
Genome Analysis and Potential Ecological Functions of Members of the Genus Ensifer from Subsurface Environments and Description of Ensifer oleiphilus sp. nov.
 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sources of Isolation of Bacterial Strains
2.2. DNA Extraction, 16S rRNA Gene Sequencing and Phylogenetic Analysis
2.3. Genome Sequencing and Analyses
2.4. Phenotypic Characterization
2.5. Chemotaxonomic Characterization
2.6. Analytical Methods
2.7. Bacterial Reduction of Radionuclides and Metals
2.8. Nucleotide Sequence Accession Numbers
3. Results and Discussion
3.1. Morphological Characterization of the Isolated Strains
3.2. Analysis of the 16S rRNA Genes
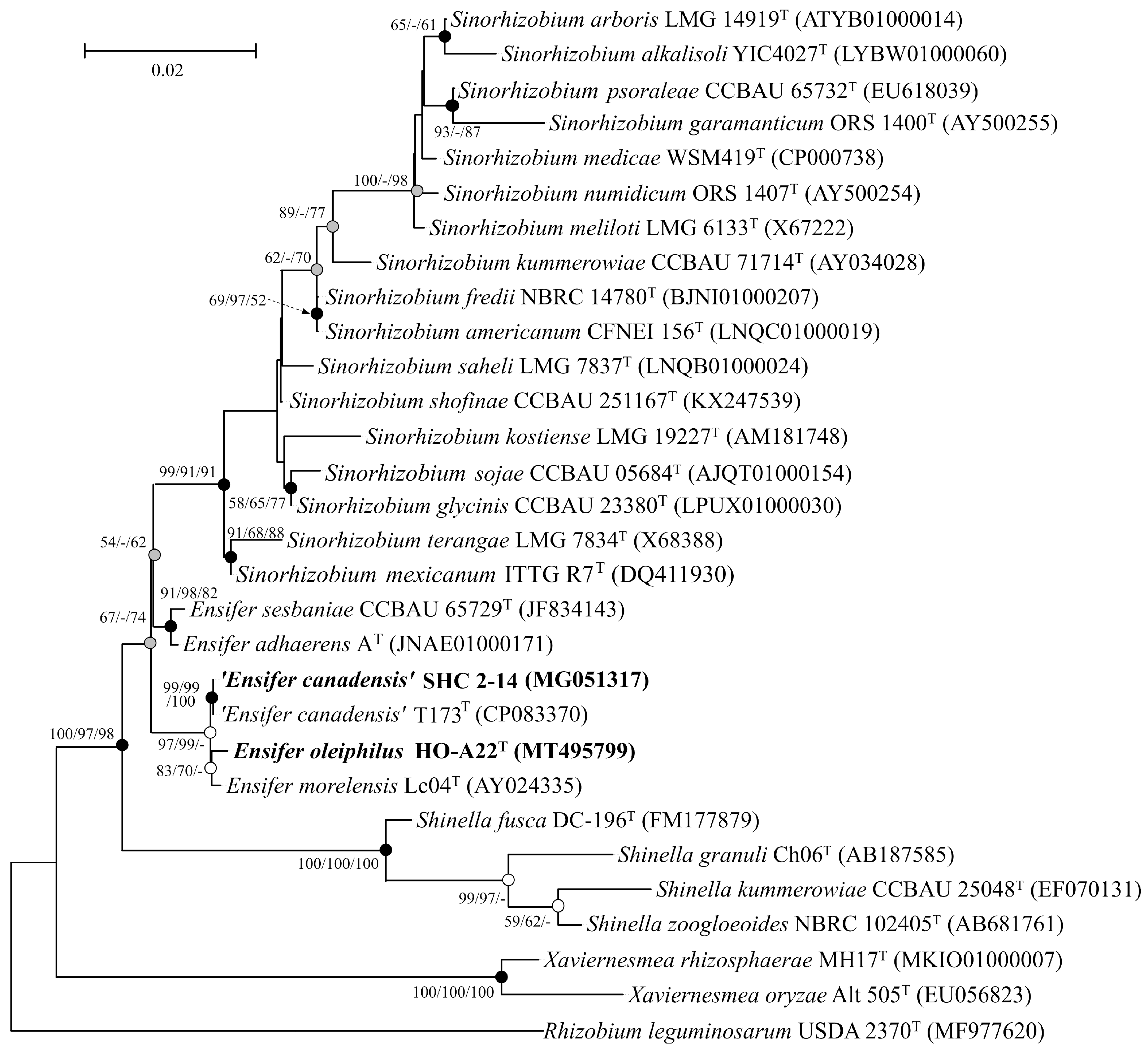
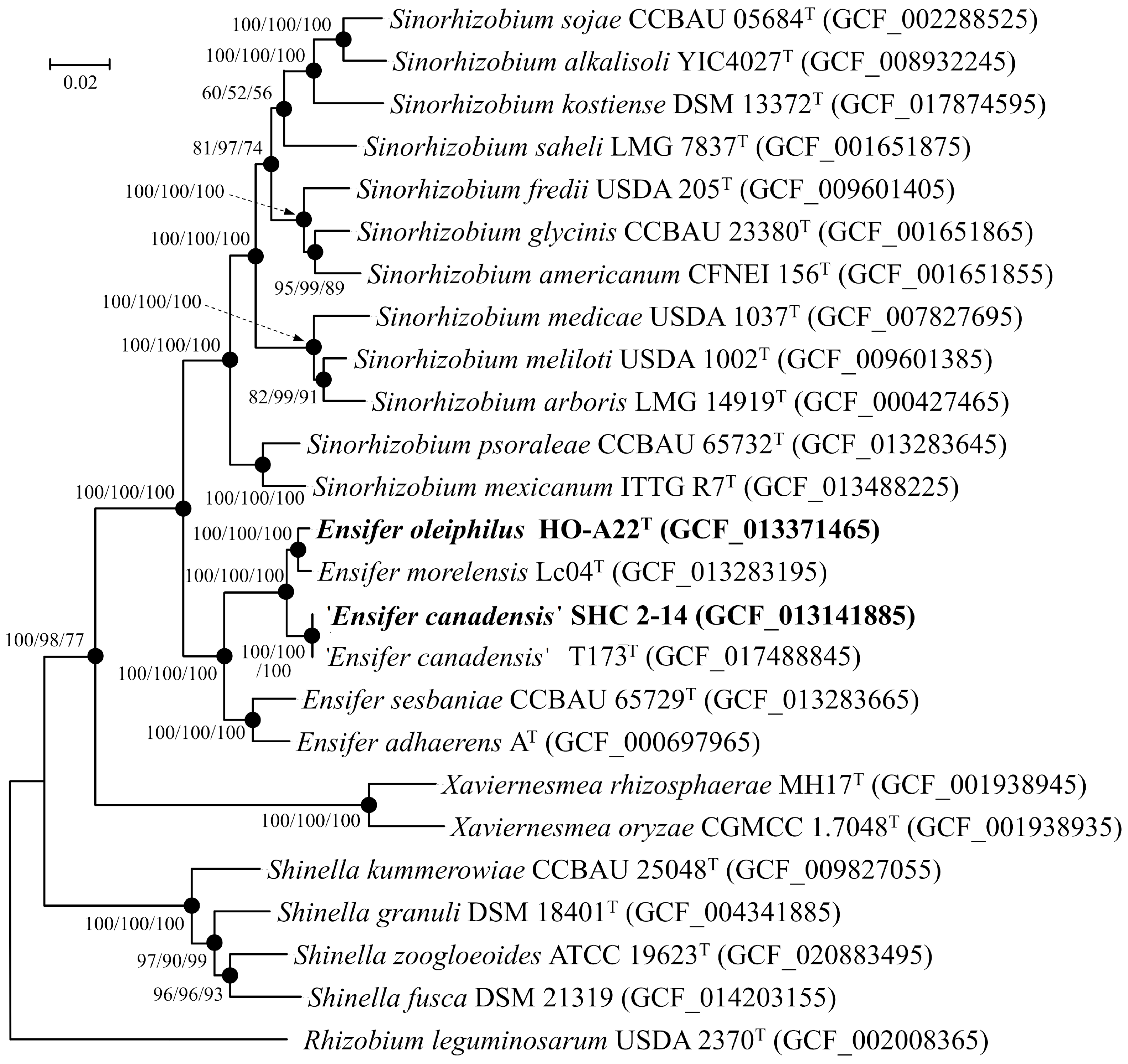
3.3. Phylogenetic Analysis
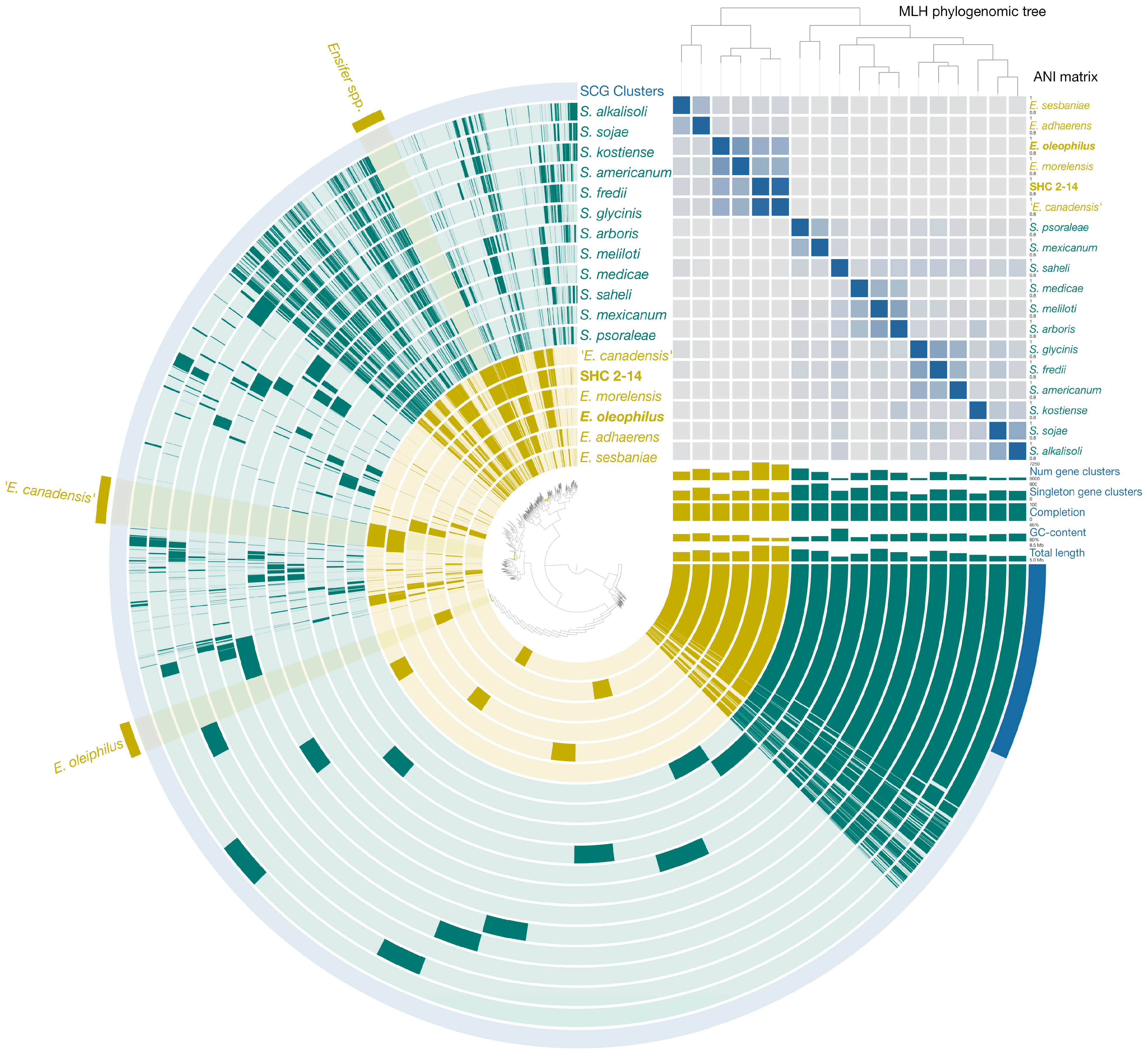
3.4. Genomic Analysis and Environmental Implications
3.5. Physiological Characterization
3.6. Chemotaxonomic Characterization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Willems, A.; Fernández-López, M.; Muñoz-Adelantado, E.; Goris, J.; De Vos, P.; Martínez-Romero, E.; Toro, N.; Gillis, M. Description of new Ensifer strains from nodules and proposal to transfer Ensifer adhaerens Casida 1982 to Sinorhizobium as Sinorhizobium adhaerens comb. nov. Request for an Opinion. Int. J. Syst. Evol. Microbiol. 2003, 53, 1207–1217. [Google Scholar] [CrossRef]
- Young, J.M. The genus name Ensifer Casida 1982 takes priority over Sinorhizobium Chen et al. 1988, and Sinorhizobium morelense Wang et al. 2002 is a later synonym of Ensifer adhaerens Casida 1982. Is the combination ‘Sinorhizobium adhaerens’ (Casida 1982) Willems et al. 2003 legitimate? Request for an Opinion. Int. J. Syst. Evol. Microbiol. 2003, 53, 2107–2110. [Google Scholar] [CrossRef]
- Young, J.M. Sinorhizobium versus Ensifer: May a taxonomy subcommittee of the ICSP contradict the Judicial Commission? Int. J. Syst. Evol. Microbiol. 2010, 60, 1711–1713. [Google Scholar] [CrossRef][Green Version]
- Wang, Y.C.; Wang, F.; Hou, B.C.; Wang, E.T.; Chen, W.F.; Sui, X.H.; Chen, W.X.; Li, Y.; Zhang, Y.B. Proposal of Ensifer psoraleae sp. nov.; Ensifer sesbaniae sp. nov., Ensifer morelense comb. nov. and Ensifer americanum comb. nov. Syst. Appl. Microbiol. 2013, 36, 467–473. [Google Scholar] [CrossRef]
- Kuzmanović, N.; Fagorzi, C.; Mengoni, A.; Lassalle, F.; diCenzo, G.C. Taxonomy of Rhizobiaceae revisited: Proposal of a new framework for genus delimitation. Int. J. Syst. Evol. Microbiol. 2022, 72, 005243. [Google Scholar] [CrossRef]
- Bromfield, E.S.P.; Cloutier, S.; Hynes, M.F. Ensifer canadensis sp. nov. strain T173T isolated from Melilotus albus (sweet clover) in Canada possesses recombinant plasmid pT173b harbouring symbiosis and type IV secretion system genes apparently acquired from Ensifer medicae. Front. Microbiol. 2023, 14, 1195755. [Google Scholar] [CrossRef] [PubMed]
- Casida, L.E., Jr. Ensifer adhaerens gen. nov., sp. nov.: A bacterial predator of bacteria in soil. Int. J. Syst. Bacteriol. 1982, 32, 339–345. [Google Scholar] [CrossRef]
- DiCenzo, G.C.; Debiec, K.; Krzysztoforski, J.; Uhrynowski, W.; Mengoni, A.; Fagorzi, C.; Gorecki, A.; Dziewit, L.; Bajda, T.; Rzepa, G.; et al. Genomic and biotechnological characterization of the heavy-metal resistant, arsenic-oxidizing bacterium Ensifer sp. M14. Genes 2018, 9, 379. [Google Scholar] [CrossRef]
- Ma, L.; Chen, S.; Yuan, J.; Yang, P.; Liu, Y.; Stewart, K. Rapid biodegradation of atrazine by Ensifer sp. strain and its degradation genes. Int. Biodeterior. Biodegrad. 2017, 116, 133–140. [Google Scholar] [CrossRef]
