

Mining Autoimmune-Disorder-Linked Molecular-Mimicry Candidates in Clostridioides difficile and Prospects of Mimic-Based Vaccine Design: An In Silico Approach

 , , , ,
, , , , 
Abstract
1. Introduction
2. Material and Methods
2.1. Homology Analysis
2.2. Mimic Region Identification
2.3. Autoimmunity Elucidation
2.4. Mimic-Based Vaccine Design
2.5. Immune Receptor Binding Study
3. Results
3.1. Homologous Sequence Identification
3.2. Autoimmunity Prediction
3.3. Sequence Mimics and Vaccine Design
3.4. Immune Response Simulation
3.5. Vaccine Interaction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martins, Y.C.; Jurberg, A.D.; Daniel-Ribeiro, C.T.J.M. Visiting Molecular Mimicry Once More: Pathogenicity, Virulence, and Autoimmunity. Microorganisms 2023, 11, 1472. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.M.; Rahal, E.A. The role of viral infections in the development of autoimmune diseases. Crit. Rev. Microbiol. 2019, 45, 394–412. [Google Scholar] [CrossRef] [PubMed]
- Topi, S.; Bottalico, L.; Charitos, I.A.; Colella, M.; Di Domenico, M.; Palmirotta, R.; Santacroce, L.J. Biomolecular Mechanisms of Autoimmune Diseases and Their Relationship with the Resident Microbiota: Friend or Foe? Pathophysiology 2022, 29, 507–536. [Google Scholar] [CrossRef]
- Guarneri, F. The Concept of Infection-Triggered Autoimmunity. In Role of Microorganisms in Pathogenesis and Management of Autoimmune Diseases: Volume I: Liver, Skin, Thyroid, Rheumatic & Myopathic Diseases; Springer: Berlin/Heidelberg, Germany, 2022; pp. 3–19. [Google Scholar]
- Mobasheri, L.; Nasirpour, M.H.; Masoumi, E.; Azarnaminy, A.F.; Jafari, M.; Esmaeili, S.-A. SARS-CoV-2 triggering autoimmune diseases. Cytokine 2022, 154, 155873. [Google Scholar] [CrossRef]
- Fugger, L.; Jensen, L.T.; Rossjohn, J.J. Challenges, progress, and prospects of developing therapies to treat autoimmune diseases. Cell 2020, 181, 63–80. [Google Scholar] [CrossRef] [PubMed]
- Burbelo, P.D.; Iadarola, M.J.; Keller, J.M.; Warner, B.M. Autoantibodies targeting intracellular and extracellular proteins in autoimmunity. Front. Immunol. 2021, 12, 548469. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, M.W. Molecular mimicry, autoimmunity, and infection: The cross-reactive antigens of group A streptococci and their sequelae. Microbiol. Spectr. 2019, 7, 20. [Google Scholar] [CrossRef]
- Adiguzel, Y.J. Molecular mimicry between SARS-CoV-2 and human proteins. Autoimmun. Rev. 2021, 20, 102791. [Google Scholar] [CrossRef]
- Venigalla, S.S.K.; Premakumar, S.; Janakiraman, V. A possible role for autoimmunity through molecular mimicry in alphavirus mediated arthritis. Sci. Rep. 2020, 10, 938. [Google Scholar] [CrossRef]
- Joubert, B.; Dalmau, J. The role of infections in autoimmune encephalitides. Rev. Neurol. 2019, 175, 420–426. [Google Scholar] [CrossRef]
- Cuan-Baltazar, Y.; Soto-Vega, E. Microorganisms associated to thyroid autoimmunity. Autoimmun. Rev. 2020, 19, 102614. [Google Scholar] [CrossRef] [PubMed]
- Youssefi, M.; Tafaghodi, M.; Farsiani, H.; Ghazvini, K.; Keikha, M. Helicobacter pylori infection and autoimmune diseases. Is there an association with systemic lupus erythematosus, rheumatoid arthritis, autoimmune atrophy gastritis and autoimmune pancreatitis? A systematic review and meta-analysis study. J. Microbiol. Immunol. Infect. 2021, 54, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Begum, S.; Aiman, S.; Ahmad, S.; Samad, A.; Almehmadi, M.; Allahyani, M.; Aljuaid, A.; Afridi, S.G.; Khan, A. Molecular Mimicry Analyses Unveiled the Human Herpes Simplex and Poxvirus Epitopes as Possible Candidates to Incite Autoimmunity. Pathogens 2022, 11, 1362. [Google Scholar] [CrossRef]
- Rahman, N.; Begum, S.; Khan, A.; Afridi, S.G.; Sahibzada, M.U.K.; Atwah, B.; Alhindi, Z.; Khan, H. An insight in Salmonella typhi associated autoimmunity candidates’ prediction by molecular mimicry. Comput. Biol. Med. 2022, 148, 105865. [Google Scholar] [CrossRef] [PubMed]
- Mashraqi, M.M.; Alzamami, A.; Alturki, N.A.; Alshamrani, S.; Alshahrani, M.M.; Almasoudi, H.H.; Basharat, Z. Molecular Mimicry Mapping in Streptococcus pneumoniae: Cues for Autoimmune Disorders and Implications for Immune Defense Activation. Pathogens 2023, 12, 857. [Google Scholar] [CrossRef]
- Gouttefangeas, C.; Klein, R.; Maia, A. The good and the bad of T cell cross-reactivity: Challenges and opportunities for novel therapeutics in autoimmunity and cancer. Front. Immunol. 2023, 14, 1212546. [Google Scholar] [CrossRef]
- Reynolds, S.; Rafeek, R.A.M.; Hamlin, A.; Lepletier, A.; Pandey, M.; Ketheesan, N.; Good, M. Streptococcus pyogenes vaccine candidates do not induce autoimmune responses in a rheumatic heart disease model. NPJ Vaccines 2023, 8, 9. [Google Scholar] [CrossRef]
- Burova, L.A.; Suvorov, A.N.; Pigarevsky, P.V.; Totolian, A.A. Autoimmune streptococcal glomerulonephritis: The problem of nephritogenicity of Streptococcus pyogenes. Russ. J. Infect. Immun. 2023, 13, 409–429. [Google Scholar] [CrossRef]
- Cutforth, T.; De Mille, M.M.; Agalliu, I.; Agalliu, D.J. CNS autoimmune disease after Streptococcus pyogenes infections: Animal models, cellular mechanisms and genetic factors. Future Neurol. 2016, 11, 63–76. [Google Scholar] [CrossRef]
- Draborg, A.H.; Duus, K.; Houen, G. Epstein-Barr virus in systemic autoimmune diseases. Clin. Dev. Immunol. 2013, 2013, 535738. [Google Scholar] [CrossRef]
- Poole, B.D.; Scofield, R.H.; Harley, J.B.; James, J. Epstein-Barr virus and molecular mimicry in systemic lupus erythematosus. Autoimmunity 2006, 39, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Jog, N.R.; James, J.A. Epstein Barr virus and autoimmune responses in systemic lupus erythematosus. Front. Immunol. 2021, 11, 623944. [Google Scholar] [CrossRef] [PubMed]
- Chiba, T.; Goto, S.; Yokosuka, O.; Imazeki, F.; Tanaka, M.; Fukai, K.; Takahashi, Y.; Tsujimura, H.; Saisho, H. Fatal chronic active Epstein–Barr virus infection mimicking autoimmune hepatitis. Eur. J. Gastroenterol. Hepatol. 2004, 16, 225–228. [Google Scholar] [CrossRef]
- Lünemann, J.D.; Münz, C. Epstein-Barr virus and multiple sclerosis. Nat. Rev. Microbiol. 2007, 7, 253–258. [Google Scholar] [CrossRef]
- Paroli, M.; Iannucci, G.; Accapezzato, D. Hepatitis C virus infection and autoimmune diseases. Int. J. Gen. Med. 2012, 5, 903–907. [Google Scholar] [CrossRef]
- Dudak, F.C.; Boyaci, I.H.; Orner, B.P. The discovery of small-molecule mimicking peptides through phage display. Molecules 2011, 16, 774–789. [Google Scholar] [CrossRef]
