
Altered Plasma microRNA Signature in Hospitalized COVID-19 Patients Requiring Oxygen Support
 , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Cohort
2.2. RNA Extraction
2.3. Small RNA Library Generation and Sequencing
2.4. miRNA Sequence Analysis
2.5. Statistical Analysis
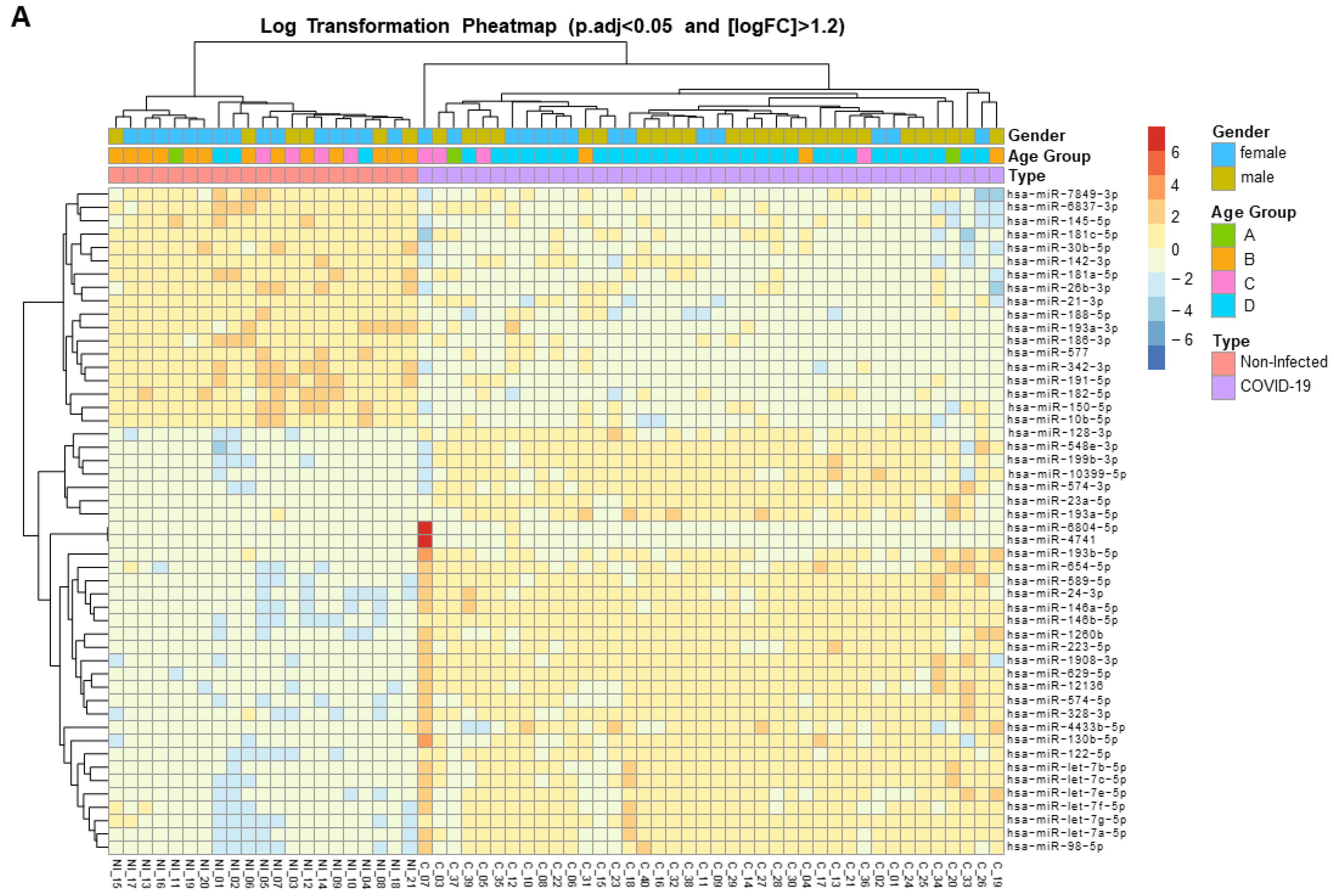
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Price-Haywood, E.G.; Burton, J.; Fort, D.; Seoane, L. Hospitalization and Mortality among Black Patients and White Patients with COVID-19. N. Engl. J. Med. 2020, 382, 2534–2543. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: Major findings, mechanisms and recommendations. Nat. Rev. Microbiol. 2023, 21, 133–146. [Google Scholar] [CrossRef]
- Nyberg, T.; Ferguson, N.M.; Nash, S.G.; Webster, H.H.; Flaxman, S.; Andrews, N.; Hinsley, W.; Bernal, J.L.; Kall, M.; Bhatt, S.; et al. Comparative analysis of the risks of hospitalisation and death associated with SARS-CoV-2 omicron (B.1.1.529) and delta (B.1.617.2) variants in England: A cohort study. Lancet 2022, 399, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Ameres, S.L.; Zamore, P.D. Diversifying microRNA sequence and function. Nat. Rev. Mol. Cell Biol. 2013, 14, 475–488. [Google Scholar] [CrossRef]
- Diener, C.; Keller, A.; Meese, E. Emerging concepts of miRNA therapeutics: From cells to clinic. Trends Genet. 2022, 38, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, X.; Wu, Z.; Huang, K.; Sun, X.; Chen, H.; Jin, M. The Downregulation of MicroRNA hsa-miR-340-5p in IAV-Infected A549 Cells Suppresses Viral Replication by Targeting RIG-I and OAS2. Mol. Ther. Nucleic Acids 2019, 14, 509–519. [Google Scholar] [CrossRef]
- Martinez, M.A.; Tural, C.; Franco, S. Circulating MicroRNAs as a Tool for Diagnosis of Liver Disease Progression in People Living with HIV-1. Viruses 2022, 14, 1118. [Google Scholar] [CrossRef]
- Martínez-Fleta, P.; Vera-Tomé, P.; Jiménez-Fernández, M.; Requena, S.; Roy-Vallejo, E.; Sanz-García, A.; Lozano-Prieto, M.; López-Sanz, C.; Vara, A.; Lancho-Sánchez, Á.; et al. A Differential Signature of Circulating miRNAs and Cytokines between COVID-19 and Community-Acquired Pneumonia Uncovers Novel Physiopathological Mechanisms of COVID-19. Front. Immunol. 2022, 12, 815651. [Google Scholar] [CrossRef]
- Farr, R.J.; Rootes, C.L.; Rowntree, L.C.; Nguyen, T.H.O.; Hensen, L.; Kedzierski, L.; Cheng, A.C.; Kedzierska, K.; Au, G.G.; Marsh, G.A.; et al. Altered microRNA expression in COVID-19 patients enables identification of SARS-CoV-2 infection. PLoS Pathog. 2021, 17, e1009759. [Google Scholar] [CrossRef]
