

The Impact of Early-Life Cecal Microbiota Transplantation on Social Stress and Injurious Behaviors in Egg-Laying Chickens

 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Genetic Lines and Study Design
2.1. Unique Production, Biology, and Behavior between the Divergently Selected Inbred Lines
2.2. Study Design and Results
2.2.1. Trial 1 [112]
2.2.2. Trial 2 [106]
2.2.3. Trial 3 [123]
3. Cecal Microbiota Transplantation, Social Stress, and Injurious Behavior in Chickens
3.1. Stress and Gut Microbiota
3.2. Possible Pathophysiological Mechanisms Underlying Injurious Behaviors in Chickens
3.3. Physiological Mechanisms of Modulation of Intestinal Microbiota to Regulate Social Stress and Related Abnormal Behaviors
3.4. Cecal Microbiota Transplantation and Injurious Behavior in Chickens
4. Conclusions and Perspectives
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Henchion, M.; Moloney, A.P.; Hyland, J.; Zimmermann, J.; McCarthy, S. Review: Trends for meat, milk and egg consumption for the next decades and the role played by livestock systems in the global production of proteins. Animal 2021, 15, 100287. [Google Scholar] [CrossRef] [PubMed]
- USDA ERS (Economic Research Service, U.S. Department of Agriculture). Poultry and Eggs, Market Outlook. Available online: https://www.ers.usda.gov/topics/animal-products/poultry-eggs/market-outlook (accessed on 30 November 2023).
- Hagelso, A.M.; Krohn, C.C. Quantifing social behavior of the individual. In Animal Genetic Resources for Adaptation to More Extensive Production Systems; Hagelso, A.M., Ed.; Commission of the European Communities: Foulum, Denmark, 1993. [Google Scholar]
- Rothschild, J. Ethical considerations of gene editing and genetic selection. J. Gen. Fam. Med. 2020, 21, 37–47. [Google Scholar] [CrossRef]
- Muir, W.M.; Cheng, H.W.; Coney, C. New selection methods for layer performance and potential impacts on behavior and management. In Proceedings of the XIVth European Poultry Conference, Stavanger, Norway, 23–27 June 2014. [Google Scholar]
- Baldauf, S.; Engqvist, L.; Weissing, F. Diversifying evolution of competitiveness. Nat. Commun. 2014, 5, 5233. [Google Scholar] [CrossRef]
- Bernhardt, J.R.; Kratina, P.; Pereira, A.L.; Tamminen, M.; Thomas, M.K.; Narwani, A. The evolution of competitive ability for essential resources. Philos. Trans. R. Soc. 2020, B375, 20190247. [Google Scholar] [CrossRef] [PubMed]
- Grether, G.F.; Anderson, C.N.; Drury, J.P.; Kirschel, A.N.; Losin, N.; Okamoto, K.; Peiman, K.S. The evolutionary consequences of interspecific aggression. Ann. N. Y. Acad. Sci. 2013, 1289, 48–68. [Google Scholar] [CrossRef] [PubMed]
- Hocking, P.M. Unexpected consequences of genetic selection in broilers and turkeys: Problems and solutions. Br. Poult. Sci. 2014, 55, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Drury, J.P.; Cowen, M.C.; Grether, G.F. Competition and Hybridization Drive Interspecific Territoriality in Birds. Proc. Natl. Acad. Sci. USA 2020, 117, 12923–12930. [Google Scholar] [CrossRef] [PubMed]
- Muir, W.M. Group Selection for Adaptation to Multiple-Hen Cages: Selection Program and Direct Responses. Poult. Sci. 1996, 75, 447–458. [Google Scholar] [CrossRef]
- Biro, P.A.; Stamps, J.A. Are animal personality traits linked to life-history productivity? Trends Ecol. Evol. 2008, 23, 361–368. [Google Scholar] [CrossRef]
- Senner, N.R.; Conklin, J.R.; Piersma, T. An ontogenetic perspective on individual differences. Proc. Biol. Sci. 2015, 282, 20151050. [Google Scholar] [CrossRef]
- Moore, M.P.; Martin, R.A. On the evolution of carry-over effects. J. Anim. Ecol. 2019, 88, 1832–1844. [Google Scholar] [CrossRef] [PubMed]
- Chunduri, A.; Reddy, S.D.M.; Jahanavi, M.; Reddy, C.N. Gut–Brain Axis, Neurodegeneration and Mental Health: A Personalized Medicine Perspective. Indian J. Microbiol. 2022, 62, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Mench, J.A.; Duncan, I.J. Poultry welfare in North America: Opportunities and challenges. Poult. Sci. 1998, 77, 1763–1765. [Google Scholar] [CrossRef]
- Cheng, H.-W. Animal Welfare: Should We Change Housing to Better Accommodate the Animal or Change the Animal to Accommodate the Housing? CABI Rev. 2007, 2, 14. [Google Scholar] [CrossRef]
- Lin, E.-J.D.; Sun, M.; Choi, E.Y.; Magee, D.; Stets, C.W.; During, M.J. Social Overcrowding as a Chronic Stress Model That Increases Adiposity in Mice. Psychoneuroendocrinology 2015, 51, 318–330. [Google Scholar] [CrossRef]
- Banich, M.T.; Floresco, S. Reward systems, cognition, and emotion: Introduction to the special issue. Cogn. Affect. Behav. Neurosci. 2019, 19, 409–414. [Google Scholar] [CrossRef]
- Kern, E.M.A.; Robinson, D.; Gass, E.; Godwin, J.; Langerhans, R.B. Correlated evolution of personality, morphology and performance. Anim. Behav. 2016, 117, 79–86. [Google Scholar] [CrossRef]
- De Boer, S.F. Animal models of excessive aggression: Implications for human aggression and violence. Curr. Opin. Psychol. 2018, 19, 81–87. [Google Scholar] [CrossRef]
- Flanigan, M.E.; Russo, S.J. Recent advances in the study of aggression. Neuropsychopharmacology 2019, 44, 241–244. [Google Scholar] [CrossRef]
- Kleszcz, A.; Cholewińska, P.; Front, G.; Pacoń, J.; Bodkowski, R.; Janczak, M.; Dorobisz, T. Review on Selected Aggression Causes and the Role of Neurocognitive Science in the Diagnosis. Animals 2022, 12, 281. [Google Scholar] [CrossRef]
- Lay, D.C., Jr.; Fulton, R.M.; Hester, P.Y.; Karcher, D.M.; Kjaer, J.B.; Mench, J.A.; Mullens, B.A.; Newberry, R.C.; Nicol, C.J.; O’Sullivan, N.P.; et al. Hen welfare in different housing systems. Poult. Sci. 2011, 90, 278–294. [Google Scholar] [CrossRef] [PubMed]
- van Staaveren, N.; Harlander, A. Cause and Prevention of Injurious Pecking in Chickens. In Understanding the Behaviour and Improving the Welfare of Chickens; Burleigh Dodds Science Publishing: Cambridge, UK, 2020; pp. 509–566. [Google Scholar]
- Meuser, V.; Weinhold, L.; Hillemacher, S.; Tiemann, I. Welfare-Related Behaviors in Chickens: Characterization of Fear and Exploration in Local and Commercial Chicken Strains. Animals 2021, 11, 679. [Google Scholar] [CrossRef] [PubMed]
- Van der Eijk, J.A.; de Vries, H.; Kjaer, J.B.; Naguib, M.; Kemp, B.; Smidt, H.; Rodenburg, T.B.; Lammers, A. Differences in gut microbiota composition of laying hen lines divergently selected on feather pecking. Poult. Sci. 2019, 98, 7009–7021. [Google Scholar] [CrossRef]
- Van der Eijk, J.A.; Rodenburg, T.B.; de Vries, H.; Kjaer, J.B.; Smidt, H.; Naguib, M.; Kemp, B.; Lammers, A. Early-life microbiota transplantation affects behavioural responses, serotonin and immune characteristics in chicken lines divergently selected on feather pecking. Sci. Rep. 2020, 10, 2750. [Google Scholar] [CrossRef] [PubMed]
- Iffland, H.; Wellmann, R.; Preuß, S.; Tetens, J.; Bessei, W.; Piepho, H.P.; Bennewitz, J. A novel model to explain extreme feather pecking behavior in laying hens. Behav. Genet. 2020, 50, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Cronin, G.; Hopcroft, R.; Groves, P.; Hall, E.; Phalen, D.; Hemsworth, P. Why Did Severe Feather Pecking and Cannibalism Outbreaks Occur? An Unintended Case Study While Investigating the Effects of Forage and Stress on Pullets during Rearing. Poult. Sci. 2018, 97, 1484–1502. [Google Scholar] [CrossRef]
- Ellen, E.D.; Van Der Sluis, M.; Siegford, J.; Guzhva, O.; Toscano, M.J.; Bennewitz, J.; Van Der Zande, L.E.; Van Der Eijk, J.A.; de Haas, E.N.; Norton, T.; et al. Review of sensor technologies in animal breeding: Phenotyping behaviors of laying hens to select against feather pecking. Animals 2019, 9, 108. [Google Scholar] [CrossRef]
- Falker-Gieske, C.; Mott, A.; Preuß, S.; Franzenburg, S.; Bessei, W.; Bennewitz, J.; Tetens, J. Analysis of the brain transcriptome in lines of laying hens divergently selected for feather pecking. BMC Genom. 2020, 21, 595. [Google Scholar] [CrossRef]
- Falker-Gieske, C.; Iffland, H.; Preuß, S.; Bessei, W.; Drögemüller, C.; Bennewitz, J.; Tetens, J. Meta-analyses of genome wide association studies in lines of laying hens divergently selected for feather pecking using imputed sequence level genotypes. BMC Genet. 2020, 21, 114. [Google Scholar] [CrossRef]
- Baker, P.E.; Nicol, C.J.; Weeks, C.A. The effect of hard pecking enrichment during rear on feather cover, feather pecking behaviour and beak length in beak-trimmed and intact-beak laying hen pullets. Animals 2022, 12, 674. [Google Scholar] [CrossRef]
- BFREPA (the British Free Range Egg Producers Association). Egg Industry Organizations Have Joined Forces to Fund Vital Research into the Latest Beak Treatment Technique. 2006. Available online: http://www.theranger.co.uk (accessed on 30 November 2023).
- Poultrysite. Pecking—The Unanswered Question. 2010. Available online: https://www.farminguk.com/news/pecking-the-unanswered-questions_432.html (accessed on 30 November 2023).
- Piepho, H.P.; Lutz, V.; Kjaer, J.B.; Grashorn, M.; Bennewitz, J.; Bessei, W. The presence of extreme feather peckers in groups of laying hens. Animal 2017, 11, 500–506. [Google Scholar] [CrossRef]
- RSPCA (The Royal Society for the Prevention of Cruelty to Animals). How Can Feather Pecking Be Managed in Cage-Free Layer Hen Systems? 2023. Available online: https://kb.rspca.org.au/knowledge-base/how-can-feather-pecking-be-managed-in-cage-free-layer-hen-systems/ (accessed on 30 November 2023).
- Cheng, H.W. Morphopathological changes and pain in beak trimming laying hens. World’s Poult. Sci. J. 2005, 62, 41–52. [Google Scholar] [CrossRef]
- Mikoni, N.A.; Guzman, D.S.; Fausak, E.; Paul-Murphy, J. Recognition and assessment of pain-related behaviors in avian species: An integrative review. J. Avian Med. Surg. 2022, 36, 153–172. [Google Scholar] [CrossRef]
- NASS (The National Agricultural Statistics Service). Chickens and Eggs. 2022. Available online: https://www.nass.usda.gov/Publications/Todays_Reports/reports/ckeg0422.pdf (accessed on 30 November 2023).
