
The Multiple Roles of LCCL Domain-Containing Proteins for Malaria Parasite Transmission


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Structure and Origin of LCCL Modules
3. Architecture of the Plasmodial LCCL Domain-Containing Proteins
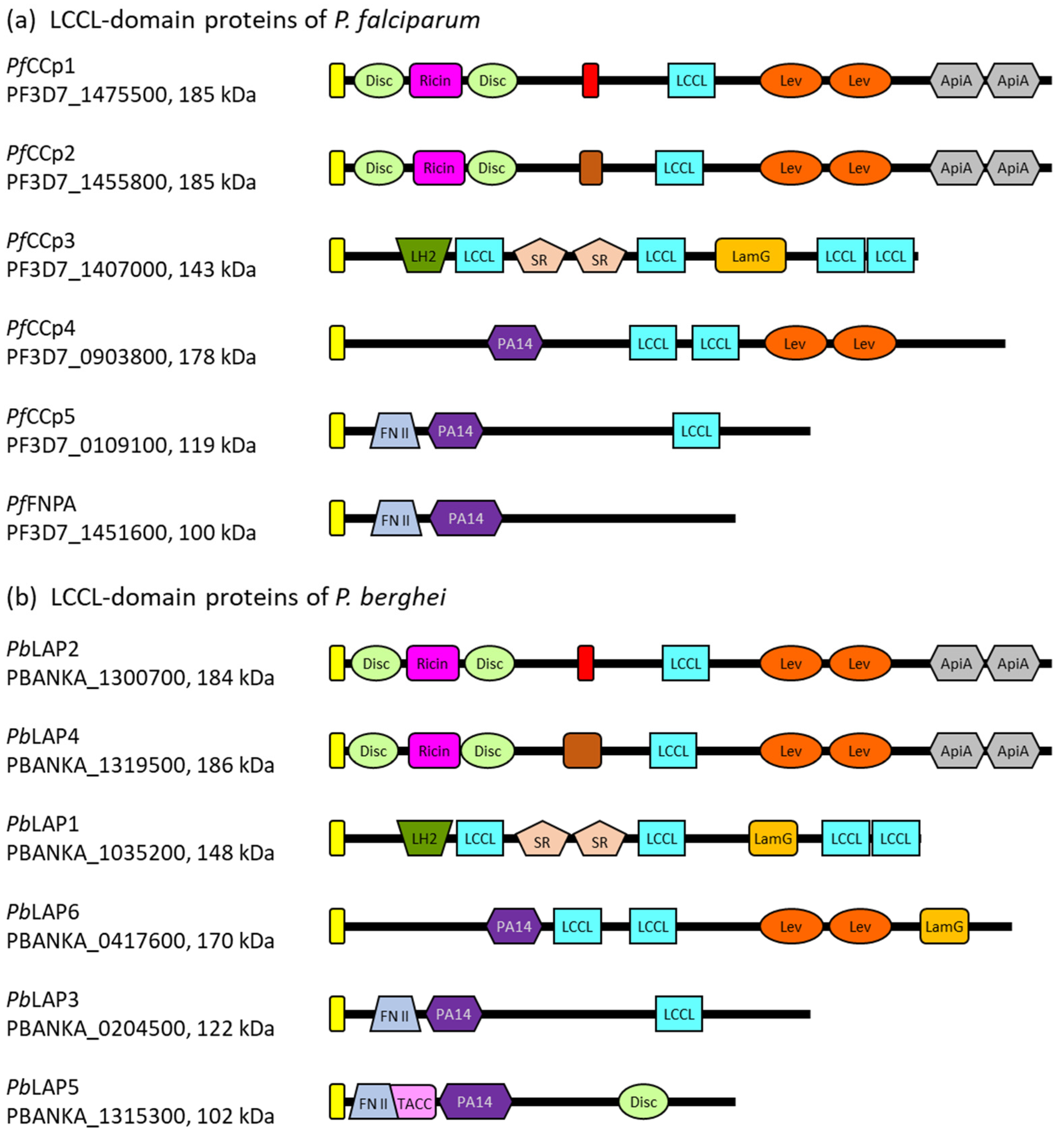
4. Expression of the Plasmodial LCCL Domain-Containing Proteins in the Transmission Stages
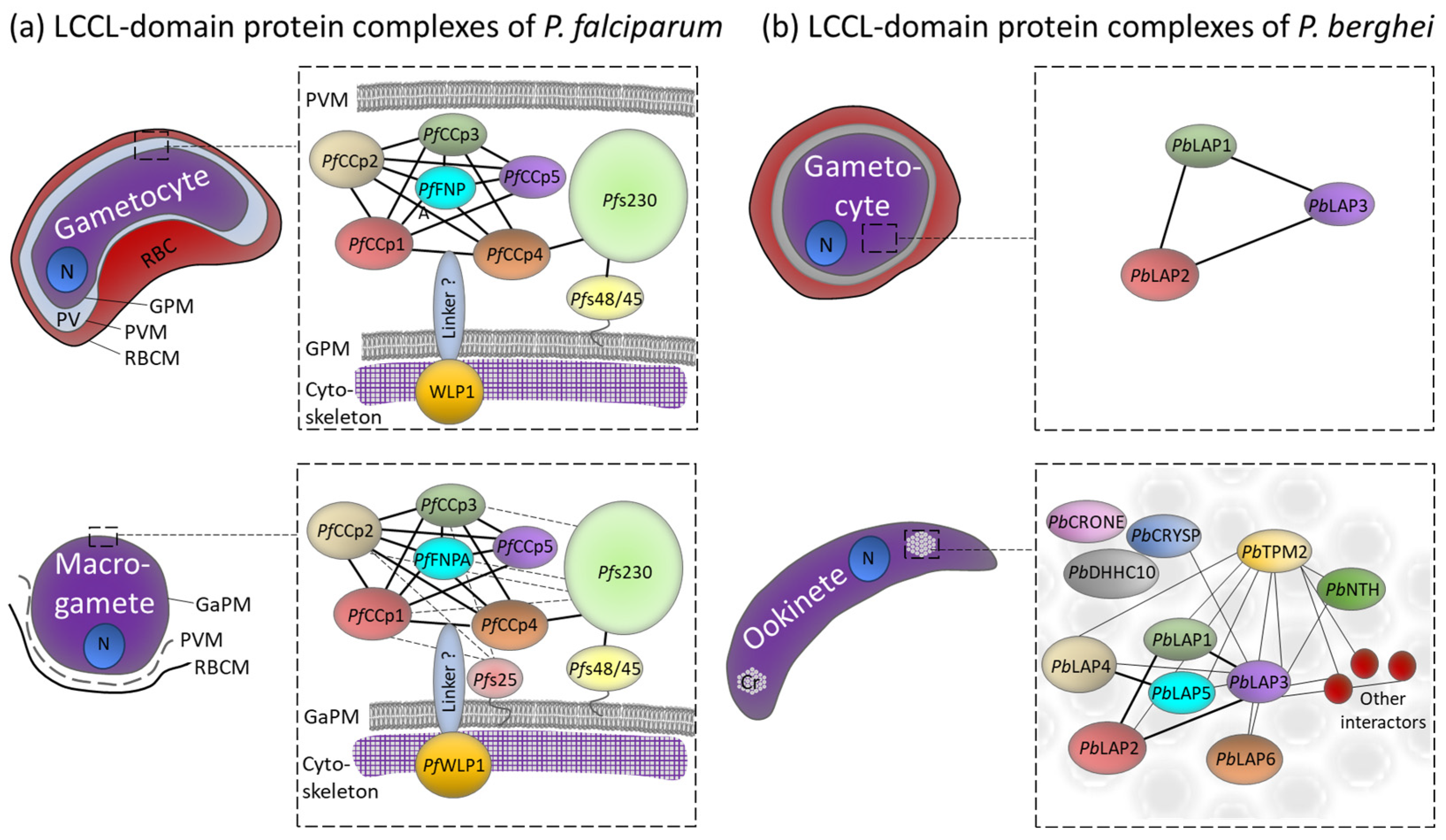
4.1. LCCL Domain-Containing Protein Complexes in P. falciparum Gametocytes
4.2. Aggregation of the LCCL Domain-Containing Proteins in the Transmission Stages of P. berghei
5. The Role of the Plasmodial LCCL Domain-Containing Proteins during Malaria Transmission
6. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- WHO (World Health Organisation). World Malaria Report 2022; WHO: Geneva, Switzerland, 2022; ISBN 9789240040496. [Google Scholar]
- Poespoprodjo, J.R.; Douglas, N.M.; Ansong, D.; Kho, S.; Anstey, N.M. Malaria. Lancet 2023, 402, 2328–2345. [Google Scholar] [CrossRef]
- Bennink, S.; Kiesow, M.J.; Pradel, G. The Development of Malaria Parasites in the Mosquito Midgut. Cell. Microbiol. 2016, 18, 905–918. [Google Scholar] [CrossRef]
- Josling, G.A.; Williamson, K.C.; Llinás, M. Regulation of Sexual Commitment and Gametocytogenesis in Malaria Parasites. Annu. Rev. Microbiol. 2018, 72, 501–519. [Google Scholar] [CrossRef]
- Bennink, S.; Pradel, G. Vesicle Dynamics during the Egress of Malaria Gametocytes from the Red Blood Cell. Mol. Biochem. Parasitol. 2021, 243, 111372. [Google Scholar] [CrossRef]
- Trexler, M.; Bányai, L.; Patthy, L. The LCCL Module. Eur. J. Biochem. 2000, 267, 5751–5757. [Google Scholar] [CrossRef]
- Nakamura, T.; Tokunaga, F.; Morita, T.; Iwanaga, S.; Kusumoto, S.; Shiba, T.; Kobayashi, T.; Inoue, K. Intracellular Serine-Protease Zymogen, Factor C, from Horseshoe Crab Hemocytes Its Activation by Synthetic Lipid A Analogues and Acidic Phospholipids. Eur. J. Biochem. 1988, 176, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Tokunaga, F.; Morita, T.; Iwanaga, S. Interaction between Lipopolysaccharide and Intracellular Serine Protease Zymogen, Factor C, from Horseshoe Crab (Tachypleus tridentatus) Hemocytes. J. Biochem. 1988, 103, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Muta, T.; Miyata, T.; Misumi, Y.; Tokunaga, F.; Nakamura, T.; Toh, Y.; Ikehara, Y.; Iwanaga, S. Limulus Factor C. An Endotoxin-Sensitive Serine Protease Zymogen with a Mosaic Structure of Complement-like, Epidermal Growth Factor-like, and Lectin-like Domains. J. Biol. Chem. 1991, 266, 6554–6561. [Google Scholar] [CrossRef] [PubMed]