- Zhao, F.; Guo, C.; Cui, Q.; Hao, Q.; Xiu, J.; Han, S.; Zhang, Y. Exopolysaccharide production by an indigenous isolate Pseudomonas stutzeri XP1 and its application potential in enhanced oil recovery. Carbohydr. Polym. 2018, 199, 375–381. [Google Scholar] [CrossRef]
- Alvarez, V.M.; Jurelevicius, D.; Serrato, R.V.; Barreto-Bergter, E.; Seldin, L. Chemical characterization and potential application of exopolysaccharides produced by Ensifer adhaerens JHT2 as a bioemulsifier of edible oils. Int. J. Biol. Macromol. 2018, 114, 18–25. [Google Scholar] [CrossRef]
- Lehman, A.P.; Long, S.R. Exopolysaccharides from Sinorhizobium meliloti can protect against H2O2-dependent damage. J. Bacteriol. 2013, 195, 5362–5369. [Google Scholar] [CrossRef] [PubMed]
- Quijada, N.M.; Abreu, I.; Reguera, M.; Bonilla, I.; Bolaños, L. Boron nutrition affects growth, adaptation to stressful environments, and exopolysaccharide synthesis of Ensifer meliloti. Rhizosphere 2022, 22, 100534. [Google Scholar] [CrossRef]
- Xu, L.; Chen, X.; Li, H.; Hu, F.; Liang, M. Characterization of the biosorption and biodegradation properties of Ensifer adhaerens: A potential agent to remove polychlorinated biphenyls from contaminated water. J. Hazard. Mater. 2016, 302, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; An, D.; Cavallaro, A.; Voordouw, G. Souring in low-temperature surface facilities of two high-temperature Argentinian oil fields. Appl. Microbiol. Biotechnol. 2014, 98, 8017–8029. [Google Scholar] [CrossRef] [PubMed]
- An, B.A.; Shen, Y.; Voordouw, G. Control of sulfide production in high salinity Bakken Shale oil reservoirs by halophilic bacteria reducing nitrate to nitrite. Front. Microbiol. 2017, 8, 1164. [Google Scholar] [CrossRef]
- Sokolova, D.S.; Semenova, E.M.; Grouzdev, D.S.; Ershov, A.P.; Bidzhieva, S.K.; Ivanova, A.E.; Babich, T.L.; Sissenbayeva, M.R.; Bisenova, M.A.; Nazina, T.N. Microbial diversity and potential sulfide producers in the Karazhanbas oilfield (Kazakhstan). Microbiology 2020, 89, 459–469. [Google Scholar] [CrossRef]
- Torres, M.J.; Avila, S.; Bedmar, E.J.; Delgado, M.J. Overexpression of the periplasmic nitrate reductase supports anaerobic growth by Ensifer meliloti. FEMS Microbiol. Lett. 2018, 365, fny041. [Google Scholar] [CrossRef]
- Liu, L.X.; Li, Q.Q.; Zhang, Y.Z.; Hu, Y.; Jiao, J.; Guo, H.J.; Zhang, X.X.; Zhang, B.; Chen, W.X.; Tian, C.F. The nitrate-reduction gene cluster components exert lineage-dependent contributions to optimization of Sinorhizobium symbiosis with soybeans. Environ. Microbiol. 2017, 19, 4926–4938. [Google Scholar] [CrossRef]
- Ruiz, B.; Le Scornet, A.; Sauviac, L.; Rémy, A.; Bruand, C.; Meilhoc, E. The nitrate assimilatory pathway in Sinorhizobium meliloti: Contribution to NO production. Front. Microbiol. 2019, 10, 1526. [Google Scholar] [CrossRef]
- Safonov, A.V.; Babich, T.L.; Sokolova, D.S.; Grouzdev, D.S.; Tourova, T.P.; Poltaraus, A.B.; Zakharova, E.V.; Merkel, A.Y.; Novikov, A.P.; Nazina, T.N. Microbial community and in situ bioremediation of groundwater by nitrate removal in the zone of a radioactive waste surface repository. Front. Microbiol. 2018, 9, 1985. [Google Scholar] [CrossRef]
- Nazina, T.N.; Babich, T.L.; Kostryukova, N.K.; Sokolova, D.S.; Abdullin, R.R.; Tourova, T.P.; Poltaraus, A.B.; Kalmykov, S.N.; Zakharova, E.V.; Myasoedov, B.F.; et al. Microbial diversity and possible activity in nitrate- and radionuclide-contaminated groundwater. In Behavior of Radionuclides in the Environment I. Function of Particles in Aquatic System; Kato, K., Konoplev, A., Kalmykov, S.N., Eds.; Springer Nature Singapore Pte. Ltd.: Singapore, 2020; pp. 35–66. [Google Scholar] [CrossRef]