- Shahbandeh, M. Total Number of Laying Hens in the U.S. 2000–2022. Statista. 2023. Available online: https://www.statista.com/statistics/195823/total-number-of-laying-hens-in-the-us-since-2000/ (accessed on 30 November 2023).
- Iqbal, A.; Moss, A.F. Review: Key tweaks to the chicken’s beak: The versatile use of the beak by avian species and potential approaches for improvements in poultry production. Animal 2021, 15, 100119. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Yoshida, S.; Matsuyama, H.; Obi, T.; Takase, K. Morphologically abnormal beaks observed in chickens that were beak-trimmed at young ages. J. Vet. Med. Sci. 2017, 79, 1466–1471. [Google Scholar] [CrossRef]
- Coton, J.; Guinebretière, M.; Guesdon, V.; Chiron, G.; Mindus, C.; Laravoire, A.; Pauthier, G.; Balaine, L.; Descamps, M.; Bignon, L.; et al. Feather pecking in laying hens housed in free-range or furnished-cage systems on French farms. Br. Poult. Sci. 2019, 60, 617–627. [Google Scholar] [CrossRef]
- Elson, A. Laying Hens Beaks: To Trim or Not to Trim. Livestock Knowledge Transfer, a DEFRA Initiative Operated by ADAS/IGER/University of Bristol. Poultry 2001. Available online: http://www.agrowebcee.net/fileadmin/content/faw/doc/reports2/BEAK_TRIMMING_REVIEW.pdf (accessed on 30 November 2023).
- Rudkin, C. Feather pecking and foraging uncorrelated—The redirection hypothesis revisited. Br. Poult. Sci. 2022, 63, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Kops, M.S.; de Haas, E.N.; Rodenburg, T.B.; Ellen, E.D.; Korte-Bouws, G.A.; Olivier, B.; Güntürkün, O.; Bolhuis, J.E.; Korte, S.M. Effects of feather pecking phenotype (severe feather peckers, victims and non-peckers) on serotonergic and dopaminergic activity in four brain areas of laying hens (Gallus gallus domesticus). Physiol. Behav. 2013, 120, 77–82. [Google Scholar] [CrossRef]
- García-Cabrerizo, R.; Carbia, C.; O’ Riordan, K.J.; Schellekens, H.; Cryan, J.F. Microbiota-gut-brain axis as a regulator of reward processes. J. Neurochem. 2021, 157, 1495–1524. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Chen, T.; Green, S.J.; Mutlu, E.; Martin, B.R.; Rumpagaporn, P.; Patterson, J.A.; Keshavarzian, A.; Hamaker, B.R. Physical inaccessibility of a resistant starch shifts mouse gut microbiota to butyrogenic firmicutes. Mol. Nutr. Food Res. 2019, 63, e1801012. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Mu, C.-L.; Farzi, A.; Zhu, W.-Y. Tryptophan metabolism: A link between the gut microbiota and brain. Adv. Nutr. 2020, 11, 709–723. [Google Scholar] [CrossRef] [PubMed]
- Rea, K.; Dinan, T.G.; Cryan, J.F. The microbiome: A key regulator of stress and neuroinflammation. Neurobiol. Stress 2016, 4, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Gwak, M.G.; Chang, S.Y. Gut-Brain Connection: Microbiome, Gut Barrier, and Environmental Sensors. Immune Netw. 2021, 21, e20. [Google Scholar] [CrossRef]
- Barton, J.R.; Londregan, A.K.; Alexander, T.D.; Entezar, A.A.; Covarrubias, M.; Waldman, S.A. Enteroendocrine cell regulation of the gut-brain axis. Front. Neurosci. 2023, 17, 1272955. [Google Scholar] [CrossRef]
- Yarandi, S.S.; Peterson, D.A.; Treisman, G.J.; Moran, T.H.; Pasricha, P.J. Modulatory effects of gut microbiota on the central nervous system: How gut could play a role in neuropsychiatric health and Diseases. J. Neurogastroenterol. Motil. 2016, 22, 201–212. [Google Scholar] [CrossRef]
- Dicks, L.M.T. Gut bacteria and neurotransmitters. Microorganisms 2022, 10, 1838. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Shim, J.O. Gut microbiota affects brain development and behavior. Clin. Exp. Pediatr. 2022, 66, 274–280. [Google Scholar] [CrossRef]
- Homer, B.; Judd, J.; Mohammadi Dehcheshmeh, M.; Ebrahimie, E.; Trott, D.J. Gut microbiota and behavioural issues in production, performance, and companion animals: A systematic review. Animals 2023, 13, 1458. [Google Scholar] [CrossRef]
- Tan, H.E. The microbiota-gut-brain axis in stress and depression. Front. Neurosci. 2023, 17, 1151478. [Google Scholar] [CrossRef]
- Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Ikeda, Y.; Tsuji, A.; Matsuda, S. A new concept of associations between gut microbiota, immunity and central nervous system for the innovative treatment of neurodegenerative disorders. Metabolites 2022, 12, 1052. [Google Scholar] [CrossRef] [PubMed]
- Marano, G.; Mazza, M.; Lisci, F.M.; Ciliberto, M.; Traversi, G.; Kotzalidis, G.D.; De Berardis, D.; Laterza, L.; Sani, G.; Gasbarrini, A.; et al. The Microbiota-Gut-Brain axis: Psychoneuroimmunological insights. Nutrients 2023, 15, 1496. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Luo, S.; Yan, C. Gut microbiota implications for health and welfare in farm animals, A review. Animals 2021, 12, 93. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wu, H.; Chen, Z.; Hao, H.; Zheng, X. Gut microbiome at the crossroad of genetic variants and behavior disorders. Gut Microbes 2023, 15, 2201156. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, M.A.H.; Ashrafudoulla, M.; Mevo, S.I.U.; Mizan, M.F.R.; Park, S.H.; Ha, S.D. Current and future interventions for improving poultry health and poultry food safety and security: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2023, 22, 1555–1596. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, L.; Feng, Y.; Liu, Y.-F.; Si, T.L.; Su, Z.; Cheung, T.; Ungvari, G.S.; Ng, C.H.; Xiang, Y.-T. Brain-gut axis and psychiatric disorders: A perspective from bibliometric and visual analysis. Front. Immunol. 2022, 13, 1047007. [Google Scholar] [CrossRef] [PubMed]
- Demin, K.A.; Zabegalov, K.A.; Kolesnikova, T.O.; Galstyan, D.S.; Kositsyn, Y.M.; Costa, F.V.; De Abreu, M.S.; Kalueff, A.V. Animal Inflammation-Based Models of Neuropsychiatric Disorders. In Neuroinflammation, Gut-Brain Axis and Immunity in Neuropsychiatric Disorders; Springer Nature: Singapore, 2023; pp. 91–104. [Google Scholar]
- Mitrea, L.; Nemeş, S.A.; Szabo, K.; Teleky, B.E.; Vodnar, D.C. Guts Imbalance Imbalances the Brain: A Review of Gut Microbiota Association with Neurological and Psychiatric Disorders. Front. Med. 2022, 9, 813204. [Google Scholar] [CrossRef] [PubMed]
- Carloni, S.; Rescigno, M. The gut-brain vascular axis in neuroinflammation. Semin. Immunol. 2023, 69, 101802. [Google Scholar] [CrossRef]
- Cheng, J.; Lei, H.; Xie, C.; Chen, J.; Yi, X.; Zhao, F.; Yuan, Y.; Chen, P.; He, J.; Luo, C.; et al. B lymphocyte development in the bursa of fabricius of young broilers is influenced by the gut microbiota. Microbiol. Spectr. 2023, 11, e0479922. [Google Scholar] [CrossRef]
- Huang, C.; Hao, E.; Yue, Q.; Liu, M.; Wang, D.; Chen, Y.; Shi, L.; Zeng, D.; Zhao, G.; Chen, H. Malfunctioned inflammatory response and serotonin metabolism at the microbiota-gut-brain axis drive feather pecking behavior in laying hens. Poult. Sci. 2023, 102, 102686. [Google Scholar] [CrossRef]