- Morales, L.; Oliveros, J.C.; Enjuanes, L.; Sola, I. Contribution of Host miRNA-223-3p to SARS-CoV-Induced Lung Inflammatory Pathology. MBio 2022, 13, e0313521. [Google Scholar] [CrossRef]
- Ying, H.; Ebrahimi, M.; Keivan, M.; Khoshnam, S.E.; Salahi, S.; Farzaneh, M. miRNAs; a novel strategy for the treatment of COVID-19. Cell Biol. Int. 2021, 45, 2045–2053. [Google Scholar] [CrossRef] [PubMed]
- Trinité, B.; Tarrés-Freixas, F.; Rodon, J.; Pradenas, E.; Urrea, V.; Marfil, S.; Rodríguez de la Concepción, M.L.; Ávila-Nieto, C.; Aguilar-Gurrieri, C.; Barajas, A.; et al. SARS-CoV-2 infection elicits a rapid neutralizing antibody response that correlates with disease severity. Sci. Rep. 2021, 11, 2608. [Google Scholar] [CrossRef] [PubMed]
- Franco, S.; Buccione, D.; Pluvinet, R.; Mothe, B.; Ruiz, L.; Nevot, M.; Jordan-Paiz, A.; Ramos, L.; Aussó, S.; Morillas, R.M.; et al. Large-scale screening of circulating microRNAs in individuals with HIV-1 mono-infections reveals specific liver damage signatures. Antiviral Res. 2018, 155, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Franco, S.; Buccione, D.; Tural, C.; Martinez, M.A. Circulating microRNA signatures that predict liver fibrosis progression in patients with HIV-1/hepatitis C virus coinfections. AIDS 2021, 35, 1355–1363. [Google Scholar] [CrossRef] [PubMed]
- Mellado-Artigas, R.; Ferreyro, B.L.; Angriman, F.; Hernández-Sanz, M.; Arruti, E.; Torres, A.; Villar, J.; Brochard, L.; Ferrando, C.; Mellado-Artigas, R.; et al. High-flow nasal oxygen in patients with COVID-19-associated acute respiratory failure. Crit. Care 2021, 25, 58. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; He, L.; Yang, Z.; Jia, N.; Chen, R.; Xie, J.; Fu, W.; Chen, H.; Lin, X.; Huang, R.; et al. Identification of Parameters Representative of Immune Dysfunction in Patients with Severe and Fatal COVID-19 Infection: A Systematic Review and Meta-analysis. Clin. Rev. Allergy Immunol. 2023, 64, 33–65. [Google Scholar] [CrossRef] [PubMed]
- Gjorgjieva, M.; Sobolewski, C.; Dolicka, D.; Correia De Sousa, M.; Foti, M. miRNAs and NAFLD: From pathophysiology to therapy. Gut 2019, 68, 2065–2079. [Google Scholar] [CrossRef]
- Franco, S.; Llibre, J.M.; Jou, T.; Tural, C.; Martínez, M.A. Normalization of circulating plasma levels of miRNAs in HIV-1/HCV co-infected patients following direct-acting antiviral-induced sustained virologic response. Heliyon 2023, 9, e12686. [Google Scholar] [CrossRef]
