
Planted Citrus Regulates the Community and Networks of phoD-Harboring Bacteria to Drive Phosphorus Availability Between Karst and Non-Karst Soils

 ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sampling
2.3. General Soil Parameters
2.4. DNA Extraction and Illumina Sequencing
2.5. Statistical Analyses
3. Results
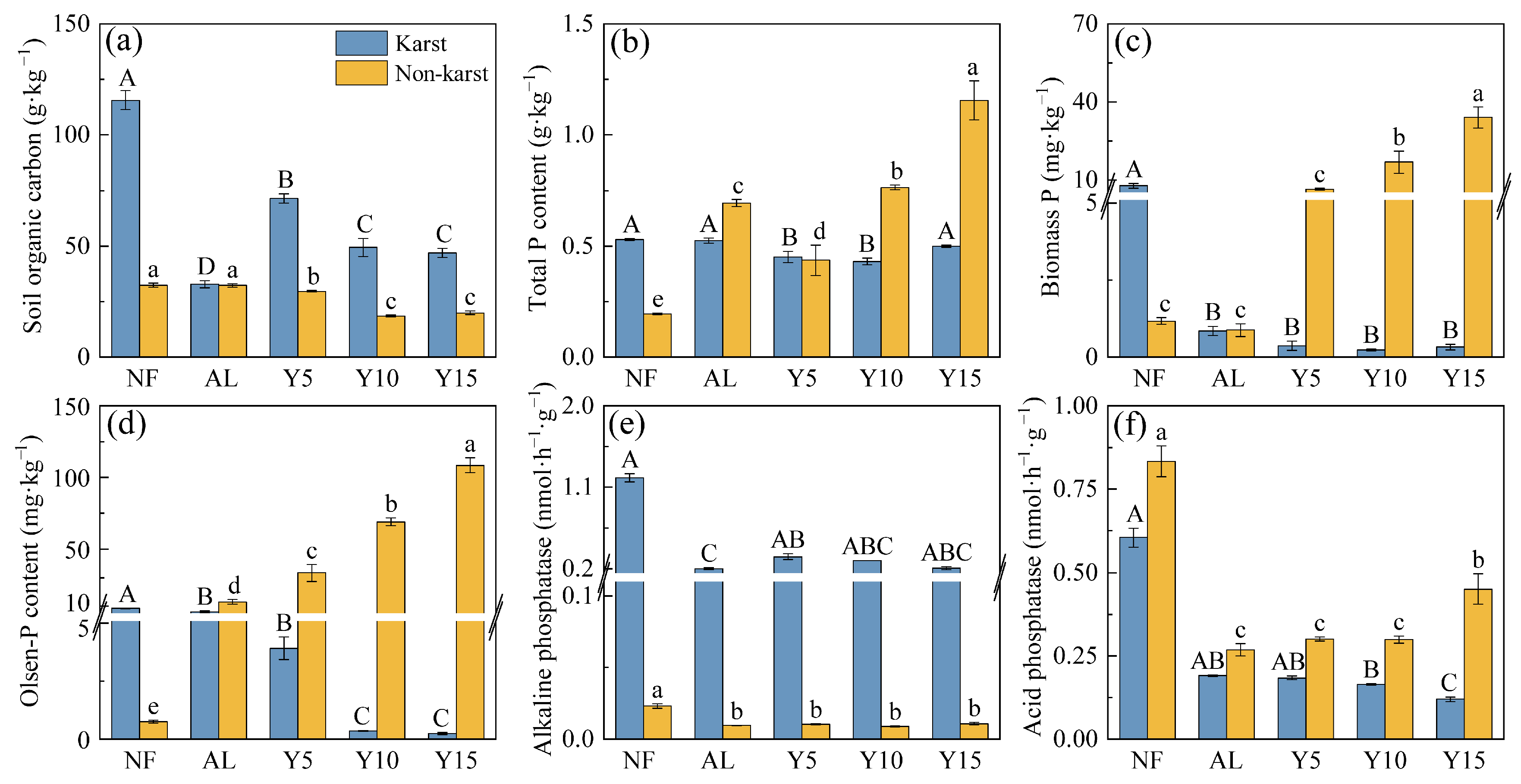
3.1. Soil P Contents and Enzyme Activities in Karst and Non-Karst Citrus Cultivations
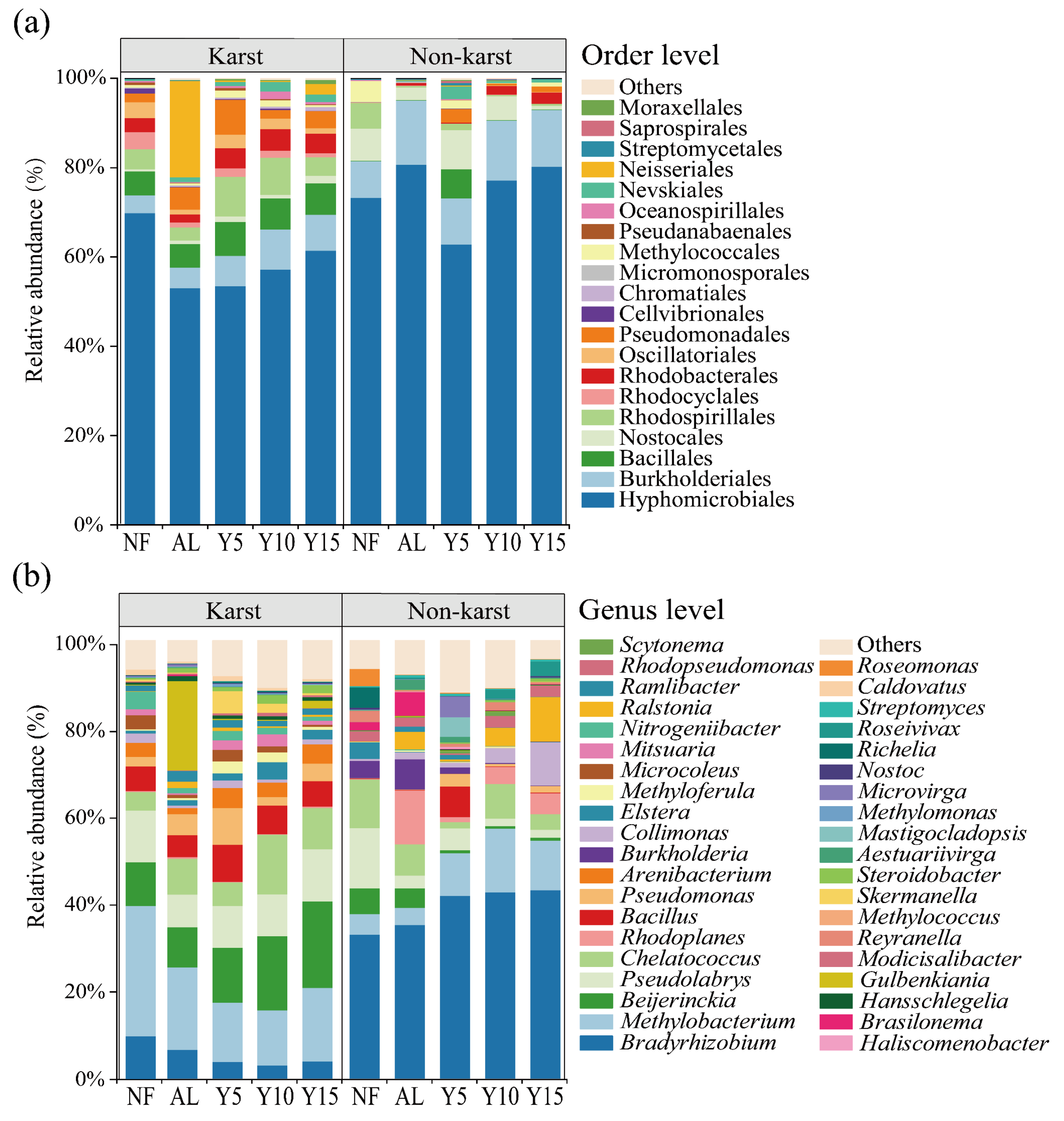
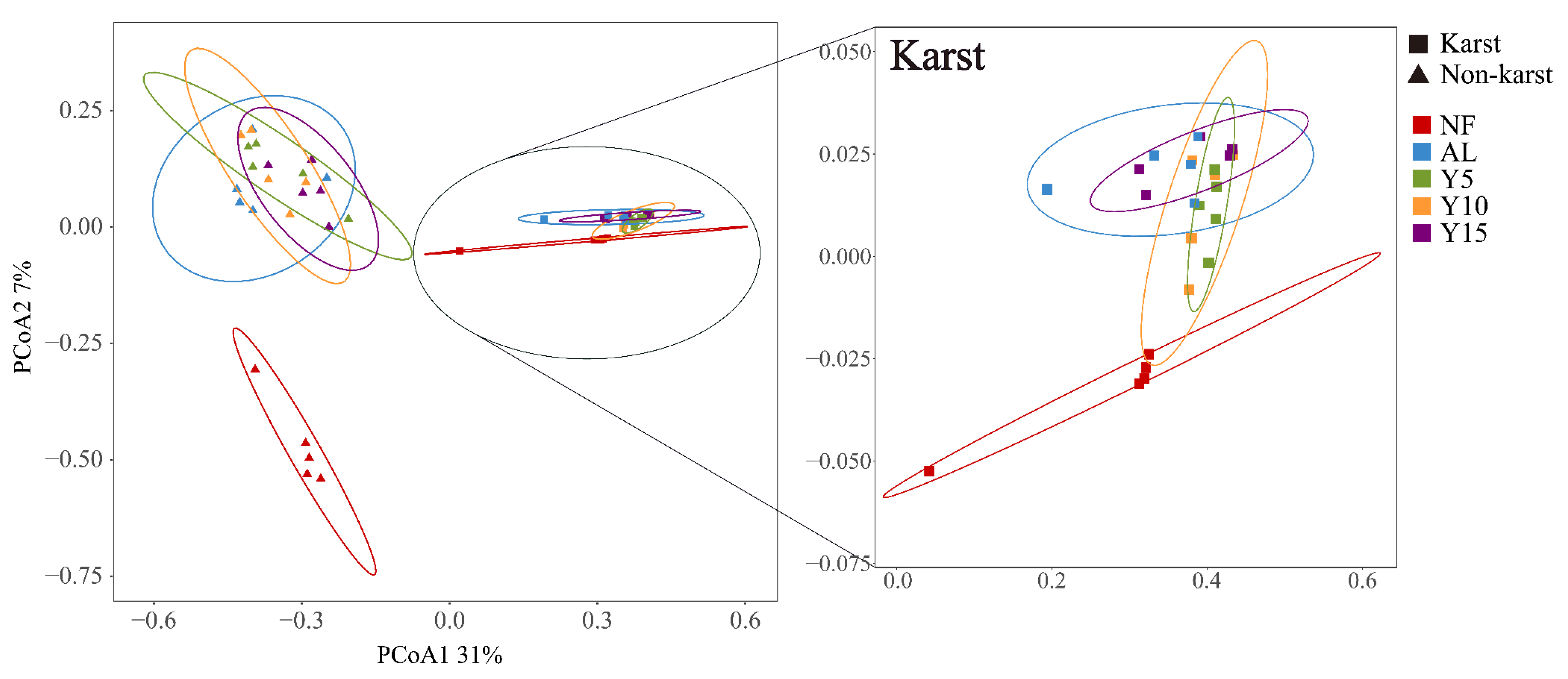
3.2. Community Structure, Diversity, and Co-Occurrence Networks of phoD-Harboring Bacteria
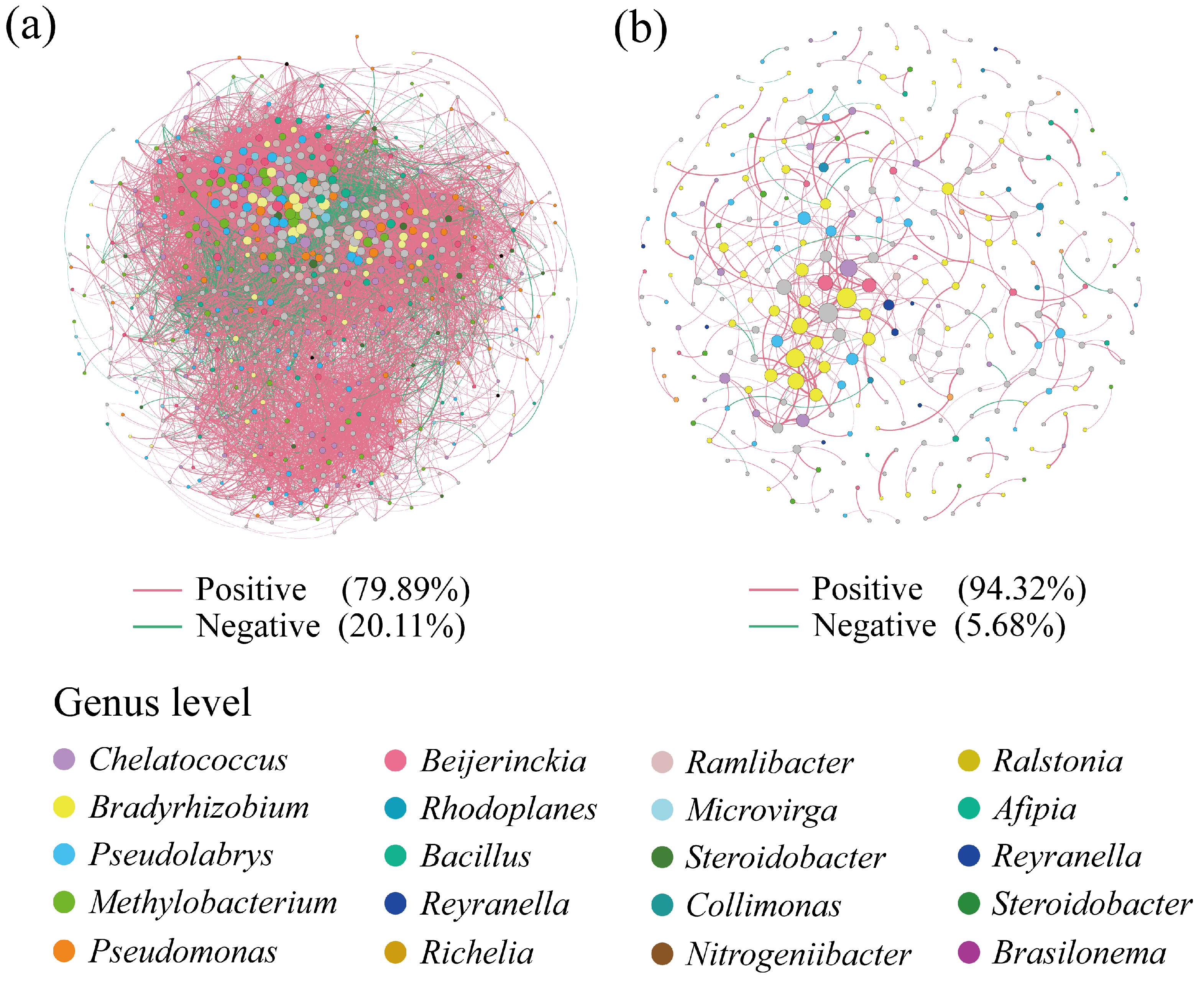
3.3. Co-Occurrence Networks of phoD-Harboring Bacteria
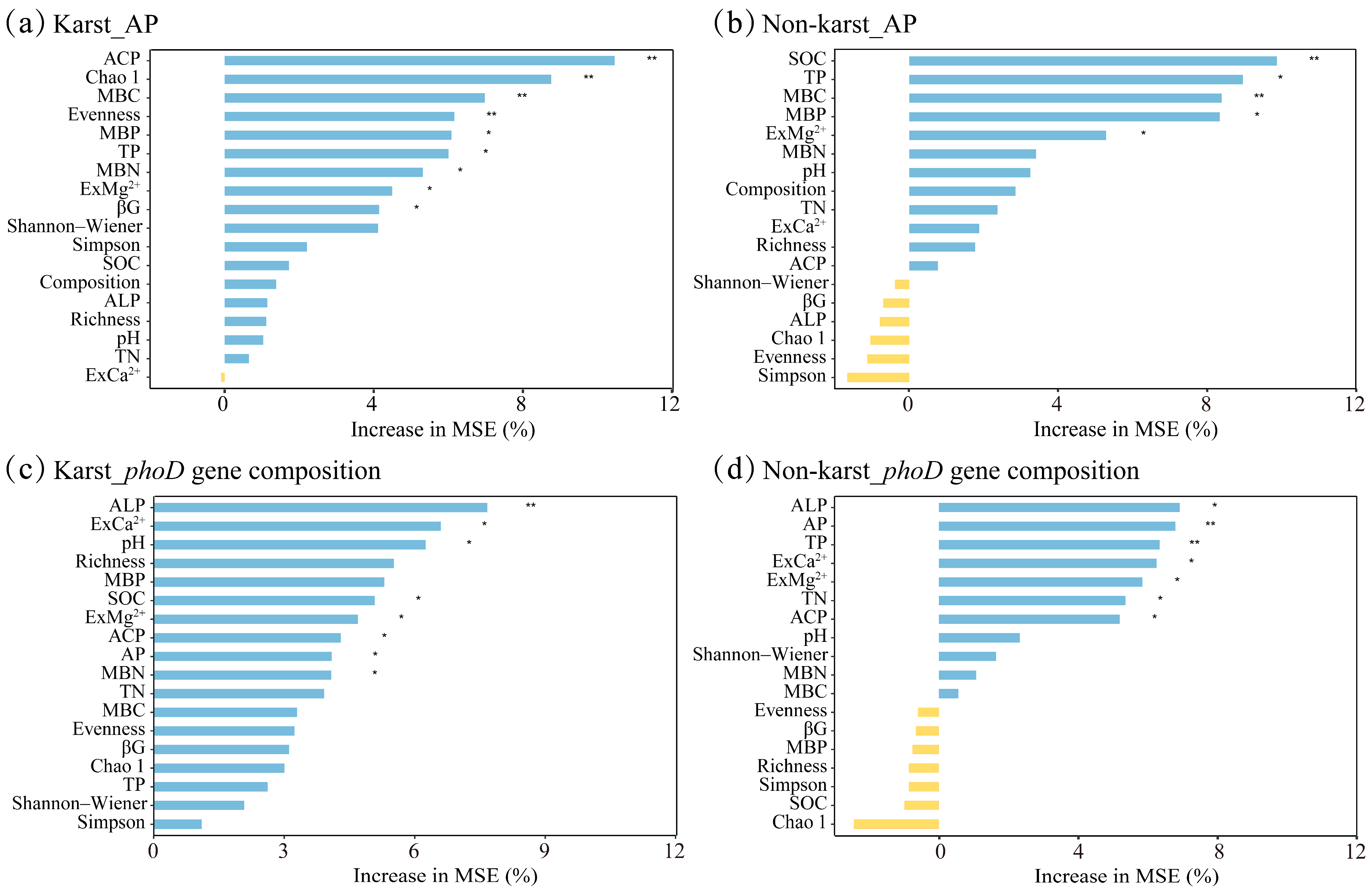
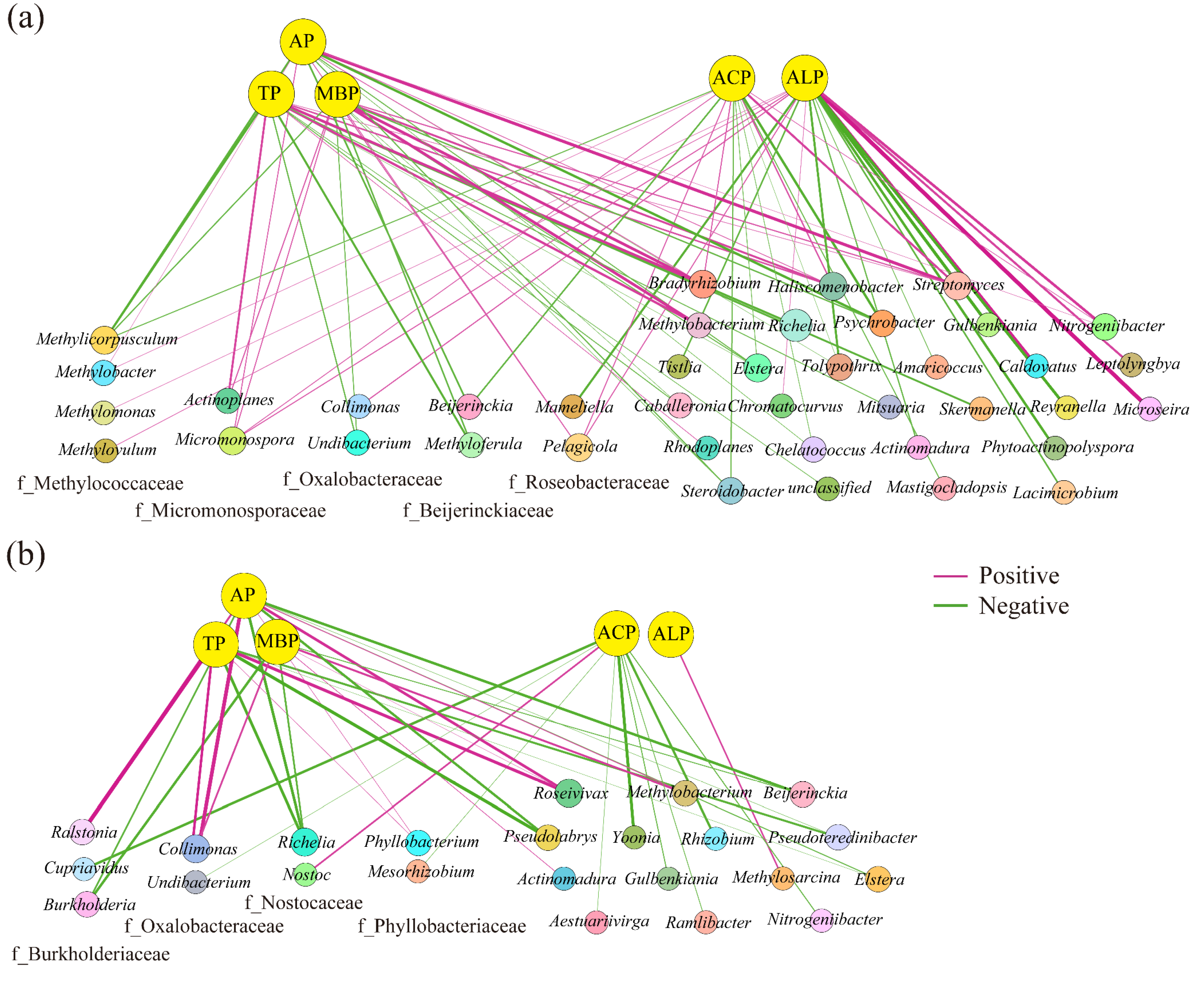
3.4. Soil phoD-Harboring Bacteria Affected P Availability
4. Discussion
4.1. Soil P Availability and Influencing Factors
4.2. phoD-Harboring Microbial Community and Co-Occurrence Networks and Influencing Factors
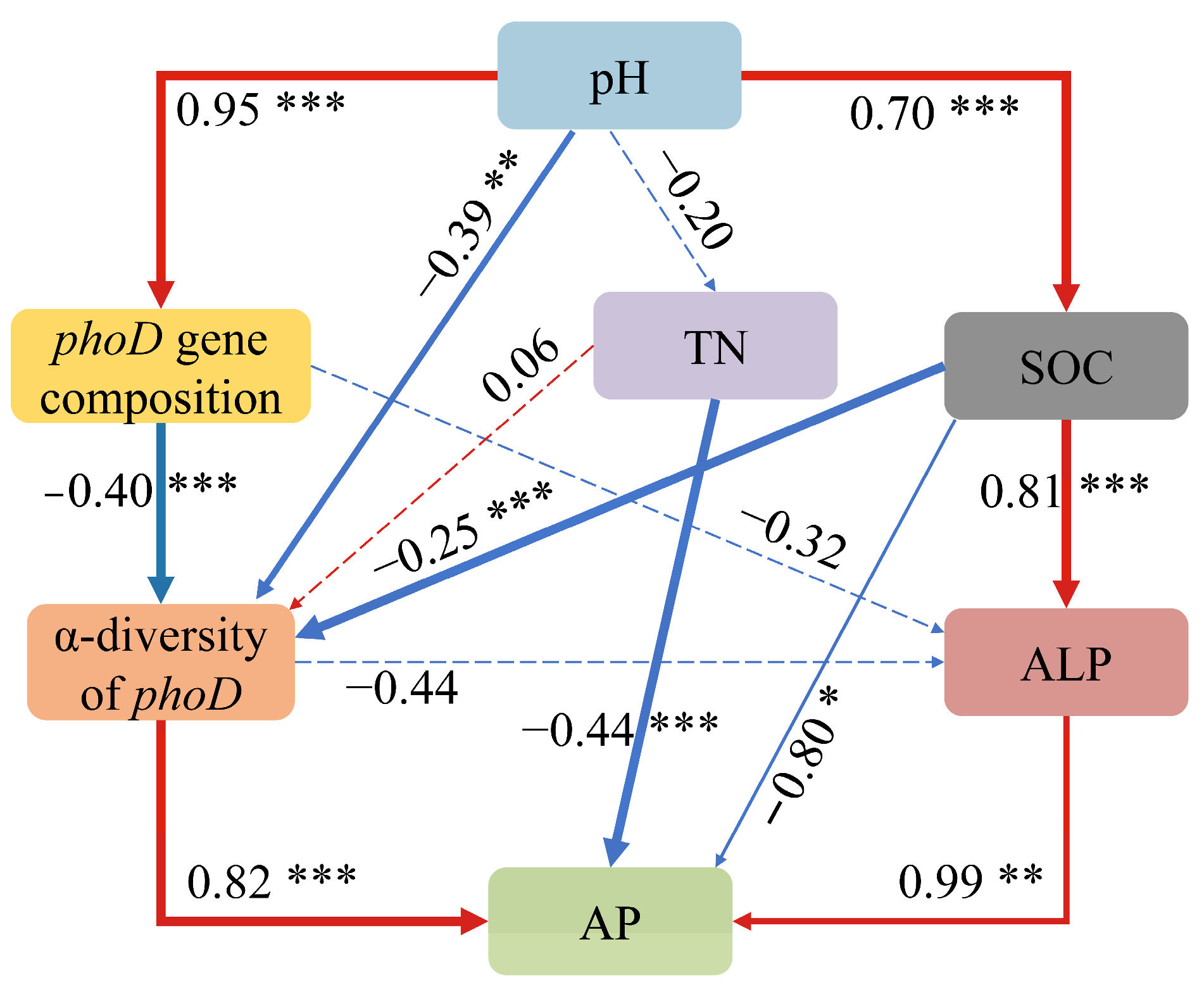
4.3. Mechanism by Which phoD-Harboring Bacteria Influenced P Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, F.; Qu, L.; Hong, X.; Sun, X. Isolation and Characterization of a Phosphate-Solubilizing Halophilic Bacterium Kushneria sp. YCWA18 from Daqiao Saltern on the Coast of Yellow Sea of China. Evid. Based Complement. Altern. Med. 2011, 2011, 615032. [Google Scholar] [CrossRef]
- Ma, X.; Li, F.; Chen, Y.; Chang, Y.; Lian, X.; Li, Y.; Ye, L.; Yin, T.; Lu, X. Effects of Fertilization Approaches on Plant Development and Fertilizer Use of Citrus. Plants 2022, 11, 2547. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-J.; Yang, M.; Zhang, Z.-Z.; Li, W.-L.; Guo, C.-Y.; Chen, X.-P.; Shi, X.-J.; Zhou, P.; Tang, X.-D.; Zhang, Y.-Q.; et al. An Ecological Research on Potential for Zero-growth of Chemical Fertilizer Use in Citrus Production in China. Ekoloji 2019, 28, 1049–1059. [Google Scholar]