- Trela, M.; Nelson, P.N.; Rylance, P.B. The role of molecular mimicry and other factors in the association of Human Endogenous Retroviruses and autoimmunity. Apmis 2016, 124, 88–104. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Feng, T.; Xu, S.; Gao, F.; Lam, T.T.; Wang, Q.; Wu, T.; Huang, H.; Zhan, L.; Li, L. ggmsa: A visual exploration tool for multiple sequence alignment and associated data. Brief. Bioinform. 2022, 23, bbac222. [Google Scholar] [CrossRef]
- Silva, L.; Antunes, A. Omics and Remote Homology Integration to Decipher Protein Functionality. In Homology Modeling: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2023; pp. 61–81. [Google Scholar]
- Lundegaard, C.; Lamberth, K.; Harndahl, M.; Buus, S.; Lund, O.; Nielsen, M. NetMHC-3.0: Accurate web accessible predictions of human, mouse and monkey MHC class I affinities for peptides of length 8–11. Nucleic Acids Res. 2008, 36, W509–W512. [Google Scholar] [CrossRef]
- Vita, R.; Mahajan, S.; Overton, J.A.; Dhanda, S.K.; Martini, S.; Cantrell, J.R.; Wheeler, D.K.; Sette, A.; Peters, B. The immune epitope database (IEDB): 2018 update. Nucleic Acids Res. 2019, 47, D339–D343. [Google Scholar] [CrossRef]
- Morsy, S.; Morsy, A. Epitope mimicry analysis of SARS-CoV-2 surface proteins and human lung proteins. J. Mol. Graph. Model. 2021, 105, 107836. [Google Scholar] [CrossRef] [PubMed]
- Lunemann, J.D.; Jelcic, I.; Roberts, S.; Lutterotti, A.; Tackenberg, B.; Martin, R.; Munz, C. EBNA1-specific T cells from patients with multiple sclerosis cross react with myelin antigens and co-produce IFN-gamma and IL-2. J. Exp. Med. 2008, 205, 1763–1773. [Google Scholar] [CrossRef]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [PubMed]
- Kyrklund, M.; Bildo, M.; Akhi, R.; Nissinen, A.E.; Pussinen, P.; Hörkkö, S.; Wang, C. Humoral immune response to heat shock protein 60 of Aggregatibacter actinomycetemcomitans and cross-reactivity with malondialdehyde acetaldehyde-modified LDL. PLoS ONE 2020, 15, e0230682. [Google Scholar] [CrossRef] [PubMed]
- Lawson, C.M. Evidence for mimicry by viral antigens in animal models of autoimmune disease including myocarditis. Cell. Mol. Life Sci. 2000, 57, 552–560. [Google Scholar] [CrossRef]
- Benvenga, S.; Guarneri, F. Molecular mimicry and autoimmune thyroid disease. Rev. Endocr. Metab. Disord. 2016, 17, 485–498. [Google Scholar] [CrossRef]
- Amedei, A.; Bergman, M.P.; Appelmelk, B.J.; Azzurri, A.; Benagiano, M.; Tamburini, C.; van der Zee, R.; Telford, J.L.; Vandenbroucke-Grauls, C.M.; D’Elios, M. Molecular mimicry between Helicobacter pylori antigens and H+, K+—adenosine triphosphatase in human gastric autoimmunity. J. Exp. Med. 2003, 198, 1147–1156. [Google Scholar] [CrossRef]
- Nora, T.; Lomma, M.; Gomez-Valero, L.; Buchrieser, C. Molecular mimicry: An important virulence strategy employed by Legionella pneumophila to subvert host functions. Future Microbiol. 2009, 4, 691–701. [Google Scholar] [CrossRef]
- Tanim, T.M. In Silico Identification of Molecular Mimicry Between T-Cell Epitopes of Borrelia Burgdorferi and the Human Proteome: Implications for Autoimmune Response in Lyme Disease; Brac University: Dhaka, Bangladesh, 2022. [Google Scholar]
- Gkoutzourelas, A.; Barmakoudi, M.; Bogdanos, D. A bioinformatics analysis reveals novel pathogens as molecular mimicry triggers of systemic sclerosis. Mediterr. J. Rheumatol. 2020, 31, 50. [Google Scholar] [CrossRef]
- Pahari, S.; Chatterjee, D.; Negi, S.; Kaur, J.; Singh, B.; Agrewala, J. Morbid sequences suggest molecular mimicry between microbial peptides and self-antigens: A possibility of inciting autoimmunity. Front. Microbiol. 2017, 8, 1938. [Google Scholar] [CrossRef]
- Vardhini, D.; Suneetha, S.; Ahmed, N.; Joshi, D.; Karuna, S.; Magee, X.; Vijayalakshmi, D.; Sridhar, V.; Karunakar, K.; Archelos, J.; et al. Comparative proteomics of the Mycobacterium leprae binding protein myelin P0: Its implication in leprosy and other neurodegenerative diseases. Infect. Genet. Evol. 2004, 4, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Ludin, P.; Nilsson, D.; Mäser, P. Genome-wide identification of molecular mimicry candidates in parasites. PLoS ONE 2011, 6, e17546. [Google Scholar] [CrossRef] [PubMed]
- Dreyfus, D.H.; Farina, A.; Farina, G. Molecular mimicry, genetic homology, and gene sharing proteomic “molecular fingerprints” using an EBV (Epstein-Barr virus)-derived microarray as a potential diagnostic method in autoimmune disease. Immunol. Res. 2018, 66, 686–695. [Google Scholar] [CrossRef] [PubMed]
- Repac, J.; Mandić, M.; Lunić, T.; Božić, B.; Božić Nedeljković, B. Mining the capacity of human-associated microorganisms to trigger rheumatoid arthritis—A systematic immunoinformatics analysis of T cell epitopes. PLoS ONE 2021, 16, e0253918. [Google Scholar] [CrossRef]
- Garg, A.; Kumari, B.; Kumar, R.; Kumar, M. miPepBase: A database of experimentally verified peptides involved in molecular mimicry. Front. Microbiol. 2017, 8, 2053. [Google Scholar] [CrossRef]
- Martínez, Y.A.; Guo, X.; Portales-Pérez, D.P.; Rivera, G.; Castañeda-Delgado, J.E.; García-Pérez, C.A.; Enciso-Moreno, J.A.; Lara-Ramírez, E. The analysis on the human protein domain targets and host-like interacting motifs for the MERS-CoV and SARS-CoV/CoV-2 infers the molecular mimicry of coronavirus. PLoS ONE 2021, 16, e0246901. [Google Scholar]
- Tayal, S.; Bhatia, V.; Mehrotra, T.; Bhatnagar, S. ImitateDB: A database for domain and motif mimicry incorporating host and pathogen protein interactions. Amino Acids 2022, 54, 923–934. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Tunyasuvunakool, K.; Ronneberger, O.; Bates, R.; Žídek, A.; Bridgland, A.; et al. AlphaFold 2. 2020. Available online: https://predictioncenter.org/casp14/doc/presentations/2020_12_01_TS_predictor_AlphaFold2.pdf (accessed on 14 June 2023).
- David, A.; Islam, S.; Tankhilevich, E.; Sternberg, M. The AlphaFold database of protein structures: A biologist’s guide. J. Mol. Biol. 2022, 434, 167336. [Google Scholar] [CrossRef]
- Gelly, J.-C.; Joseph, A.P.; Srinivasan, N.; de Brevern, A. iPBA: A tool for protein structure comparison using sequence alignment strategies. Nucleic Acids Res. 2011, 39, W18–W23. [Google Scholar] [CrossRef]
- Zhang, Y.; Skolnick, J. TM-align: A protein structure alignment algorithm based on the TM-score. Nucleic Acids Res. 2005, 33, 2302–2309. [Google Scholar] [CrossRef]
- Rahmati, S.; Abovsky, M.; Pastrello, C.; Kotlyar, M.; Lu, R.; Cumbaa, C.A.; Rahman, P.; Chandran, V.; Jurisica, I. pathDIP 4: An extended pathway annotations and enrichment analysis resource for human, model organisms and domesticated species. Nucleic Acids Res. 2020, 48, D479–D488. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.-T.; Mathias, S.; Bologa, C.; Brunak, S.; Fernandez, N.; Gaulton, A.; Hersey, A.; Holmes, J.; Jensen, L.J.; Karlsson, A. Pharos: Collating protein information to shed light on the druggable genome. Nucleic Acids Res. 2017, 45, D995–D1002. [Google Scholar] [CrossRef] [PubMed]