- Li, Y.; Yang, L.; Li, J.; Gao, W.; Zhao, Z.; Dong, K.; Duan, W.; Dai, B.; Guo, R. Antidepression of Xingpijieyu formula targets gut microbiota derived from depressive disorder. CNS Neurosci. Ther. 2023, 29, 669–681. [Google Scholar] [CrossRef]
- Kim, J.E.; Tun, H.M.; Bennett, D.C.; Leung, F.C.; Cheng, K.M. Microbial diversity and metabolic function in duodenum, jejunum and ileum of emu (Dromaius novaehollandiae). Sci. Rep. 2023, 13, 4488. [Google Scholar] [CrossRef]
- Koyasu, H.; Takahashi, H.; Yoneda, M.; Naba, S.; Sakawa, N.; Sasao, I.; Nagasawa, M.; Kikusui, T. Correlations between behavior and hormone concentrations or gut microbiome imply that domestic cats (Felis silvestris catus) living in a group are not like ‘groupmates’. PLoS ONE 2022, 17, e0269589. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, S.A.L.; Ton, A.M.M.; Pereira, T.M.C.; Campagnaro, B.P.; Martinelli, L.; Picos, A.; Campos-Toimil, M.; Vasquez, E.C. The gut microbiota-brain axis: A new frontier on neuropsychiatric disorders. Front. Psychiatry 2022, 13, 872594. [Google Scholar] [CrossRef] [PubMed]
- Accettulli, A.; Corbo, M.R.; Sinigaglia, M.; Speranza, B.; Campaniello, D.; Racioppo, A.; Altieri, C.; Bevilacqua, A. Psycho-microbiology, a new frontier for probiotics: An exploratory overview. Microorganisms 2022, 10, 2141. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, N.Y.; Jalgaonkar, M.P.; Hargude, A.B.; Sherje, A.P.; Oza, M.J.; Doshi, G.M. Gut-Brain axis and neurological disorders-how microbiomes affect our mental health. CNS Neurol. Disord. -Drug Targets 2023, 22, 1008–1030. [Google Scholar] [CrossRef] [PubMed]
- Garvey, M. The association between dysbiosis and neurological conditions often manifesting with chronic pain. Biomedicines 2023, 11, 748. [Google Scholar] [CrossRef]
- Handajani, Y.S.; Hengky, A.; Schröder-Butterfill, E.; Hogervorst, E.; Turana, Y. Probiotic supplementation improved cognitive function in cognitively impaired and healthy older adults: A systematic review of recent trials. Neurol. Sci. 2023, 44, 1163–1169. [Google Scholar] [CrossRef]
- Johnson, D.; Letchumanan, V.; Thum, C.C.; Thurairajasingam, S.; Lee, L.H. A microbial-based approach to mental health: The potential of probiotics in the treatment of depression. Nutrients 2023, 15, 1382. [Google Scholar] [CrossRef]
- Kim, I.B.; Park, S.C.; Kim, Y.K. Microbiota-Gut-Brain axis in major depression: A new therapeutic approach. Adv. Exp. Med. Biol. 2023, 1411, 209–224. [Google Scholar]
- Mohan, A.; Godugu, S.; Joshi, S.S.; Shah, K.B.; Vanka, S.C.; Shakil, H.; Dhanush, P.; Veliginti, S.; Sure, P.S.; Goranti, J. Gut-brain axis: Altered microbiome and depression—Review. Ann. Med. Surg. 2023, 85, 1784–1789. [Google Scholar]
- Rathour, D.; Shah, S.; Khan, S.; Singh, P.K.; Srivastava, S.; Singh, S.B.; Khatri, D.K. Role of gut microbiota in depression: Understanding molecular pathways, recent research, and future direction. Behav. Brain Res. 2023, 436, 114081. [Google Scholar] [CrossRef] [PubMed]
- Sikorska, M.; Antosik-Wójcińska, A.Z.; Dominiak, M. Probiotics as a tool for regulating molecular mechanisms in depression: A systematic review and meta-analysis of randomized clinical trials. Int. J. Mol. Sci. 2023, 24, 3081. [Google Scholar] [CrossRef] [PubMed]
- Varesi, A.; Campagnoli, L.I.M.; Chirumbolo, S.; Candiano, B.; Carrara, A.; Ricevuti, G.; Esposito, C.; Pascale, A. The brain-gut-microbiota interplay in depression: A key to design innovative therapeutic approaches. Pharmacol. Res. 2023, 192, 106799. [Google Scholar] [CrossRef]
- Hashimoto, K. Emerging role of the host microbiome in neuropsychiatric disorders: Overview and future directions. Mol. Psychiatry. 2023, 28, 3625–3637. [Google Scholar] [CrossRef] [PubMed]
- Dinan, K.; Dinan, T.G. Gut microbes and neuropathology: Is there a causal nexus? Pathogens 2022, 11, 796. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.V.; Watson, K.K.; Dunbar, R.I.M.; Burnet, P.W.J. Sociability in a non-captive macaque population is associated with beneficial gut bacteria. Front. Microbiol. 2022, 13, 1032495. [Google Scholar] [CrossRef]
- Clench, M.H.; Mathias, J.R. The Avian cecum: A review. Wilson Bull. 1995, 107, 93–121. [Google Scholar]
- Stanley, D.; Geier, M.S.; Chen, H.; Hughes, R.J.; Moore, R.J. Comparison of fecal and cecal microbiotas reveals qualitative similarities but quantitative differences. BMC Microbiol. 2015, 15, 51. [Google Scholar] [CrossRef]
- Hunt, A.; Al-Nakkash, L.; Lee, A.H.; Smith, H.F. Phylogeny and herbivory are related to avian cecal size. Sci. Rep. 2019, 9, 4243. [Google Scholar] [CrossRef]
- Di Marcantonio, L.; Marotta, F.; Vulpiani, M.P.; Sonntag, Q.; Iannetti, L.; Janowicz, A.; Di Serafino, G.; Di Giannatale, E.; Garofolo, G. Investigating the cecal microbiota in broiler poultry farms and its potential relationships with animal welfare. Res. Vet. Sci. 2022, 144, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Campos, P.M.; Schreier, L.L.; Proszkowiec-Weglarz, M.; Dridi, S. Cecal microbiota composition differs under normal and high ambient temperatures in genetically distinct chicken lines. Sci. Rep. 2023, 13, 16037. [Google Scholar] [CrossRef]
- Cazals, A.; Estellé, J.; Bruneau, N.; Coville, J.L.; Menanteau, P.; Rossignol, M.N.; Jardet, D.; Bevilacqua, C.; Rau, A.; Bed’Hom, B.; et al. Differences in caecal microbiota composition and Salmonella carriage between experimentally infected inbred lines of chickens. Genet. Sel. Evol. 2022, 54, 7. [Google Scholar] [CrossRef]
- Van der Eijk, J.A.; Lammers, A.; Kjaer, J.B.; Rodenburg, T.B. Stress response, peripheral serotonin and natural antibodies in feather pecking genotypes and phenotypes and their relation with coping style. Physiol. Behav. 2019, 199, 1–10. [Google Scholar] [CrossRef]
- Van der Eijk, J.A.; Verwoolde, M.B.; de Vries Reilingh, G.; Jansen, C.A.; Rodenburg, T.B.; Lammers, A. Chicken lines divergently selected on feather pecking differ in immune characteristics. Physiol. Behav. 2019, 212, 112680. [Google Scholar] [CrossRef]
- Meyer, B.; Zentek, J.; Harlander-Matauschek, A. Differences in intestinal microbial metabolites in laying hens with high and low levels of repetitive feather-pecking behavior. Physiol. Behav. 2013, 110, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, Y.; Wang, H.; Li, M.; Rong, J.; Liao, X.; Wu, Y.; Wang, Y. Differences in peripheral and central metabolites and gut microbiome of laying hens with different feather-pecking phenotypes. Front. Microbiol. 2023, 14, 1132866. [Google Scholar] [CrossRef]
- Borda-Molina, D.; Iffland, H.; Schmid, M.; Müller, R.; Schad, S.; Seifert, J.; Tetens, J.; Bessei, W.; Bennewitz, J.; Camarinha-Silva, A. Gut microbial composition and predicted functions are not associated with feather pecking and antagonistic behavior in laying hens. Life 2021, 11, 235. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.A. Possibilities of early life programming in broiler chickens via intestinal microbiota modulation. Poult. Sci. 2019, 98, 695–706. [Google Scholar] [CrossRef]