- White, P.J.; Brown, P.H. Plant nutrition for sustainable development and global health. Ann. Bot. 2010, 105, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Zancanaro, L.E.; Nunes, R.; Sousa, D.; Busato, J.; Figueiredo, C. Response of Maize to Different Soil Residual Phosphorus Conditions. Agron. J. 2019, 111, 3291–3300. [Google Scholar] [CrossRef]
- Carpenter, S. Eutrophication of Aquatic Ecosystems: Bistability and Soil Phosphorus. Proc. Natl. Acad. Sci. USA 2005, 102, 10002–10005. [Google Scholar] [CrossRef]
- Chen, X.; Condron, L.M.; Dunfield, K.E.; Wakelin, S.A.; Chen, L. Impact of grassland afforestation with contrasting tree species on soil phosphorus fractions and alkaline phosphatase gene communities. Soil Biol. Biochem. 2021, 159, 108274. [Google Scholar] [CrossRef]
- Zeng, Q.; Mei, T.; Delgado-Baquerizo, M.; Wang, M.; Tan, W. Suppressed phosphorus-mineralizing bacteria after three decades of fertilization. Agric. Ecosyst. Environ. 2022, 323, 107679. [Google Scholar] [CrossRef]
- Bibi, S.; Irshad, M.; Ullah, F.; Mahmood, Q.; Shahzad, M.; Tariq, M.A.U.R.; Hussain, Z.; Mohiuddin, M.; An, P.; Ng, A.W.M.; et al. Phosphorus extractability in relation to soil properties in different fields of fruit orchards under similar ecological conditions of Pakistan. Front. Ecol. Evol. 2023, 10, 1077270. [Google Scholar] [CrossRef]
- MacDonald, G.K.; Bennett, E.M.; Taranu, Z.E. The influence of time, soil characteristics, and land-use history on soil phosphorus legacies: A global meta-analysis. Glob. Change Biol. 2012, 18, 1904–1917. [Google Scholar] [CrossRef]