- Rahmati, S.; Abovsky, M.; Pastrello, C.; Jurisica, I. pathDIP: An annotated resource for known and predicted human gene-pathway associations and pathway enrichment analysis. Nucleic Acids Res. 2017, 45, D419–D426. [Google Scholar] [CrossRef]
- Doytchinova, I.A.; Flower, D. VaxiJen: A server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinform. 2007, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.e.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Grote, A.; Hiller, K.; Scheer, M.; Münch, R.; Nörtemann, B.; Hempel, D.C.; Jahn, D. JCat: A novel tool to adapt codon usage of a target gene to its potential expression host. Nucleic Acids Res. 2005, 33, W526–W531. [Google Scholar] [CrossRef] [PubMed]
- Castiglione, F.; Bernaschi, M. C-immsim: Playing with the immune response. In Proceedings of the Sixteenth International Symposium on Mathematical Theory of Networks and Systems (MTNS2004), KU Leuven, Belgium, 5–9 July 2004. [Google Scholar]
- Jiang, S.; Bottazzi, M.E.; Du, L.; Lustigman, S.; Tseng, C.-T.K.; Curti, E.; Jones, K.; Zhan, B.; Hotez, P. Roadmap to developing a recombinant coronavirus S protein receptor-binding domain vaccine for severe acute respiratory syndrome. Expert Rev. Vaccines 2012, 11, 1405–1413. [Google Scholar] [CrossRef]
- Delgado, M.F.; Coviello, S.; Monsalvo, A.C.; Melendi, G.A.; Hernandez, J.Z.; Batalle, J.P.; Diaz, L.; Trento, A.; Chang, H.-Y.; Mitzner, W. Lack of antibody affinity maturation due to poor Toll-like receptor stimulation leads to enhanced respiratory syncytial virus disease. Nat. Med. 2009, 15, 34–41. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 1–8. [Google Scholar] [CrossRef]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro web server for protein–protein docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef]
- Comeau, S.R.; Gatchell, D.W.; Vajda, S.; Camacho, C. ClusPro: A fully automated algorithm for protein–protein docking. Nucleic Acids Res. 2004, 32, W96–W99. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.C.; Rodrigues, J.P.; Kastritis, P.L.; Bonvin, A.M.; Vangone, A.J.B. PRODIGY: A web server for predicting the binding affinity of protein–protein complexes. Bioinformatics 2016, 32, 3676–3678. [Google Scholar] [CrossRef]
- Loshaj-Shala, A.; Regazzoni, L.; Daci, A.; Orioli, M.; Brezovska, K.; Panovska, A.P.; Beretta, G.; Suturkova, L. Guillain Barré syndrome (GBS): New insights in the molecular mimicry between C. jejuni and human peripheral nerve (HPN) proteins. J. Neuroimmunol. 2015, 289, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Yokota, S.-i.; Chiba, S.; Furuyama, H.; Fujii, N. Cerebrospinal fluids containing anti-HSP70 autoantibodies from multiple sclerosis patients augment HSP70-induced proinflammatory cytokine production in monocytic cells. J. Neuroimmunol. 2010, 218, 129–133. [Google Scholar] [CrossRef]
- Mosenson, J.A.; Zloza, A.; Nieland, J.D.; Garrett-Mayer, E.; Eby, J.M.; Huelsmann, E.J.; Kumar, P.; Denman, C.J.; Lacek, A.T.; Kohlhapp, F. Mutant HSP70 reverses autoimmune depigmentation in vitiligo. Sci. Transl. Med. 2013, 5, 174ra128. [Google Scholar] [CrossRef] [PubMed]
- Lemos, J.; Giambiagi-Demarval, M.; Castro, A. Expression of heat-shock proteins in Streptococcus pyogenes and their immunoreactivity with sera from patients with streptococcal diseases. J. Med. Microbiol. 1998, 47, 711–715. [Google Scholar] [CrossRef] [PubMed]
- Shraibman, B.; Barnea, E.; Kadosh, D.M.; Haimovich, Y.; Slobodin, G.; Rosner, I.; López-Larrea, C.; Hilf, N.; Kuttruff, S.; Song, C.; et al. Identification of tumor antigens among the HLA peptidomes of glioblastoma tumors and plasma. Mol. Cell. Proteom. 2018, 17, 2132–2145. [Google Scholar] [CrossRef] [PubMed]
- Sudhir, P.-R.; Lin, T.-D.; Zhang, Q. HLA allele-specific quantitative profiling of type 1 diabetic b lymphocyte immunopeptidome. J. Proteome Res. 2021, 21, 250–264. [Google Scholar] [CrossRef] [PubMed]
- Shoda, H.; Hanata, N.; Sumitomo, S.; Okamura, T.; Fujio, K.; Yamamoto, K. Immune responses to Mycobacterial heat shock protein 70 accompany self-reactivity to human BiP in rheumatoid arthritis. Sci. Rep. 2016, 6, 22486. [Google Scholar] [CrossRef]
- Requena, J.M.; Soto, M.; Guzman, F.; Maekelt, A.; Noya, O.; Patarroyo, M.E.; Alonso, C. Mapping of antigenic determinants of the T. cruzi hsp70 in chagasic and healthy individuals. Mol. Immunol. 1993, 30, 1115–1121. [Google Scholar] [CrossRef]
- Adams, E.; Britton, W.; Morgan, A.; Sergeantson, S.; Basten, A. Individuals from Different Populations Identify Multiple and Diverse T-Cell Determinants on Mycobacterial HSP70. Scand. J. Immunol. 1994, 39, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Sigdel, K.R.; Cheng, A.; Wang, Y.; Duan, L.; Zhang, Y. The emerging functions of long noncoding RNA in immune cells: Autoimmune diseases. J. Immunol. Res. 2015, 2015, 848790. [Google Scholar] [CrossRef] [PubMed]
- Read, S.A.; Tay, E.; Shahidi, M.; George, J.; Douglas, M. Hepatitis C virus infection mediates cholesteryl ester synthesis to facilitate infectious particle production. J. Gen. Virol. 2014, 95, 1900–1910. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Leung, E.L.-H.; Pan, H.; Yao, X.; Huang, Q.; Wu, M.; Xu, T.; Wang, Y.; Cai, J.; Li, R. Identification of potential genetic causal variants for rheumatoid arthritis by whole-exome sequencing. Oncotarget 2017, 8, 111119. [Google Scholar] [CrossRef] [PubMed]
- Dudek, N.L.; Tan, C.T.; Gorasia, D.G.; Croft, N.P.; Illing, P.T.; Purcell, A. Constitutive and inflammatory immunopeptidome of pancreatic β-cells. Diabetes 2012, 61, 3018–3025. [Google Scholar] [CrossRef]
- Wong, S.M.; Jackson, M.D.; Akerley, B. Suppression of alternative lipooligosaccharide glycosyltransferase activity by UDP-galactose epimerase enhances murine lung infection and evasion of serum IgM. Front. Cell. Infect. Microbiol. 2019, 9, 160. [Google Scholar] [CrossRef]
- Maji, A.; Misra, R.; Kumar Mondal, A.; Kumar, D.; Bajaj, D.; Singhal, A.; Arora, G.; Bhaduri, A.; Sajid, A.; Bhatia, S. Expression profiling of lymph nodes in tuberculosis patients reveal inflammatory milieu at site of infection. Sci. Rep. 2015, 5, 15214. [Google Scholar] [CrossRef]
- Zhang, Z. Genomic biomarker heterogeneities between SARS-CoV-2 and COVID-19. Vaccines 2022, 10, 1657. [Google Scholar] [CrossRef]
- Hu, J.; Zhou, S.; Guo, W. Construction of the coexpression network involved in the pathogenesis of thyroid eye disease via bioinformatics analysis. Hum. Genom. 2022, 16, 1–15. [Google Scholar] [CrossRef]
- Peng, S.; Wang, J.; Wei, S.; Li, C.; Zhou, K.; Hu, J.; Ye, X.; Yan, J.; Liu, W.; Gao, G. Endogenous cellular microRNAs mediate antiviral defense against influenza A virus. Mol. Ther. Nucleic Acids 2018, 10, 361–375. [Google Scholar] [CrossRef]