- Safonov, A.V.; Andryushchenko, N.D.; Ivanov, P.V.; Boldyrev, K.A.; Babich, T.L.; German, K.E.; Zakharova, E.V. Biogenic factors of radionuclide immobilization on sandy rocks of upper aquifers. Radiochemistry 2019, 61, 99–108. [Google Scholar] [CrossRef]
- Gadd, G.M. Metals, minerals and microbes: Geomicrobiology and bioremediation. Microbiology 2010, 156, 609–643. [Google Scholar] [CrossRef] [PubMed]
- Nazina, T.N.; Luk’yanova, E.A.; Zakharova, E.V.; Konstantinova, L.I.; Kalmykov, S.N.; Poltaraus, A.B.; Zubkov, A.A. Microorganisms in a disposal site for liquid radioactive wastes and their influence on radionuclides. Geomicrobiol. J. 2010, 27, 473–486. [Google Scholar] [CrossRef]
- Shukla, A.; Parmar, P.; Saraf, M. Radiation, radionuclides and bacteria: An in-perspective review. J. Environ. Radioact. 2017, 180, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R.; Phillips, E.J.P.; Gorby, Y.A.; Landa, E.R. Microbial reduction of uranium. Nature 1991, 350, 413–416. [Google Scholar] [CrossRef]
- Lloyd, J.R. Microbial reduction of metals and radionuclides. FEMS Microbiol. Rev. 2003, 27, 411–425. [Google Scholar] [CrossRef]
- Nazina, T.; Babich, T.; Kostryukova, N.; Sokolova, D.; Abdullin, R.; Tourova, T.; Kadnikov, V.; Mardanov, A.; Ravin, N.; Grouzdev, D.; et al. Ultramicrobacteria from nitrate- and radionuclide-contaminated groundwater. Sustainability 2020, 12, 1239. [Google Scholar] [CrossRef]
- Nazina, T.N.; Sokolova, D.S.; Babich, T.L.; Semenova, E.M.; Ershov, A.P.; Bidzhieva, S.K.; Borzenkov, I.A.; Poltaraus, A.B.; Khisametdinov, M.R.; Tourova, T.P. Microorganisms of low-temperature heavy oil reservoirs (Russia) and their possible application for enhanced oil recovery. Microbiology 2017, 86, 773–785. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Vinh, L.S.; Flouri, T.; Stamatakis, A.; von Haeseler, A.; Minh, B.Q. MPBoot: Fast phylogenetic maximum parsimony tree inference and bootstrap approximation. BMC Evol. Biol. 2018, 18, 11. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Conesa, A.; García-Alcalde, F. Qualimap 2: Advanced multi-sample quality control for high-throughput sequencing data. Bioinformatics 2016, 32, 292–294. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Chaumeil, P.; Mussig, A.J.; Parks, D.H.; Hugenholtz, P. GTDB-Tk: A toolkit to classify genomes with the Genome Taxonomy Database. Bioinformatics 2020, 36, 1925–1927. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinf. 2013, 14, 60. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef]
- Grouzdev, D.S.; Rysina, M.S.; Bryantseva, I.A.; Gorlenko, V.M.; Gaisin, V.A. Draft genome sequences of ‘Candidatus Chloroploca asiatica’ and ‘Candidatus Viridilinea mediisalina’, candidate representatives of the Chloroflexales order: Phylogenetic and taxonomic implications. Stand. Genomic. Sci. 2018, 13, 24. [Google Scholar] [CrossRef]
- Qin, Q.-L.; Xie, B.-B.; Zhang, X.-Y.; Chen, X.-L.; Zhou, B.-C.; Zhou, J.; Oren, A.; Zhang, Y.-Z. A proposed genus boundary for the Prokaryotes based on genomic insights. J. Bacteriol. 2014, 196, 2210–2215. [Google Scholar] [CrossRef]
- Delmont, T.O.; Eren, E.M. Linking pangenomes and metagenomes: The Prochlorococcus metapangenome. PeerJ 2018, 6, e4320. [Google Scholar] [CrossRef]