- Jiang, Y.; Yang, X.; Ni, K.; Ma, L.; Shi, Y.; Wang, Y.; Cai, Y.; Ma, Q.; Ruan, J. Nitrogen addition reduces phosphorus availability and induces a shift in soil phosphorus cycling microbial community in a tea (Camellia sinensis L.) plantation. J. Environ. Manag. 2023, 342, 118207. [Google Scholar] [CrossRef] [PubMed]
- Fraser, T.D.; Lynch, D.H.; Bent, E.; Entz, M.H.; Dunfield, K.E. Soil bacterial phoD gene abundance and expression in response to applied phosphorus and long-term management. Soil Biol. Biochem. 2015, 88, 137–147. [Google Scholar] [CrossRef]
- Tan, H.; Barret, M.; Mooij, M.J.; Rice, O.; Morrissey, J.P.; Dobson, A.; Griffiths, B.; O’Gara, F. Long-term phosphorus fertilisation increased the diversity of the total bacterial community and the phoD phosphorus mineraliser group in pasture soils. Biol. Fertil. Soils 2013, 49, 661–672. [Google Scholar] [CrossRef]
- Daneshgar, S.; Callegari, A.; Capodaglio, A.G.; Vaccari, D. The Potential Phosphorus Crisis: Resource Conservation and Possible Escape Technologies: A Review. Resources 2018, 7, 37. [Google Scholar] [CrossRef]
- Lynch, M.D.J.; Neufeld, J.D. Ecology and exploration of the rare biosphere. Nat. Rev. Microbiol. 2015, 13, 217–229. [Google Scholar] [CrossRef]
- Wei, X.; Hu, Y.; Razavi, B.S.; Zhou, J.; Shen, J.; Nannipieri, P.; Wu, J.; Ge, T. Rare taxa of alkaline phosphomonoesterase-harboring microorganisms mediate soil phosphorus mineralization. Soil Biol. Biochem. 2019, 131, 62–70. [Google Scholar] [CrossRef]
- Tong, X.; Brandt, M.; Yue, Y.; Horion, S.; Wang, K.; Keersmaecker, W.D.; Tian, F.; Schurgers, G.; Xiao, X.; Luo, Y.; et al. Increased vegetation growth and carbon stock in China karst via ecological engineering. Nat. Sustain. 2018, 1, 44–50. [Google Scholar] [CrossRef]
- Wang, Y.-P.; Huang, Y.; Augusto, L.; Goll, D.S.; Helfenstein, J.; Hou, E. Toward a Global Model for Soil Inorganic Phosphorus Dynamics: Dependence of Exchange Kinetics and Soil Bioavailability on Soil Physicochemical Properties. Glob. Biogeochem. Cycles 2022, 36, e2021GB007061. [Google Scholar] [CrossRef]