- Forloni, M.; Dogra, S.K.; Dong, Y.; Conte, D.; Ou, J.; Zhu, L.J.; Deng, A.; Mahalingam, M.; Green, M.R.; Wajapeyee, N. miR-146a promotes the initiation and progression of melanoma by activating Notch signaling. Elife 2014, 2014, e01460. [Google Scholar] [CrossRef]
- Pinacchio, C.; Scordio, M.; Santinelli, L.; Frasca, F.; Sorrentino, L.; Bitossi, C.; Oliveto, G.; Viscido, A.; Ceci, F.M.; Celani, L.; et al. Analysis of serum microRNAs and rs2910164 GC single-nucleotide polymorphism of miRNA-146a in COVID-19 patients. J. Immunoass. Immunochem. 2022, 43, 347–364. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Fan, L.; Mou, J.; Yin, Y.; Peng, C.; Chen, Y.; Lu, H.; Zhao, L.; Tao, Z.; et al. MiR-629-5p promotes the invasion of lung adenocarcinoma via increasing both tumor cell invasion and endothelial cell permeability. Oncogene 2020, 39, 3473–3488. [Google Scholar] [CrossRef] [PubMed]
- Khodadadi-Jamayran, A.; Akgol-Oksuz, B.; Afanasyeva, Y.; Heguy, A.; Thompson, M.; Ray, K.; Giro-Perafita, A.; Sánchez, I.; Wu, X.; Tripathy, D.; et al. Prognostic role of elevated mir-24-3p in breast cancer and its association with the metastatic process. Oncotarget 2018, 9, 12868–12878. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, N.; Kulshreshtha, R. miR-191: An emerging player in disease biology. Front. Genet. 2014, 5, 99. [Google Scholar] [CrossRef]
- Lian, J.; Zhu, X.; Du, J.; Huang, B.; Zhao, F.; Ma, C.; Guo, R.; Zhang, Y.; Ji, L.; Yahaya, B.H.; et al. Extracellular vesicle-transmitted miR-671-5p alleviates lung inflammation and injury by regulating the AAK1/NF-κB axis. Mol. Ther. 2023, 31, 1365–1382. [Google Scholar] [CrossRef] [PubMed]
- Greco, F.; Lorefice, E.; Carissimi, C.; Laudadio, I.; Ciccosanti, F.; Di Rienzo, M.; Colavita, F.; Meschi, S.; Maggi, F.; Fimia, G.M.; et al. A microRNA Arising from the Negative Strand of SARS-CoV-2 Genome Targets FOS to Reduce AP-1 Activity. Non-Coding RNA 2023, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Pawlica, P.; Yario, T.A.; White, S.; Wang, J.; Moss, W.N.; Hui, P.; Vinetz, J.M.; Steitz, J.A. SARS-CoV-2 expresses a microRNA-like small RNA able to selectively repress host genes. Proc. Natl. Acad. Sci. USA 2021, 118, e2116668118. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Giralt, N.; Du, J.; Marin-Corral, J.; Bódalo-Torruella, M.; Blasco-Hernando, F.; Muñoz-Bermúdez, R.; Clarós, M.; Nonell, L.; Perera-Bel, J.; Fernandez-González, M.; et al. Circulating microRNA profiling is altered in the acute respiratory distress syndrome related to SARS-CoV-2 infection. Sci. Rep. 2022, 12, 6926. [Google Scholar] [CrossRef] [PubMed]