- Eren, A.M.; Esen, O.C.; Quince, C.; Vineis, J.H.; Morrison, H.G.; Sogin, M.L.; Delmont, T.O. Anvi’o: An advanced analysis and visualization platform for ’omics data. PeerJ 2015, 3, e1319. [Google Scholar] [CrossRef]
- Sokolova, D.S.; Semenova, E.M.; Grouzdev, D.S.; Bidzhieva, S.K.; Babich, T.L.; Loiko, N.G.; Ershov, A.P.; Kadnikov, V.V.; Beletsky, A.V.; Mardanov, A.V.; et al. Sulfidogenic microbial communities of the Uzen high-temperature oil field in Kazakhstan. Microorganisms 2021, 9, 1818. [Google Scholar] [CrossRef] [PubMed]
- Adkins, J.P.; Cornell, L.A.; Tanner, R.S. Microbial composition of carbonate petroleum reservoir fluids. Geomicrobiol. J. 1992, 10, 87–97. [Google Scholar] [CrossRef]
- Pfennig, N.; Lippert, K.D. Über das Vitamin B12-Bedürfnis phototropher Schwefelbakterien. Arch. Mikrobiol. 1966, 55, 245–256. [Google Scholar] [CrossRef]
- Bidzhieva, S.K.; Sokolova, D.S.; Grouzdev, D.S.; Kostrikina, N.A.; Poltaraus, A.B.; Tourova, T.P.; Shcherbakova, V.A.; Troshina, O.Y.; Nazina, T.N. Sphaerochaeta halotolerans sp. nov., a novel spherical halotolerant spirochete from a Russian heavy oil reservoir, emended description of the genus Sphaerochaeta, reclassification of Sphaerochaeta coccoides to a new genus Parasphaerochaeta gen. nov. as Parasphaerochaeta coccoides comb. nov. and proposal of Sphaerochaetaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 4748–4759. [Google Scholar] [CrossRef]
- Collins, M.D.; Jones, D. Distribution of isoprenoid quinone structural types in bacteria and their taxonomic implication. Microbiol. Rev. 1981, 45, 316–354. [Google Scholar] [CrossRef] [PubMed]
- Minnikin, D.E.; O’Donnell, A.G.; Goodfellow, M.; Alderson, G.; Athalye, M.; Schaal, A.; Parlett, J.H. An integrated procedure for the extraction of bacterial isoprenoid quinones and polar lipids. J. Microbiol. Methods 1984, 2, 233–241. [Google Scholar] [CrossRef]
- Semenova, E.M.; Grouzdev, D.S.; Sokolova, D.S.; Tourova, T.P.; Poltaraus, A.B.; Potekhina, N.V.; Shishina, P.N.; Bolshakova, M.A.; Avtukh, A.N.; Ianutsevich, E.A.; et al. Physiological and genomic characterization of Actinotalea subterranea sp. nov. from oil-degrading methanogenic enrichment and reclassification of the family Actinotaleaceae. Microorganisms 2022, 10, 378. [Google Scholar] [CrossRef]
- Semenova, E.M.; Babich, T.L.; Sokolova, D.S.; Ershov, A.P.; Raievska, Y.I.; Bidzhieva, S.K.; Stepanov, A.L.; Korneykova, M.V.; Myazin, V.A.; Nazina, T.N. Microbial communities of seawater and coastal soil of Russian Arctic region and their potential for bioremediation from hydrocarbon pollutants. Microorganisms 2022, 10, 1490. [Google Scholar] [CrossRef]
- Koziaeva, V.; Dziuba, M.; Leão, P.; Uzun, M.; Krutkina, M.; Grouzdev, D. Genome-based metabolic reconstruction of a novel uncultivated freshwater magnetotactic coccus “Ca. Magnetaquicoccus inordinatus” UR-1, and proposal of a candidate family “Ca. Magnetaquicoccaceae”. Front. Microbiol. 2019, 10, 2290. [Google Scholar] [CrossRef]
- Li, Y.; Xue, H.; Sang, S.; Lin, C.; Wang, X. Phylogenetic analysis of family Neisseriaceae based on genome sequences and description of Populibacter corticis gen. nov., sp. nov., a member of the family Neisseriaceae, isolated from symptomatic bark of Populus × euramericana canker. PLoS ONE 2017, 12, e0174506. [Google Scholar] [CrossRef] [PubMed]
- Orata, F.D.; Meier-Kolthoff, J.P.; Sauvageau, D.; Stein, L.Y. Phylogenomic analysis of the gammaproteobacterial methanotrophs (order Methylococcales) calls for the reclassification of members at the genus and species levels. Front. Microbiol. 2018, 9, 3162. [Google Scholar] [CrossRef] [PubMed]