- Huang, T.; Huang, X.; Yao, M. miR-143 inhibits intracellular salmonella growth by targeting ATP6V1A in macrophage cells in pig. Res. Vet. Sci. 2018, 117, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, F.; Zhang, J.; Wang, L.; Wang, J.; Wen, Z.; Wang, Z.; Shuai, L.; Wang, X.; Ge, J. The ATPase ATP6V1A facilitates rabies virus replication by promoting virion uncoating and interacting with the viral matrix protein. J. Biol. Chem. 2021, 296. [Google Scholar] [CrossRef] [PubMed]
- Grodzki, M.; Bluhm, A.P.; Schaefer, M.; Tagmount, A.; Russo, M.; Sobh, A.; Rafiee, R.; Vulpe, C.D.; Karst, S.M.; Norris, M. Genome-scale CRISPR screens identify host factors that promote human coronavirus infection. Genome Med. 2022, 14, 1–18. [Google Scholar] [CrossRef]
- Blischak, J.D.; Tailleux, L.; Mitrano, A.; Barreiro, L.B.; Gilad, Y. Mycobacterial infection induces a specific human innate immune response. Sci. Rep. 2015, 5, 16882. [Google Scholar] [CrossRef] [PubMed]
- Raphael, I.; Webb, J.; Gomez-Rivera, F.; Chase Huizar, C.A.; Gupta, R.; Arulanandam, B.P.; Wang, Y.; Haskins, W.E.; Forsthuber, T. Serum neuroinflammatory disease-induced central nervous system proteins predict clinical onset of experimental autoimmune encephalomyelitis. Front. Immunol. 2017, 8, 812. [Google Scholar] [CrossRef] [PubMed]
- Gerstner, C.; Dubnovitsky, A.; Sandin, C.; Kozhukh, G.; Uchtenhagen, H.; James, E.A.; Ronnelid, J.; Ytterberg, A.J.; Pieper, J.; Reed, E. Functional and Structural Characterization of a Novel HLA-DRB1* 04: 01-Restricted alpha-Enolase T Cell Epitope in Rheumatoid Arthritis. Front. Immunol. 2016, 7, 494. [Google Scholar] [CrossRef] [PubMed]
- Moreno, I., Jr.; Rodríguez-Sánchez, I.; Schafer, X.; Munger, J. Human cytomegalovirus induces neuronal enolase to support virally mediated metabolic remodeling. Proc. Natl. Acad. Sci. USA 2022, 119, e2205789119. [Google Scholar] [CrossRef]
- Gaseitsiwe, S.; Valentini, D.; Mahdavifar, S.; Magalhaes, I.; Hoft, D.F.; Zerweck, J.; Schutkowski, M.; Andersson, J.; Reilly, M.; Maeurer, M. Pattern recognition in pulmonary tuberculosis defined by high content peptide microarray chip analysis representing 61 proteins from M. tuberculosis. PLoS ONE 2008, 3, e3840. [Google Scholar] [CrossRef]
- Zhao, M.; Wu, J.; Li, X.; Gao, Y. Urinary candidate biomarkers in an experimental autoimmune myocarditis rat model. J. Proteom. 2018, 179, 71–79. [Google Scholar] [CrossRef]
- Mishra, N.; Huang, X.; Joshi, S.; Guo, C.; Ng, J.; Thakkar, R.; Wu, Y.; Dong, X.; Li, Q.; Pinapati, R. Immunoreactive peptide maps of SARS-CoV-2. Commun. Biol. 2021, 4, 225. [Google Scholar] [CrossRef]
- González-Sapienza, G.; Cachau, R. Identification of critical residues of an immunodominant region of Echinococcus granulosus antigen B. J. Biol. Chem. 2003, 278, 20179–20184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Minardi, L.M.; Todd Kuenstner, J.; Zekan, S.M.; Zhu, F.; Hu, Y.; Kruzelock, R. Cross—Reactivity of antibodies against microbial proteins to human tissues as basis of Crohn’s disease and other autoimmune diseases. bioRxiv 2017, 116574. [Google Scholar]
- Zhang, P.; Minardi, L.M.; Kuenstner, J.T.; Zekan, S.M.; Kruzelock, R.J.B. Seroprevalence of anti-microbial antibodies in the normal healthy population with implications in chronic diseases. bioRxiv 2019, 693655. [Google Scholar]
- Syed, S.; Viazmina, L.; Mager, R.; Meri, S.; Haapasalo, K. Streptococci and the complement system: Interplay during infection, inflammation and autoimmunity. FEBS Lett. 2020, 594, 2570–2585. [Google Scholar] [CrossRef] [PubMed]
- Harvey, K.L.; Jarocki, V.M.; Charles, I.G.; Djordjevic, S. The diverse functional roles of elongation factor Tu (EF-Tu) in microbial pathogenesis. Front. Microbiol. 2019, 10, 2351. [Google Scholar]
- Lo, K.C.; Sullivan, E.; Bannen, R.M.; Jin, H.; Rowe, M.; Li, H.; Pinapati, R.S.; Cartwright, A.J.; Tan, J.C.; Patel, J.; et al. Comprehensive profiling of the rheumatoid arthritis antibody repertoire. Arthritis Rheumatol. 2020, 72, 242–250. [Google Scholar] [CrossRef]
- Venema, W.J.; Hiddingh, S.; de Boer, J.H.; Claas, F.H.; Mulder, A.; den Hollander, A.I.; Stratikos, E.; Sarkizova, S.; van der Veken, L.T.; Janssen, G. ERAP2 increases the abundance of a peptide submotif highly selective for the birdshot uveitis-associated HLA-A29. Front. Immunol. 2021, 12, 634441. [Google Scholar] [CrossRef]
- Caradonna, S.J.; Cheng, Y. Induction of uracil-DNA glycosylase and dUTP nucleotidohydrolase activity in herpes simplex virus-infected human cells. J. Biol. Chem. 1981, 256, 9834–9837. [Google Scholar] [CrossRef]
- Stuart, D.; Upton, C.; Higman, M.; Niles, E.; McFadden, G. A poxvirus-encoded uracil DNA glycosylase is essential for virus viability. J. Virol. 1993, 67, 2503–2512. [Google Scholar] [CrossRef]
- James, E.A.; Gates, T.J.; LaFond, R.E.; Yamamoto, S.; Ni, C.; Mai, D.; Gersuk, V.H.; O’Brien, K.; Nguyen, Q.-A.; Zeitner, B. Neuroinvasive West Nile infection elicits elevated and atypically polarized T cell responses that promote a pathogenic outcome. PLoS Pathog. 2016, 12, e1005375. [Google Scholar] [CrossRef]
- Life, P.; Hassell, A.; Williams, K.; Young, S.; Bacon, P.; Southwood, T.; Gaston, J. Responses to gram negative enteric bacterial antigens by synovial T cells from patients with juvenile chronic arthritis: Recognition of heat shock protein HSP60. J. Rheumatol. 1993, 20, 1388–1396. [Google Scholar] [PubMed]
- Ford, P.; Gemmell, E.; Hamlet, S.; Hasan, A.; Walker, P.; West, M.; Cullinan, M.; Seymour, G. Cross-reactivity of GroEL antibodies with human heat shock protein 60 and quantification of pathogens in atherosclerosis. Oral Microbiol. Immunol. 2005, 20, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Rodolico, V.; Tomasello, G.; Zerilli, M.; Martorana, A.; Pitruzzella, A.; Marino Gammazza, A.; David, S.; Zummo, G.; Damiani, P.; Accomando, S.; et al. Hsp60 and Hsp10 increase in colon mucosa of Crohn’s disease and ulcerative colitis. Cell Stress Chaperones 2010, 15, 877–884. [Google Scholar] [CrossRef] [PubMed]
- De Jong, H.; Lafeber, F.F.; de Jager, W.; Haverkamp, M.H.; Kuis, W.; Bijlsma, J.W.; Prakken, B.J.; Albani, S. PAN–DR-Binding Hsp60 self epitopes induce an interleukin-10–mediated immune response in rheumatoid arthritis. Arthritis Rheum. 2009, 60, 1966–1976. [Google Scholar] [CrossRef] [PubMed]
- Dieudé, M.; Senécal, J.L.; Raymond, Y. Induction of endothelial cell apoptosis by heat-shock protein 60–reactive antibodies from anti–endothelial cell autoantibody–positive systemic lupus erythematosus patients. Arthritis Rheum. 2004, 50, 3221–3231. [Google Scholar] [CrossRef] [PubMed]