- Green, S.M.; Dungait, J.A.J.; Tu, C.; Buss, H.L.; Sanderson, N.; Hawkes, S.J.; Xing, K.; Yue, F.; Hussey, V.L.; Peng, J.; et al. Soil functions and ecosystem services research in the Chinese karst Critical Zone. Chem. Geol. 2019, 527, 119107. [Google Scholar] [CrossRef]
- Zhou, M.; Yang, H.; Zhu, T.; Zhang, C.; Zhu, D. Preliminary Research on Agricultural Cultivation Decreasing Amino Sugar Accumulation in Calcareous Soils in Subtropical Karst Region of China. Land 2022, 11, 1684. [Google Scholar] [CrossRef]
- Sun, M.; Yang, R.; Tang, Y.; Xiao, D.; Zhang, W.; Xu, Z.; Shi, Z.; Hu, P.; Wu, H.; Wang, K. Lithologic control of soil C:N:P stoichiometry across a climatic gradient in southwest China. J. Soil Sediment. 2023, 23, 1662–1673. [Google Scholar] [CrossRef]
- Pan, F.; Yu, X.; Chen, M.; Liang, Y. Vegetation recovery reshapes the composition and enhances the network connectivity of phoD-harboring microorganisms to promote P availability in a karst ecosystem. Sci. Total Environ. 2024, 918, 170561. [Google Scholar] [CrossRef] [PubMed]
- Finzi, A.C.; Sinsabaugh, R.L.; Long, T.M.; Osgood, M.P. Microbial Community Responses to Atmospheric Carbon Dioxide Enrichment in a Warm-Temperate Forest. Ecosystems 2006, 9, 215–226. [Google Scholar] [CrossRef]
- Sakurai, M.; Wasaki, J.; Tomizawa, Y.; Shinano, T.; Osaki, M. Analysis of bacterial communities on alkaline phosphatase genes in soil supplied with organic matter. Soil Sci. Plant Nutr. 2008, 54, 62–71. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Fish, J.A.; Chai, B.; Wang, Q.; Sun, Y.; Brown, C.T.; Tiedje, J.M.; Cole, J.R. FunGene: The functional gene pipeline and repository. Front. Microbiol. 2013, 4, 291. [Google Scholar] [CrossRef]