- Giannella, A.; Riccetti, S.; Sinigaglia, A.; Piubelli, C.; Razzaboni, E.; Di Battista, P.; Agostini, M.; Dal Molin, E.; Manganelli, R.; Gobbi, F.; et al. Circulating microRNA signatures associated with disease severity and outcome in COVID-19 patients. Front. Immunol. 2022, 13, 968991. [Google Scholar] [CrossRef]
- Grehl, C.; Schultheiß, C.; Hoffmann, K.; Binder, M.; Altmann, T.; Grosse, I.; Kuhlmann, M. Detection of SARS-CoV-2 Derived Small RNAs and Changes in Circulating Small RNAs Associated with COVID-19. Viruses 2021, 13, 1593. [Google Scholar] [CrossRef]
- Meidert, A.S.; Hermann, S.; Brandes, F.; Kirchner, B.; Buschmann, D.; Billaud, J.N.; Klein, M.; Lindemann, A.; Aue, E.; Schelling, G.; et al. Extracellular Vesicle Associated miRNAs Regulate Signaling Pathways Involved in COVID-19 Pneumonia and the Progression to Severe Acute Respiratory Corona Virus-2 Syndrome. Front. Immunol. 2021, 12, 784028. [Google Scholar] [CrossRef]
- Zeng, Q.; Qi, X.; Ma, J.; Hu, F.; Wang, X.; Qin, H.; Li, M.; Huang, S.; Yang, Y.; Li, Y.; et al. Distinct miRNAs associated with various clinical presentations of SARS-CoV-2 infection. iScience 2022, 25, 104309. [Google Scholar] [CrossRef]
- Willan, J.; Agarwal, G.; Bienz, N. Mortality and burden of post-COVID-19 syndrome have reduced with time across SARS-CoV-2 variants in haematology patients. Br. J. Haematol. 2023, 201, 640–644. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| SARS-CoV-2 Infected Patients | Uninfected Individuals | p-Value | |
|---|---|---|---|
| N | 40 | 21 | - |
| Age yr, median (IQR) a | 56 (50.25–64) | 37 (35–44.5) | <0.0001 |
| Female no. (%) b | 14 (35) | 15 (71.4) | 0.0068 |
| Albumin (g/L), median (IQR) a | 34.95 (31.10–37.78) | - | - |
| Bilirubin (mg/dL), median (IQR) a | 0.54 (0.42–0.64) | 0.47 (0.37–0.74) | 0.5944 |
| Creatinine mg/dL, median (IQR) a | 0.75 (0.66–0.89) | 0.78 (0.68–0.90) | 0.7790 |
| D-Dimer (µg/L), median (IQR) a | 453 (217–809) | - | - |
| Ferritin (ng/mL), median (IQR) a | 671 (315.1–1035) | - | - |
| Fibrinogen (mg/L), median (IQR) a | 654 (540–726) | - | - |
| Glucose (mg/dL), median (IQR) a | 102.20 (90.33–115) | - | - |
| ALT (U/L), median (IQR) a | 32 (19.75–58.50) | 15 (12–19.50) | <0.0001 |
| Potassium (mmol/L), median (IQR) a | 4.00 (3.71–4.36) | - | - |
| Sodium (mmol/L), median (IQR) a | 139.10 (136.80–141) | - | - |
| Urea (mg/dL), median (IQR) a | 32 (25–49) | - | - |
| Hematocrit %, median (IQR) a | 39.60 (34.60–44.90) | 41.25 (38.40–43.63) | 0.4708 |