- Robertson, N.G.; Skvorak, A.B.; Yin, Y.; Weremowicz, S.; Johnson, K.R.; Kovatch, K.A.; Battey, J.F.; Bieber, F.R.; Morton, C.C. Mapping and Characterization of a Novel Cochlear Gene in Human and in Mouse: A Positional Candidate Gene for a Deafness Disorder, DFNA9. Genomics 1997, 46, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Yoo, J.E.; Choe, Y.H.; Park, S.C.; Lee, H.J.; Lee, H.J.; Noh, B.; Kim, S.H.; Kang, G.-Y.; Lee, K.-M.; et al. Cleaved Cochlin Sequesters Pseudomonas aeruginosa and Activates Innate Immunity in the Inner Ear. Cell Host Microbe 2019, 25, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Verdoodt, D.; Van Camp, G.; Ponsaerts, P.; Van Rompaey, V. On the Pathophysiology of DFNA9: Effect of Pathogenic Variants in the COCH Gene on Inner Ear Functioning in Human and Transgenic Mice. Hear. Res. 2021, 401, 108162. [Google Scholar] [CrossRef]
- Bae, S.-H.; Robertson, N.G.; Cho, H.-J.; Morton, C.C.; Jung, D.J.; Baek, J.-I.; Choi, S.-Y.; Lee, J.; Lee, K.-Y.; Kim, U.-K. Identification of Pathogenic Mechanisms of COCH Mutations, Abolished Cochlin Secretion, and Intracellular Aggregate Formation: Genotype-Phenotype Correlations in DFNA9 Deafness and Vestibular Disorder Formation and Retention of Dimers and Large Multimeric. Hum. Mutat. 2014, 35, 1506–1513. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.H.; Yoo, J.E.; Hong, J.W.; Park, H.R.; Noh, B.; Kim, H.; Kang, M.; Hyun, Y.-M.; Gee, H.Y.; Choi, J.Y.; et al. LCCL Peptide Cleavage after Noise Exposure Exacerbates Hearing Loss and Is Associated with the Monocyte Infiltration in the Cochlea. Hear. Res. 2021, 412, 108378. [Google Scholar] [CrossRef] [PubMed]
- Py, B.F.; Gonzalez, S.F.; Long, K.; Kim, M.-S.; Kim, Y.-A.; Zhu, H.; Yao, J.; Degauque, N.; Villet, R.; Ymele-Leki, P.; et al. Cochlin Produced by Follicular Dendritic Cells Promotes Antibacterial Innate Immunity. Immunity 2013, 38, 1063–1072. [Google Scholar] [CrossRef]
- Zhang, H.; Sweezey, N.B.; Kaplan, F. LGL1 Modulates Proliferation, Apoptosis, and Migration of Human Fetal Lung Fibroblasts. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 308, L391–L402. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-Q.; Xing, W.-M.; Fan, H.-H.; Wang, K.-S.; Zhang, H.-K.; Wang, Q.-W.; Qi, J.; Yang, H.-M.; Yang, J.; Ren, Y.-N.; et al. The Novel Lipopolysaccharide-Binding Protein CRISPLD2 Is a Critical Serum Protein to Regulate Endotoxin Function 1. J. Immunol. 2009, 183, 6646–6656. [Google Scholar] [CrossRef] [PubMed]
- Vásárhelyi, V.; Trexler, M.; Patthy, L. Both LCCL-Domains of Human CRISPLD2 Have High Affinity for Lipid A. Biochimie 2014, 97, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Claudianos, C.; Dessens, J.T.; Trueman, H.E.; Arai, M.; Mendoza, J.; Butcher, G.A.; Crompton, T.; Sinden, R.E. A Malaria Scavenger Receptor-like Protein Essential for Parasite Development. Mol. Microbiol. 2002, 45, 1473–1484. [Google Scholar] [CrossRef] [PubMed]