- Aragona, P.; Magazzu, G.; Macchia, G.; Bartolone, S.; Di Pasquale, G.; Vitali, C.; Ferreri, G. Presence of antibodies against Helicobacter pylori and its heat-shock protein 60 in the serum of patients with Sjögren’s syndrome. J. Rheumatol. 1999, 26, 1306–1311. [Google Scholar]
- Jones, D.B.; Coulson, A.F.; Duff, G. Sequence homologies between hsp60 and autoantigens. Immunol. Today 1993, 14, 115–118. [Google Scholar] [CrossRef]
- Marino Gammazza, A.; Rizzo, M.; Citarrella, R.; Rappa, F.; Campanella, C.; Bucchieri, F.; Patti, A.; Nikolic, D.; Cabibi, D.; Amico, G.; et al. Elevated blood Hsp60, its structural similarities and cross-reactivity with thyroid molecules, and its presence on the plasma membrane of oncocytes point to the chaperonin as an immunopathogenic factor in Hashimoto’s thyroiditis. Cell Stress Chaperones 2014, 19, 343–353. [Google Scholar] [CrossRef]
- Francesco, C.; Everly, C.; Alberto, J. HSPD1 (heat shock 60 kDa protein 1). Atlas Genet. Cytogenet. Oncol. Haematol. 2015, 19, 575–585. [Google Scholar]
- Vojdani, A.; Bazargan, M.; Vojdani, E.; Samadi, J.; Nourian, A.A.; Eghbalieh, N.; Cooper, E. Heat shock protein and gliadin peptide promote development of peptidase antibodies in children with autism and patients with autoimmune disease. Clin. Vaccine Immunol. 2004, 11, 515–524. [Google Scholar] [CrossRef]
- Gonciarz, W.; Matusiak, A.; Rudnicka, K.; Rechciński, T.; Chałubiński, M.; Czkwianianc, E.; Broncel, M.; Gajewski, A.; Chmiela, M. Autoantibodies to a specific peptide epitope of human Hsp60 (ATVLA) with homology to Helicobacter pylori HspB in H. pylori-infected patients. APMIS J. Pathol. Microbiol. Immunol. 2019, 127, 139–149. [Google Scholar]
- Lopez, J.A.M.; Cordero, J.F.B.; Russo, U.D.B.; Villarreal, M.A.G.; Hernández, F.A.T.; Bonilla, J.A.A.; Amaya, G.P.B.; Dumaguala, M.R.J.; Valencia, C.C.R. Molecular Mimetism between Cystic Fibrosis and P. aeruginosa and S. aureus, Possible Cause of Autoimmunity? In-Silico Analysis. J. Biomed. Sci. 2022, 4, 2066–2072. [Google Scholar]
- Perschinka, H.; Mayr, M.; Millonig, G.; Mayerl, C.; van der Zee, R.; Morrison, S.G.; Morrison, R.P.; Xu, Q.; Wick, G. Cross-reactive B-cell epitopes of microbial and human heat shock protein 60/65 in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Abulafia-Lapid, R.; Elias, D.; Raz, I.; Keren-Zur, Y.; Atlan, H.; Cohen, I. T cell proliferative responses of type 1 diabetes patients and healthy individuals to human hsp60 and its peptides. J. Autoimmun. 1999, 12, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Wizrah, M.S.; Chua, S.M.; Luo, Z.; Manik, M.K.; Pan, M.; Whyte, J.M.; Robertson, A.A.; Kappler, U.; Kobe, B.; Fraser, J. AICAR transformylase/IMP cyclohydrolase (ATIC) is essential for de novo purine biosynthesis and infection by Cryptococcus neoformans. J. Biol. Chem. 2022, 298, 102453. [Google Scholar] [CrossRef] [PubMed]
- Guvenel, A.; Jozwik, A.; Ascough, S.; Ung, S.K.; Paterson, S.; Kalyan, M.; Gardener, Z.; Bergstrom, E.; Kar, S.; Habibi, M. Epitope-specific airway-resident CD4+ T cell dynamics during experimental human RSV infection. J. Clin. Investig. 2020, 130, 523–538. [Google Scholar] [CrossRef]
- Cagnazzo, T.d.O.; Nogueira, C.T.; Castro, C.A.d.; Neris, D.M.; Fattori, A.C.M.; Correia, R.d.O.; Albuquerque, Y.R.; Fragelli, B.D.d.L.; Mendes, T.M.F.; Allegretti, S. Investigating Immunization With Nucleotide Enzymes of Schistosoma Mansoni: Nucleoside Diphosphate Kinase and Adenylosuccinate Lyase as New Antigenic Targets Against Schistosomiasis. Front. Immunol. 2020, 11, 569988. [Google Scholar] [CrossRef]
- Rahman, K.S.; Darville, T.; Russell, A.N.; O’Connell, C.M.; Wiesenfeld, H.C.; Hillier, S.L.; Chowdhury, E.U.; Juan, Y.-C.; Kaltenboeck, B. Discovery of human-specific immunodominant Chlamydia trachomatis B cell epitopes. mSphere 2018, 3. [Google Scholar] [CrossRef]
- Lin, S.Y.-H.; Cheng, C.-W.; Su, E. Prediction of B-cell epitopes using evolutionary information and propensity scales. BMC Bioinformatics 2013, 14, S10. [Google Scholar] [CrossRef]
- Chauvin, A.; Bouvet, G.; Boulard, C. Humoral and cellular immune responses to Fasciola hepatica experimental primary and secondary infection in sheep. Int. J. Parasitol. 1995, 25, 1227–1241. [Google Scholar] [CrossRef]
- Boes, M. Role of natural and immune IgM antibodies in immune responses. Mol. Immunol. 2000, 37, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Coutelier, J.-P.; Van Der Logt, J.; Heessen, F. IgG subclass distribution of primary and secondary immune responses concomitant with viral infection. J. Immunol. 1991, 147, 1383–1386. [Google Scholar] [CrossRef] [PubMed]
- Guillerey, C.; Nakamura, K.; Vuckovic, S.; Hill, G.R.; Smyth, M. Immune responses in multiple myeloma: Role of the natural immune surveillance and potential of immunotherapies. Cell. Mol. Life Sci. 2016, 73, 1569–1589. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; O’Connell, R.M.; Mazmanian, S. Coordination of tolerogenic immune responses by the commensal microbiota. J. Autoimmun 2010, 34, J220–J225. [Google Scholar] [CrossRef]
- Nikolich-Žugich, J. The twilight of immunity: Emerging concepts in aging of the immune system. Nat. Immunol. 2018, 19, 10–19. [Google Scholar] [CrossRef]
- Siegrist, C. Vaccine Immunology. 2008, Volume 5, pp. 17–36. Available online: http://www.who.int/immunization/documents/Elsevier_Vaccine_immunology.pdf (accessed on 16 June 2023).
- Souza-Fonseca-Guimaraes, F.; Adib-Conquy, M.; Cavaillon, J. Natural killer (NK) cells in antibacterial innate immunity: Angels or devils? Mol. Med. 2012, 18, 270–285. [Google Scholar] [CrossRef]
- Studer, G.; Rempfer, C.; Waterhouse, A.M.; Gumienny, R.; Haas, J.; Schwede, T. QMEANDisCo—Distance constraints applied on model quality estimation. Bioinformatics 2020, 36, 1765–1771. [Google Scholar] [CrossRef]
- Monirinasab, H.; Zakariazadeh, M.; Kohestani, H.; Kouhestani, M.; Fathi, F. Study of β-lactam-based drug interaction with albumin protein using optical, sensing, and docking methods. J. Biol. Phys. 2022, 48, 177–194. [Google Scholar] [CrossRef]
- Holler, P.D.; Kranz, D. Quantitative analysis of the contribution of TCR/pepMHC affinity and CD8 to T cell activation. Immunity 2003, 18, 255–264. [Google Scholar] [CrossRef]
- Vangone, A.; Bonvin, A. Contacts-based prediction of binding affinity in protein–protein complexes. eLife 2015, 4, e07454. [Google Scholar] [CrossRef]
- Czepiel, J.; Dróżdż, M.; Pituch, H.; Kuijper, E.J.; Perucki, W.; Mielimonka, A.; Goldman, S.; Wultańska, D.; Garlicki, A.; Biesiada, G.; et al. Clostridium difficile infection. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1211–1221. [Google Scholar] [CrossRef]
