudomonas azotoformans (MK883209) |
| BGH1-3 | MW079843 | 559 | 100 | 97.50 | Serratia liquefaciens (MN326772) |
| BGH2-2 | MW008870 | 301 | 99 | 97.00 | S. nematodiphila (MH669373) |
| G2C | MW008604 | 645 | 99 | 96.57 | S. nematodiphila (MN691578) |
| Bacterial Isolates | Leptopeptide-Encoding Genes | |||
|---|---|---|---|---|
| BamC | Itup | FenD | Sfp | |
| G1A | + | + | + | - |
| G2C | + | - | + | + |
| G1D * | + | + | + | + |
| G3F | + | + | - | + |
| G1B * | + | + | + | + |
| BGH 1-5 | + | - | + | - |
| BGH2-1 | + | + | + | + |
| G3C | - | + | + | - |
| BGH4-1 | - | + | + | + |
| G2B | - | + | + | + |
| BGH2-3 | + | - | + | - |
| G4A | + | + | + | - |
| G3D | + | - | + | + |
| BGH 2-7 | + | + | + | - |
| BGH 1-3 * | + | + | + | + |
| BGH1-6 | + | + | - | - |
| BGH2-2 * | + | + | + | + |
| BGH2-5 | + | + | + | - |
| Bacteria Isolates | ICLa | IPC a | IPR a | ISP a | IAM a | ICH a | AIA | HCN |
|---|---|---|---|---|---|---|---|---|
| G2c | 0 | 1.37 ± 0.08 | 4.12 ± 0.28 | 1.02 ± 0.16 | 1.26 ± 0.06 | 0 | +++ | ++ |
| BGH 1-5 | 0 | 0 | 1.17 ± 0.41 | 1.49 ± 0.16 | 0 | 5.21 ± 0.71 | +++ | − |
| BGH 2-1 | 0 | 0 | 0 | 0 | 0 | 1.94 ± 0.47 | +++ | +++ |
| BGH 2-5 | 0 | 0 | 0 | 1.01 ± 0.43 | 0 | 0 | ++ | + |
| BGH 1-6 | 1.5 ± 0.04 | 1.98 ± 0.79 | 0 | 1.16 ± 0.62 | 0 | 0 | + | + |
| G3f | 0 | 0 | 3.76 ± 0.22 | 1.37 ± 0.08 | 0 | 0 | + | − |
| G2b | 0 | 0 | 0 | 1.40 ± 0.12 | 0 | 0 | + | ++ |
| BGH 2-2 | 0 | 0 | 1.21 ± 0.46 | 1.25 ± 0.19 | 0 | 0 | + | ++ |
| BGH 1-3 | 1.61 ± 0.09 | 1.36 ± 0.34 | 4.12 ± 0.30 | 0 | 1.28 ± 0.04 | 1.53 ± 0.01 | + | +++ |
| G3d | 1.57 ± 0.06 | 0 | 3.09 ± 1.45 | 0 | 0 | 0 | + | + |
| G4a | 1.54 ± 0.01 | 1.58 ± 0.11 | 1.82 ± 0.10 | 0 | 1.14 ± 0.00 | 0 | + | + |
| BGH 2-7 | 0 | 2.03 ± 0.24 | 5.0 ± 0.21 | 1.41 ± 0.26 | 1.06 ± 0.00 | 0 | + | ++ |
| G1b | 0 | 0 | 0 | 0 | 1.19 ± 0.01 | 0 | − | − |
| BGH 2-3 | 0 | 0 | 0 | 1.55 ± 0.01 | 0 | 0 | − | + |
| BGH 4-1 | 0 | 0 | 4.85 ± 0.10 | 1.14 ± 0.20 | 1.24 ± 0.05 | 0 | − | +++ |
| G1d | 0 | 0 | 0 | 1.19 ± 0.47 | 0 | 0 | − | + |
| G1a | 0 | 0 | 0 | 0 | 0 | 0 | − | + |
| G3c | 1.62 ± 0.05 | 0 | 1.42 ± 0.11 | 0 | 0 | 0 | − | − |
| Treatments | Identity (Closest BLAST Match) a | AUDPC b | Efficiency (%) | Significance |
|---|---|---|---|---|
| BGH 2-7 | Bacillus vallismortis | 10.31 ± 0.12 | 77.42% | b |
| BGH 1-3 | Serratia liquefaciens | 11.94 ± 0.14 | 73.86% | b |
| BGH 2-2 | Serratia nematodiphila | 17.61 ± 0.24 | 61.45% | c |
| BGH 1-6 | Pantoea agglomerans | 10.91 ± 0.33 | 76.10% | b |
| Difenoconazole | 5.25 ± 0.43 | 88.51% | a | |
| Control | 45.68 ± 0.96 | d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Housni, Z.; Ezrari, S.; Radouane, N.; Tahiri, A.; Ouijja, A.; Errafii, K.; Hijri, M. Evaluating Rhizobacterial Antagonists for Controlling Cercospora beticola and Promoting Growth in Beta vulgaris. Microorganisms 2024, 12, 668. https://doi.org/10.3390/microorganisms12040668
El Housni Z, Ezrari S, Radouane N, Tahiri A, Ouijja A, Errafii K, Hijri M. Evaluating Rhizobacterial Antagonists for Controlling Cercospora beticola and Promoting Growth in Beta vulgaris. Microorganisms. 2024; 12(4):668. https://doi.org/10.3390/microorganisms12040668
Chicago/Turabian StyleEl Housni, Zakariae, Said Ezrari, Nabil Radouane, Abdessalem Tahiri, Abderrahman Ouijja, Khaoula Errafii, and Mohamed Hijri. 2024. "Evaluating Rhizobacterial Antagonists for Controlling Cercospora beticola and Promoting Growth in Beta vulgaris" Microorganisms 12, no. 4: 668. https://doi.org/10.3390/microorganisms12040668
APA StyleEl Housni, Z., Ezrari, S., Radouane, N., Tahiri, A., Ouijja, A., Errafii, K., & Hijri, M. (2024). Evaluating Rhizobacterial Antagonists for Controlling Cercospora beticola and Promoting Growth in Beta vulgaris. Microorganisms, 12(4), 668. https://doi.org/10.3390/microorganisms12040668