- Bacon, L.D.; Hunt, H.D.; Cheng, H.H. Genetic resistance to Marek’s disease. Curr. Top. Microbiol. Immunol. 2001, 255, 121–141. [Google Scholar]
- Boodhoo, N.; Gurung, A.; Sharif, S.; Behboudi, S. Marek’s disease in chickens: A review with focus on immunology. Vet. Res. 2016, 47, 119. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; He, Y.; Ding, Y.; Liu, G.E.; Zhang, H.; Cheng, H.H.; Taylor, R.L.; Song, J. Genetic assessment of inbred chicken lines indicates genomic signatures of resistance to Marek’s disease. J. Anim. Sci. Biotechnol. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dennis, R.L.; Zhang, H.W.; Bacon, L.D.; Estevez, I.; Cheng, H.W. Behavioral and physiological features of chickens diversely selected for resistance to avian disease: I. Selected inbred lines differ for behavioral and physical responses to social stress. Poult. Sci. 2004, 83, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Dennis, R.L.; Zhang, H.M.; Cheng, H.W. Effect of selection for resistance and susceptibility to viral diseases on concentrations of dopamine and immunological parameters in six-week-old chickens. Poult. Sci. 2006, 85, 2135–2140. [Google Scholar] [CrossRef]
- Dennis, R.L.; Cheng, H.W. Differential serotonergic mediation of aggression in roosters bred for resistance and susceptibility to Marek’s disease. Br. Poult. Sci. 2014, 55, 13–20. [Google Scholar] [CrossRef]
- Fu, Y.; Hu, J.Y.; Erasmus, M.; Johnson, T.; Cheng, H.W. Effects of early-life cecal microbiota transplantation from divergently selected inbred chicken lines on growth, gut serotonin, and immune parameters in recipient chickens. Poult. Sci. 2022, 101, 101925. [Google Scholar] [CrossRef]
- Bacon, L.D.; Hunt, H.D.; Cheng, H.H. A review of the development of chicken lines to resolve genes determining resistance to diseases. Poult. Sci. 2000, 79, 1082–1093. [Google Scholar] [CrossRef] [PubMed]
- Bacon, L.D.; Palmquist, D. Chicken lines differ in production of interferon-like activity by peripheral white blood cells stimulated with phytohemagglutinin. Poult. Sci. 2002, 81, 1629–1636. [Google Scholar] [CrossRef]
- Yonash, N.I.; Bacon, L.D.; Smith, E.J. Concentration of immunoglobulin G in plasma varies among 6C. 7 recombinant congenic strains of chickens. Poult. Sci. 2002, 81, 1104–1108. [Google Scholar] [CrossRef]
- Warren, W.C.; Rice, E.S.; Meyer, A.; Hearn, C.J.; Steep, A.; Hunt, H.D.; Monson, M.S.; Lamont, S.J.; Cheng, H.H. The immune cell landscape and response of Marek’s disease resistant and susceptible chickens infected with Marek’s disease virus. Sci. Rep. 2023, 13, 5355. [Google Scholar] [CrossRef]
- Dennis, R.L.; Muir, M.W.; Cheng, H.W. Effects of raclopride on aggression and stress in diversely selected chicken lines. Behav. Brain Res. 2006, 175, 104–111. [Google Scholar] [CrossRef]
- Hu, J.Y.; Johnson, T.A.; Zhang, H.; Cheng, H.W. The microbiota-gut-brain axis: Gut microbiota modulates conspecific aggression in diversely selected laying hens. Microorganisms 2022, 10, 1081. [Google Scholar] [CrossRef]
- Burgess, S.C.; Basaran, B.H.; Davison, T.F. Resistance to marek’s disease herpesvirus-induced lymphoma is multiphasic and dependent on host genotype. Vet. Pathol. 2002, 38, 129–142. [Google Scholar] [CrossRef]
- Perumbakkam, S.; Hunt, H.D.; Cheng, H.H. Differences in CD8αα and cecal microbiome community during proliferation and late cytolytic phases of Marek’s disease virus infection are associated with genetic resistance to Marek’s disease. FEMS Microbiol. Ecol. 2016, 92, 188. [Google Scholar] [CrossRef] [PubMed]
- Heidari, M.; Wang, D.; Delekta, P.; Sun, S. Marek’s disease virus immunosuppression alters host cellular responses and immune gene expression in the skin of infected chickens. Vet. Immunol. Immunopathol. 2016, 180, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, P.; Underwood, G.; Davison, F. Differential cytokine responses following Marek’s disease virus infection of chickens differing in resistance to Marek’s disease. J. Virol. 2003, 77, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Haunshi, S.; Cheng, H.H. Differential expression of Toll-like receptor pathway genes in chicken embryo fibroblasts from chickens resistant and susceptible to Marek’s disease. Poult. Sci. 2014, 93, 550–555. [Google Scholar] [CrossRef] [PubMed]
- De Haas, E.N.; van der Eijk, J.A. Where in the serotonergic system does it go wrong? Unravelling the route by which the serotonergic system affects feather pecking in chickens. Neurosci. Biobehav. Rev. 2018, 95, 170–188. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, V.V.; Han, J.; Fasina, Y.; Harrison, S.H. Connecting gut microbiomes and short chain fatty acids with the serotonergic system and behavior in Gallus gallus and other avian species. Front. Physiol. 2022, 13, 1035538. [Google Scholar] [CrossRef]
- Krakowski, M. Violence and Serotonin: Influence of Impulse Control, Affect Regulation, and Social Functioning. J. Neuropsych. Clin. Neurosci. 2003, 15, 3. [Google Scholar] [CrossRef] [PubMed]
- Blake, P.; Grafman, J. The neurobiology of aggression. Lancet 2004, 364, 12–13. [Google Scholar] [CrossRef]
- Da Cunha-Bang, S.; Knudsen, G.M. The Modulatory Role of Serotonin on Human Impulsive Aggression. Biol. Psychiatry 2021, 90, 447–457. [Google Scholar] [CrossRef]
- Fu, Y.; Hu, J.; Erasmus, M.A.; Zhang, H.; Johnson, T.A.; Cheng, H. Cecal microbiota transplantation: Unique influence of cecal microbiota from two divergently selected inbred donor lines on cecal microbial profile, serotonergic activity, and aggressive behavior of recipient chickens. J. Anim. Sci. Biotechnol. 2023, 14, 1–16. [Google Scholar]
- Webster, A.B. Behavior of chickens. In Commercial Chicken Meat and Egg Production; Bell, D.D., Weaver, W.D., Eds.; Springer: Boston, MA, USA, 2002. [Google Scholar]
- Dennis, R.L.; Fahey, A.G.; Cheng, H.W. Alterations to embryonic serotonin change aggression and fearfulness. Aggress. Behav. 2013, 39, 91–98. [Google Scholar] [CrossRef]
- Daigle, C.L.; Rodenburg, T.B.; Bolhuis, J.E.; Swanson, J.C.; Siegford, J.M. Use of dynamic and rewarding environmental enrichment to alleviate feather pecking in non-cage laying hens. Appl. Anim. Behav. Sci. 2014, 161, 75–85. [Google Scholar] [CrossRef]
- Dennis, R.L.; Cheng, H.W. The dopaminergic system and aggression in laying hens. Poult. Sci. 2011, 90, 2440–2448. [Google Scholar] [CrossRef]
- Dennis, R.L.; Lay, D.J., Jr.; Cheng, H.W. Effects of early serotonin programming on behavioral and central monoamine concentrations in an avian model. Behav. Brain Res. 2013, 253, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Lourenço-Silva, M.I.; Ulans, A.; Campbell, A.M.; Almeida, I.C.L.; Jacobs, L. Social-pair judgment bias testing in slow-growing broiler chickens raised in low- or high-complexity environments. Sci. Rep. 2023, 13, 9393. [Google Scholar] [CrossRef] [PubMed]
- Koolhaas, J.M.; Coppens, C.M.; de Boer, S.F.; Buwalda, B.; Meerlo, P.; Timmermans, P.J. The resident-intruder paradigm: A standardized test for aggression, violence and social stress. J. Vis. Exp. 2013, 77, e4367. [Google Scholar]