- diCenzo, G.C.; Muhammed, Z.; Østerås, M.; O’Brien, S.A.; Finan, T.M. A key regulator of the glycolytic and gluconeogenic central metabolic pathways in Sinorhizobium meliloti. Genetics 2017, 207, 961–974. [Google Scholar] [CrossRef]
- Draghi, W.O.; Del Papa, M.F.; Barsch, A.; Albicoro, F.J.; Lozano, M.J.; Pühler, A.; Niehaus, K.; Lagares, A. A metabolomic approach to characterize the acid-tolerance response in Sinorhizobium meliloti. Metabolomics 2017, 13, 71. [Google Scholar] [CrossRef]
- Geddes, B.A.; Oresnik, I.J. Physiology, genetics, and biochemistry of carbon metabolism in the alphaproteobacterium Sinorhizobium meliloti. Can. J. Microbiol. 2014, 60, 491–507. [Google Scholar] [CrossRef]
- Hawkins, J.P.; Ordonez, P.A.; Oresnik, I.J. Characterization of mutations that affect the nonoxidative pentose phosphate pathway in Sinorhizobium meliloti. J. Bacteriol. 2018, 200, e00436-17. [Google Scholar] [CrossRef]
- Yurgel, S.N.; Qu, Y.; Rice, J.T.; Ajeethan, N.; Zink, E.M.; Brown, J.M.; Purvine, S.; Lipton, M.S.; Kahn, M.L. Specialization in a nitrogen-fixing symbiosis: Proteome differences between Sinorhizobium medicae bacteria and bacteroids. Mol. Plant-Microbe Interact. 2021, 34, 1409–1422. [Google Scholar] [CrossRef]
- Torres, M.J.; Rubia, M.I.; Bedmar, E.J.; Delgado, M.J. Denitrification in Sinorhizobium meliloti. Biochem. Soc. Trans. 2011, 39, 1886–1889. [Google Scholar] [CrossRef]
- Torres, M.J.; Rubia, M.I.; Coba de la Peña, T.; Pueyo, J.J.; Bedmar, E.J.; Delgado, M.J. Genetic basis for denitrification in Ensifer meliloti. BMC Microbiol. 2014, 14, 142. [Google Scholar] [CrossRef]
- Rivers, D.M.; Kim, D.D.; Oresnik, I.J. Inability to catabolize rhamnose by Sinorhizobium meliloti Rm1021 affects competition for nodule occupancy. Microorganisms 2022, 10, 732. [Google Scholar] [CrossRef]
- Zivkovic, M.; Miljkovic, M.; Ruas-Madiedo, P.; Strahinic, I.; Tolinacki, M.; Golic, N.; Kojic, M. Exopolysaccharide production and ropy phenotype are determined by two gene clusters in putative probiotic strain Lactobacillus paraplantarum BGCG11. Appl. Environ. Microbiol. 2015, 81, 1387–1396. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; D’Haeze, W.; De Rycke, R.; Wolucka, B.; Holsters, M. Knockout of an azorhizobial dTDP-L-rhamnose synthase affects lipopolysaccharide and extracellular polysaccharide production and disables symbiosis with Sesbania rostrata. Mol. Plant-Microbe Interact. 2001, 14, 857–866. [Google Scholar] [CrossRef]
- Jofré, E.; Lagares, A.; Mori, G. Disruption of dTDP-rhamnose biosynthesis modifies lipopolysaccharide core, exopolysaccharide production, and root colonization in Azospirillum brasilense. FEMS Microbiol. Lett. 2004, 231, 267–275. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhan, H.J.; Leigh, J.A. Two genes that regulate exopolysaccharide production in Rhizobium meliloti. J. Bacteriol. 1990, 172, 5254–5259. [Google Scholar] [CrossRef]
- Pourbabaee, A.A.; Khazaei, M.; Alikhani, H.A.; Emami, S. Root nodulation of alfalfa by Ensifer meliloti in petroleum contaminated soil. Rhizosphere 2021, 17, 100305. [Google Scholar] [CrossRef]
- Keum, Y.S.; Seo, J.S.; Hu, Y.; Li, Q.X. Degradation pathways of phenanthrene by Sinorhizobium sp. C4. Appl. Microbiol. Biotechnol. 2006, 71, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Muratova, A.; Pozdnyakova, N.; Makarov, O.; Baboshin, M.; Baskunov, B.; Myasoedova, N.; Golovleva, L.; Turkovskaya, O. Degradation of phenanthrene by the rhizobacterium Ensifer meliloti. Biodegradation 2014, 25, 787–795. [Google Scholar] [CrossRef]