- Balsells, E.; Shi, T.; Leese, C.; Lyell, I.; Burrows, J.; Wiuff, C.; Campbell, H.; Kyaw, M.H.; Nair, H. Global burden of Clostridium difficile infections: A systematic review and meta-analysis. J. Glob. Health 2019, 9, 010407. [Google Scholar] [CrossRef]
- Revolinski, S.L.; Munoz-Price, L. Clostridium difficile in immunocompromised hosts: A review of epidemiology, risk factors, treatment, and prevention. Clin. Infect. Dis. 2019, 68, 2144–2153. [Google Scholar] [CrossRef] [PubMed]
- Teneberg, S.; Lönnroth, I.; López, J.F.T.; Galili, U.; Halvarsson, M.Ö.; Ångström, J.; Karlsson, K. Molecular mimicry in the recognition of glycosphingolipids by Galα3Galβ4GlcNAcβ-binding Clostridium difficile toxin A, human natural anti α-galactosyl IgG and the monoclonal antibody Gal-13: Characterization of a binding-active human glycosphingolipid, non-identical with the animal receptor. Glycobiology 1996, 6, 599–609. [Google Scholar] [PubMed]
- Mindur, J.E.; Yadav, S.K.; Ito, N.; Senoh, M.; Kato, H.; Dhib-Jalbut, S.; Ito, K. Surface layer protein a expressed in Clostridioides difficile DJNS06-36 possesses an encephalitogenic mimotope of myelin basic protein. Microorganisms 2020, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Alam, J.; Kim, Y.C.; Choi, Y. Potential role of bacterial infection in autoimmune diseases: A new aspect of molecular mimicry. Immune Netw. 2014, 14, 7–13. [Google Scholar] [CrossRef]
- Mayo, I.; Arizti, P.; Parés, A.; Oliva, J.; Doforno, R.A.; de Sagarra, M.R.; Rodés, J.; Castaño, J. Antibodies against the COOH-terminal region of E. coli ClpP protease in patients with primary biliary cirrhosis. J. Hepatol. 2000, 33, 528–536. [Google Scholar]
- Zhang, C.; Zhang, T.-X.; Liu, Y.; Jia, D.; Zeng, P.; Du, C.; Yuan, M.; Liu, Q.; Wang, Y.; Shi, F. B-cell compartmental features and molecular basis for therapy in autoimmune disease. Neurol. Neuroimmunol. Neuroinflamm. 2021, 8, e1070. [Google Scholar] [CrossRef] [PubMed]
- Mansilla, M.J.; Montalban, X.; Espejo, C. Heat shock protein 70: Roles in multiple sclerosis. Mol. Med. 2012, 18, 1018–1028. [Google Scholar] [CrossRef]
- Fouani, M.; Basset, C.A.; Mangano, G.D.; Leone, L.G.; Lawand, N.B.; Leone, A.; Barone, R. Heat shock proteins alterations in rheumatoid arthritis. Int. J. Mol. Sci. 2022, 23, 2806. [Google Scholar] [CrossRef]
- Lucchese, G.; Flöel, A. Chaperones. SARS-CoV-2 and Guillain-Barré syndrome: Molecular mimicry with human heat shock proteins as potential pathogenic mechanism. Cell Stress Chaperones 2020, 25, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Tukaj, S. Heat shock protein 70 as a double agent acting inside and outside the cell: Insights into autoimmunity. Int. J. Mol. Sci. 2020, 21, 5298. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Eguchi, T. Heat Shock Proteins and Periodontitis–Cross-Reaction Between Bacterial and Human HSP in Periodontal Infection Linking with Cardiovascular Diseases. In Heat Shock Proteins in Inflammatory Diseases; Springer: Berlin/Heidelberg, Germany, 2020; pp. 19–32. [Google Scholar]
- Malik, J.A.; Lone, R. Heat shock proteins with an emphasis on HSP 60. Mol. Biol. Rep. 2021, 48, 6959–6969. [Google Scholar] [CrossRef] [PubMed]
- Repetto, O.; Zanussi, S.; Casarotto, M.; Canzonieri, V.; De Paoli, P.; Cannizzaro, R.; De Re, V. Differential proteomics of Helicobacter pylori associated with autoimmune atrophic gastritis. Mol. Med. 2014, 20, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Qeshmi, S.A.; Dabbagh, F.; Haghighi, A.B.; Ghasemi, Y. Bioinformatics evaluation of the possibility of heat shock proteins as autoantigens in multiple sclerosis based on molecular mimicry hypothesis. J. Neuroimmunol. 2016, 295, 100–121. [Google Scholar] [CrossRef]
- Lee, Y.M.; Huh, K.C.; Yoon, S.M.; Jang, B.I.; Shin, J.E.; Koo, H.S.; Jung, Y.; Kim, S.H.; Moon, H.S.; Lee, S.; et al. Incidence and clinical outcomes of Clostridium difficile infection after treatment with tuberculosis medication. Gut Liver 2016, 10, 250. [Google Scholar] [CrossRef]
- Kullin, B.R.; Reid, S.; Abratt, V. Clostridium difficile in patients attending tuberculosis hospitals in Cape Town, South Africa, 2014–2015. Afr. J. Lab. Med. 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Obuch-Woszczatyński, P.; Dubiel, G.; Harmanus, C.; Kuijper, E.; Duda, U.; Wultańska, D.; Van Belkum, A.; Pituch, H. Emergence of Clostridium difficile infection in tuberculosis patients due to a highly rifampicin-resistant PCR ribotype 046 clone in Poland. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1027–1030. [Google Scholar] [CrossRef]
- Chang, K.; Leung, C.; Yew, W.; Lam, F.; Ho, P.; Chau, C.; Cheng, V.; Yuen, K. Analyses of fluoroquinolones and Clostridium difficile-associated diarrhoea in tuberculosis patients. Int. J. Tuberc. Lung Dis. 2009, 13, 341–346. [Google Scholar]
- Collini, P.J.; Kuijper, E.; Dockrell, D. Clostridium difficile infection in patients with HIV/AIDS. Curr. HIV/AIDS Rep. 2013, 10, 273–282. [Google Scholar] [CrossRef]
- Clem, A. Fundamentals of vaccine immunology. J. Glob. Infect. Dis. 2011, 3, 73. [Google Scholar] [CrossRef] [PubMed]
- Irvine, D.J.; Swartz, M.A.; Szeto, G. Engineering synthetic vaccines using cues from natural immunity. Nat. Mater. 2013, 12, 978–990. [Google Scholar] [PubMed]
- Vadalà, M.; Poddighe, D.; Laurino, C.; Palmieri, B. Vaccination and autoimmune diseases: Is prevention of adverse health effects on the horizon? EPMA J. 2017, 8, 295–311. [Google Scholar] [PubMed]
- Horwitz, D.A.; Fahmy, T.M.; Piccirillo, C.A.; La Cava, A. Rebalancing immune homeostasis to treat autoimmune diseases. Trends Immunol. 2019, 40, 888–908. [Google Scholar] [CrossRef] [PubMed]
- Dangkoub, F.; Sankian, M.; Tafaghodi, M.; Jaafari, M.R.; Badiee, A. The impact of nanocarriers in the induction of antigen-specific immunotolerance in autoimmune diseases. J. Control. Release 2021, 339, 274–283. [Google Scholar] [CrossRef]
- Divangahi, M.; Aaby, P.; Khader, S.A.; Barreiro, L.B.; Bekkering, S.; Chavakis, T.; van Crevel, R.; Curtis, N.; DiNardo, A.R.; Dominguez-Andres, J. Trained immunity, tolerance, priming and differentiation: Distinct immunological processes. Nat. Immunol. 2021, 22, 2–6. [Google Scholar] [CrossRef]
- Chong, C.R.; Park, V.J.; Cohen, B.; Postow, M.A.; Wolchok, J.D.; Kamboj, M. Safety of inactivated influenza vaccine in cancer patients receiving immune checkpoint inhibitors. Clin. Infect. Dis. 2020, 70, 193–199. [Google Scholar] [CrossRef]
- Ke, Y.; Jiang, G.; Sun, D.; Kaplan, H.J.; Shao, H. Anti-CD3 antibody ameliorates experimental autoimmune uveitis by inducing both IL-10 and TGF-β dependent regulatory T cells. Clin. Immunol. 2011, 138, 311–320. [Google Scholar] [CrossRef]
- Wraith, D. Therapeutic peptide vaccines for treatment of autoimmune diseases. Immunol. Lett. 2009, 122, 134–136. [Google Scholar] [CrossRef][Green Version]