- Foster, J.A.; Rinaman, L.; Cryan, J.F. Stress & the gut-brain axis: Regulation by the microbiome. Neurobiol. Stress 2017, 7, 124–136. [Google Scholar] [PubMed]
- Freimer, D.; Yang, T.T.; Ho, T.C.; Tymofiyeva, O.; Leung, C. The gut microbiota, HPA axis, and brain in adolescent-onset depression: Probiotics as a novel treatment. Brain Behav. Immun. Health 2022, 26, 100541. [Google Scholar] [CrossRef] [PubMed]
- Senchukova, M.A. Microbiota of the gastrointestinal tract: Friend or foe? World J. Gastroenterol. 2023, 29, 19–42. [Google Scholar] [CrossRef] [PubMed]
- Shakhar, K. The inclusive behavioral immune system. Front. Psychol. 2019, 10, 1004. [Google Scholar] [CrossRef]
- Parashar, A.; Udayabanu, M. Gut microbiota regulates key modulators of social behavior. Eur. Neuropsychopharmacol. 2016, 26, 78–91. [Google Scholar] [CrossRef]
- Stilling, R.M.; Bordenstein, S.R.; Dinan, T.G.; Cryan, J.F. Friends with social benefits: Host-microbe interactions as a driver of brain evolution and development? Front. Cell Infect. Microbiol. 2014, 4, 147. [Google Scholar] [CrossRef]
- Gulledge, L.; Oyebode, D.; Donaldson, J.R. The influence of the microbiome on aggressive behavior: An insight into age-related aggression. FEMS Microbiol. Lett. 2023, 370, fnac114. [Google Scholar] [CrossRef]
- Johnson, K.V. Gut microbiome composition and diversity are related to human personality traits. Hum. Microb. J. 2020, 15, 100069. [Google Scholar] [CrossRef]
- Aatsinki, A.K.; Lahti, L.; Uusitupa, H.M.; Munukka, E.; Keskitalo, A.; Nolvi, S.; O’Mahony, S.; Pietilä, S.; Elo, L.L.; Eerola, E.; et al. Gut microbiota composition is associated with temperament traits in infants. Brain Behav. Immun. 2019, 80, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Alving-Jessep, E.; Botchway, E.; Wood, A.G.; Hilton, A.C.; Blissett, J.M. The development of the gut microbiome and temperament during infancy and early childhood: A systematic review. Dev. Psychobiol. 2022, 64, e22306. [Google Scholar] [CrossRef]
- Saleena, L.A.K.; Teo, M.Y.M.; How, Y.H.; In, L.L.A.; Pui, L.P. Immunomodulatory action of Lactococcus lactis. J. Biosci. Bioeng. 2023, 135, 1–9. [Google Scholar] [CrossRef]
- Obianwuna, U.E.; Agbai Kalu, N.; Wang, J.; Zhang, H.; Qi, G.; Qiu, K.; Wu, S. Recent trends on mitigative effect of probiotics on oxidative-stress-induced gut dysfunction in broilers under necrotic enteritis challenge: A review. Antioxidants 2023, 12, 911. [Google Scholar] [CrossRef] [PubMed]
- Vaccaro, R.; Casini, A.; Severi, C.; Lamazza, A.; Pronio, A.; Palma, R. Serotonin and melatonin in human lower gastrointestinal tract. Diagnostics 2023, 13, 204. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, Z.; Wang, G.; Song, X.; Qian, Y.; Liao, Z.; Sui, L.; Ai, L.; Xia, Y. Understanding the connection between gut homeostasis and psychological stress. J. Nutr. 2023, 153, 924–939. [Google Scholar] [CrossRef] [PubMed]
- Soares, I.; Belote, B.L.; Santin, E.; Dal Pont, G.C.; Kogut, M.H. Morphological assessment and biomarkers of low-grade, chronic intestinal inflammation in production animals. Animals 2022, 12, 3036. [Google Scholar] [CrossRef]
- Riehl, L.; Furst, J.; Kress, M.; Rykalo, N. The importance of the gut microbiome and its signals for a healthy nervous system and the multifaceted mechanisms of neuropsychiatric disorders. Front. Neurosci. 2023, 17, 1302957. [Google Scholar] [CrossRef] [PubMed]
- Boehme, M.; Guzzetta, K.E.; Wasén, C.; Cox, L.M. The gut microbiota is an emerging target for improving brain health during ageing. Gut Microbiome 2023, 4, E2. [Google Scholar] [CrossRef]
- Laman, J.D. Cutting edge technologies in chronic inflammation research. Exp. Dermatol. 2022, 31, 17–21. [Google Scholar] [CrossRef]
- Wu, W.L.; Adame, M.D.; Liou, C.W.; Barlow, J.T.; Lai, T.T.; Sharon, G.; Schretter, C.E.; Needham, B.D.; Wang, M.I.; Tang, W.; et al. Microbiota regulate social behaviour via stress response neurons in the brain. Nature 2021, 595, 409–414. [Google Scholar] [CrossRef]
- Kasarello, K.; Cudnoch-Jedrzejewska, A.; Czarzasta, K. Communication of gut microbiota and brain via immune and neuroendocrine signaling. Front. Microbiol. 2023, 14, 1118529. [Google Scholar] [CrossRef]
- Bhatt, S.; Kanoujia, J.; Mohana Lakshmi, S.; Patil, C.R.; Gupta, G.; Chellappan, D.K.; Dua, K. Role of brain-gut-microbiota axis in depression: Emerging therapeutic avenues. CNS Neurol. Disord. -Drug Targets 2023, 22, 276–288. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. Regulation of the stress response by the gut microbiota: Implications for psychoneuroendocrinology. Psychoneuroendocrinology 2012, 37, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- Ilchmann-Diounou, H.; Menard, S. Psychological stress, intestinal barrier dysfunctions, and autoimmune disorders: An overview. Front. Immunol. 2020, 11, 1823. [Google Scholar] [CrossRef]
- Feng, S.; Meng, C.; Liu, Y.; Yi, Y.; Liang, A.; Zhang, Y.; Hao, Z. Bacillus licheniformis prevents and reduces anxiety-like and depression-like behaviours. Appl. Microbiol. Biotechnol. 2023, 107, 4355–4368. [Google Scholar] [CrossRef] [PubMed]
- Bloemendaal, M.; Szopinska-Tokov, J.; Belzer, C.; Boverhoff, D.; Papalini, S.; Michels, F.; van Hemert, S.; Arias Vasquez, A.; Aarts, E. Probiotics-induced changes in gut microbial composition and its effects on cognitive performance after stress: Exploratory analyses. Transl. Psychiatry 2021, 11, 300. [Google Scholar] [CrossRef] [PubMed]
- De Haas, E.N.; Newberry, R.C.; Edgar, J.; Riber, A.B.; Estevez, I.; Ferrante, V.; Hernandez, C.E.; Kjaer, J.B.; Ozkan, S.; Dimitrov, I.; et al. Prenatal and early postnatal behavioural programming in laying hens, with possible implications for the development of injurious pecking. Front. Vet. Sci. 2021, 8, 678500. [Google Scholar] [CrossRef] [PubMed]
- Kraimi, N.; Dawkins, M.; Gebhardt-Henrich, S.G.; Velge, P.; Rychlik, I.; Volf, J.; Creach, P.; Smith, A.; Colles, F.; Leterrier, C. Influence of the microbiota-gut-brain axis on behavior and welfare in farm animals: A review. Physiol. Behav. 2019, 210, 112658. [Google Scholar] [CrossRef] [PubMed]
- Davidson, G.L.; Raulo, A.; Knowles, S.C. Identifying Microbiome-Mediated Behaviour in Wild Vertebrates. Trends Ecol. Evol. 2020, 35, 972–980. [Google Scholar] [CrossRef]
- Mikami, K.; Watanabe, N.; Tochio, T.; Kimoto, K.; Akama, F.; Yamamoto, K. Impact of Gut Microbiota on Host Aggression: Potential Applications for Therapeutic Interventions Early in Development. Microorganisms 2023, 11, 1008. [Google Scholar] [CrossRef]
- Lalonde, R.; Strazielle, C. Probiotic effects on anxiety-like behavior in animal models. Rev. Neurosci. 2022, 33, 691–701. [Google Scholar] [CrossRef]
- Huang, C.; Yue, Q.; Sun, L.; Di, K.; Yang, D.; Hao, E.; Wang, D.; Chen, Y.; Shi, L.; Zhou, R.; et al. Restorative effects of Lactobacillus rhamnosus LR-32 on the gut microbiota, barrier integrity, and 5-HT metabolism in reducing feather-pecking behavior in laying hens with antibiotic-induced dysbiosis. Front. Microbiol. 2023, 14, 1173804. [Google Scholar] [CrossRef]