- Turkovskaya, O.; Muratova, A. Plant–bacterial degradation of polyaromatic hydrocarbons in the rhizosphere. Trends Biotechnol. 2019, 37, 926–930. [Google Scholar] [CrossRef]
- Sherpa, M.T.; Najar, I.N.; Das, S.; Thakur, N. Distribution of antibiotic and metal resistance genes in two glaciers of North Sikkim, India. Ecotoxicol. Environ. Saf. 2020, 203, 111037. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Deng, W.; Liu, S.; Yu, X.; Mustafa, G.R.; Chen, S.; Chen, S.; He, L.; Ao, X.; Yang, Y.; et al. Presence of heavy metal resistance genes in Escherichia coli and Salmonella isolates and analysis of resistance gene structure in E. coli E308. J. Glob. Antimicrob. Resist. 2020, 21, 420–426. [Google Scholar] [CrossRef]
- García-Cañas, R.; Giner-Lamia, J.; Florencio, F.J.; López-Maury, L. A protease-mediated mechanism regulates the cytochrome c6/plastocyanin switch in Synechocystis sp. PCC 6803. Proc. Natl. Acad. Sci. USA 2021, 118, e2017898118. [Google Scholar] [CrossRef]
- Ge, Q.; Zhu, X.; Cobine, P.A.; De La Fuente, L. The copper-binding protein CutC is involved in copper homeostasis and affects virulence in the xylem-limited pathogen Xylella fastidiosa. Phytopathology 2022, 112, 1620–1629. [Google Scholar] [CrossRef]
- Hyre, A.; Casanova-Hampton, K.; Subashchandrabose, S. Copper homeostatic mechanisms and their role in the virulence of Escherichia coli and Salmonella enterica. EcoSal Plus 2021, 9, eESP-0014-2020. [Google Scholar] [CrossRef]
- Salam, L.B. Metagenomic insights into the microbial community structure and resistomes of a tropical agricultural soil persistently inundated with pesticide and animal manure use. Folia Microbiol. 2022, 67, 707–719. [Google Scholar] [CrossRef] [PubMed]
- Toulouse, C.; Metesch, K.; Pfannstiel, J.; Steuber, J. Metabolic reprogramming of Vibrio cholerae impaired in respiratory NADH oxidation is accompanied by increased copper sensitivity. J. Bacteriol. 2018, 200, e00761-17. [Google Scholar] [CrossRef]
- Al-Ansari, M.M. Biodetoxification mercury by using a marine bacterium Marinomonas sp. RS3 and its merA gene expression under mercury stress. Environ. Res. 2022, 205, 112452. [Google Scholar] [CrossRef]
- Babich, T.L.; Grouzdev, D.S.; Sokolova, D.S.; Tourova, T.P.; Poltaraus, A.B.; Nazina, T.N. Genome analysis of Pollutimonas subterranea gen. nov.; sp. nov. and Pollutimonas nitritireducens sp. nov.; isolated from nitrate-and radionuclide-contaminated groundwater, and transfer of several Pusillimonas species into three new genera Allopusillimonas, Neopusillimonas, and Mesopusillimonas. Antonie Van Leeuwenhoek 2023, 116, 109–127. [Google Scholar] [CrossRef]
- Binish, M.B.; Shini, S.; Sinha, R.K.; Krishnan, K.P.; Mohan, M. Mercuric reductase gene (merA) activity in a mercury tolerant sulphate reducing bacterium isolated from the Kongsfjorden. Arctic. Polar Sci. 2021, 30, 100745. [Google Scholar] [CrossRef]
- Lo, K.H.; Lu, C.W.; Liu, F.G.; Kao, C.M.; Chen, S.C. Draft genome sequence of Pseudomonas sp. A46 isolated from mercury-contaminated wastewater. J. Basic Microbiol. 2022, 62, 1193–1201. [Google Scholar] [CrossRef]
- Almeida, M.C.; Branco, R.; Morais, P.V. Response to vanadate exposure in Ochrobactrum tritici strains. PLoS ONE 2020, 15, e0229359. [Google Scholar] [CrossRef] [PubMed]
- Princy, S.; Subramanian, D.; Natarajan, J.; Prabagaran, S.R. Analysis of chromate transporters in bacterial species for Cr(VI) reduction isolated from tannery effluent contaminated site of Dindigul district, Tamil Nadu, India. Geomicrobiol. J. 2021, 38, 598–606. [Google Scholar] [CrossRef]