| Hemoglobin (g/dL), median (IQR) a | 13.40 (11.93–15.48) | 14.20 (13.05–14.53) | 0.4353 |
| Mean Corpuscular Hemoglobin Concentration (g/dL), median (IQR) a | 30.20 (29.25–31.35) | 30.35 (29.65–31.93) | 0.4204 |
| Leucocyte count (×109/L), median (IQR) a | 5.50 (4.12–7.45) | 6.50 (5.70–8.92) | 0.0752 |
| Lymphocyte count (×109/L), median (IQR) a | 1.25(0.87–1.60) | 2.00 (1.77–2.67) | <0.0001 |
| Monocyte count (×109/L), median (IQR) a | 0.45 (0.30–0.70) | 0.50 (0.37–0.80) | 0.3963 |
| Platelet Distribution Width %, median (IQR) a | 17.20 (16.63–17.60) | 16.10 (15.78–16.55) | <0.0001 |
| Platelet (×109/L), median (IQR) a | 214 (169.30–301.50) | 253.50 (195.30–347.80) | 0.1798 |
| Interleukin 6 (pg/mL), median (IQR) a | 20.06 (9.53–52.69) | - | - |
| High Flow | Low Flow | No Oxygen | p-Value a | p-Value b | p-Value c | |
|---|---|---|---|---|---|---|
| N (%) | 11 (27.50) | 23 (57.5) | 6 (15) | - | - | - |
| Age, yr, median (IQR) d | 56 (44–65) | 58 (51–64) | 52 (35–58) | 0.6174 | 0.362 | 0.0866 |
| Female sex (%) e | 4 (36.40) | 8 (34.8) | 2 (33.33) | 0.9281 | 0.9006 | 0.9470 |
| Baseline SaO2 (%), median (IQR) d | 94 (91–96) | 94(93–96) | 96 (95.5–97) | 0.4683 | 0.0376 | 0.0991 |
| Baseline FIO2, median (IQR) d | 0.21 (0.21–0.21) | 0.21 (0.21–0.21) | 0.21 (0.21–0.21) | 0.0980 | 0.5147 | >0.9999 |
| Baseline ABG PaO2 (mmHg), median (IQR) d | 52 (49–69) | 69.5 (62.5–76.25) | 76 (71–86) | 0.0807 | 0.0353 | 0.1247 |
| Baseline ABG FIO2, median (IQR) d | 0.21 (0.21–0.24) | 0.21 (0.21–0.21) | 0.21 (0.21–0.21) | 0.0891 | 0.5089 | >0.9999 |
| Baseline ABG PaO2/FIO2, median (IQR) d | 246 (158–328.6) | 317 (283.8–363.1) | 361.9 (338–409) | 0.0365 | 0.038 | 0.1091 |
| 48 h SaO2 (%), median (IQR) d | 95 (92–96) | 96 (95–97) | 96 (93.5–97.75) | 0.0693 | 0.3848 | 0.6678 |
| 48 h FIO2, median (IQR) d | 0.4 (0.28–0.5) | 0.28 (0.275–0.32) | 0.21 (0.21–0.2275) | 0.0171 | 0.0006 | 0.0058 |
| 48 h ABG PaO2 (mmHg), median (IQR) d | 75 (69–90) | 79 (75–104) | 74 (64–107) | 0.3681 | >0.9999 | 0.4982 |
| 48 h ABG FIO2, median (IQR) d | 0.4 (0.28–0.8) | 0.28 (0.225–0.32) | 0.21 (0.21–0.21) | 0.0939 | 0.0165 | 0.0571 |
| 48 h ABG PaO2/FIO2, median (IQR) d | 194 (118–314) | 304 (237–382.5) | 352 (304–510) | 0.0257 | 0.0385 | 0.2839 |
| MDW (U), median (IQR) d | 22.24 (20.33–26.48) | 23.76 (21–28.17) | 24.57 (21.18–25.55) | 0.3171 | 0.5035 | 0.7455 |
| aPTT (s), median (IQR) d | 12.9 (11.7–14.15) | 12.5 (11.8–13.2) | 12.9 (12.28–14.08) | 0.5675 | 0.7714 | 0.2284 |