- Delrieu, I.; Waller, C.C.; Mota, M.M.; Grainger, M.; Langhorne, J.; Holder, A.A. PSLAP, a Protein with Multiple Adhesive Motifs, Is Expressed in Plasmodium falciparum Gametocytes. Mol. Biochem. Parasitol. 2002, 121, 11–20. [Google Scholar] [CrossRef]
- Pradel, G.; Hayton, K.; Aravind, L.; Iyer, L.M.; Abrahamsen, M.S.; Bonawitz, A.; Mejia, C.; Templeton, T.J. A Multidomain Adhesion Protein Family Expressed in Plasmodium falciparum Is Essential for Transmission to the Mosquito. J. Exp. Med. 2004, 199, 1533–1544. [Google Scholar] [CrossRef]
- Templeton, T.J.; Iyer, L.M.; Anantharaman, V.; Enomoto, S.; Abrahante, J.E.; Subramanian, G.M.; Hoffman, S.L.; Abrahamsen, M.S.; Aravind, L. Comparative Analysis of Apicomplexa and Genomic Diversity in Eukaryotes. Genome Res. 2004, 14, 1686–1695. [Google Scholar] [CrossRef]
- Tosini, F.; Agnoli, A.; Mele, R.; Gomez Morales, M.A.; Pozio, E. A New Modular Protein of Cryptosporidium parvum, with Ricin B and LCCL Domains, Expressed in the Sporozoite Invasive Stage. Mol. Biochem. Parasitol. 2004, 134, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Trueman, H.E.; Raine, J.D.; Florens, L.; Dessens, J.T.; Mendoza, J.; Johnson, J.; Waller, C.C.; Delrieu, I.; Holder, A.A.; Langhorne, J.; et al. Functional Characterization of an LCCL-Lectin Domain Containing Protein Family in Plasmodium berghei. J. Parasitol. 2004, 90, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Ozubek, S.; Alzan, H.F.; Bastos, R.G.; Laughery, J.M.; Suarez, C.E. Identification of CCp5 and FNPA as Novel Non-Canonical Members of the CCp Protein Family in Babesia bovis. Front. Vet. Sci. 2022, 9, 833183. [Google Scholar] [CrossRef] [PubMed]
- Wickstead, B.; Ersfeld, K.; Gull, K. Repetitive Elements in Genomes of Parasitic Protozoa. Microbiol. Mol. Biol. Rev. 2003, 67, 360–375. [Google Scholar] [CrossRef] [PubMed]
- Dessens, J.T.; Sinden, R.E.; Claudianos, C. LCCL Proteins of Apicomplexan Parasites. Trends Parasitol. 2004, 20, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Pradel, G.; Templeton, T.J. Genomics of Pathogenic Parasites. In Pathogenomics; Hacker, J., Dobrindt, U., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2006; pp. 417–444. [Google Scholar]
- Rutenber, E.; Ready, M.; Robertus, J.D. Structure and Evolution of Ricin B Chain. Nature 1987, 326, 624–626. [Google Scholar] [CrossRef] [PubMed]
- Kane, W.H.; Davie, E.W. Blood Coagulation Factors V and VIII: Structural and Functional Similarities and Their Relationship to Hemorrhagic and Thrombotic Disorders. Blood 1988, 71, 539–555. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, S.; Hofmann, K.; Chiquet-Ehrismann, R.; Bucher, P. The Discoidin Domain Family Revisited: New Members from Prokaryotes and a Homology-Based Fold Prediction. Protein Sci. 1998, 7, 1626–1631. [Google Scholar] [CrossRef]