- Faust, K. Open challenges for microbial network construction and analysis. ISME J. 2021, 15, 3111–3118. [Google Scholar] [CrossRef]
- Berry, D.; Widder, S. Deciphering microbial interactions and detecting keystone species with co-occurrence networks. Front. Microbiol. 2014, 5, 219. [Google Scholar] [CrossRef]
- Sun, Y.; Li, X.; Cao, N.; Duan, C.; Ding, C.; Huang, Y.; Wang, J. Biodegradable microplastics enhance soil microbial network complexity and ecological stochasticity. J. Hazard. Mater. 2022, 439, 129610. [Google Scholar] [CrossRef]
- Guimerà, R.; Nunes Amaral, L.A. Functional cartography of complex metabolic networks. Nature 2005, 433, 895–900. [Google Scholar] [CrossRef]
- Xiao, X.; Fan, M.; Wang, E.; Chen, W.; Wei, G. Interactions of plant growth-promoting rhizobacteria and soil factors in two leguminous plants. Appl. Microbiol. Biotechnol. 2017, 101, 8485–8497. [Google Scholar] [CrossRef] [PubMed]
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: A review. Plant Soil 2001, 237, 173–195. [Google Scholar] [CrossRef]
- Yang, R.; Li, J.; Long, J.; Liao, H.; Wang, X.; Li, Y. Structural characteristics of bacterial community in rhizosphere soil of Zanthoxylum bungeamun in different planting years in Karst Areas of Guizhou. Ecol. Environ. Sci. 2021, 30, 81–91. (In Chinese) [Google Scholar] [CrossRef]
- Liao, L.; Shi, F.; Zhang, N.; Chen, X.; Bu, H.; Sun, F. Effects of Different Planting Years on Rhizosphere Soil Physiochemical Properties and Microbial Community of Zanthoxylum bungeanum. Bull. Bot. Res. 2022, 42, 466–474. (In Chinese) [Google Scholar] [CrossRef]
- Peng, S.; Kuang, X.; Cheng, H.; Wei, K.; Cai, K.; Tian, J. Post-agricultural succession affects the accumulation and enzymatic transformation of organic phosphorus in a karst area, southwest China. Plant Soil 2024, 498, 5–20. [Google Scholar] [CrossRef]