| aPTT (%), median (IQR) d | 78.5 (69–89.25) | 82 (74–88) | 77.5 (70.5–82.25) | 0.6522 | 0.7717 | 0.1876 |
| PT/INR, median (IQR) d | 1.21 (1.07–1.305) | 1.15 (1.088–1.223) | 1.29 (1.16–1.31) | 0.5452 | 0.6273 | 0.1059 |
| pH, median (IQR) d | 7.44 (7.435–7.47) | 7.45 (7.4–7.475) | 7.43 (7.365–7.465) | 0.6684 | 0.3776 | 0.4402 |
| Partial Pressure CO2 (mmHg), median (IQR) d | 43 (36.5–83) | 38 (35.5–47.5) | 35.5 (34.25–47.25) | 0.3203 | 0.2567 | 0.4605 |
| Partial Pressure O2 (mmHg), median (IQR) d | 73.5 (41.5–83.5) | 69 (36–88) | 90 (37.25–132.3) | 0.7161 | 0.4356 | 0.4101 |
| HCO3 (mmol/L), median (IQR)d | 26.3 (23.5–29.38) | 26 (24.05–27.75) | 24.9 (22.55–27.7) | 0.7313 | 0.4336 | 0.5992 |
| Base excess (mmol/L), median (IQR) d | 3.2 (0.05–4.7) | 2.1 (0.05–3.35) | 0.6 (−1.375–2.275) | 0.4220 | 0.1878 | 0.2553 |
| O2 Saturation (%), median (IQR) d | 97.6 (96.18–99.7) | 95.6 (78.2–98.3) | 97.5 (35.4–98.2) | 0.0302 | 0.5035 | >0.9999 |
| High Flow | Low Flow | No Oxygen | p-Value a | p-Value b | p-Value c | |
|---|---|---|---|---|---|---|
| N (%) | 11 (27.50) | 23 (57.50) | 6 (15) | - | - | - |
| Age yr, median (IQR) d | 56 (44–65) | 58 (51–64) | 52 (35–58) | 0.6174 | 0.3620 | 0.0866 |
| Female sex no. (%) e | 4 (36.40) | 8 (34.80) | 2 (33.33) | 0.9281 | 0.9006 | 0.9470 |
| Albumin (g/L), median (IQR)d | 35.9 (25.75–42.85) | 33.8 (30.23–36.2) | 36.40 (32.90–36.90) | 0.6778 | >0.9999 | 0.3728 |
| Bilirubin (mg/dL), median (IQR) d | 0.63 (0.54–0.7) | 0.54 (0.41–0.61) | 0.46 (0.35–0.55) | 0.1058 | 0.0513 | 0.3789 |
| Creatinine mg/dL, median (IQR) d | 0.71 (0.66–0.88) | 0.75 (0.66–1.10) | 0.77 (0.57–0.92) | 0.5693 | 0.9578 | 0.8031 |
| D-Dimer (µg/L), median (IQR) d | 704 (365–1546) | 447 (266.5–764.8) | 386.50(177.50–1415) | 0.2513 | 0.5556 | 0.8272 |
| Ferritin (ng/mL), median (IQR) d | 501 (375–901) | 654 (309–994) | 1005 (445.80–2025) | 0.9543 | 0.3290 | 0.2300 |
| Fibrinogen (mg/L), median (IQR) d | 619 (487–699) | 598 (480–726) | 685 (585.30–787.80) | 0.7209 | 0.2635 | 0.2361 |
| Glucose (mg/dL), median (IQR) d | 102.60 (88.45–131.60) | 102.50 (91–120.30) | 96.90 (82.93–142.30) | >0.9999 | 0.4559 | 0.4831 |
| ALT (U/L), median (IQR) d | 30 (20.25–57.50) | 32 (15.78–69.50) | 45 (25.50–68.03) | 0.9741 | 0.5440 | 0.5618 |
| Potassium (mmol/L), median (IQR) d | 3.99 (3.79–4.28) | 4.09 (3.71–4.43) | 3.76 (3.53–4.12) | 0.7265 | 0.2198 | 0.1018 |
| Sodium (mmol/L), median (IQR) d | 138.10 (136.20–141.20) | 139.50 (136.70–141.20) | 138.70 (136.50–139.20) | 0.7921 | 0.7472 | 0.2595 |