- Aurrecoechea, C.; Brestelli, J.; Brunk, B.P.; Dommer, J.; Fischer, S.; Gajria, B.; Gao, X.; Gingle, A.; Grant, G.; Harb, O.S.; et al. PlasmoDB: A Functional Genomic Database for Malaria Parasites. Nucleic Acids Res. 2009, 37, D539–D543. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The Conserved Domain Database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- Tremp, A.Z.; Sharma, V.; Carter, V.; Lasonder, E.; Dessens, J.T. LCCL Protein Complex Formation in Plasmodium Is Critically Dependent on LAP1. Mol. Biochem. Parasitol. 2017, 214, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Sandford, R. The PLAT Domain: A New Piece in the PKD1 Puzzle. Curr. Biol. 1999, 9, R588–R590. [Google Scholar] [CrossRef] [PubMed]
- Hohenester, E.; Sasaki, T.; Timpl, R. Crystal Structure of a Scavenger Receptor Cysteine-Rich Domain Sheds Light on an Ancient Superfamily. Nat. Struct. Biol. 1999, 6, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Resnick, D.; Pearson, A.; Krieger, M. The SRCR Superfamily: A Family Reminiscent of the Ig Superfamily. Trends Biochem. Sci. 1994, 19, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Vuolteenaho, R.; Chow, L.T.; Tryggvason, K. Structure of the Human Laminin Bl Chain Gene. J. Biol. Chem. 1990, 265, 15611–15616. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, N.; White, H.E.; Emsley, J.; Wood, S.P.; Pepys, M.B.; Blundell, T.L. Comparative Analyses of Pentraxins: Implications for Protomer Assembly and Ligand Binding. Structure 1994, 2, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Gewurz, H.; Zhang, X.-H.; Lint, T.F. Structure and Function of the Pentraxins. Curr. Opin. Immunol. 1995, 7, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, A.; Simon, N.; Pradel, G. Family Members Stick Together: Multi-Protein Complexes of Malaria Parasites. Med. Microbiol. Immunol. 2010, 199, 209–226. [Google Scholar] [CrossRef]
- Rigden, D.J.; Mello, L.V.; Galperin, M.Y. The PA14 Domain, a Conserved All-β Domain in Bacterial Toxins, Enzymes, Adhesins and Signaling Molecules. Trends Biochem. Sci. 2004, 29, 335–339. [Google Scholar] [CrossRef]
- Gergely, F.; Karlsson, C.; Still, I.; Cowell, J.; Kilmartin, J.; Raff, J.W. The TACC Domain Identifies a Family of Centrosomal Proteins That Can Interact with Microtubules. Proc. Natl. Acad. Sci. USA 2000, 97, 14352–14357. [Google Scholar] [CrossRef]
- Lasonder, E.; Ishihama, Y.; Andersen, J.S.; Vermunt, A.M.W.; Pain, A.; Sauerwein, R.W.; Eling, W.M.C.; Hall, N.; Waters, A.P.; Stunnenberg, H.G.; et al. Analysis of the Plasmodium falciparum Proteome by High-Accuracy Mass Spectrometry. Nature 2002, 419, 537–542. [Google Scholar] [CrossRef]
- Scholz, S.M.; Simon, N.; Lavazec, C.; Dude, M.A.; Templeton, T.J.; Pradel, G. PfCCp Proteins of Plasmodium falciparum: Gametocyte-Specific Expression and Role in Complement-Mediated Inhibition of Exflagellation. Int. J. Parasitol. 2008, 38, 327–340. [Google Scholar] [CrossRef]