- Villageliu, D.N.; Lyte, M. Microbial endocrinology: Why the intersection of microbiology and neurobiology matters to poultry health. Poult. Sci. 2017, 96, 2501–2508. [Google Scholar] [CrossRef]
- Sun, W.; Zhang, L. Antioxidant indexes and immune function of the intestinal flora of compound microecological preparations. Oxidative Med. Cell. Longev. 2022, 2022, 5498514. [Google Scholar] [CrossRef]
- Birkl, P.; Bharwani, A.; Kjaer, J.B.; Kunze, W.; McBride, P.; Forsythe, P.; Harlander-Matauschek, A. Differences in cecal microbiome of selected high and low feather-pecking laying hens. Poult. Sci. 2018, 97, 3009–3014. [Google Scholar] [CrossRef]
- Campbell, C.; Kandalgaonkar, M.R.; Golonka, R.M.; Yeoh, B.S.; Vijay-Kumar, M.; Saha, P. Crosstalk between gut microbiota and host immunity: Impact on inflammation and immunotherapy. Biomedicines 2023, 11, 294. [Google Scholar] [CrossRef] [PubMed]
- Jach, M.E.; Serefko, A.; Szopa, A.; Sajnaga, E.; Golczyk, H.; Santos, L.S.; Borowicz-Reutt, K.; Sieniawska, E. The role of probiotics and their metabolites in the treatment of depression. Molecules 2023, 28, 3213. [Google Scholar] [CrossRef] [PubMed]
- Mazziotta, C.; Tognon, M.; Martini, F.; Torreggiani, E.; Rotondo, J.C. Probiotics mechanism of action on immune cells and beneficial effects on human health. Cells 2023, 12, 184. [Google Scholar] [CrossRef] [PubMed]
- Ribaldone, D.G.; Pellicano, R.; Fagoonee, S.; Actis, G.C. Modulation of the gut microbiota: Opportunities and regulatory aspects. Minerva Gastroenterol. 2023, 69, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Liu, F.; Mao, Y.; Zhang, Y.; Zeng, H.; Ren, S.; Guo, L.; Chen, Z.; Hrabchenko, N.; et al. Mechanisms and applications of probiotics in prevention and treatment of swine diseases. Porcine Health Manag. 2023, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, Z.; Tang, P.; Wu, Y.; Zhang, A.; Li, D.; Wang, C.Z.; Wan, J.Y.; Yao, H.; Yuan, C.S. Probiotics fortify intestinal barrier function: A systematic review and meta-analysis of randomized trials. Front. Immunol. 2023, 14, 1143548. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.M.; Kassam, Z.; Bercik, P. The adoptive transfer of behavioral phenotype via the intestinal microbiota: Experimental evidence and clinical implications. Curr. Opin. Microbiol. 2013, 16, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Ait-Belgnaoui, A.; Colom, A.; Braniste, V.; Ramalho, L.; Marrot, A.; Cartier, C.; Houdeau, E.; Theodorou, V.; Tompkins, T. Probiotic gut effect prevents the chronic psychological stress-induced brain activity abnormality in mice. Neurogastroenterol. Motil. 2014, 26, 510–520. [Google Scholar] [CrossRef]
- Parois, S.; Calandreau, L.; Kraimi, N.; Gabriel, I.; Leterrier, C. The influence of a probiotic supplementation on memory in quail suggests a role of gut microbiota on cognitive abilities in birds. Behav. Brain Res. 2017, 331, 47–53. [Google Scholar] [CrossRef]
- Naglaa, M. Do probiotics affect the behavior of turkey poults? J. Vet. Med. Anim. Health 2013, 5, 144–148. [Google Scholar]
- Mindus, C.; van Staaveren, N.; Bharwani, A.; Fuchs, D.; Gostner, J.M.; Kjaer, J.B.; Kunze, W.; Mian, M.F.; Shoveller, A.K.; Forsythe, P.; et al. Ingestion of Lactobacillus rhamnosus modulates chronic stress-induced feather pecking in chickens. Sci. Rep. 2021, 11, 17119. [Google Scholar] [CrossRef] [PubMed]
- Mindus, C.; van Staaveren, N.; Fuchs, D.; Gostner, J.M.; Kjaer, J.B.; Kunze, W.; Mian, M.F.; Shoveller, A.K.; Forsythe, P.; Harlander-Matauschek, A. Regulatory T cell modulation by lactobacillus rhamnosus improves feather damage in chickens. Front. Vet. Sci. 2022, 9, 855261. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Hu, J.Y.; Cheng, H.W. The Impact of Probiotic Bacillus subtilis on Injurious Behavior in Laying Hens. Animals 2022, 12, 870. [Google Scholar] [CrossRef] [PubMed]
- Allegretti, J.R.; Kelly, C.R.; Grinspan, A.; Mullish, B.H.; Hurtado, J.; Carrellas, M.; Marcus, J.; Marchesi, J.R.; McDonald, J.A.; Gerardin, Y.; et al. Inflammatory Bowel Disease Outcomes Following Fecal Microbiota Transplantation for Recurrent C. difficile Infection. Inflamm. Bowel Dis. 2021, 27, 1371–1378. [Google Scholar] [CrossRef] [PubMed]
- Tariq, R.; Syed, T.; Yadav, D.; Prokop, L.J.; Singh, S.; Loftus, E.V., Jr.; Pardi, D.S.; Khanna, S. Outcomes of fecal microbiota transplantation for C. difficile infection in inflammatory bowel disease: A systematic review and meta-analysis. J. Clin. Gastroenterol. 2023, 57, 285–293. [Google Scholar] [CrossRef]
- Anand, N.; Gorantla, V.R.; Chidambaram, S.B. The Role of Gut Dysbiosis in the Pathophysiology of Neuropsychiatric Disorders. Cells 2022, 12, 54. [Google Scholar] [CrossRef]
- Hamamah, S.; Gheorghita, R.; Lobiuc, A.; Sirbu, I.O.; Covasa, M. Fecal microbiota transplantation in non-communicable diseases: Recent advances and protocols. Front. Med. 2022, 9, 1060581. [Google Scholar] [CrossRef]
- Qi, R.; Zhang, Z.; Wang, J.; Qiu, X.; Wang, Q.; Yang, F.; Huang, J.; Liu, Z. Introduction of colonic and fecal microbiota from an adult pig differently affects the growth, gut health, intestinal microbiota and blood metabolome of newborn piglets. Front. Microbiol. 2021, 12, 623673. [Google Scholar] [CrossRef] [PubMed]
- Shehata, E.; Parker, A.; Suzuki, T.; Swann, J.R.; Suez, J.; Kroon, P.A.; Day-Walsh, P. Microbiomes in physiology: Insights into 21st-century global medical challenges. Exp. Physiol. 2022, 107, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Medel-Matus, J.S.; Shin, D.; Dorfman, E.; Sankar, R.; Mazarati, A. Facilitation of kindling epileptogenesis by chronic stress may be mediated by intestinal microbiome. Epilepsia Open 2018, 3, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Pittman, Q.J. Stress co-opts the gut to affect epileptogenesis. Commentary on “Facilitation of kindling epileptogenesis by chronic stress may be mediated by intestinal microbiome”. Epilepsia Open 2019, 4, 230–231. [Google Scholar] [CrossRef]
- Kragsnaes, M.S.; Kjeldsen, J.; Horn, H.C.; Munk, H.L.; Pedersen, J.K.; Just, S.A.; Ahlquist, P.; Pedersen, F.M.; de Wit, M.; Möller, S.; et al. Safety and efficacy of faecal microbiota transplantation for active peripheral psoriatic arthritis: An exploratory randomised placebo-controlled trial. Ann. Rheum. Dis. 2021, 80, 1158–1167. [Google Scholar] [CrossRef]
- Ting, N.L.; Lau, H.C.; Yu, J. Cancer pharmacomicrobiomics: Targeting microbiota to optimise cancer therapy outcomes. Gut 2022, 71, 1412–1425. [Google Scholar] [CrossRef]
- Drugs.com. Rebyota. 2022. Available online: https://www.drugs.com/rebyota.html (accessed on 30 November 2023).