| Urea (mg/dL), median (IQR) d | 33.50 (30.50–49.75) | 31.95 (24.75–52) | 26.10 (19.50–33.45) | 0.6372 | 0.1274 | 0.1606 |
| Hematocrit %, median (IQR) d | 44.90 (33–45.90) | 38.70 (34.50–43.50) | 39.75 (36.63–43.08) | 0.3876 | 0.5740 | 0.6489 |
| Hemoglobin (g/dL), median (IQR) d | 15.40 (11.10–15.90) | 12.90 (11.90–14.50) | 13.60 (12.25–14.95) | 0.4397 | 0.6416 | 0.4855 |
| Mean Corpuscular Hemoglobin Concentration (g/dL), median (IQR) d | 34 (33.60–34.70) | 34.10 (33.40–34.50) | 34.25 (33.73–34.65) | 0.6303 | 0.7893 | 0.5190 |
| Leucocyte count (×109/L), median (IQR) d | 6.90 (4.10–8.70) | 5.40 (4.20–7.50) | 4.75 (3.67–6.95) | 0.4183 | 0.2657 | 0.4214 |
| Lymphocyte count (×109/L), median (IQR) d | 1.30 (0.85–1.62) | 1.25 (0.85–1.67) | 1.35 (0.72–1.77) | >0.9999 | 0.9800 | 0.9239 |
| Monocyte count (×109/L), median (IQR) d | 0.70 (0.37–0.80) | 0.45 (0.27–0.60) | 0.40 (0.27–0.62) | 0.0803 | 0.1485 | 0.8995 |
| Platelet Distribution Width %, median (IQR) d | 17.40 (17.10–17.60) | 17.20 (16.60–17.60) | 16.90 (16.23–18) | 0.2234 | 0.4461 | 0.9446 |
| Platelet (×109/L), median (IQR) d | 261 (145–316) | 214 (168.30–303) | 210.50 (175.30–222.30) | 0.7108 | 0.3676 | 0.5003 |
| Interleukin 6 (pg/mL), median (IQR) d | 26.60 (12.03–63.91) | 19.10 (6.98–48.90) | 19.06 (15.07–46.95) | 0.3725 | >0.9999 | 0.5808 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franco, S.; Mateu, L.; Pluvinet, R.; Sanchez-Herrero, J.F.; Toledo, R.; Sumoy, L.; Massanella, M.; Martinez, M.A. Altered Plasma microRNA Signature in Hospitalized COVID-19 Patients Requiring Oxygen Support. Microorganisms 2024, 12, 440. https://doi.org/10.3390/microorganisms12030440
Franco S, Mateu L, Pluvinet R, Sanchez-Herrero JF, Toledo R, Sumoy L, Massanella M, Martinez MA. Altered Plasma microRNA Signature in Hospitalized COVID-19 Patients Requiring Oxygen Support. Microorganisms. 2024; 12(3):440. https://doi.org/10.3390/microorganisms12030440
Chicago/Turabian StyleFranco, Sandra, Lourdes Mateu, Raquel Pluvinet, Jose Francisco Sanchez-Herrero, Ruth Toledo, Lauro Sumoy, Marta Massanella, and Miguel Angel Martinez. 2024. "Altered Plasma microRNA Signature in Hospitalized COVID-19 Patients Requiring Oxygen Support" Microorganisms 12, no. 3: 440. https://doi.org/10.3390/microorganisms12030440
APA StyleFranco, S., Mateu, L., Pluvinet, R., Sanchez-Herrero, J. F., Toledo, R., Sumoy, L., Massanella, M., & Martinez, M. A. (2024). Altered Plasma microRNA Signature in Hospitalized COVID-19 Patients Requiring Oxygen Support. Microorganisms, 12(3), 440. https://doi.org/10.3390/microorganisms12030440