- Lasonder, E.; Rijpma, S.R.; van Schaijk, B.C.L.; Hoeijmakers, W.A.M.; Kensche, P.R.; Gresnigt, M.S.; Italiaander, A.; Vos, M.W.; Woestenenk, R.; Bousema, T.; et al. Integrated Transcriptomic and Proteomic Analyses of P. falciparum Gametocytes: Molecular Insight into Sex-Specific Processes and Translational Repression. Nucleic Acids Res. 2016, 44, 6087–6101. [Google Scholar] [CrossRef]
- Pradel, G.; Wagner, C.; Mejia, C.; Templeton, T.J. Plasmodium falciparum: Co-Dependent Expression and Co-Localization of the PfCCp Multi-Adhesion Domain Proteins. Exp. Parasitol. 2006, 112, 263–268. [Google Scholar] [CrossRef]
- Simon, N.; Scholz, S.M.; Moreira, C.K.; Templeton, T.J.; Kuehn, A.; Dude, M.-A.; Pradel, G. Sexual Stage Adhesion Proteins Form Multi-Protein Complexes in the Malaria Parasite Plasmodium falciparum. J. Biol. Chem. 2009, 284, 14537–14546. [Google Scholar] [CrossRef]
- Simon, N.; Kuehn, A.; Williamson, K.C.; Pradel, G. Adhesion Protein Complexes of Malaria Gametocytes Assemble Following Parasite Transmission to the Mosquito. Parasitol. Int. 2016, 65, 27–30. [Google Scholar] [CrossRef]
- Eksi, S.; Czesny, B.; van Gemert, G.-J.; Sauerwein, R.W.; Eling, W.; Williamson, K.C. Malaria Transmission-Blocking Antigen, Pfs230, Mediates Human Red Blood Cell Binding to Exflagellating Male Parasites and Oocyst Production. Mol. Microbiol. 2006, 61, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N. Target Antigens of Malaria Transmission Blocking Immunity Exist as a Stable Membrane Bound Complex. Parasite Immunol. 1987, 9, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Wizel, B. Further Characterization of Interactions between Gamete Surface Antigens of Plasmodium falciparum. Mol. Biochem. Parasitol. 1992, 53, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Tremp, A.Z.; Saeed, S.; Sharma, V.; Lasonder, E.; Dessens, J.T. Plasmodium berghei LAPs Form an Extended Protein Complex That Facilitates Crystalloid Targeting and Biogenesis. J. Proteom. 2020, 227, 103925. [Google Scholar] [CrossRef]
- Bennink, S.; Pradel, G. The Molecular Machinery of Translational Control in Malaria Parasites. Mol. Microbiol. 2019, 112, 1658–1673. [Google Scholar] [CrossRef]
- Brooks, S.R.; Williamson, K.C. Proteolysis of Plasmodium falciparum Surface Antigen, Pfs230, during Gametogenesis. Mol. Biochem. Parasitol. 2000, 106, 77–82. [Google Scholar] [CrossRef]
- von Bohl, A.; Kuehn, A.; Simon, N.; Ngongang, V.N.; Spehr, M.; Baumeister, S.; Przyborski, J.M.; Fischer, R.; Pradel, G. A WD40-Repeat Protein Unique to Malaria Parasites Associates with Adhesion Protein Complexes and Is Crucial for Blood Stage Progeny. Malar. J. 2015, 14, 435. [Google Scholar] [CrossRef]