- Pauwels, J.; Taminiau, B.; Janssens, G.P.; De Beenhouwer, M.; Delhalle, L.; Daube, G.; Coopman, F. Cecal drop reflects the chickens’ cecal microbiome, fecal drop does not. J. Microbiol. Methods 2015, 117, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Panasevich, M.R.; Wankhade, U.D.; Chintapalli, S.V.; Shankar, K.; Rector, R.S. Cecal versus fecal microbiota in Ossabaw swine and implications for obesity. Physiol. Genom. 2018, 50, 355–368. [Google Scholar] [CrossRef]
- Kozik, A.J.; Nakatsu, C.H.; Chun, H.; Jones-Hall, Y.L. Comparison of the fecal, cecal, and mucus microbiome in male and female mice after TNBS-induced colitis. PLoS ONE 2019, 14, e0225079. [Google Scholar] [CrossRef]
- Løtvedt, P.; Fallahshahroudi, A.; Bektic, L.; Altimiras, J.; Jensen, P. Chicken domestication changes expression of stress-related genes in brain, pituitary and adrenals. Neurobiol. Stress 2017, 7, 113–121. [Google Scholar] [CrossRef]
- Williams, C.L.; Garcia-Reyero, N.; Martyniuk, C.J.; Tubbs, C.W.; Bisesi, J.H., Jr. Regulation of endocrine systems by the microbiome: Perspectives from comparative animal models. Gen. Comp. Endocrinol. 2020, 292, 113437. [Google Scholar] [CrossRef]
- Kalia, V.C.; Shim, W.Y.; Patel, S.K.S.; Gong, C.; Lee, J.K. Recent developments in antimicrobial growth promoters in chicken health: Opportunities and challenges. Sci. Total Environ. 2022, 834, 155300. [Google Scholar] [CrossRef]
- Lynch, C.M.K.; Cowan, C.S.M.; Bastiaanssen, T.F.S.; Moloney, G.M.; Theune, N.; van de Wouw, M.; Florensa Zanuy, E.; Ventura-Silva, A.P.; Codagnone, M.G.; Villalobos-Manríquez, F.; et al. Critical windows of early-life microbiota disruption on behaviour, neuroimmune function, and neurodevelopment. Brain Behav. Immun. 2023, 108, 309–327. [Google Scholar] [CrossRef]
- Goldstein, D.L. Absorption by the cecum of wild birds: Is there interspecific variation. J. Exp. Zool. 1989, 252 (Suppl. S3), 103–110. [Google Scholar] [CrossRef]
- Clench, M.H. The avian cecum: Update and motility review. J. Exp. Zool. 1999, 83, 441–447. [Google Scholar] [CrossRef]
- Svihus, B. Function of the digestive system. J. Appl. Poul. Res. 2014, 23, 306–314. [Google Scholar] [CrossRef]
- Svihus, B.; Choct, M.; Classen, H. Function and nutritional roles of the avian caeca: A review. Worlds Poult. Sci. J. 2013, 69, 249–264. [Google Scholar] [CrossRef]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Rychlik, I. Composition and function of chicken gut microbiota. Animals 2020, 10, 103. [Google Scholar] [CrossRef]
- Plata, G.; Baxter, N.T.; Susanti, D.; Volland-Munson, A.; Gangaiah, D.; Nagireddy, A.; Mane, S.P.; Balakuntla, J.; Hawkins, T.B.; Kumar Mahajan, A. Growth promotion and antibiotic induced metabolic shifts in the chicken gut microbiome. Commun. Biol. 2022, 5, 293. [Google Scholar] [CrossRef]
- Such, N.; Farkas, V.; Csitári, G.; Pál, L.; Márton, A.; Menyhárt, L.; Dublecz, K. Relative effects of dietary administration of a competitive exclusion culture and a synbiotic product, age and sampling site on intestinal microbiota maturation in broiler chickens. Vet. Sci. 2021, 8, 187. [Google Scholar] [CrossRef]
- Xiao, S.S.; Mi, J.D.; Mei, L.; Liang, J.; Feng, K.X.; Wu, Y.B.; Liao, X.D.; Wang, Y. Microbial diversity and community variation in the intestines of layer chickens. Animals 2021, 11, 840. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Jiang, Y.; Tang, J.; Chang, G.; Zhao, W.; Hou, S.; Chen, G. Comparison of cecal microbiota and performance indices between lean-type and fatty-type pekin ducks. Front. Microbiol. 2022, 13, 820569. [Google Scholar] [CrossRef] [PubMed]
- Konopka, A. What is microbial community ecology? ISME J. 2009, 3, 1223–1230. [Google Scholar] [CrossRef]
- Shah, T.; Baloch, Z.; Shah, Z.; Cui, X.; Xia, X. The intestinal microbiota: Impacts of antibiotics therapy, colonization resistance, and diseases. Int. J. Mol. Sci. 2021, 22, 6597. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Hu, Y.; Wu, S.; Shi, S. Comparative metagenomic analysis of chicken gut microbial community, function, and resistome to evaluate noninvasive and cecal sampling resources. Animals 2021, 11, 1718. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Du, P.; Huang, S.; He, D.; Chen, J.; Wen, X.; Yang, J.; Xian, S.; Cheng, Z. Comparison of the caecal microbial community structure and physiological indicators of healthy and infection Eimeria tenella chickens during peak of oocyst shedding. Avian Pathol. 2023, 52, 51–61. [Google Scholar] [CrossRef]
- Ye, J.; Yang, H.; Hu, W.; Tang, K.; Liu, A.; Bi, S. Changed cecal microbiota involved in growth depression of broiler chickens induced by immune stress. Poult. Sci. 2023, 102, 102598. [Google Scholar] [CrossRef]
- Yin, Z.; Ji, S.; Yang, J.; Guo, W.; Li, Y.; Ren, Z.; Yang, X. Cecal Microbial Succession and Its Apparent Association with Nutrient Metabolism in Broiler Chickens. mSphere 2023, 8, e0061422. [Google Scholar] [CrossRef]
- Franco, L.; Boulianne, M.; Parent, E.; Barjesteh, N.; Costa, M.C. Colonization of the gastrointestinal tract of chicks with different bacterial microbiota profiles. Animals 2023, 13, 2633. [Google Scholar] [CrossRef]
- Gong, Y.; Yang, H.; Wang, X.; Xia, W.; Lv, W.; Xiao, Y.; Zou, X. Early intervention with cecal fermentation broth regulates the colonization and development of gut microbiota in broiler chickens. Front. Microbiol. 2019, 10, 1422. [Google Scholar] [CrossRef]
- Volf, J.; Polansky, O.; Varmuzova, K.; Gerzova, L.; Sekelova, Z.; Faldynova, M.; Babak, V.; Medvecky, M.; Smith, A.L.; Kaspers, B.; et al. Transient and prolonged response of chicken cecum mucosa to colonization with different gut microbiota. PLoS ONE 2016, 11, e0163932. [Google Scholar] [CrossRef]
- van Hierden, Y.M.; de Boer, S.F.; Koolhaas, J.M.; Korte, S.M. The control of feather pecking by serotonin. Behav. Neurosci. 2004, 118, 575–583. [Google Scholar] [CrossRef]
- Falker-Gieske, C.; Bennewitz, J.; Tetens, J. The light response in chickens divergently selected for feather pecking behavior reveals mechanistic insights towards psychiatric disorders. Mol. Biol. Rep. 2022, 49, 1649–1654. [Google Scholar] [CrossRef] [PubMed]
- Pasquaretta, C.; Gómez-Moracho, T.; Heeb, P.; Lihoreau, M. Exploring interactions between the gut microbiota and social behavior through nutrition. Genes 2018, 9, 534. [Google Scholar] [CrossRef] [PubMed]
- Maraci, Ö.; Antonatou-Papaioannou, A.; Jünemann, S.; Engel, K.; Castillo-Gutiérrez, O.; Busche, T.; Kalinowski, J.; Caspers, B.A. Timing matters: Age-dependent impacts of the social environment and host selection on the avian gut microbiota. Microbiome 2022, 10, 202. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, G.A.; Richardson, E.; Clark, J.; Keshri, J.; Drechsler, Y.; Berrang, M.E.; Meinersmann, R.J.; Cox, N.A.; Oakley, B.B. Broiler chickens and early life programming: Microbiome transplant-induced cecal community dynamics and phenotypic effects. PLoS ONE 2020, 15, e0242108. [Google Scholar] [CrossRef]