- Kumar, M.; Kour, D.; Yadav, A.N.; Saxena, R.; Rai, P.K.; Jyoti, A.; Tomar, R.S. Biodiversity of methylotrophic microbial communities and their potential role in mitigation of abiotic stresses in plants. Biologia 2019, 74, 287–308. [Google Scholar] [CrossRef]
- Wielbo, J.; Kidaj, D.; Koper, P.; Kubik-Komar, A.; Skorupska, A. The effect of biotic and physical factors on the competitive ability of Rhizobium leguminosarum. Cent. Eur. J. Biol. 2012, 7, 13–24. [Google Scholar] [CrossRef]
- Luo, G.; Ling, N.; Nannipieri, P.; Chen, H.; Raza, W.; Wang, M.; Guo, S.; Shen, Q. Long-term fertilisation regimes affect the composition of the alkaline phosphomonoesterase encoding microbial community of a vertisol and its derivative soil fractions. Biol. Fertil. Soils 2017, 53, 375–388. [Google Scholar] [CrossRef]
- Zhu, X.; Zhao, X.; Lin, Q.; Li, G. Distribution Characteristics of phoD-Harbouring Bacterial Community Structure and Its Roles in Phosphorus Transformation in Steppe Soils in Northern China. J. Soil Sci. Plant Nutr. 2021, 21, 1531–1541. [Google Scholar] [CrossRef]
- Fan, Z.; Lu, S.; Liu, S.; Guo, H.; Wang, T.; Zhou, J.; Peng, X. Changes in Plant Rhizosphere Microbial Communities under Different Vegetation Restoration Patterns in Karst and Non-karst Ecosystems. Sci. Rep. 2019, 9, 8761. [Google Scholar] [CrossRef]
- Barber, J.N.; Nicholson, L.C.; Woods, L.C.; Judd, L.M.; Sezmis, A.L.; Hawkey, J.; Holt, K.E.; McDonald, M.J. Species interactions constrain adaptation and preserve ecological stability in an experimental microbial community. ISME J. 2022, 16, 1442–1452. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, J.; Yuan, J.; Tang, Z.; Wang, J.; Zhang, Y. Long-Term Organic Fertilization Strengthens the Soil Phosphorus Cycle and Phosphorus Availability by Regulating the pqqC- and phoD-Harboring Bacterial Communities. Microb. Ecol. 2023, 86, 2716–2732. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; He, X.; Zhang, W.; Cheng, M.; Hu, P.; Wang, K. Diazotroph and arbuscular mycorrhizal fungal diversity and community composition responses to karst and non-karst soils. Appl. Soil Ecol. 2022, 170, 104227. [Google Scholar] [CrossRef]
- Wagg, C.; Schlaeppi, K.; Banerjee, S.; Kuramae, E.E.; van der Heijden, M.G.A. Fungal-bacterial diversity and microbiome complexity predict ecosystem functioning. Nat. Commun. 2019, 10, 4841. [Google Scholar] [CrossRef]
- Xiao, D.; He, X.; Zhang, W.; Hu, P.; Sun, M.; Wang, K. Comparison of bacterial and fungal diversity and network connectivity in karst and non-karst forests in southwest China. Sci. Total Environ. 2022, 822, 153179. [Google Scholar] [CrossRef]
- Xu, L.; Cao, H.; Li, C.; Wang, C.; He, N.; Hu, S.; Yao, M.; Wang, C.; Wang, J.; Zhou, S.; et al. The importance of rare versus abundant phoD-harboring subcommunities in driving soil alkaline phosphatase activity and available P content in Chinese steppe ecosystems. Soil Biol. Biochem. 2022, 164, 108491. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and interactions in plant growth promotion. Appl. Microbiol. Biotechnol. 2019, 103, 1179–1188. [Google Scholar] [CrossRef]
- Xun, W.; Liu, Y.; Li, W.; Ren, Y.; Xiong, W.; Xu, Z.; Zhang, N.; Miao, Y.; Shen, Q.; Zhang, R. Specialized metabolic functions of keystone taxa sustain soil microbiome stability. Microbiome 2021, 9, 35. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, J.-J.; Banerjee, S.; Zhou, N.; Zhao, Z.-Y.; Zhang, K.; Tian, C.-Y. Soil pH is equally important as salinity in shaping bacterial communities in saline soils under halophytic vegetation. Sci. Rep. 2018, 8, 4550. [Google Scholar] [CrossRef]