- Roling, L.; Flammersfeld, A.; Pradel, G.; Bennink, S. The WD40-Protein Pf WLP1 Ensures Stability of the Pf CCp-Based Adhesion Protein Complex in Plasmodium falciparum Gametocytes. Front. Cell. Infect. Microbiol. 2022, 12, 942364. [Google Scholar] [CrossRef]
- Baum, J.; Richard, D.; Healer, J.; Rug, M.; Krnajski, Z.; Gilberger, T.-W.; Green, J.L.; Holder, A.A.; Cowman, A.F. A Conserved Molecular Motor Drives Cell Invasion and Gliding Motility across Malaria Life Cycle Stages and Other Apicomplexan Parasites. J. Biol. Chem. 2006, 281, 5197–5208. [Google Scholar] [CrossRef] [PubMed]
- Bargieri, D.Y.; Thiberge, S.; Tay, C.L.; Carey, A.F.; Rantz, A.; Hischen, F.; Lorthiois, A.; Straschil, U.; Singh, P.; Singh, S.; et al. Plasmodium Merozoite TRAP Family Protein Is Essential for Vacuole Membrane Disruption and Gamete Egress from Erythrocytes. Cell Host Microbe 2016, 20, 618–630. [Google Scholar] [CrossRef]
- Kehrer, J.; Frischknecht, F.; Mair, G.R. Proteomic Analysis of the Plasmodium berghei Gametocyte Egressome and Vesicular BioID of Osmiophilic Body Proteins Identifies Merozoite TRAP-like Protein (MTRAP) as an Essential Factor for Parasite Transmission. Mol. Cell. Proteom. 2016, 15, 2852–2862. [Google Scholar] [CrossRef]
- Flieger, A.; Frischknecht, F.; Häcker, G.; Hornef, M.W.; Pradel, G. Pathways of Host Cell Exit by Intracellular Pathogens. Microb. Cell 2018, 5, 525–544. [Google Scholar] [CrossRef] [PubMed]
- Lavazec, C.; Moreira, C.K.; Mair, G.R.; Waters, A.P.; Janse, C.J.; Templeton, T.J. Analysis of Mutant Plasmodium berghei Parasites Lacking Expression of Multiple PbCCp Genes. Mol. Biochem. Parasitol. 2009, 163, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.; Carter, V.; Tremp, A.Z.; Dessens, J.T. Translational Repression Controls Temporal Expression of the Plasmodium berghei LCCL Protein Complex. Mol. Biochem. Parasitol. 2013, 189, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.M.; Franke-Fayard, B.; Mair, G.R.; Lasonder, E.; Janse, C.J.; Mann, M.; Waters, A.P. Proteome Analysis of Separated Male and Female Gametocytes Reveals Novel Sex-Specific Plasmodium Biology. Cell 2005, 121, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Raine, J.D.; Ecker, A.; Mendoza, J.; Tewari, R.; Stanway, R.R.; Sinden, R.E. Female Inheritance of Malarial Lap Genes Is Essential for Mosquito Transmission. PLoS Pathog. 2007, 3, e30. [Google Scholar] [CrossRef]
- Carter, V.; Shimizu, S.; Arai, M.; Dessens, J.T. PbSR Is Synthesized in Macrogametocytes and Involved in Formation of the Malaria Crystalloids. Mol. Microbiol. 2008, 68, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.; Carter, V.; Tremp, A.Z.; Dessens, J.T. Plasmodium berghei Crystalloids Contain Multiple LCCL Proteins. Mol. Biochem. Parasitol. 