- Ishina, I.A.; Zakharova, M.Y.; Kurbatskaia, I.N.; Mamedov, A.E.; Belogurov, A.A., Jr.; Gabibov, A. MHC Class II Presentation in Autoimmunity. Cells 2023, 12, 314. [Google Scholar]
- Liu, J.; Liu, Z.; Pang, Y.; Zhou, H. The interaction between nanoparticles and immune system: Application in the treatment of inflammatory diseases. J. Nanobiotechnol. 2022, 20, 1–25. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
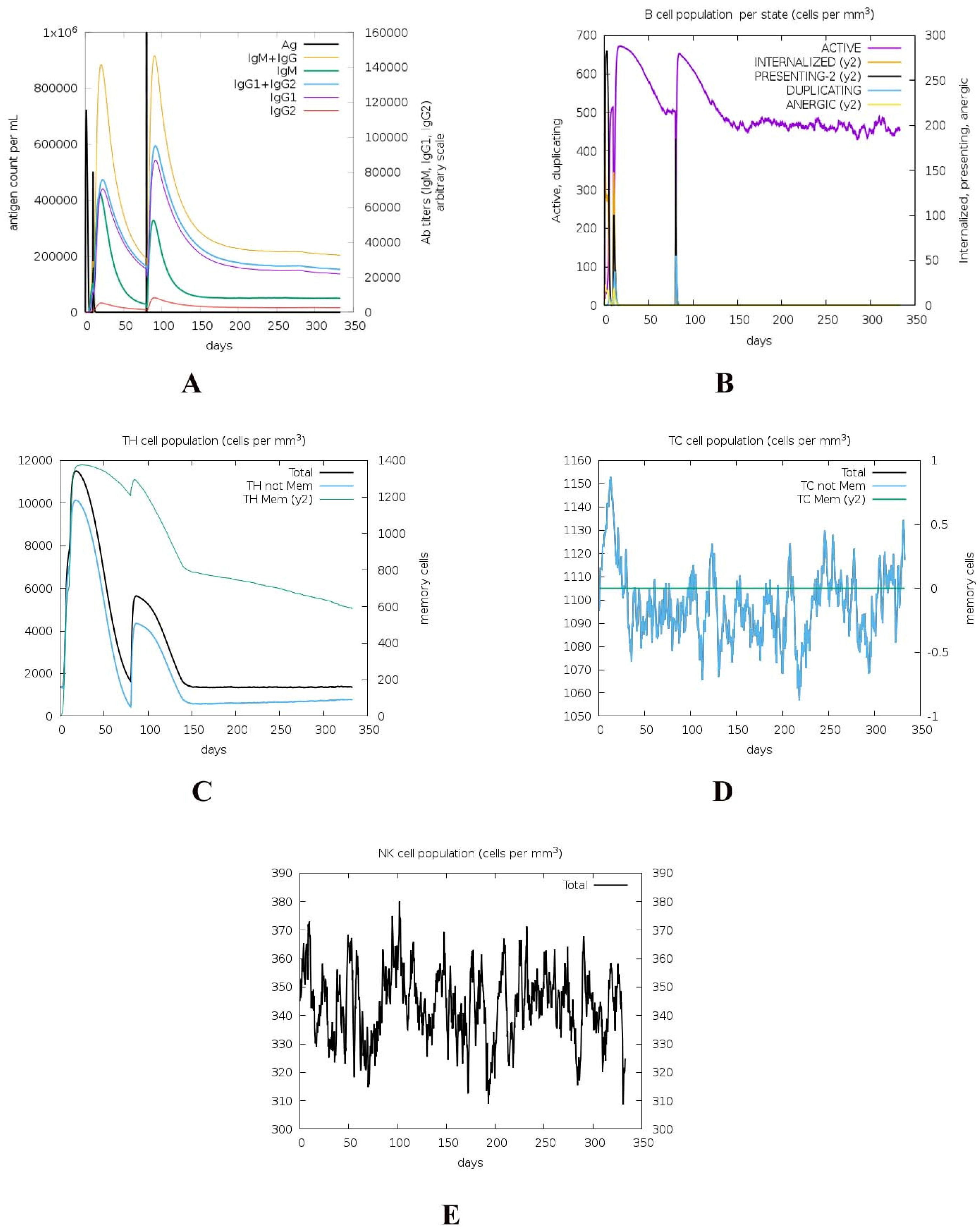{kind=link}
{kind=link}
| Serial no. | Name | UniProt ID of Human Homolog | NCBI Accession of Bacterial Homolog | Bacterial Protein Structure AlphaFold ID | No. of Similar Peptides (Length ≥ 10) | Molecular Mimic Region (Length ≥ 10) | Superposed Protein RMSD (iPBA) | TM-Align RMSD |
|---|---|---|---|---|---|---|---|---|
| 1 | Molecular chaperone DnaK | P38646 | QWS53804.1 | Q182E8 | 6 | GIDLGTTNSCVAV RTTPSVVAFT INEPTAAALAYG LLLDVTPLSLGIET RNTTIPTKKSQ PQIEVTFDIDANGIV | 1.13 | 2.0 |
| 2 | Translation elongation factor 4 | Q8N442 | QWS53810.1 | Q182F4 | 3 | DHGKSTLADRL LNLIDTPGHVDF VLAKCYGGDI | 1.25 | 1.55 |
| 3 | Uracil-DNA glycosylase | P13051 | QWS53824.1 | Q182G9 | 0 | - | 1.01 | 1.13 |
| 4 | Acetyl-CoA C-acetyltransferase | Q9BWD1 | QWS54012.1 | Q18AR0 | 1 | NASGINDGAA | 0.70 | 0.75 |
| 5 | 3-oxoacid CoA-transferase subunit B | E9PDW2 | QWS54014.1 | Q183B1 | 0 | - | 1.00 | 1.74 |
| 6 | UDP-glucose 4-epimerase GalE | Q14376 | QWS54050.1 | Q183E8 | 3 | GGAGYIGSHT VFSSSATVYG DGTGVRDYIHV | 0.79 | 0.79 |
| 7 | V-type proton ATPase subunit B | P21281 | QWS54278.1 | Q184E3 | 2 | YAEALREVSAA THPIPDLTGYITEGQI | 0.81 | 0.91 |
| 8 | V-type ATP synthase catalytic unit A | P38606 | QWS54279.1 | Q184E7 | 2 | MPVAAREASIYTGIT MADSTSRWAEALRE | 0.98 | 1.03 |
| 9 | Phosphopyruvate hydratase | P09104 | QWS54488.1 | Q181T5 | 3 | LDSRGNPTVEV QEFMILPVGA VGDEGGFAPN | 0.87 | 1.39 |
| 10 | ATP-dependent Clp endopeptidase proteolytic subunit ClpP | Q16740 | QWS54701.1 | Q180F0 | 1 | VVEQTGRGER | 0.51 | 0.51 |
| 11 | ATP-dependent Clp endopeptidase proteolytic subunit ClpP | Q16740 | QWS54730.1 | Q180J6 | 0 | - | 0.56 | 0.56 |
| 12 | F0F1 ATP synthase subunit beta | P06576 | QWS54823.1 | Q184E3 | 11 | TGIKVVDLLAPY KGGKIGLFGGAGVGKTVLI G VGERTREGNDLY GQMNEPPGAR DNIFRFTQAGSEVSALLGR PSAVGYQPTLAT TKKGSITSVQA YVPADDLTDPAPATTF LGIYPAVDPL LQDIIAILGMDELS RARKIQRFLSQ | 2.11 | 2.73 |
| 13 | F0F1 ATP synthase subunit alpha | P25705 | QWS54825.1 | Q184E7 | 7 | IKEGDIVKRTG PIGRGQRELIIGDRQTGKTSI CIYVAIGQKRST YTIVVSATAS YDDLSKQAVAYR MSLLLRRPPGREAYPGDVFYLHSRLLERAAK IETQAGDVSAYIPTNVISITDGQI | 1.52 | 2.69 |
| 14 | Elongation factor Tu | P49411 | QWS55098.1 | Q18CE4 | 3 | GTIGHVDHGKTTLTAAITK DCPGHADYVKNMITG DGPMPQTREH | 0.98 | 1.12 |
| 15 | Elongation factor Tu | P49411 | QWS55112.1 | Q18CE4 | 3 | GTIGHVDHGKTTLTAAITK DCPGHADYVKNMITG DGPMPQTREH | 0.98 | 1.12 |
| 16 | Methionine adenosyltransferase | P31153 | QWS55174.1 | Q18CL7 | 4 | EGHPDKICDQISD RFVIGGPQGD HGGGAFSGKD TKVDRSAAYAAR | 0.72 | 0.93 |
| 17 | Chaperonin GroEL | P10809 | QWS55239.1 | Q18CT5 | 3 | AGDGTTTATVLA VVAVKAPGFGD DALNATRAAVEEGIV | 1.22 | 3.61 |
| 18 | Isocitrate/isopropylmalate dehydrogenase family protein | P50213 | QWS55807.1 | Q18A33 | 5 | VTLIPGDGIGPE VMPNLYGDILSDL AGDGTTTATVLA VVAVKAPGFGD DALNATRAAVEEGIV | 0.89 | 1.48 |
| 19 | Phosphoribosylaminoimidazolecarboxamide formyltransferase | P31939 | QWS55813.1 | Q18A34 | 5 | WQLVKELKEA SFKHVSPAGAAVG REVSDGIIAPGY KYTQSNSVCYAK GAGQQSRIHCTRLAG | 0.73 | 1.23 |
| 20 | Acyl-CoA dehydrogenase | P16219 | QWS55947.1 | Q18AQ1 | 2 | LIFEDCRIPK ITEIYEGTSE | 0.72 | 0.99 |
| 21 | Acetyl-CoA C-acetyltransferase | Q9BWD1 | QWS55952.1 | Q18AR0 | 2 | NASGINDGAA | 0.70 | 0.75 |
| 22 | Fe-S cluster assembly scaffold protein NifU | Q9H1K1 | QWS56138.1 | Q18BE3 | 2 | GCGSAIASSS | 1.01 | 1.39 |
| 23 | Adenylosuccinate lyase | P30566 | QWS56192.1 | Q18BJ9 | 2 | RGVKGTTGTQASFL YKRNPMRSER | 0.81 | 0.94 |
| Serial no. | Protein Homolog | PHAROS | PATHDIP | Literature | ||||
|---|---|---|---|---|---|---|---|---|
| Autoimmunity Pathway | Infection Pathway | Autoimmune Pathway | Infection Pathway | Autoimmune Pathway | Infection Pathway | |||
| 1 | P38646 | Molecular chaperone DnaK | Autoimmune disease, Parkinson’s disease | Perinatal necrotizing enterocolitis, HIV, Tuberculosis | Parkinson’s disease, Huntington’s disease, Diabetes mellitus, Alzheimer’s | Tuberculosis, HIV, Papillomavirus, E. coli, Cytomegalovirus, Staphylococcus sp., Legionellosis, Chagas, Leishmaniasis, Measles | Guillain–Barré syndrome [69], Multiple sclerosis [70], Vitiligo [71], Systemic lupus erythematosus [72], Ankylosing spondylitis [73], Type I Diabetes mellitus [74], Rheumatoid arthritis [75] | Trypanosoma cruzi [76], Mycobacterium leprae [77] |
| 2 | Q8N442 | Translation elongation factor 4 | - | - | Tuberculosis, Rheumatoid arthritis | - | - | |
| 3 | P13051 | Uracil-DNA glycosylase | - | - | - | HIV, Viral carcinogenesis | - | - |
| 4 | Q9BWD1 | Acetyl-CoA C-acetyltransferase | - | - | - | HBV, Viral carcinogenesis | Systemic lupus erythematosus [78] | HCV [79] |
| 5 | E9PDW2 | 3-oxoacid CoA-transferase subunit B | Crohn’s disease | - | - | - | Rheumatoid arthritis [80] | - |
| 6 | Q14376 | UDP-glucose 4-epimerase GalE | Psoriasis, Interstitial cystitis | Tinea corporis, Tinea pedis | - | - | Type I Diabetes mellitus [81] | Hemophilus influenzae [82] |
| 7 | P21281 | V-type proton ATPase subunit B | IgA glomerulonephritis | - | Huntington’s disease, Rheumatoid arthritis | Helicobacter pylori infection, HPV, Tuberculosis, Viral carcinogenesis, Vibrio chloerae, HIV | - | Tuberculosis [83], SARS-CoV-2 [84] |
| 8 | P38606 | V-type ATP synthase catalytic unit A | Psoriasis, Myopathy | - | Alzheimer’s disease, Huntington’s disease, Parkinson’s disease, Rheumatoid arthritis, | HPV, H. pylori, Tuberculosis, Vibrio cholerae, Viral carcinogenesis, HIV | Thyroid eye disease [85] | Influenza H1N1 [86], Salmonellosis [87], Rabies virus [88], SARS-CoV-2 [89], Tuberculosis [90] |
| 9 | P09104 | Phosphopyruvate hydratase | Psoriasis | - | - | - | Autoimmune encephalomyelitis [91], Rheumatoid arthritis [92] | Cytomegalovirus [93] |
| 10 | Q16740 | ATP-dependent Clp endopeptidase proteolytic subunit ClpP | Psoriasis | - | Alzheimer’s disease, Huntington’s disease, Parkinson’s disease | - | - | Tuberculosis [94] |
| 11 | P06576 | F0F1 ATP synthase subunit beta | - | - | Alzheimer’s, Huntington’s, Parkinson’s, Non-alcoholic fatty-acid liver diseases | Epstein–Barr virus infection, HBV, HCV, HPV, Measles, Legionellosis, E. coli | Autoimmune myocarditis [95] | MERS coronavirus [96], Echinococcus granulosus [97] |
| 12 | P25705 | F0F1 ATP synthase subunit alpha | Alzheimer’s disease | - | - | - | Sjogren’s syndrome [98], Crohn’s disease [98], Ankolysing spondolytis [73] | - |