- Semenova, E.M.; Ershov, A.P.; Sokolova, D.S.; Tourova, T.P.; Nazina, T.N. Diversity and biotechnological potential of nitrate-reducing bacteria from heavy-oil reservoirs (Russia). Microbiology 2020, 89, 685–696. [Google Scholar] [CrossRef]
- Tóth, E.; Szuróczki, S.; Kéki, Z.; Bóka, K.; Szili-Kovács, T.; Schumann, P. Gellertiella hungarica gen. nov., sp. nov., a novel bacterium of the family Rhizobiaceae isolated from a spa in Budapest. Int. J. Syst. Evol. Microbiol. 2017, 67, 4565–4571. [Google Scholar] [CrossRef]
- Wang, E.T.; Tan, Z.Y.; Willems, A.; Fernández-López, M.; Reinhold-Hurek, B.; Martínez-Romero, E. Sinorhizobium morelense sp. nov.; a Leucaena leucocephala-associated bacterium that is highly resistant to multiple antibiotics. Int. J. Syst. Evol. Microbiol. 2002, 52, 1687–1693. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genome | HO-A22T | SHC 2-14 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 16S rRNA | dDDH | ANI | AAI | POCP | 16S rRNA | dDDH | ANI | AAI | POCP | |
| HO-A22T | 100 | 100 | 100 | 100 | 100 | 99.8 | 35.8 | 88.7 | 91.6 | 80.6 |
| SHC 2-14 | 99.8 | 35.8 | 88.7 | 91.7 | 80.6 | 100 | 100 | 100 | 100 | 100 |
| E. adhaerens AT | 99.1 | 25.4 | 84.1 | 84.1 | 79.7 | 99.2 | 25.4 | 83.8 | 83.5 | 78.9 |
| E. canadensis T173T | 99.8 | 35.8 | 87.9 | 91.8 | 80.8 | 100 | 92.8 | 99.1 | 99.1 | 88.7 |
| E. morelensis Lc04T | 99.9 | 45.9 | 92.4 | 95.1 | 87.4 | 99.9 | 35.6 | 88.7 | 91.4 | 79.3 |
| E. sesbaniae CCBAU 65729T | 99.0 | 25.6 | 83.7 | 84.3 | 75.6 | 99.1 | 25.8 | 83.7 | 83.8 | 74.1 |
| Characteristics | ‘E. canadensis’ | ‘E. canadensis’ | E. adhaerens | E. morelensis | E. sesbaniae | |
|---|---|---|---|---|---|---|
| Strain | HO-A22T | SHC 2-14 | T173T | AT | Lc04T | CCBAU 65729T |
| Catalase | + | + | ND | w | + | – |
| Citrate utilization | – | – | ND | – | – | + |
| Urease | – | – | ND | + | – | + |
| Growth at: | ||||||
| 2% NaCl | + | – | + | + | + | – |
| 3% NaCl | w | – | – | w | w | – |
| 15 °C | + | + | + | + | + | – |
| 37 °C | – | – | w | w | w | + |
| pH 5 | – | – | + | – | – | – |
| pH 10 | w | – | + | w | w | – |
| Acid production (API® 50CH) from: | ||||||
| D-fucose | + | + | – | + | + | ND |
| D-galactose | + | w | – | + | + | ND |
| D-lactose | + | – | w | + | + | + |
| D-melicitose | – | – | ND | w | – | + |
| D-turanose | + | w | – | + | + | ND |
| Dulcitol | + | – | ND | – | w | – |
| Gentiobiose | + | – | w | w | + | ND |
| L-rhamnose | + | w | – | + | + | + |
| L-sorbose | + | – | ND | – | w | + |
| Salicin | + | – | – | – | + | + |
| Major fatty acids | C18:1, C16:0, C14:0 3-OH, C16:1, C17:1 | C18:1, C16:0, C16:1, C14:0 3-OH, C18:0 | C18:1, C16:0, C12:0 aldehyde, C18:0, C18:0 cyclo ω8c | C18:1, C16:0, C16:1, C14:0 3-OH, C18:0, C18:0 3-OH | C18:1, C16:0, C16:0 3-OH, C18:0, C16:1 | C18:1, C12:0 aldehyde, C16:0, C16:1, C18:0 |
| Major polar lipids | PC, PE, DPG, PG | PC, DPG, PE, APL4, PG | ND | PE, PC, PG, DPG | PE, DPG, PC, PG | PC, PE, PG |
| G+C content, % | 61.7 | 61.0 | 61.0 | 62.8 a | 61.7 b | 60.4 |
| Isolation source | Injection water of an oil field | Radionuclide-contaminated groundwater | Root nodule of Melilotus albus | Soil a | Root nodule of Leucaena leucocephala b | Root nodule of Sesbania cannabina |
| Parameter | Ensifer oleiphilus sp. nov. |
|---|---|
| Genus name | Ensifer |
| Species name | Ensifer oleiphilus |
| Species status | sp. nov. |