- Glendinning, L.; Chintoan-Uta, C.; Stevens, M.P.; Watson, M. Effect of cecal microbiota transplantation between different broiler breeds on the chick flora in the first week of life. Poult. Sci. 2022, 101, 101624. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, X.; Wu, Y.; Peng, J.; Wei, H. Effect of the microbiome on intestinal innate immune development in early life and the potential strategy of early intervention. Front. Immunol. 2022, 13, 936300. [Google Scholar] [CrossRef]
- Brett, B.E.; de Weerth, C. The microbiota-gut-brain axis: A promising avenue to foster healthy developmental outcomes. Dev. Psychobiol. 2019, 61, 772–782. [Google Scholar] [CrossRef]
- Forssberg, H. Microbiome programming of brain development: Implications for neurodevelopmental disorders. Dev. Med. Child. Neurol. 2019, 61, 744–749. [Google Scholar] [CrossRef] [PubMed]
- AABP (American Association of Bovine Practitioners). Dehorning Guidelines. 2019. Available online: https://www.aabp.org/resources/aabp_guidelines/dehorning-2019.pdf (accessed on 30 November 2023).
- AVMA (The American Veterinary Medical Association). Supersede Policy on Castration and Dehorning of Cattle. Resolution #4—2023. 2023. Available online: https://www.avma.org/sites/default/files/2023-03/2023W_Resolution4F.pdf (accessed on 30 November 2023).
- AVMA (the American Veterinary Medical Association). Teeth Clipping, Tail Docking and Permanent Identification of Piglet. 2014. Available online: https://www.avma.org/sites/default/files/resources/practices_piglets_bgnd.pdf (accessed on 30 November 2023).
- D’Eath, R.B.; O’Driscoll, K.; Fàbrega, E. Editorial: Holistic prevention strategies for tail biting in pigs; from farm to slaughterhouse. Front. Vet. Sci. 2023, 10, 1296461. [Google Scholar] [CrossRef] [PubMed]
- Bleibel, L.; Dziomba, S.; Waleron, K.F.; Kowalczyk, E.; Karbownik, M.S. Deciphering psychobiotics’ mechanism of action: Bacterial extracellular vesicles in the spotlight. Front. Microbiol. 2023, 14, 1211447. [Google Scholar] [CrossRef]
- Ross, K. Psychobiotics: Are they the future intervention for managing depression and anxiety? A literature review. Explore 2023, 9, 669–680. [Google Scholar] [CrossRef] [PubMed]
- NAMI (National Alliance on Mental Illness). Mental Health by the Numbers. 2023. Available online: https://www.nami.org/mhstats (accessed on 30 November 2023).
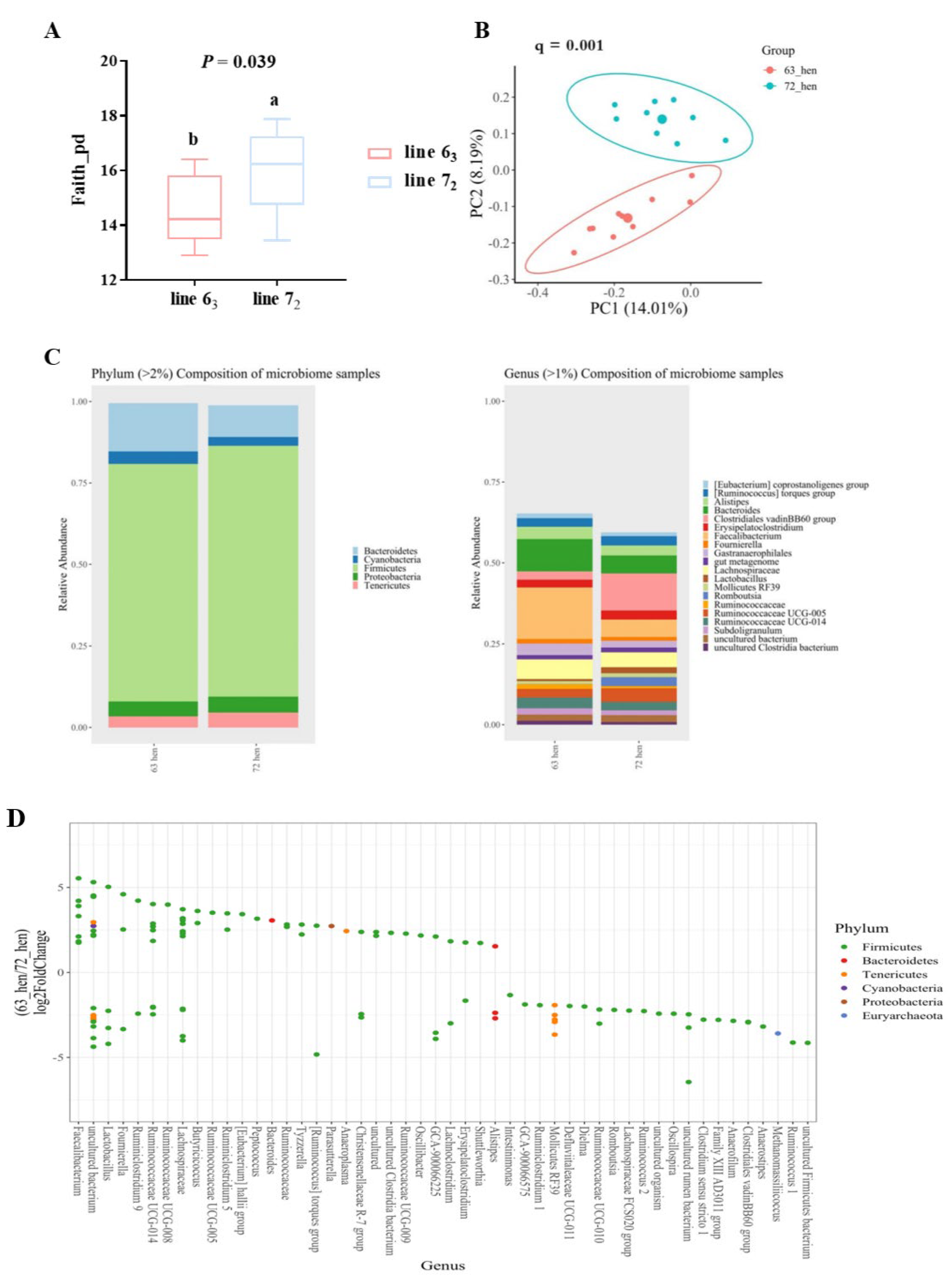
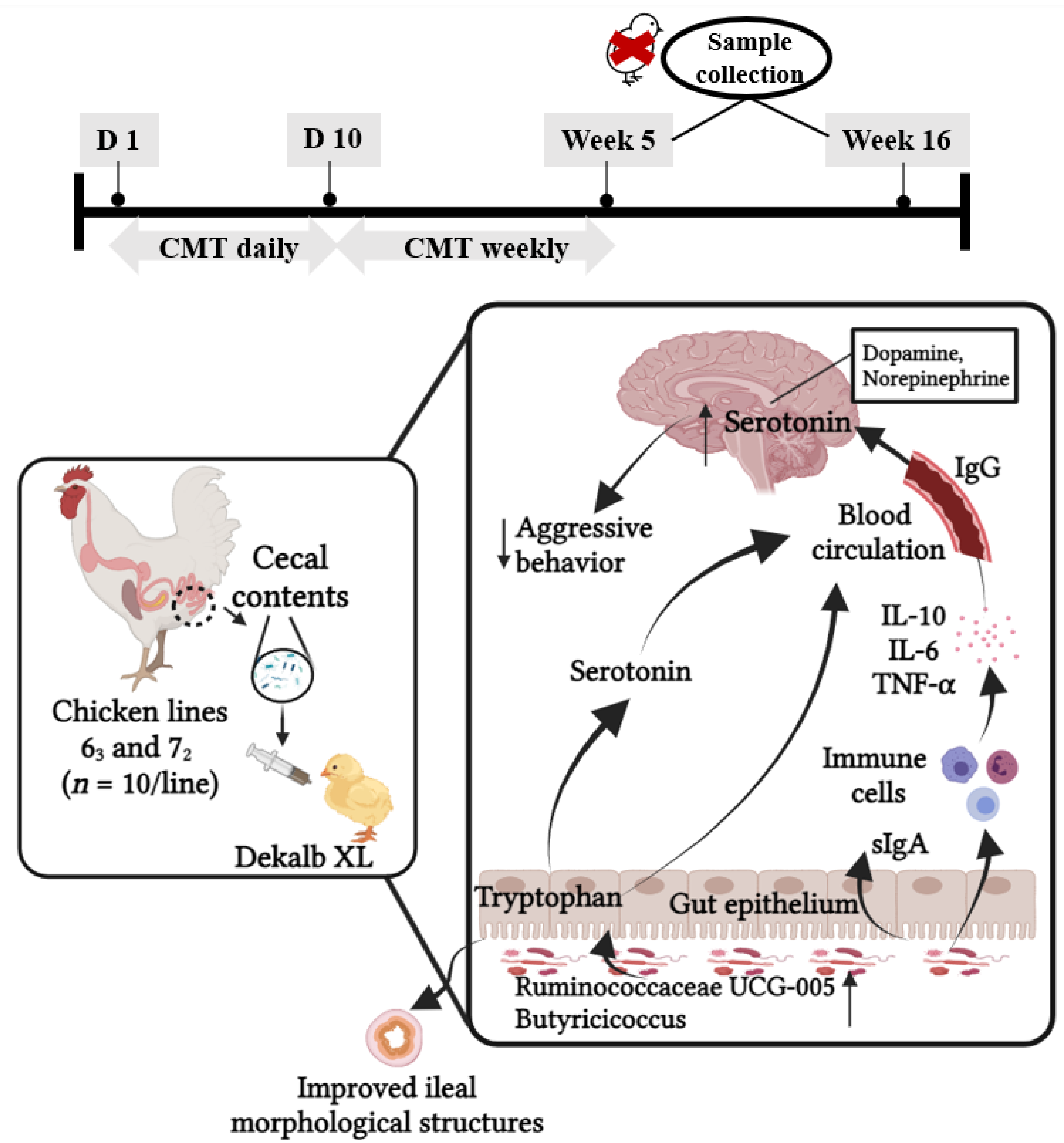
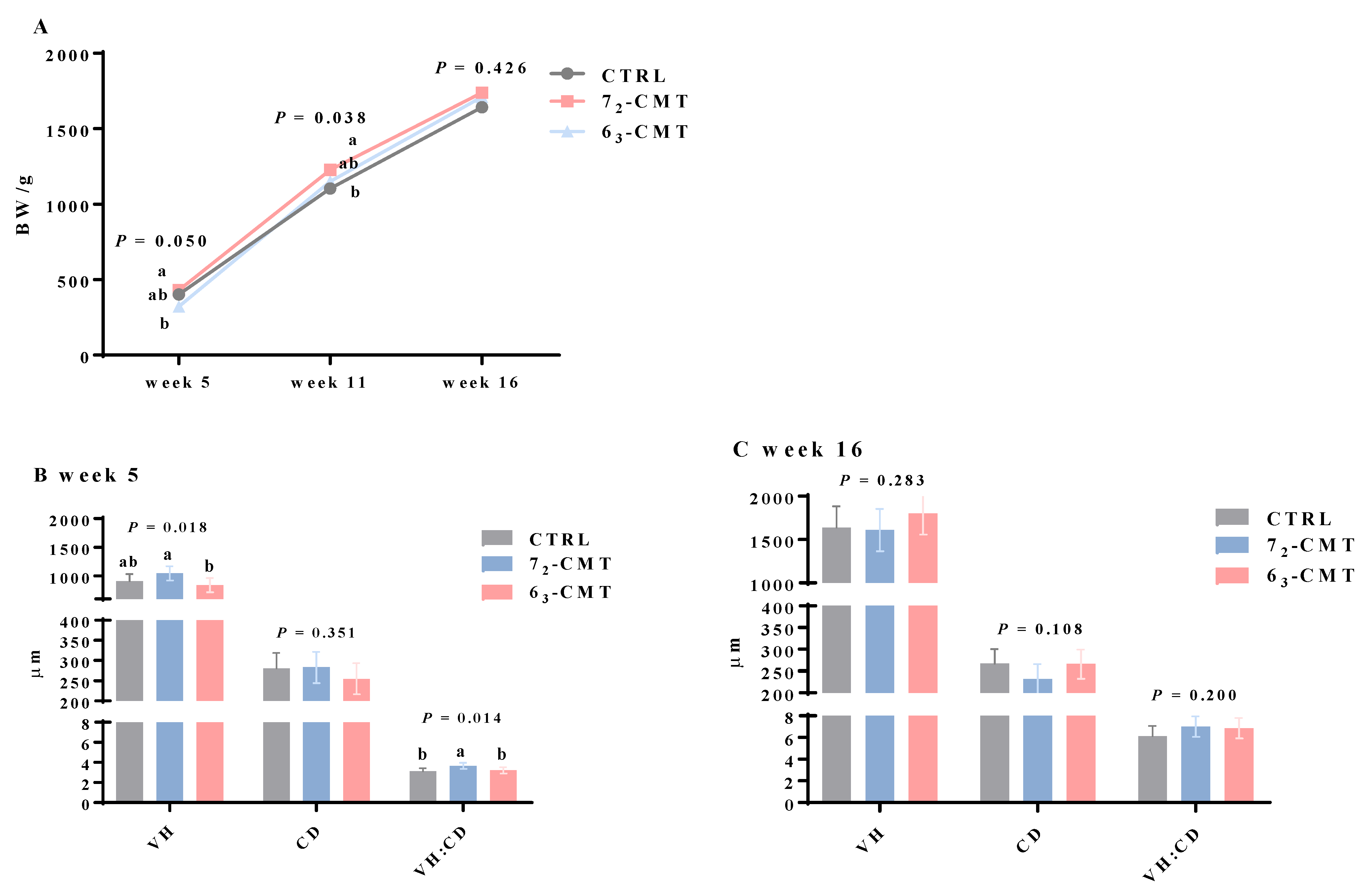
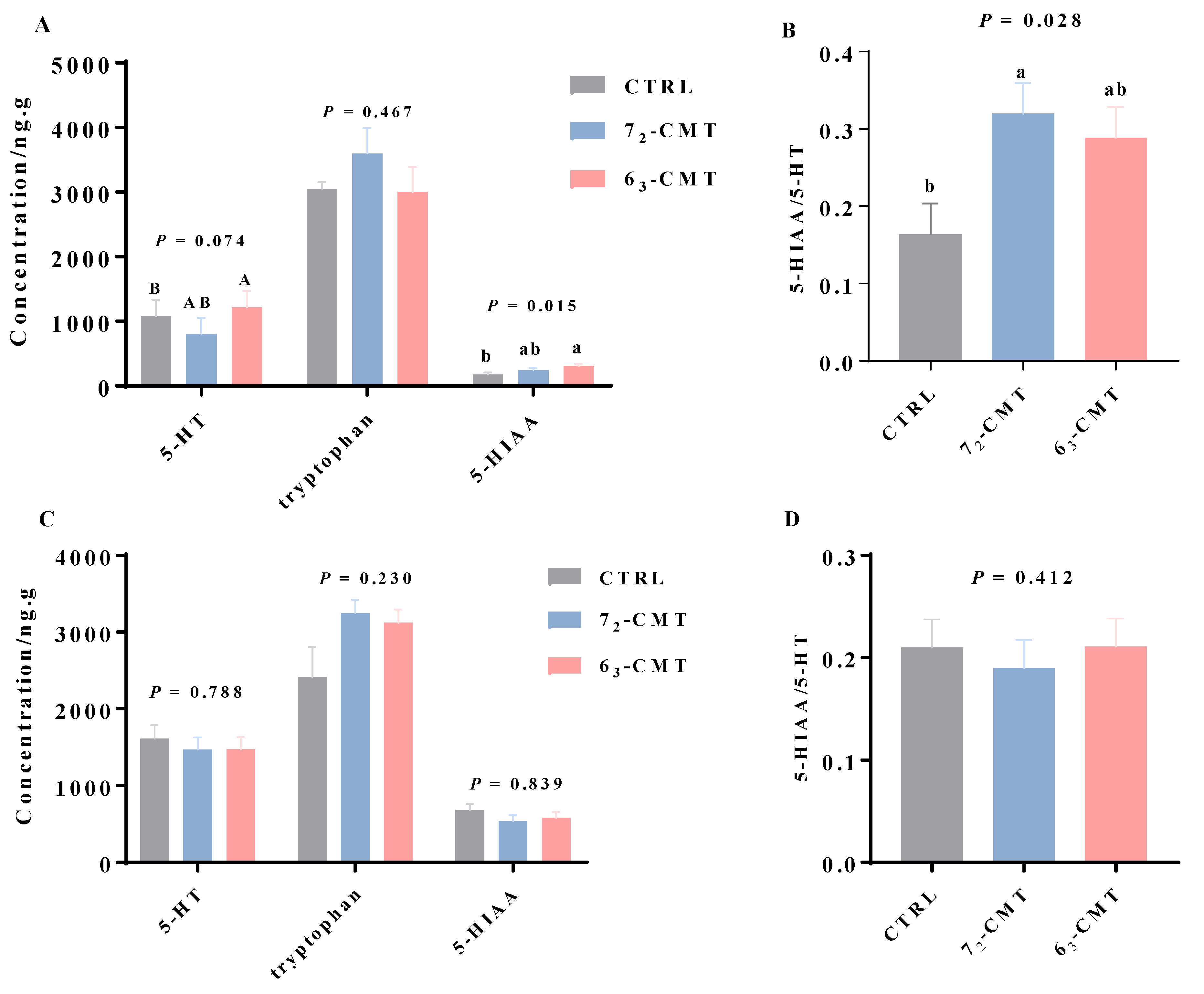
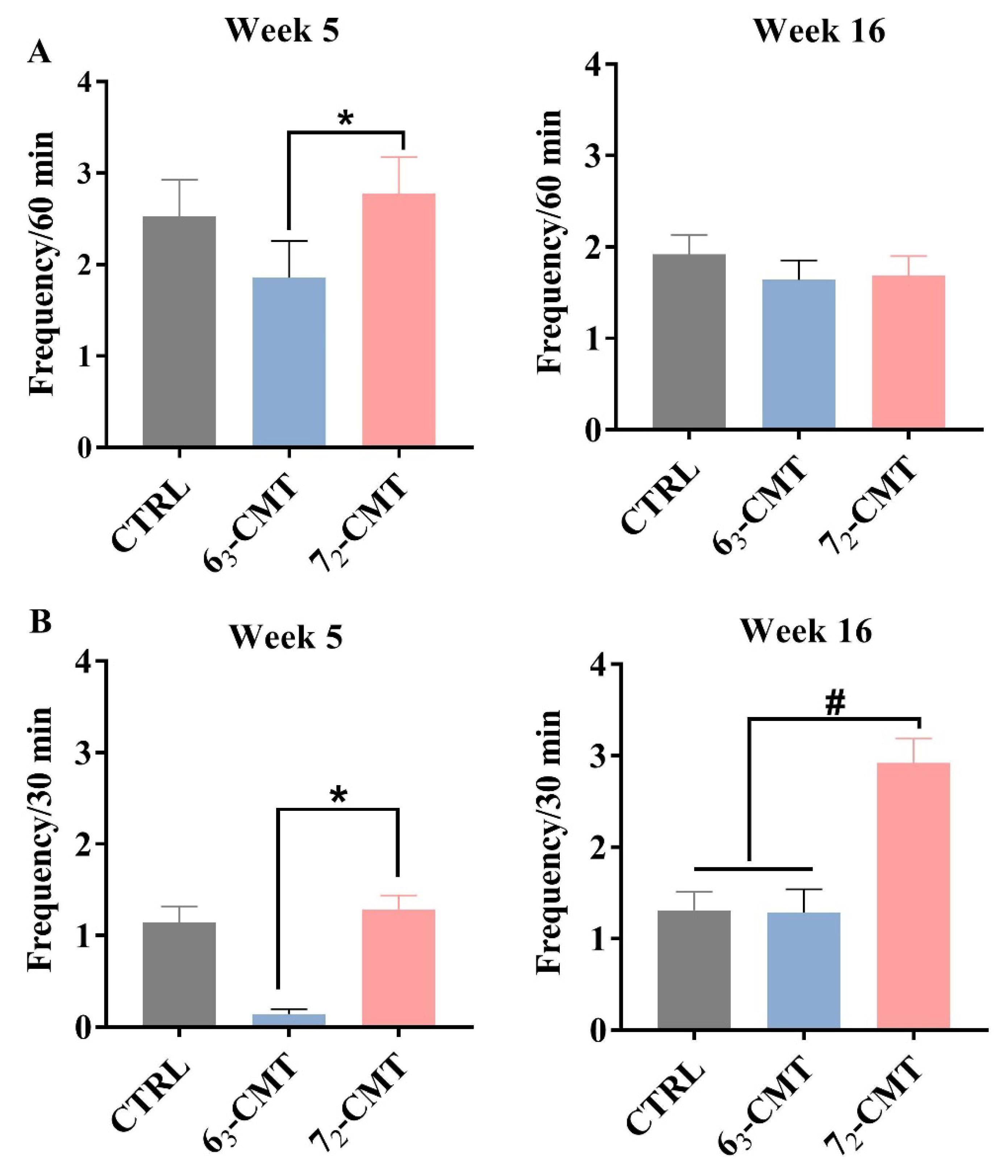
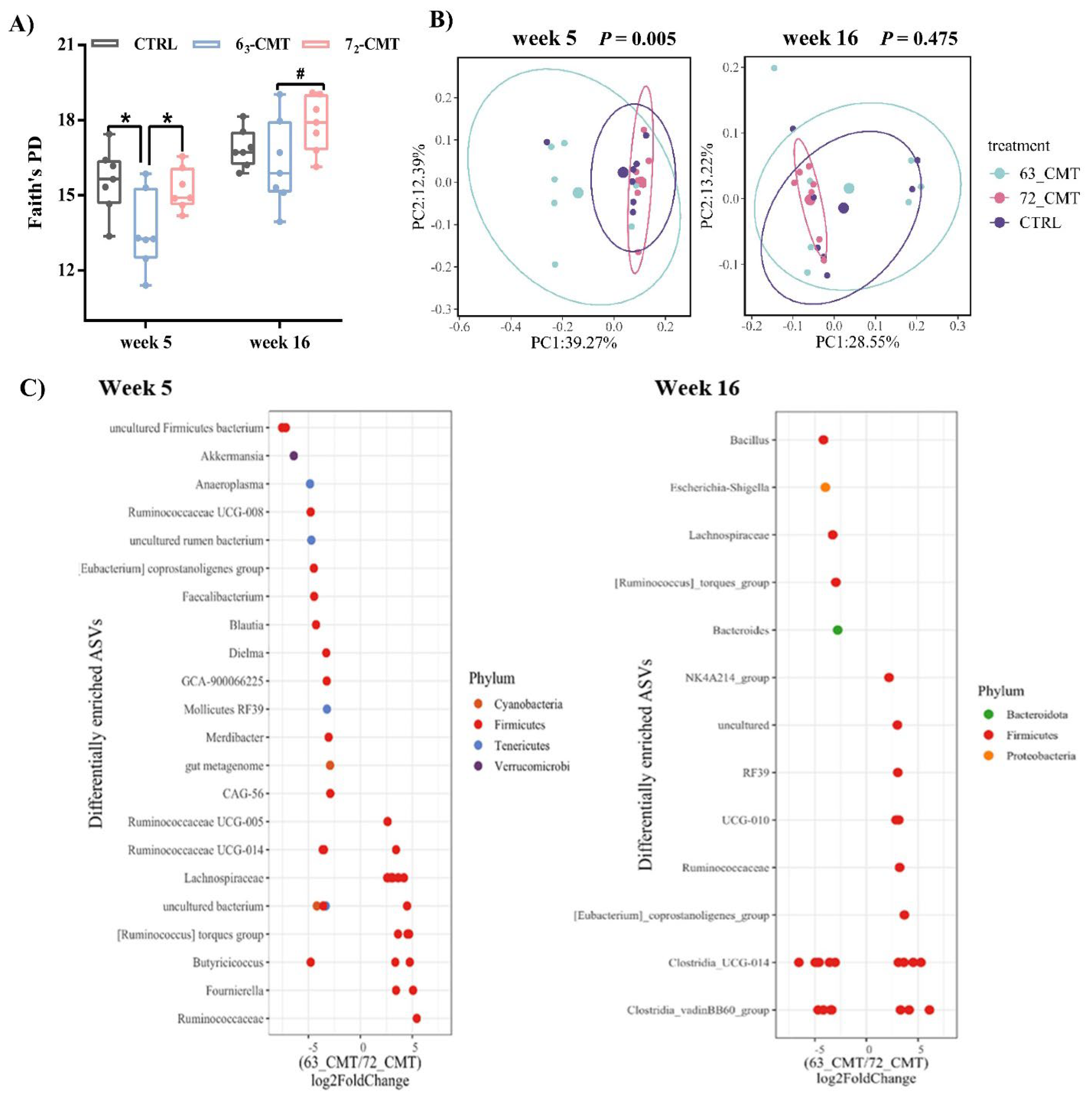

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) | |||||
|---|---|---|---|---|---|
| Treatment | 5-HT (ng/g) | 5-HIAA (ng/g) | 5-HT/5-HIAA | TRP | |
| Line 63 | 512.6 a | 151.8 | 3.2 b | 1183.8 a | |
| Line 72 | 352.7 b | 168.9 | 4.9 a | 963.2 b | |
| SEM | 8.2 | 12.9 | 0.2 | 22.4 | |
| p-value | 0.01 | 0.62 | 0.04 | 0.08 | |
| (B) | |||||
| Treatment | 5-HT (ng/g) | TRP (ng/g) | CORT (ng/mL) | H/L ratio | |
| Line 63 | 61.38 | 171.52 a | 8.44 b | 0.16 b | |
| Line 72 | 59.46 | 121.42 b | 9.75 a | 0.50 a | |
| SEM | 3.79 | 15.37 | 1.51 | 0.04 | |
| p-value | 0.73 | 0.03 | 0.05 | <0.0001 | |
| (C) | |||||
| Treatment | IgG (mg/mL) | IL-6 (pg/mL) | IL-2 (pg/mL) | IL-10 (pg/mL) | TNF-α (ng/mL) |
| Line 63 | 12.0 | 28.14 | 60.09 | 9.37 | 36.65 A |
| Line 72 | 12.9 | 27.56 | 71.65 | 13.13 | 30.73 B |
| SEM | 0.73 | 1.63 | 12.8 | 1.64 | 2.37 |
| p-value | 0.54 | 0.81 | 0.54 | 0.12 | 0.09 |
| Measures | Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| CTRL | 72-CMT | 63-CMT | |||
| Body weight | 1642.5 | 1738.6 | 1711.3 | 34.3 | 0.426 |
| Adrenal gland 1 | 4.181 AB | 4.762 A | 3.306 B | 0.420 | 0.090 |
| H/L ratio | 0.327 ab | 0.367 a | 0.243 b | 0.029 | 0.024 |
| Corticosterone (ng/mL) | 4.235 | 4.678 | 3.697 | 0.900 | 0.789 |
| Testosterone (ng/mL) | 1.423 | 1.132 | 1.744 | 0.277 | 0.345 |
| Treatment | IgG (mg/mL) | IL-6 (pg/mL) | TNF-α (pg/mL) | IL-10 (pg/mL) |
|---|---|---|---|---|
| Week 5 | ||||
| CTRL | 5.197 | 38.532 | 22.846 | 42.569 |
| 72-CMT | 5.412 | 37.109 | 26.495 | 33.259 |
| 63-CMT | 5.245 | 32.903 | 26.211 | 37.503 |
| SEM | 0.624 | 2.014 | 2.597 | 5.254 |
| p-value | 0.565 | 0.118 | 0.293 | 0.499 |
| Week 16 | ||||
| CTRL | 15.032 ab | 43.128 AB | 16.660 | 27.467 ab |
| 72-CMT | 17.993 a | 47.523 A | 21.706 | 26.928 b |
| 63-CMT | 13.716 b | 38.597 B | 16.161 | 33.835 a |
| SEM | 1.176 | 3.294 | 1.896 | 1.997 |
| p-value | 0.046 | 0.070 | 0.107 | 0.045 |
| Treatment | sIgA (mg/g) | Relative mRNA Abundance | ||
|---|---|---|---|---|
| IL-6 | TNF-α | IL-10 | ||
| Week 5 | ||||
| CTRL | 2.167 ab | 0.806 | 0.905 | 0.396 |
| 72-CMT | 1.757 b | 0.763 | 1.378 | 0.461 |
| 63-CMT | 3.473 a | 0.673 | 1.280 | 0.258 |
| SEM | 0.440 | 0.141 | 0.175 | 0.153 |
| p-value | 0.045 | 0.796 | 0.296 | 0.456 |
| Week 16 | ||||
| CTRL | 6.433 | 1.133 AB | 2.390 AB | 0.879 |
| 72-CMT | 7.989 | 1.694 A | 2.741 A | 0.739 |
| 63-CMT | 9.914 | 0.832 B | 2.217 B | 0.816 |
| SEM | 1.369 | 0.263 | 0.149 | 0.266 |
| p-value | 0.249 | 0.080 | 0.065 | 0.722 |
| Treatment | MAOA | 5-HT (ng/g) | 5-HIAA (ng/g) | 5-HIAA/5-HT | Tryptophan (ng/g) | Dopamine (ng/g) | Norepinephrine (ng/g) |
|---|---|---|---|---|---|---|---|
| Week 5 | |||||||
| 63-CMT | 1.51 | 496 a | 122.3 a | 0.225 | 1784 A | 55.7 ab | 394 AB |
| 72-CMT | 1.56 | 388 b | 86.7 b | 0.231 | 1532 AB | 44.5 b | 324 B |
| CTRL | 1.45 | 482 ab | 108.3 ab | 0.225 | 1454 B | 70.6 a | 494 A |
| SEM | 0.11 | 29.4 | 6.9 | 0.014 | 103.6 | 6.6 | 28.1 |
| p-value | 0.80 | 0.04 | 0.007 | 0.500 | 0.09 | 0.03 | 0.07 |
| Week 16 | |||||||
| 63-CMT | 2.52 a | 397 | 59.3 | 0.164 a | 2760 | 121 | 540 |
| 72-CMT | 1.80 b | 368 | 50.9 | 0.131 ab | 2140 | 121 | 517 |
| CTRL | 1.90 ab | 384 | 42 | 0.110 b | 2480 | 117 | 506 |
| SEM | 0.05 | 29.5 | 5.2 | 0.012 | 176 | 9.6 | 30.7 |
| p-value | 0.02 | 0.80 | 0.23 | 0.011 | 0.06 | 0.94 | 0.71 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Hu, J.; Zhang, H.; Erasmus, M.A.; Johnson, T.A.; Cheng, H.-W. The Impact of Early-Life Cecal Microbiota Transplantation on Social Stress and Injurious Behaviors in Egg-Laying Chickens. Microorganisms 2024, 12, 471. https://doi.org/10.3390/microorganisms12030471
Fu Y, Hu J, Zhang H, Erasmus MA, Johnson TA, Cheng H-W. The Impact of Early-Life Cecal Microbiota Transplantation on Social Stress and Injurious Behaviors in Egg-Laying Chickens. Microorganisms. 2024; 12(3):471. https://doi.org/10.3390/microorganisms12030471
Chicago/Turabian StyleFu, Yuechi, Jiaying Hu, Huanmin Zhang, Marisa A. Erasmus, Timothy A. Johnson, and Heng-Wei Cheng. 2024. "The Impact of Early-Life Cecal Microbiota Transplantation on Social Stress and Injurious Behaviors in Egg-Laying Chickens" Microorganisms 12, no. 3: 471. https://doi.org/10.3390/microorganisms12030471
APA StyleFu, Y., Hu, J., Zhang, H., Erasmus, M. A., Johnson, T. A., & Cheng, H.-W. (2024). The Impact of Early-Life Cecal Microbiota Transplantation on Social Stress and Injurious Behaviors in Egg-Laying Chickens. Microorganisms, 12(3), 471. https://doi.org/10.3390/microorganisms12030471