2010, 170, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.; Tremp, A.Z.; Dessens, J.T. Conformational Co-Dependence between Plasmodium berghei LCCL Proteins Promotes Complex Formation and Stability. Mol. Biochem. Parasitol. 2012, 185, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.; Tremp, A.Z.; Dessens, J.T. Biogenesis of the Crystalloid Organelle in Plasmodium Involves Microtubule-Dependent Vesicle Transport and Assembly. Int. J. Parasitol. 2015, 45, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Dessens, J.T.; Saeed, S.; Tremp, A.Z.; Carter, V. Malaria Crystalloids: Specialized Structures for Parasite Transmission? Trends Parasitol. 2011, 27, 106–110. [Google Scholar] [CrossRef]
- Dessens, J.T.; Tremp, A.Z.; Saeed, S. Crystalloids: Fascinating Parasite Organelles Essential for Malaria Transmission. Trends Parasitol. 2021, 37, 581–584. [Google Scholar] [CrossRef]
- Saeed, S.; Tremp, A.Z.; Dessens, J.T. The Plasmodium LAP Complex Affects Crystalloid Biogenesis and Oocyst Cell Division. Int. J. Parasitol. 2018, 48, 1073–1078. [Google Scholar] [CrossRef]
- Guerreiro, A.; Deligianni, E.; Santos, J.M.; Silva, P.A.G.C.; Louis, C.; Pain, A.; Janse, C.J.; Franke-Fayard, B.; Carret, C.K.; Siden-Kiamos, I.; et al. Genome-Wide RIP-Chip Analysis of Translational Repressor-Bound MRNAs in the Plasmodium Gametocyte. Genome Biol. 2014, 15, 493. [Google Scholar] [CrossRef] [PubMed]
- Ukegbu, C.V.; Gomes, A.R.; Giorgalli, M.; Campos, M.; Bailey, A.J.; Besson, T.R.B.; Billker, O.; Vlachou, D.; Christophides, G.K. Identification of Genes Required for Plasmodium Gametocyte-to-Sporozoite Development in the Mosquito Vector. Cell Host Microbe 2023, 31, 1539–1551.e6. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.M.; Duarte, N.; Kehrer, J.; Ramesar, J.; Avramut, M.C.; Koster, A.J.; Dessens, J.T.; Frischknecht, F.; Chevalley-Maurel, S.; Janse, C.J.; et al. Maternally Supplied S-Acyl-Transferase Is Required for Crystalloid Organelle Formation and Transmission of the Malaria Parasite. Proc. Natl. Acad. Sci. USA 2016, 113, 7183–7188. [Google Scholar] [CrossRef] [PubMed]
- Ecker, A.; Bushell, E.S.C.; Tewari, R.; Sinden, R.E. Reverse Genetics Screen Identifies Six Proteins Important for Malaria Development in the Mosquito. Mol. Microbiol. 2008, 70, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.; Lau, C.I.; Tremp, A.Z.; Crompton, T.; Dessens, J.T. Dysregulated Gene Expression in Oocysts of Plasmodium berghei LAP Mutants. Mol. Biochem. Parasitol. 2019, 229, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Yuda, M.; Iwanaga, S.; Shigenobu, S.; Kato, T.; Kaneko, I. Transcription Factor AP2-Sp and Its Target Genes in Malarial Sporozoites. Mol. Microbiol. 2010, 75, 854–863. [Google Scholar] [CrossRef]