- Li, D.; Chen, L.; Xu, J.; Ma, L.; Olk, D.C.; Zhao, B.; Zhang, J.; Xin, X. Chemical nature of soil organic carbon under different long-term fertilization regimes is coupled with changes in the bacterial community composition in a Calcaric Fluvisol. Biol. Fertil. Soils 2018, 54, 999–1012. [Google Scholar] [CrossRef]
- Pan, F.J.; Yang, Q.; Liang, Y.M.; Yu, X.; Hu, P.L.; Zhang, W.; Pang, Y.L. Lithology and elevated temperature impact phoD-harboring bacteria on soil available P enhancing in subtropical forests. Sci. Total Environ. 2024, 948, 174815. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Feng, L.; Huang, Y.; Liang, Y.; Pan, F.; Zhang, W.; Zhao, Y.; Xiao, Y. Planted Citrus Regulates the Community and Networks of phoD-Harboring Bacteria to Drive Phosphorus Availability Between Karst and Non-Karst Soils. Microorganisms 2024, 12, 2582. https://doi.org/10.3390/microorganisms12122582
Yu X, Feng L, Huang Y, Liang Y, Pan F, Zhang W, Zhao Y, Xiao Y. Planted Citrus Regulates the Community and Networks of phoD-Harboring Bacteria to Drive Phosphorus Availability Between Karst and Non-Karst Soils. Microorganisms. 2024; 12(12):2582. https://doi.org/10.3390/microorganisms12122582
Chicago/Turabian StyleYu, Xuan, Lulu Feng, Yuan Huang, Yueming Liang, Fujing Pan, Wei Zhang, Yuan Zhao, and Yuexin Xiao. 2024. "Planted Citrus Regulates the Community and Networks of phoD-Harboring Bacteria to Drive Phosphorus Availability Between Karst and Non-Karst Soils" Microorganisms 12, no. 12: 2582. https://doi.org/10.3390/microorganisms12122582
APA StyleYu, X., Feng, L., Huang, Y., Liang, Y., Pan, F., Zhang, W., Zhao, Y., & Xiao, Y. (2024). Planted Citrus Regulates the Community and Networks of phoD-Harboring Bacteria to Drive Phosphorus Availability Between Karst and Non-Karst Soils. Microorganisms, 12(12), 2582. https://doi.org/10.3390/microorganisms12122582