| Species etymology | o.le.i.phi’lus. L. neut. n. oleum oil; N.L. masc. adj. philus (from Gr. masc. adj. philos) loving; N.L. masc. adj. oleiphilus loving oil, referring to the ability of type strain to utilize hydrocarbons of crude oil |
| Description of the new taxon and diagnostic traits | Cells are Gram-negative, motile, aerobic, non-spore-forming rods. Colonies up to 8.0 in diameter are creamy, smooth, shiny, and round after 5 days of cultivation in the TEG medium at 22 °C. Growth occurs on TEG, PCA, and LB media, in the presence of 0–3.0% (w/v) NaCl (optimum, 0–1.2% NaCl), at pH 6.0–9.8 (optimum, pH 6.6–8.0) and at 15–33 °C (optimum, 22–28 °C). Catalase-positive, but negative for urease and citrate utilization. Chemoorganoheterotrophic, facultatively anaerobic. Nitrate is reduced to the nitrous oxide. Acid is produced from D-fucose, D-galactose, D-lactose, D-turanose, dulcitol, gentiobiose, L-rhamnose, L-sorbose, and salicin, but not from D-melicitose. The predominant fatty acids are C18:1, C16:0, C14:0 3-OH, C16:1, C17:1, and C18:0. Major menaquinone is Q10. The major polar lipids are phosphatidylcholine, phosphatidylethanolamine, diphosphatidylglycerol, and phosphatidylglycerol. The genome size of the type strain is 6.76 Mb with a G+C value of 61.7%. The strain type, HO-A22T (=VKM B-3646T = KCTC 92427T), was isolated from a denitrifying enrichment obtained from an injection water of the Cheremukhovskoe oil field (Tatarstan, Russia). The GenBank/EMBL/DDBJ accession number for the 16S rRNA gene sequence is MT495799 and the genomic assembly accession number is GCF_013371465.1. |
| Country of origin | Russian Federation |
| Region of origin | Nurlat region, Tatarstan |
| Date of isolation | 2018 |
| Source of isolation | Injection water of an oil field |
| Sampling date | 2016 |
| Latitude | 55°0′ N |
| Longitude | 51°1′ E |
| Depth (meters below sea level) | 850–1300 |
| Number of strains in study | 1 |
| Information related to the Nagoya Protocol | Not applicable |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ershov, A.P.; Babich, T.L.; Grouzdev, D.S.; Sokolova, D.S.; Semenova, E.M.; Avtukh, A.N.; Poltaraus, A.B.; Ianutsevich, E.A.; Nazina, T.N. Genome Analysis and Potential Ecological Functions of Members of the Genus Ensifer from Subsurface Environments and Description of Ensifer oleiphilus sp. nov. Microorganisms 2023, 11, 2314. https://doi.org/10.3390/microorganisms11092314
Ershov AP, Babich TL, Grouzdev DS, Sokolova DS, Semenova EM, Avtukh AN, Poltaraus AB, Ianutsevich EA, Nazina TN. Genome Analysis and Potential Ecological Functions of Members of the Genus Ensifer from Subsurface Environments and Description of Ensifer oleiphilus sp. nov. Microorganisms. 2023; 11(9):2314. https://doi.org/10.3390/microorganisms11092314
Chicago/Turabian StyleErshov, Alexey P., Tamara L. Babich, Denis S. Grouzdev, Diyana S. Sokolova, Ekaterina M. Semenova, Alexander N. Avtukh, Andrey B. Poltaraus, Elena A. Ianutsevich, and Tamara N. Nazina. 2023. "Genome Analysis and Potential Ecological Functions of Members of the Genus Ensifer from Subsurface Environments and Description of Ensifer oleiphilus sp. nov." Microorganisms 11, no. 9: 2314. https://doi.org/10.3390/microorganisms11092314
APA StyleErshov, A. P., Babich, T. L., Grouzdev, D. S., Sokolova, D. S., Semenova, E. M., Avtukh, A. N., Poltaraus, A. B., Ianutsevich, E. A., & Nazina, T. N. (2023). Genome Analysis and Potential Ecological Functions of Members of the Genus Ensifer from Subsurface Environments and Description of Ensifer oleiphilus sp. nov. Microorganisms, 11(9), 2314. https://doi.org/10.3390/microorganisms11092314