- Modrzynska, K.; Pfander, C.; Chappell, L.; Yu, L.; Suarez, C.; Dundas, K.; Gomes, A.R.; Goulding, D.; Rayner, J.C.; Choudhary, J.; et al. A Knockout Screen of ApiAP2 Genes Reveals Networks of Interacting Transcriptional Regulators Controlling the Plasmodium Life Cycle. Cell Host Microbe 2017, 21, 11–22. [Google Scholar] [CrossRef]
- Ménard, R.; Sultan, A.A.; Cortes, C.; Altszuler, R.; Van Dijk, M.R.; Janse, C.J.; Waters, A.P.; Nussenzweig, R.S.; Nussenzweig, V. Circumsporozoite Protein Is Required for Development of Malaria Sporozoites in Mosquitoes. Nature 1997, 385, 336–340. [Google Scholar] [CrossRef]
- Khater, E.I.; Sinden, R.E.; Dessens, J.T. A Malaria Membrane Skeletal Protein Is Essential for Normal Morphogenesis, Motility, and Infectivity of Sporozoites. J. Cell Biol. 2004, 167, 425–432. [Google Scholar] [CrossRef]
- Al-Khattaf, F.S.; Tremp, A.Z.; El-Houderi, A.; Dessens, J.T. The Plasmodium Alveolin IMC1a Is Stabilised by Its Terminal Cysteine Motifs and Facilitates Sporozoite Morphogenesis and Infectivity in a Dose-Dependent Manner. Mol. Biochem. Parasitol. 2017, 211, 48–56. [Google Scholar] [CrossRef]
- Spaccapelo, R.; Naitza, S.; Robson, K.J.; Crisanti, A. Thrombospondin-Related Adhesive Protein (TRAP) of Plasmodium berghei and Parasite Motility. Lancet 1997, 350, 335. [Google Scholar] [CrossRef] [PubMed]
- Sultan, A.A.; Thathy, V.; Frevert, U.; Robson, K.J.H.; Crisanti, A.; Nussenzweig, V.; Nussenzweig, R.S.; Ménard, R. TRAP Is Necessary for Gliding Motility and Infectivity of Plasmodium Sporozoites. Cell 1997, 90, 511–522. [Google Scholar] [CrossRef]
- Ishino, T.; Chinzei, Y.; Yuda, M. A Plasmodium Sporozoite Protein with a Membrane Attack Complex Domain Is Required for Breaching the Liver Sinusoidal Cell Layer Prior to Hepatocyte Infection. Cell. Microbiol. 2005, 7, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.; Tremp, A.Z.; Sharma, V.; Lasonder, E.; Dessens, J.T. NAD(P) Transhydrogenase Has Vital Non-Mitochondrial Functions in Malaria Parasite Transmission. EMBO Rep. 2020, 21, e47832. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bennink, S.; Pradel, G. The Multiple Roles of LCCL Domain-Containing Proteins for Malaria Parasite Transmission. Microorganisms 2024, 12, 279. https://doi.org/10.3390/microorganisms12020279
Bennink S, Pradel G. The Multiple Roles of LCCL Domain-Containing Proteins for Malaria Parasite Transmission. Microorganisms. 2024; 12(2):279. https://doi.org/10.3390/microorganisms12020279
Chicago/Turabian StyleBennink, Sandra, and Gabriele Pradel. 2024. "The Multiple Roles of LCCL Domain-Containing Proteins for Malaria Parasite Transmission" Microorganisms 12, no. 2: 279. https://doi.org/10.3390/microorganisms12020279
APA StyleBennink, S., & Pradel, G. (2024). The Multiple Roles of LCCL Domain-Containing Proteins for Malaria Parasite Transmission. Microorganisms, 12(2), 279. https://doi.org/10.3390/microorganisms12020279