| 13 | P49411 | Elongation factor Tu | - | - | Huntington’s, Parkinson’s | HCV, HBV, Legionellosis, E. coli, V. cholerae | Sjogren’s syndrome [99], Crohn’s disease [98], Ankolysing spondylitis [73] | Streptococcus pneumoniae [100], bacteria like Bacillus anthracis, Francisella talurensis, Staphylococcus sp., E. coli, H. pylori, etc. [101] |
| 14 | P31153 | Methionine adenosyltransferase | Type 2 diabetes mellitus, demyelinating diseases, MODY, Psoriasis, fatty liver, or non-alcoholic steatohepatitis | - | - | - | Rheumatoid arthritis [102], Uveitis [103] | Herpes simplex type 1 [104], Poxvirus [105], West Nile virus [106] |
| 15 | P10809 | Chaperonin GroEL | Allergic rhinitis | Tuberculosis, HIV | - | - | Type I Diabetes [103], Juvenile chronic arthritis [107], Atherosclerosis [108], Crohn’s disease [109], Rheumatoid arthritis [110], Systemic lupus erythematosus [111], Sjogren syndrome [112], Hashimoto thyroiditis [113,114], and myasthenia gravis [115], Autism [116] | H. pylori [117], P. aeruginosa and S. aureus [118] |
| 16 | P50213 | Isocitrate dehydrogenase [NAD] subunit alpha | Psoriasis | - | Huntington’s disease, Parkinson’s disease | Epstein–Barr virus, HCV, Legionellosis | Atherosclerosis [119], Type I diabetes [120] | H. pylori [117] |
| 17 | P31939 | Phosphoribosylaminoimidazolecarboxamide formyltransferase | Rheumatoid arthritis, Psoriatic arthritis, Erythrodermic psoriasis, Pustular psoriasis, Plaque psoriasis, Diabetes mellitus, Juvenile idiopathic arthritis | - | - | - | - | C. neoformans [121] |
| 18 | P16219 | Acyl-CoA dehydrogenase | Allergic rhinitis, Ulcerative colitis, Crohn’s disease | - | - | - | - | - |
| 19 | Q9BWD1 | Acetyl-CoA C-acetyltransferase | - | - | Parkinson’s disease | HBV, Viral carcinogenesis | - | - |
| 20 | Q9H1K1 | Fe-S cluster assembly scaffold protein NifU | - | - | Parkinson’s disease | Influenza, HIV | - | Human respiratory syncytial virus [122], SARS-CoV-1 [96] |
| 21 | P30566 | Adenylosuccinate lyase | Psoriasis | - | - | - | - | Schistosomiasis [123], Chlamydia sp. [124] |
| Serial no. | Antigenic Score | Sequence | Length | Immunogenicity Score | SVM Score for Toxicity | Hydrophobicity | Hydropathicity | Hydrophilicity | Charge | Mol Wt | Allergenicity |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. | 1.09 | GIDLGTTNSCVAV | 15 | 0.65 | 0.07 | 0.85 | −0.42 | −1 | 1249.59 | 0.07 | No |
| 2. | 0.58 | RTTPSVVAFT | 12 | 0.35 | −0.07 | 0.4 | −0.39 | 1 | 1078.36 | −0.07 | No |
| 3. | 0.89 | LLLDVTPLSLGIET | 12 | 0.31 | 0.15 | 1.18 | −0.49 | −2 | 1483.99 | 0.15 | yes |
| 4. | 1.39 | PQIEVTFDIDANGIV | 11 | 0.29 | 0.04 | 0.42 | −0.16 | −3 | 1631.04 | 0.04 | yes |
| 5. | 0.80 | DHGKSTLADRL | 12 | 0.26 | −0.33 | −1.01 | 0.66 | 0.5 | 1212.48 | −0.33 | No |
| 6. | 1.22 | NASGINDGAA | 12 | 0.26 | −0.05 | −0.22 | 0.04 | −1 | 889.02 | −0.05 | No |
| 7. | 0.65 | GGAGYIGSHT | 24 | 0.21 | 0.08 | −0.13 | −0.52 | 0.5 | 919.11 | 0.08 | No |
| 8. | 1.47 | DGTGVRDYIHV | 10 | 0.18 | −0.15 | −0.42 | 0.09 | −0.5 | 1231.49 | −0.15 | No |
| 9. | 1.20 | LDSRGNPTVEV | 10 | 0.18 | −0.23 | −0.57 | 0.39 | −1 | 1186.44 | −0.23 | yes |
| 10. | 1.48 | VVEQTGRGER | 11 | 0.14 | −0.42 | −1.26 | 0.88 | 0 | 1130.37 | −0.42 | No |
| 11. | 0.54 | TGIKVVDLLAPY | 10 | 0.12 | 0.1 | 0.91 | −0.47 | 0 | 1288.73 | 0.1 | No |
| 12. | 1.25 | VGERTREGNDLY | 11 | 0.12 | −0.41 | −1.48 | 0.77 | −1 | 1408.66 | −0.41 | No |
| 13. | 1.03 | PSAVGYQPTLAT | 10 | 0.11 | 0.03 | 0.08 | −0.58 | 0 | 1204.51 | 0.03 | No |
| 14. | 1.04 | TKKGSITSVQA | 14 | 0.11 | −0.19 | −0.38 | 0.2 | 2 | 1119.44 | −0.19 | no |
| 15. | 0.54 | LGIYPAVDPL | 10 | 0.09 | 0.19 | 0.97 | −0.67 | −1 | 1057.4 | 0.19 | yes |
| 16. | 0.65 | IKEGDIVKRTG | 10 | 0.06 | −0.29 | −0.69 | 0.86 | 1 | 1215.58 | −0.29 | No |
| 17. | 0.70 | CIYVAIGQKRST | 12 | 0.05 | −0.13 | 0.2 | −0.23 | 2 | 1338.76 | −0.13 | No |
| 18. | 0.53 | YTIVVSATAS | 10 | 0.02 | 0.14 | 1.22 | −0.83 | 0 | 1011.27 | 0.14 | No |
| 19. | 0.84 | IETQAGDVSAYIPTNVISITDGQI | 12 | 0.01 | 0.02 | 0.25 | −0.26 | −3 | 2506.13 | 0.02 | No |
| 20. | 0.56 | DGPMPQTREH | 13 | −0.05 | −0.41 | −2.06 | 0.7 | −0.5 | 1167.4 | −0.41 | No |
| 21. | 1.41 | EGHPDKICDQISD | 10 | −0.05 | −0.28 | −1.22 | 0.8 | −2.5 | 1456.73 | −0.28 | No |
| 22. | 1.22 | HGGGAFSGKD | 12 | −0.06 | −0.1 | −0.84 | 0.28 | 0.5 | 932.1 | −0.1 | No |
| 23. | 0.57 | TKVDRSAAYAAR | 15 | −0.12 | −0.35 | −0.65 | 0.51 | 2 | 1308.6 | −0.35 | No |
| 24. | 1.75 | AGDGTTTATVLA | 12 | −0.13 | 0.06 | 0.52 | −0.28 | −1 | 1077.32 | 0.06 | No |
| 25. | 1.07 | VTLIPGDGIGPE | 10 | −0.16 | 0.11 | 0.41 | −0.11 | −2 | 1167.51 | 0.11 | No |
| 26. | 1.76 | AGDGTTTATVLA | 14 | −0.17 | 0.06 | 0.52 | −0.28 | −1 | 1077.32 | 0.06 | No |
| 27. | 1.62 | GAGQQSRIHCTRLAG | 13 | −0.18 | −0.23 | −0.5 | 0.01 | 2.5 | 1554.97 | −0.23 | No |
| 28. | 1.22 | NASGINDGAA | 10 | −0.19 | −0.05 | −0.22 | 0.04 | −1 | 889.02 | −0.05 | No |
| 29. | 1.42 | GCGSAIASSS | 10 | −0.20 | 0.06 | 0.66 | −0.26 | 0 | 839.01 | 0.06 | No |
| 30. | 1.65 | RGVKGTTGTQASFL | 11 | −0.22 | −0.14 | −0.24 | −0.07 | 2 | 1422.81 | −0.14 | No |
| 31. | 0.55 | YKRNPMRSER | 11 | −0.25 | −0.77 | −2.62 | 1.19 | 3 | 1336.66 | −0.77 | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alshamrani, S.; Mashraqi, M.M.; Alzamami, A.; Alturki, N.A.; Almasoudi, H.H.; Alshahrani, M.A.; Basharat, Z. Mining Autoimmune-Disorder-Linked Molecular-Mimicry Candidates in Clostridioides difficile and Prospects of Mimic-Based Vaccine Design: An In Silico Approach. Microorganisms 2023, 11, 2300. https://doi.org/10.3390/microorganisms11092300
Alshamrani S, Mashraqi MM, Alzamami A, Alturki NA, Almasoudi HH, Alshahrani MA, Basharat Z. Mining Autoimmune-Disorder-Linked Molecular-Mimicry Candidates in Clostridioides difficile and Prospects of Mimic-Based Vaccine Design: An In Silico Approach. Microorganisms. 2023; 11(9):2300. https://doi.org/10.3390/microorganisms11092300
Chicago/Turabian StyleAlshamrani, Saleh, Mutaib M. Mashraqi, Ahmad Alzamami, Norah A. Alturki, Hassan H. Almasoudi, Mohammed Abdulrahman Alshahrani, and Zarrin Basharat. 2023. "Mining Autoimmune-Disorder-Linked Molecular-Mimicry Candidates in Clostridioides difficile and Prospects of Mimic-Based Vaccine Design: An In Silico Approach" Microorganisms 11, no. 9: 2300. https://doi.org/10.3390/microorganisms11092300
APA StyleAlshamrani, S., Mashraqi, M. M., Alzamami, A., Alturki, N. A., Almasoudi, H. H., Alshahrani, M. A., & Basharat, Z. (2023). Mining Autoimmune-Disorder-Linked Molecular-Mimicry Candidates in Clostridioides difficile and Prospects of Mimic-Based Vaccine Design: An In Silico Approach. Microorganisms, 11(9), 2300. https://doi.org/10.3390/microorganisms11092300